- 5. Ядро, Организация генетического материала эукариот (2)

Содержание
- 2. Эукариотная ДНК Расположена: В ядре В митохондриях
- 3. Ядерная ДНК Длинные линейные молекулы; В виде хроматина ↔ хромосом; Около 10% ДНК – кодирующая, остальная
- 4. Митохондриальная ДНК Небольшие кольцевые молекулы Количество – от 2-х до 10 в одной митохондрии Количество мт-ДНК
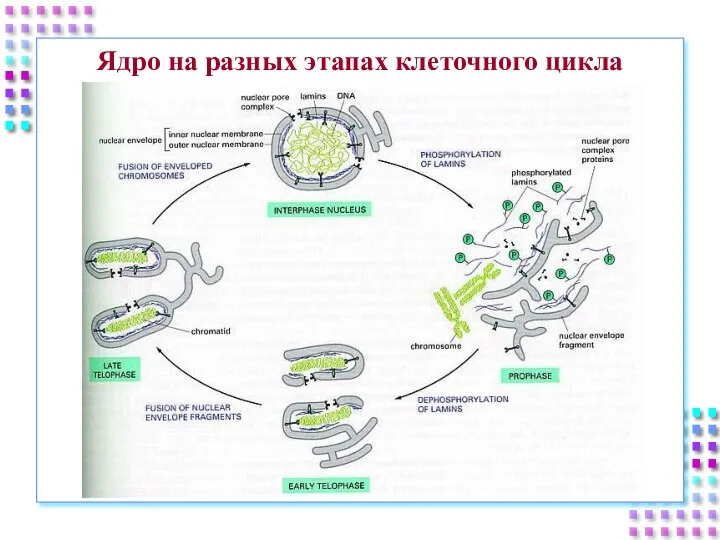
- 5. Ядро на разных этапах клеточного цикла
- 6. Функции ядра Хранение и функциональная упаковка ДНК: Эухроматин – активная ГИ; Гетерохроматин – неактивная ДНК. Контроль
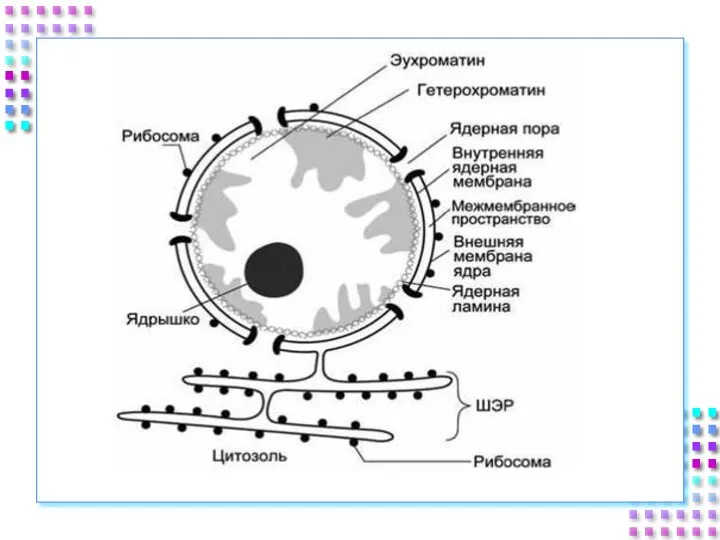
- 7. Строение ядра 1) Ядерная оболочка: Внешняя ядерная мембрана Внутренняя ядерная мембрана Перинуклеарное пространство Поровый комплекс Ядерная
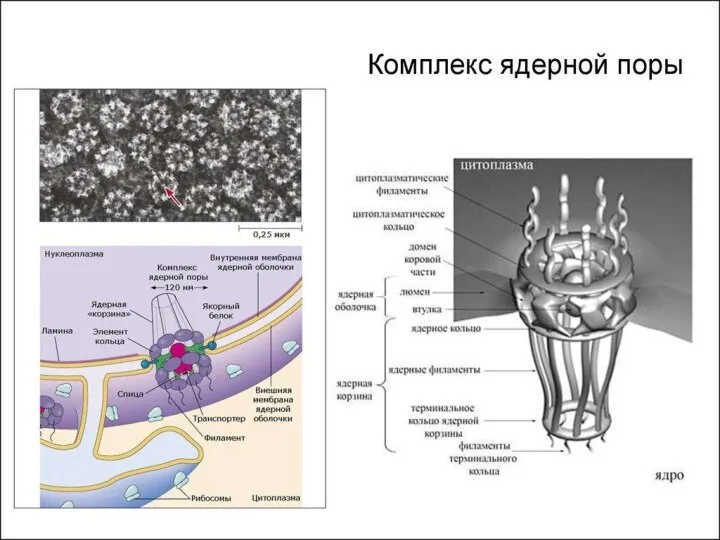
- 9. 1. Ядерная оболочка Состоит из двух мембран липопротеиновой природы Внешняя мембрана продолжается в мембраны шЭПС, на
- 11. Ядерная ламина (1) Поддерживает целостность и форму ядра, Отвечает за дезинтеграцию и реорганизацию ядерной оболочки в
- 12. Ядерная ламина (2)
- 13. LMNA - нормальный ген
- 14. Ядерная ламина, болезни и старение человека Мутации гена ламина A/C (LMNA) у пациентов с дилатационной кардиомиопатией
- 15. 2. Кариоплазма и ядерный матрикс внутреннее содержимое ядра, содержит жидкую фракцию и матрикс жидкая фракция ядра
- 16. Ядерный матрикс Собственно матрикс (нерастворимые белки – негистоны, ферменты, нуклеоплазмины, матрины и др.) Внутриядерная сеть фибриллярных
- 17. Функции ядерного матрикса Обеспечивает форму и внутреннюю организацию ядра Пространственное распределение хроматина и его конденсация(упаковка) Передвижение
- 18. 3. Ядрышко Ядрышковый организатор – фрагменты ДНК содержащие информацию о рРНК; рРНК: первичные транскрипты 45S; рРНК
- 19. Ядрышко – область биогенеза рибосом: Транскрипция рибосомных генов и синтез про-рРНК (45S); Процессинг про-рРНК 45S с
- 21. Нобелевская премия в области химии 2009 года «за изучение структуры и функций рибосом“ Venkatraman Ramakrishnan, MRC
- 22. 4. ХРОМАТИН ХРОМОСОМЫ Хроматин – это частично упакованный генетический материал в интерфазе; Хромосома – максимально упакованный
- 23. Хромосомная ДНК – содержит, передает и реализует ГИ; Длинные, линейные молекулы; Гетерогенные молекулы: Кодирующие последовательности (гены):
