- ВЕЩЕСТВО НАСЛЕДСТВЕННОСТИ

Содержание
- 2. 1. «Белковая» гипотеза о строении генов До середины 40-х годов ХХ в. о было установлено, что
- 3. Рис.1. Гипотетическая схема хромосомы по Н.К. Кольцову. Внутри хромосомы (контурная линия) — две генонемы, два пучка
- 4. Таким образом, Кольцов первый предположил, что генетический «каркас» хромосомы составляет гигантская линейная макромолекула, построенная из ограниченного
- 5. 2. Доказательство генетической роли ДНК Исследования, показавшие, что генетическим материалом является ДНК, лежали в стороне от
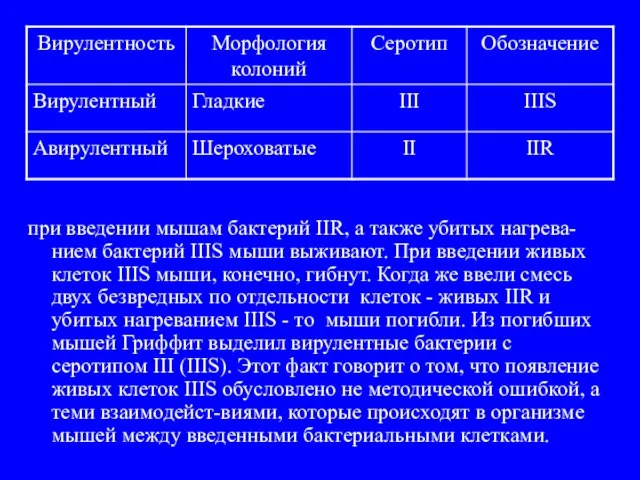
- 6. при введении мышам бактерий IIR, а также убитых нагрева-нием бактерий IIIS мыши выживают. При введении живых
- 7. Убитые нагреванием бактерии IIIS каким-то образом превращают живые клетки IIR в вирулентные клетки IIIS. Гриффит назвал
- 8. В 1944 г. Эвери, Мак-Леоду и Мак-Карти осуществили химическую идентификацию трансформирующего начала. Они экстрагировали и очистили
- 9. 2.2. Опыты Херши и Чейз (фаговая инфекция) Основные компоненты фага — ДНК и белок (рис. 1).
- 10. Полученные меченые фаги, использовали для заражения и лизиса бактерий (3). Авторы показали, что именно фаговая ДНК
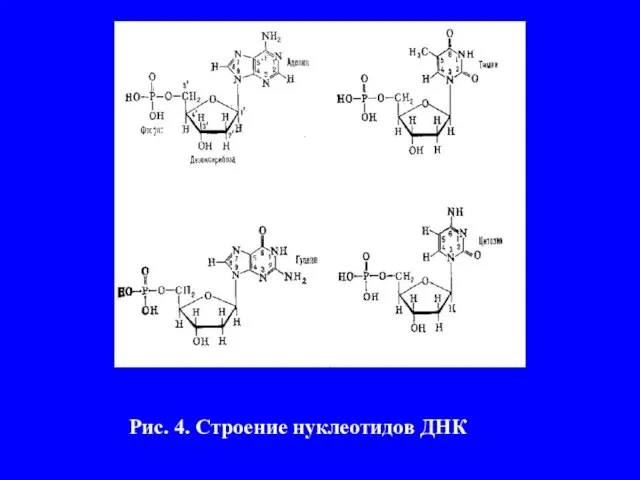
- 11. 3. Изучение химического состава молекулы ДНК Впервые нуклеиновые кислоты были обнаружены в 1869 г. швейцарским биохимиком
- 12. Рис. 4. Строение нуклеотидов ДНК
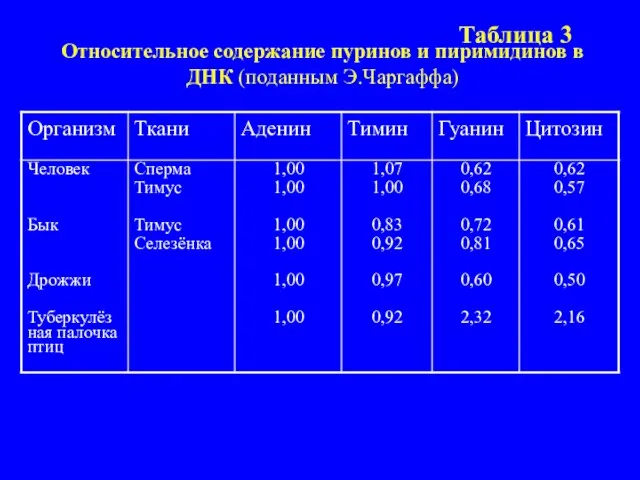
- 13. Э. Чаргафф в 1950 г. выделил высокоочищенную ДНК из разных тканей различных организмов. Образцы ДНК гидро-лизовались
- 14. Таблица 3 Относительное содержание пуринов и пиримидинов в ДНК (поданным Э.Чаргаффа)
- 16. Скачать презентацию