- 24. Гистоновые белки: Глобулярные Богатые Liz и Arg; 5 классов: H1, H2A, H2B, H3, H4 Функция: Упаковка
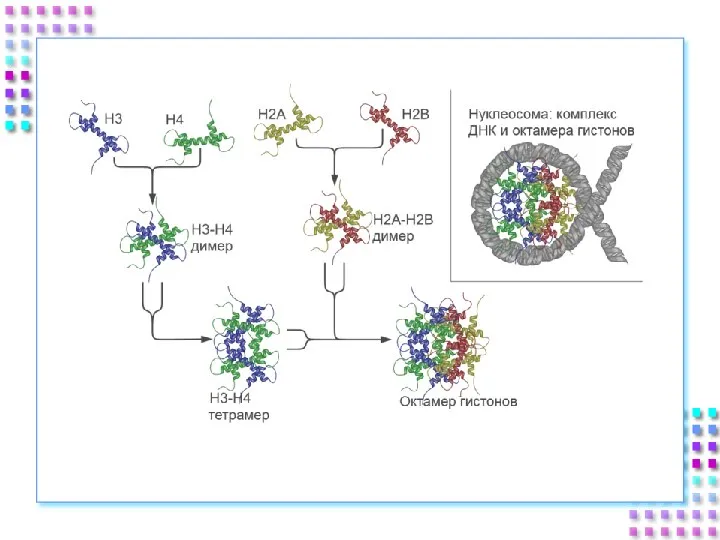
- 25. Гистоновые белки ДНК + гистоны = нуклеосома a. Нуклеосомное ядро: 2H2A 2H2B 2H3 2H4 b. 200
- 27. Негистоновые белки Крайне гетерогенные белки; Ядерные ферменты для: репликации (праймаза, ДНК-пол., ДНК-лиг.); репарации (эндонуклеазы, ДНК-пол); транскрипции
- 28. Helix-Turn-Helix "спираль-поворот- спираль". Zink finger "цинковые пальцы".
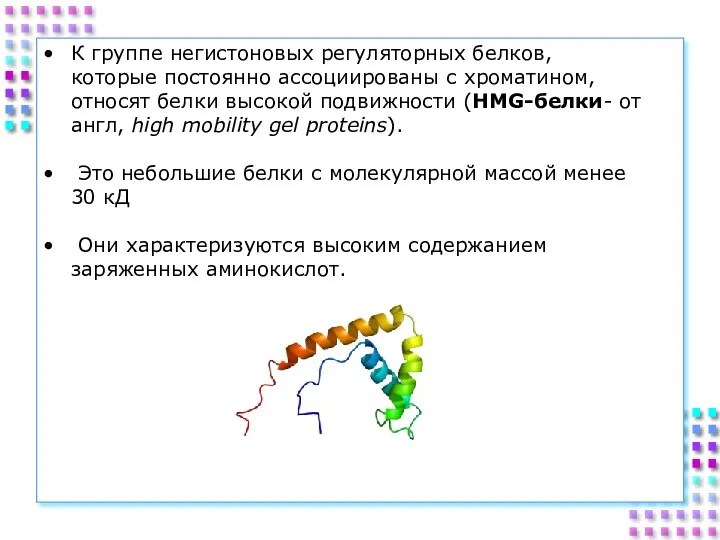
- 29. К группе негистоновых регуляторных белков, которые постоянно ассоциированы с хроматином, относят белки высокой подвижности (HMG-белки- от
- 30. Хромосомная РНК Первичные транскрипты микроРНК в составе ядерных ферментов
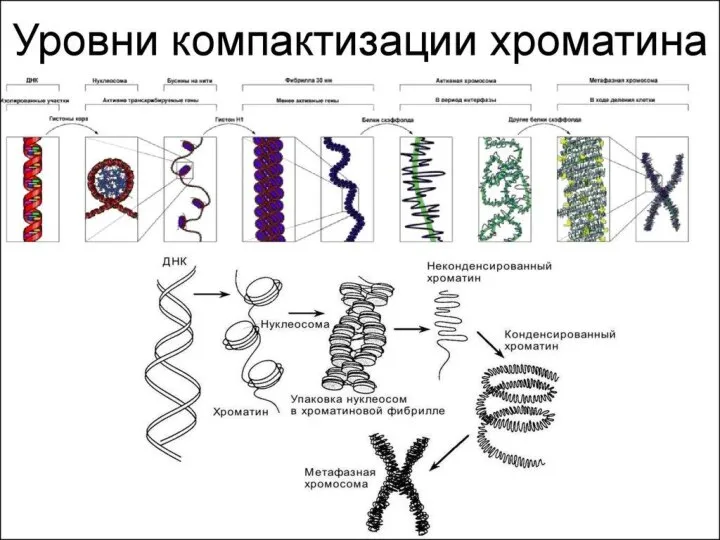
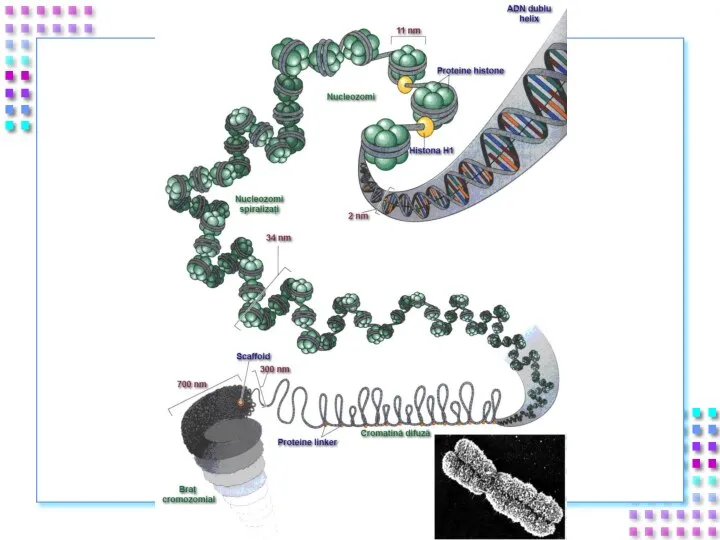
- 31. Уровни упаковки ДНК I уровень – нуклеосомный = нити хроматина; II уровень – соленоид = нити
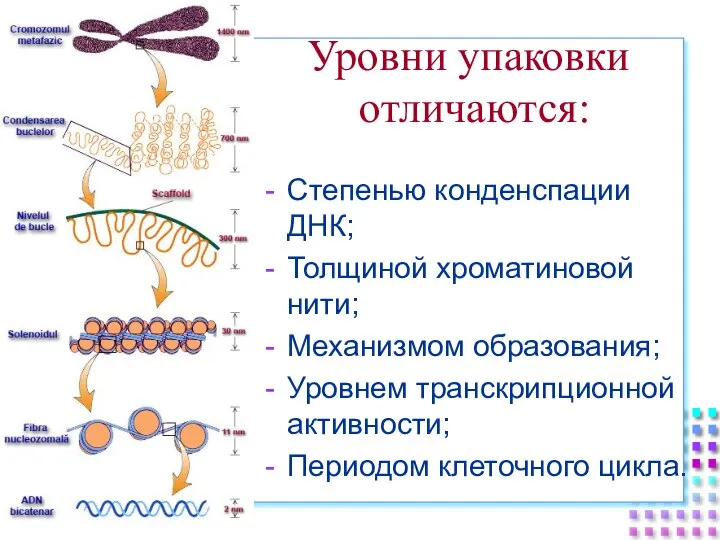
- 33. Степенью конденспации ДНК; Толщиной хроматиновой нити; Механизмом образования; Уровнем транскрипционной активности; Периодом клеточного цикла. Уровни упаковки
- 34. I уровень- нити хроматина: Полинуклеосомная нить 11nm; Степень конденсации – x 6 раз; Механизм образования: Взаимодействие
- 36. Гистоны в октамере имеют подвижный N-концевой фрагмент («хвост») из 20 аминокислот, который выступает из нуклеосом и
- 37. I уровень- нити хроматина 10 nm Сокращение гистона при старении
- 38. Некоторые модификации гистонов (фосфорилированиеНекоторые модификации гистонов (фосфорилирование и ацетилированиеНекоторые модификации гистонов (фосфорилирование и ацетилирование)локализованы преимущественно на
- 39. Биологи нашли слабое место в ДНК человека и других животных Ученые Института биомедицинских исследований обнаружили, что
- 40. Метилирование H3 (Lys4) – активная экспрессия гена Метилирование H3 (Lys9) – атенуация транскрипции Ацетилирование гистонов –
- 41. Пора переходить от слов к делу… Как повлиять на ацетилирование гистонов для защиты от стресса и
- 42. II уровень – соленоид Нить хроматина 30 nm; Степень конденсации – x40 раз; Механизм образования: Фосфорилирование
- 44. Нить хроматина Соленоид– 30 nm Нуклеосомная нить – 10 nm
- 45. III уровень- петли Петли большие и малые, диффузные или компактные, толщина – 300 nm; Степень конденсации
- 47. !!! SAR (MAR) - (Scaffold /Matrix Associated Region) – специфические сайты ДНК ответственные за присоединение к
- 48. IV уровень – метафазная хромосома Наивысший уровень упаковки ДНК – x10000 раз; Толщина хроматиды ~700nm; Механизм
- 51. Период клеточного цикла и степень конденсации хроматина Интерфаза: Период G1; Период S; Период G2; Митоз: Профаза
- 52. Функциональная классификация хроматина: Эухроматин: Активные фрагменты ДНК; Содержит кодогенную ДНК. Гетерохроматин: Неактивные фрагменты ДНК; Содержит некодогенную
- 53. Гетерохроматин Конститутивный: Некодирующая, репетитивная ДНК; Ценромеры, теломеры, сателлиты, спейсеры. Факультативный: Кодирующая, неактивная ДНК; Может переходить в
- 54. Центромера = первичная перетяжка Теломера
- 55. Центромера Повторяющиеся последовательности, богатые A/T Конститутивный гетерохроматин H3 замещен CENP-A Связывает хроматиды до анафазы митоза (коэзин)
- 56. Теломеры Участки на концах хромосом, которые не несут генетической информации Каждая клетка человека имеет 92 теломеры
- 57. https://nauchforum.ru/archive/MNF_nature/5(23).pdf (дата обращения: 03.10.2018) В 1930 году Герман Меллер и Барбара Мак-Клинток пришли к заключению, что
- 60. https://nauchforum.ru/archive/MNF_nature/5(23).pdf (дата обращения: 03.10.2018) В 1971г. Алексей Оловников предложил теорию, которая объясняла механизм работы такого «счетчика»
- 61. Теломераза – удивительный фермент Способна достраивать короткие теломерные участки Состоит из РНК-матрицы и белкового компонента
- 62. Молекулярная организация теломеров Последовательности богатая G/C Образует петли на концах хромосом У человека теломеры содержат единственный
- 63. The Nobel Prize in Medicine 2009 "for the discovery of how chromosomes are protected by telomeres
- 64. Функции теломер Обеспечивают целостность хромосом и препятствуют слипанию их концов Защищают концы хромосом от действия экзонуклеаз
- 66. Скачать презентацию
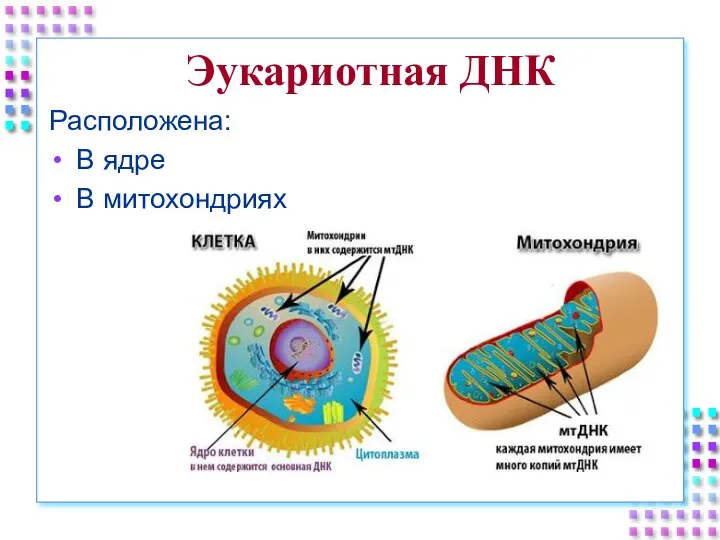
Слайд 3Ядерная ДНК
Длинные линейные молекулы;
В виде хроматина ↔ хромосом;
Около 10% ДНК – кодирующая,
Ядерная ДНК
Длинные линейные молекулы;
В виде хроматина ↔ хромосом;
Около 10% ДНК – кодирующая,