Слайд 3Рис.1. Гипотетическая схема хромосомы по Н.К. Кольцову.
Внутри хромосомы (контурная линия) —
Рис.1. Гипотетическая схема хромосомы по Н.К. Кольцову.
Внутри хромосомы (контурная линия) —

Слайд 4 Таким образом, Кольцов первый предположил, что генетический «каркас» хромосомы составляет гигантская
Таким образом, Кольцов первый предположил, что генетический «каркас» хромосомы составляет гигантская

Слайд 52. Доказательство генетической роли ДНК
Исследования, показавшие, что генетическим материалом является ДНК,
2. Доказательство генетической роли ДНК
Исследования, показавшие, что генетическим материалом является ДНК,

Слайд 6при введении мышам бактерий IIR, а также убитых нагрева-нием бактерий IIIS мыши
при введении мышам бактерий IIR, а также убитых нагрева-нием бактерий IIIS мыши
Слайд 7 Убитые нагреванием бактерии IIIS каким-то образом превращают живые клетки IIR в
Убитые нагреванием бактерии IIIS каким-то образом превращают живые клетки IIR в

Слайд 8 В 1944 г. Эвери, Мак-Леоду и Мак-Карти осуществили химическую идентификацию трансформирующего
В 1944 г. Эвери, Мак-Леоду и Мак-Карти осуществили химическую идентификацию трансформирующего

Слайд 92.2. Опыты Херши и Чейз (фаговая инфекция)
Основные компоненты фага — ДНК
2.2. Опыты Херши и Чейз (фаговая инфекция)
Основные компоненты фага — ДНК

Слайд 10Полученные меченые фаги, использовали для заражения и лизиса бактерий (3). Авторы показали,
Полученные меченые фаги, использовали для заражения и лизиса бактерий (3). Авторы показали,

Слайд 113. Изучение химического состава молекулы ДНК
Впервые нуклеиновые кислоты были обнаружены
3. Изучение химического состава молекулы ДНК
Впервые нуклеиновые кислоты были обнаружены

Слайд 12Рис. 4. Строение нуклеотидов ДНК
Рис. 4. Строение нуклеотидов ДНК
Слайд 13 Э. Чаргафф в 1950 г. выделил высокоочищенную ДНК из разных тканей
Э. Чаргафф в 1950 г. выделил высокоочищенную ДНК из разных тканей

Слайд 14 Таблица 3
Относительное содержание пуринов и пиримидинов в ДНК (поданным Э.Чаргаффа)
Таблица 3 Относительное содержание пуринов и пиримидинов в ДНК (поданным Э.Чаргаффа)
 Экономика семьи Д/З: § 18, читать
Экономика семьи Д/З: § 18, читать Презентация на тему Синтаксическая синонимия. Открытый урок
Презентация на тему Синтаксическая синонимия. Открытый урок Kazaerostat. Your dreams are real
Kazaerostat. Your dreams are real Анализ методической работы
Анализ методической работы Гигиена девочки. Уход за кожей
Гигиена девочки. Уход за кожей Тренажер лексики
Тренажер лексики Планирование развёртывания полевого магистрального трубопровода
Планирование развёртывания полевого магистрального трубопровода Команда СШОР Русичи г. Орла: ФК Орел – 2008. Для спонсоров и партнеров
Команда СШОР Русичи г. Орла: ФК Орел – 2008. Для спонсоров и партнеров Мониторинг ТВСостояние проектов
Мониторинг ТВСостояние проектов Ochrona pracownikow placowek dyplomatycznych i konsularnych
Ochrona pracownikow placowek dyplomatycznych i konsularnych Акции: продукты гаммы Fusio-Dose
Акции: продукты гаммы Fusio-Dose алгоритм
алгоритм 1 апреля – праздник смеха
1 апреля – праздник смеха Особенности кадровой политики в неустойчивых деловых организациях. Тема 5
Особенности кадровой политики в неустойчивых деловых организациях. Тема 5 Циклический алгоритм 6 класс
Циклический алгоритм 6 класс Творческое ассорти
Творческое ассорти Маска, я тебя знаю, или Тайны сценического грима
Маска, я тебя знаю, или Тайны сценического грима Практический опыт использования ИКТ в преподавании биологии 6,8 классы
Практический опыт использования ИКТ в преподавании биологии 6,8 классы Воинский учет в 2022 году
Воинский учет в 2022 году Сила трения в природе
Сила трения в природе Путешествие
Путешествие Создание букета на собственных стеблях без использования каркаса
Создание букета на собственных стеблях без использования каркаса Презентация дидактического материала для работы с одарёнными детьми на уроках литературного чтения ( развитие творческого вообр
Презентация дидактического материала для работы с одарёнными детьми на уроках литературного чтения ( развитие творческого вообр Презентация на тему Русские иконописцы
Презентация на тему Русские иконописцы Компания Commencal
Компания Commencal Сохраним мир живой природы
Сохраним мир живой природы Аудио эксперимент. Гид по генным ключам
Аудио эксперимент. Гид по генным ключам Абсолютные и относительные статистические величины
Абсолютные и относительные статистические величины