Слайд 4Митохондриальная ДНК
Небольшие кольцевые молекулы
Количество – от 2-х до 10 в одной
Митохондриальная ДНК
Небольшие кольцевые молекулы
Количество – от 2-х до 10 в одной

Слайд 5Ядро на разных этапах клеточного цикла
Ядро на разных этапах клеточного цикла
Слайд 6Функции ядра
Хранение и функциональная упаковка ДНК:
Эухроматин – активная ГИ;
Гетерохроматин – неактивная ДНК.
Контроль
Функции ядра
Хранение и функциональная упаковка ДНК:
Эухроматин – активная ГИ;
Гетерохроматин – неактивная ДНК.
Контроль

Слайд 7Строение ядра
1) Ядерная оболочка:
Внешняя ядерная мембрана
Внутренняя ядерная мембрана
Перинуклеарное пространство
Поровый комплекс
Ядерная ламина
2) Кариоплазма
Лабильная
Строение ядра
1) Ядерная оболочка:
Внешняя ядерная мембрана
Внутренняя ядерная мембрана
Перинуклеарное пространство
Поровый комплекс
Ядерная ламина
2) Кариоплазма
Лабильная

Слайд 91. Ядерная оболочка
Состоит из двух мембран липопротеиновой природы
Внешняя мембрана продолжается в мембраны
1. Ядерная оболочка
Состоит из двух мембран липопротеиновой природы
Внешняя мембрана продолжается в мембраны

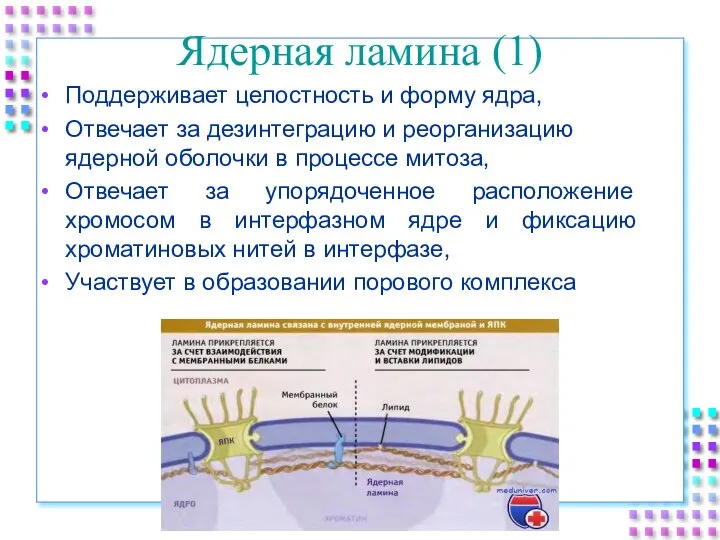
Слайд 11Ядерная ламина (1)
Поддерживает целостность и форму ядра,
Отвечает за дезинтеграцию и реорганизацию ядерной
Ядерная ламина (1)
Поддерживает целостность и форму ядра,
Отвечает за дезинтеграцию и реорганизацию ядерной
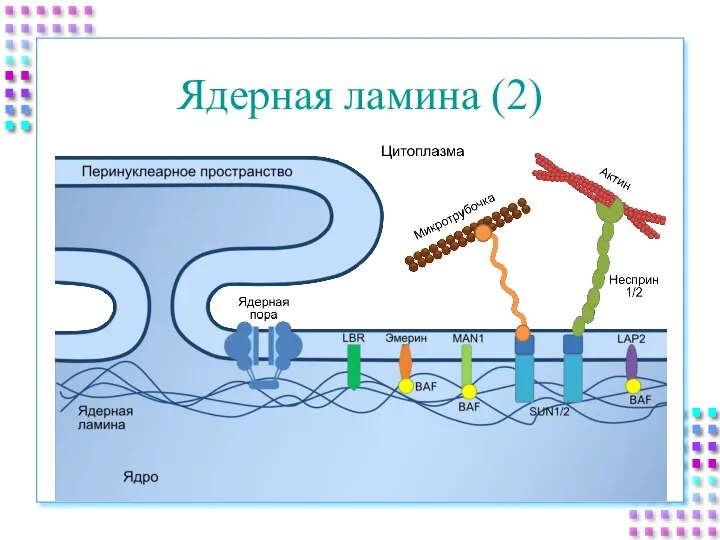
Слайд 12Ядерная ламина (2)
Ядерная ламина (2)
Слайд 13LMNA - нормальный ген
LMNA - нормальный ген
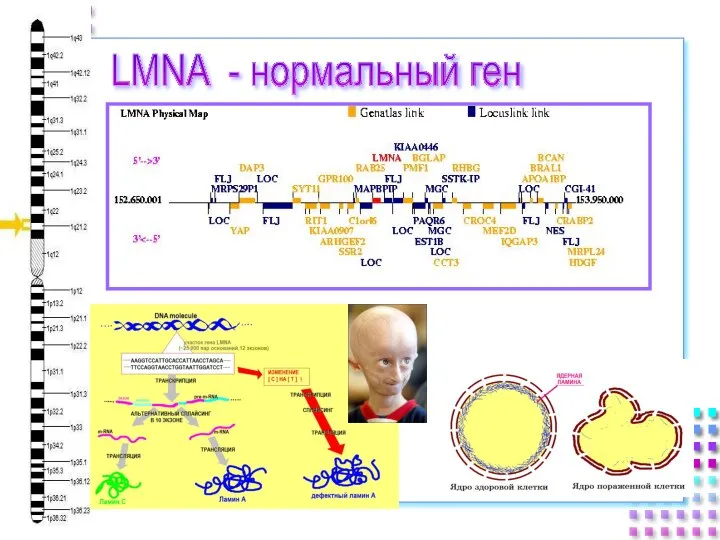
Слайд 14Ядерная ламина, болезни и старение человека
Мутации гена ламина A/C (LMNA) у пациентов
Ядерная ламина, болезни и старение человека
Мутации гена ламина A/C (LMNA) у пациентов

Слайд 152. Кариоплазма и ядерный матрикс
внутреннее содержимое ядра,
содержит жидкую фракцию и матрикс
жидкая фракция
2. Кариоплазма и ядерный матрикс
внутреннее содержимое ядра,
содержит жидкую фракцию и матрикс
жидкая фракция

Слайд 16 Ядерный матрикс
Собственно матрикс (нерастворимые белки – негистоны, ферменты, нуклеоплазмины, матрины и
Ядерный матрикс
Собственно матрикс (нерастворимые белки – негистоны, ферменты, нуклеоплазмины, матрины и

Слайд 17Функции ядерного матрикса
Обеспечивает форму и внутреннюю организацию ядра
Пространственное распределение хроматина и его
Функции ядерного матрикса
Обеспечивает форму и внутреннюю организацию ядра
Пространственное распределение хроматина и его

Слайд 183. Ядрышко
Ядрышковый организатор – фрагменты ДНК содержащие информацию о рРНК;
рРНК: первичные транскрипты
3. Ядрышко
Ядрышковый организатор – фрагменты ДНК содержащие информацию о рРНК;
рРНК: первичные транскрипты

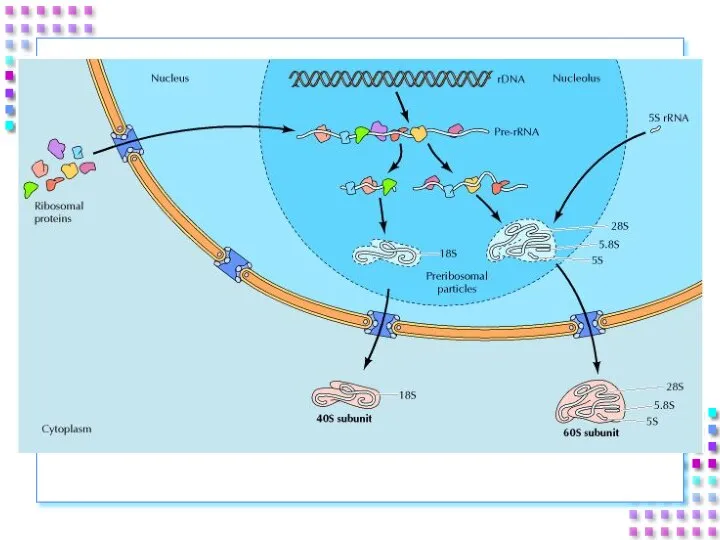
Слайд 19Ядрышко –
область биогенеза рибосом:
Транскрипция рибосомных генов и синтез про-рРНК (45S);
Процессинг про-рРНК
Ядрышко –
область биогенеза рибосом:
Транскрипция рибосомных генов и синтез про-рРНК (45S);
Процессинг про-рРНК

Слайд 21Нобелевская премия в области химии 2009 года
«за изучение структуры и функций рибосом“
Venkatraman
Нобелевская премия в области химии 2009 года
«за изучение структуры и функций рибосом“
Venkatraman

Слайд 224. ХРОМАТИН ХРОМОСОМЫ
Хроматин – это частично упакованный генетический материал в интерфазе;
Хромосома –
4. ХРОМАТИН ХРОМОСОМЫ
Хроматин – это частично упакованный генетический материал в интерфазе;
Хромосома –

Слайд 23Хромосомная ДНК – содержит, передает и реализует ГИ;
Длинные, линейные молекулы;
Гетерогенные молекулы:
Кодирующие последовательности
Хромосомная ДНК – содержит, передает и реализует ГИ;
Длинные, линейные молекулы;
Гетерогенные молекулы:
Кодирующие последовательности

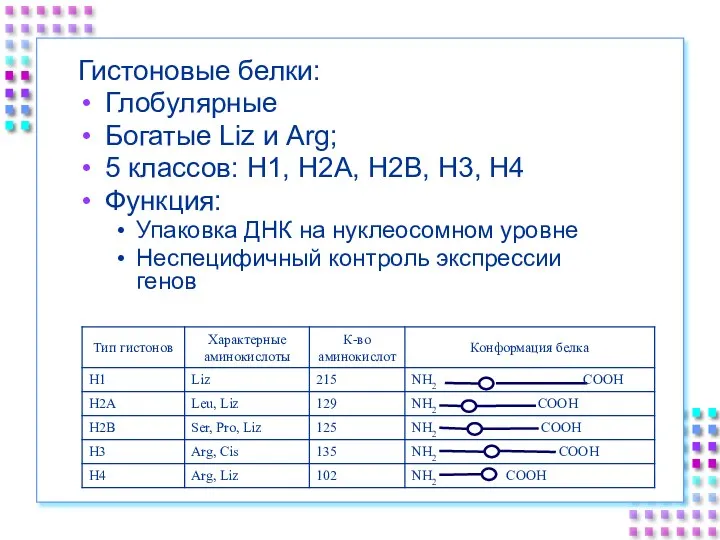
Слайд 24Гистоновые белки:
Глобулярные
Богатые Liz и Arg;
5 классов: H1, H2A, H2B, H3, H4
Функция:
Упаковка
Гистоновые белки:
Глобулярные
Богатые Liz и Arg;
5 классов: H1, H2A, H2B, H3, H4
Функция:
Упаковка
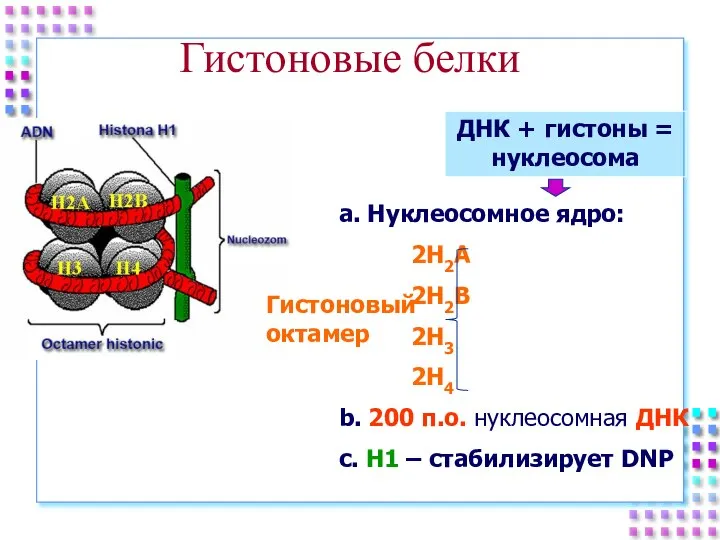
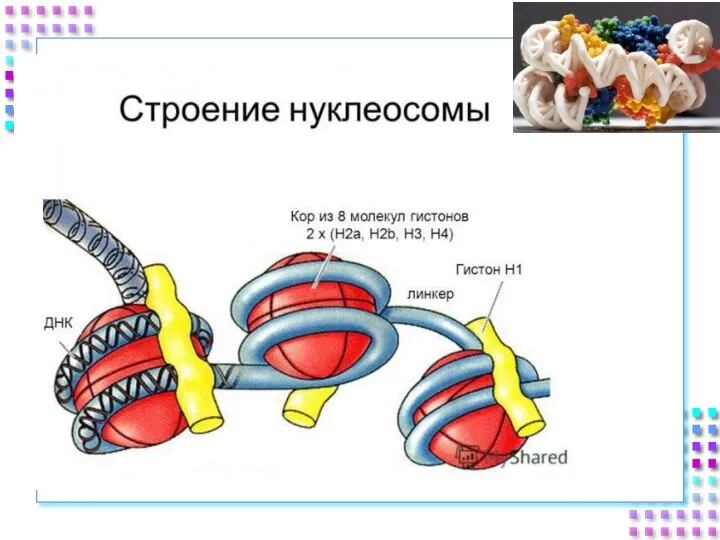
Слайд 25Гистоновые белки
ДНК + гистоны = нуклеосома
a. Нуклеосомное ядро:
2H2A
2H2B
2H3
2H4
b. 200 п.о.
Гистоновые белки
ДНК + гистоны = нуклеосома
a. Нуклеосомное ядро:
2H2A
2H2B
2H3
2H4
b. 200 п.о.
Слайд 27Негистоновые белки
Крайне гетерогенные белки;
Ядерные ферменты для:
репликации (праймаза, ДНК-пол., ДНК-лиг.);
репарации (эндонуклеазы, ДНК-пол);
транскрипции (РНК-пол.,
Негистоновые белки
Крайне гетерогенные белки;
Ядерные ферменты для:
репликации (праймаза, ДНК-пол., ДНК-лиг.);
репарации (эндонуклеазы, ДНК-пол);
транскрипции (РНК-пол.,

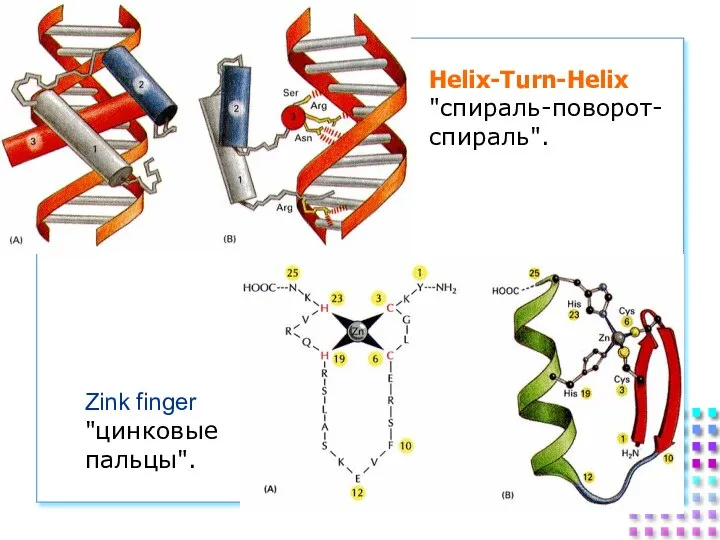
Слайд 28Helix-Turn-Helix
"спираль-поворот-
спираль".
Zink finger
"цинковые пальцы".
Helix-Turn-Helix
"спираль-поворот-
спираль".
Zink finger
"цинковые пальцы".
Слайд 29К группе негистоновых регуляторных белков, которые постоянно ассоциированы с хроматином, относят белки
К группе негистоновых регуляторных белков, которые постоянно ассоциированы с хроматином, относят белки
Слайд 30Хромосомная РНК
Первичные транскрипты
микроРНК в составе ядерных ферментов
Хромосомная РНК
Первичные транскрипты
микроРНК в составе ядерных ферментов

Слайд 31Уровни упаковки ДНК
I уровень – нуклеосомный = нити хроматина;
II уровень – соленоид
Уровни упаковки ДНК
I уровень – нуклеосомный = нити хроматина;
II уровень – соленоид

Слайд 33Степенью конденспации ДНК;
Толщиной хроматиновой нити;
Механизмом образования;
Уровнем транскрипционной активности;
Периодом клеточного цикла.
Уровни упаковки
отличаются:
Степенью конденспации ДНК;
Толщиной хроматиновой нити;
Механизмом образования;
Уровнем транскрипционной активности;
Периодом клеточного цикла.
Уровни упаковки
отличаются:
Слайд 34I уровень- нити хроматина:
Полинуклеосомная нить 11nm;
Степень конденсации – x 6 раз;
Механизм образования:
Взаимодействие
I уровень- нити хроматина:
Полинуклеосомная нить 11nm;
Степень конденсации – x 6 раз;
Механизм образования:
Взаимодействие

Слайд 36Гистоны в октамере имеют подвижный N-концевой фрагмент («хвост») из 20 аминокислот, который
Гистоны в октамере имеют подвижный N-концевой фрагмент («хвост») из 20 аминокислот, который

Слайд 37I уровень- нити хроматина
10 nm
Сокращение гистона при старении
I уровень- нити хроматина
10 nm
Сокращение гистона при старении

Слайд 38Некоторые модификации гистонов (фосфорилированиеНекоторые модификации гистонов (фосфорилирование и ацетилированиеНекоторые модификации гистонов (фосфорилирование и ацетилирование)локализованы преимущественно на
Некоторые модификации гистонов (фосфорилированиеНекоторые модификации гистонов (фосфорилирование и ацетилированиеНекоторые модификации гистонов (фосфорилирование и ацетилирование)локализованы преимущественно на

Слайд 39Биологи нашли слабое место в ДНК человека и других животных
Ученые Института
Биологи нашли слабое место в ДНК человека и других животных
Ученые Института

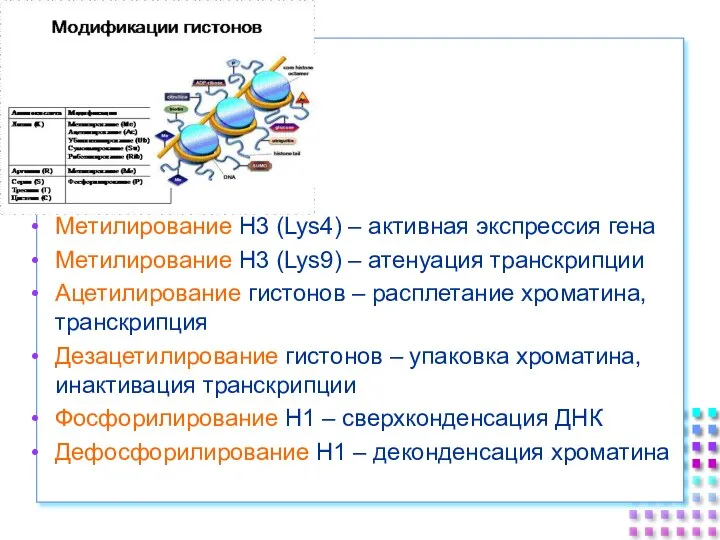
Слайд 40Метилирование H3 (Lys4) – активная экспрессия гена
Метилирование H3 (Lys9) – атенуация транскрипции
Ацетилирование
Метилирование H3 (Lys4) – активная экспрессия гена
Метилирование H3 (Lys9) – атенуация транскрипции
Ацетилирование
Слайд 41Пора переходить от слов к делу…
Как повлиять на ацетилирование гистонов для защиты
Пора переходить от слов к делу…
Как повлиять на ацетилирование гистонов для защиты

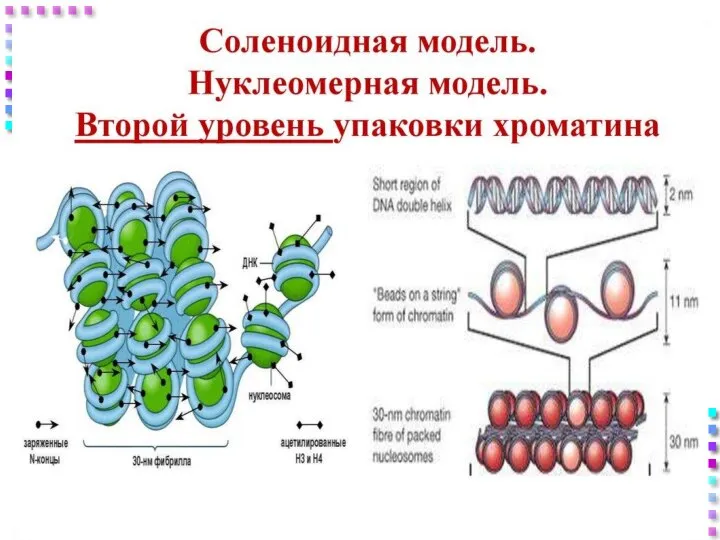
Слайд 42II уровень – соленоид
Нить хроматина 30 nm;
Степень конденсации – x40 раз;
Механизм образования:
Фосфорилирование
II уровень – соленоид
Нить хроматина 30 nm;
Степень конденсации – x40 раз;
Механизм образования:
Фосфорилирование

Слайд 44Нить хроматина
Соленоид– 30 nm
Нуклеосомная нить
– 10 nm
Нить хроматина
Соленоид– 30 nm
Нуклеосомная нить
– 10 nm

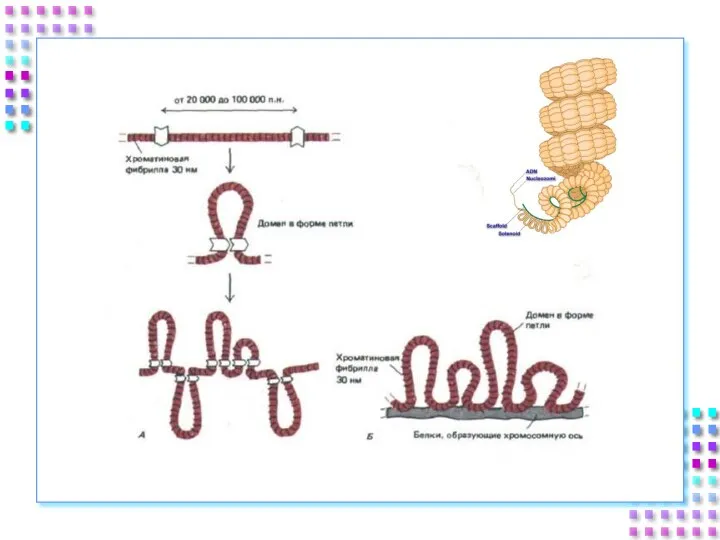
Слайд 45III уровень- петли
Петли большие и малые, диффузные или компактные, толщина – 300
III уровень- петли
Петли большие и малые, диффузные или компактные, толщина – 300

Слайд 47!!! SAR (MAR) - (Scaffold /Matrix Associated Region) – специфические сайты ДНК
!!! SAR (MAR) - (Scaffold /Matrix Associated Region) – специфические сайты ДНК

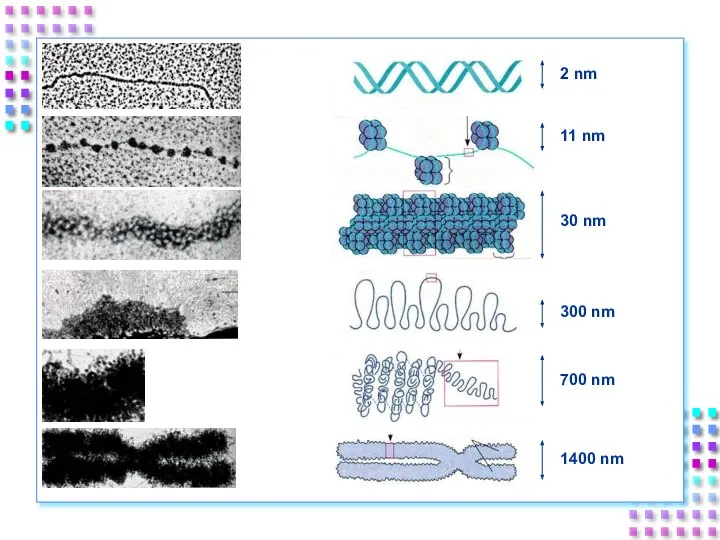
Слайд 48IV уровень –
метафазная хромосома
Наивысший уровень упаковки ДНК – x10000 раз;
Толщина хроматиды
IV уровень –
метафазная хромосома
Наивысший уровень упаковки ДНК – x10000 раз;
Толщина хроматиды

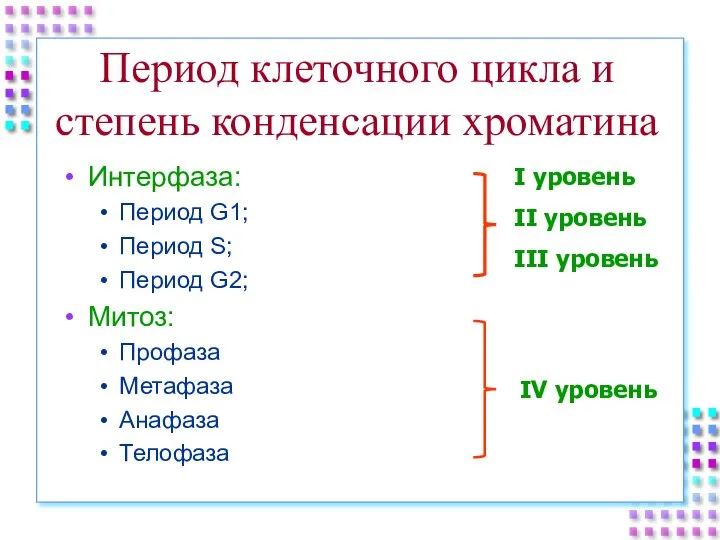
Слайд 51Период клеточного цикла и степень конденсации хроматина
Интерфаза:
Период G1;
Период S;
Период G2;
Митоз:
Профаза
Метафаза
Анафаза
Телофаза
I уровень
II уровень
III
Период клеточного цикла и степень конденсации хроматина
Интерфаза:
Период G1;
Период S;
Период G2;
Митоз:
Профаза
Метафаза
Анафаза
Телофаза
I уровень
II уровень
III
Слайд 52Функциональная классификация хроматина:
Эухроматин:
Активные фрагменты ДНК;
Содержит кодогенную ДНК.
Гетерохроматин:
Неактивные фрагменты ДНК;
Содержит некодогенную
Функциональная классификация хроматина:
Эухроматин:
Активные фрагменты ДНК;
Содержит кодогенную ДНК.
Гетерохроматин:
Неактивные фрагменты ДНК;
Содержит некодогенную

Слайд 53Гетерохроматин
Конститутивный:
Некодирующая, репетитивная ДНК;
Ценромеры, теломеры, сателлиты, спейсеры.
Факультативный:
Кодирующая, неактивная ДНК;
Может переходить в эухроматин;
Может быть
Гетерохроматин
Конститутивный:
Некодирующая, репетитивная ДНК;
Ценромеры, теломеры, сателлиты, спейсеры.
Факультативный:
Кодирующая, неактивная ДНК;
Может переходить в эухроматин;
Может быть

Слайд 54Центромера = первичная перетяжка
Теломера
Центромера = первичная перетяжка
Теломера
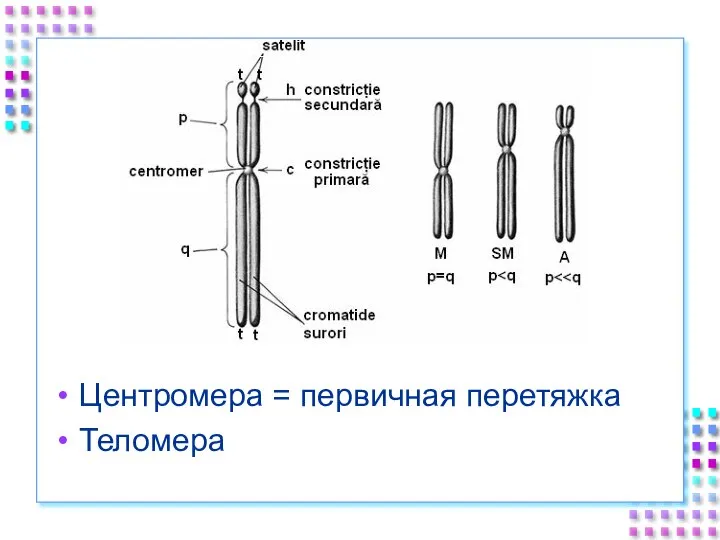
Слайд 55Центромера
Повторяющиеся последовательности, богатые A/T
Конститутивный гетерохроматин
H3 замещен CENP-A
Связывает хроматиды до анафазы митоза (коэзин)
Обеспечивает
Центромера
Повторяющиеся последовательности, богатые A/T
Конститутивный гетерохроматин
H3 замещен CENP-A
Связывает хроматиды до анафазы митоза (коэзин)
Обеспечивает

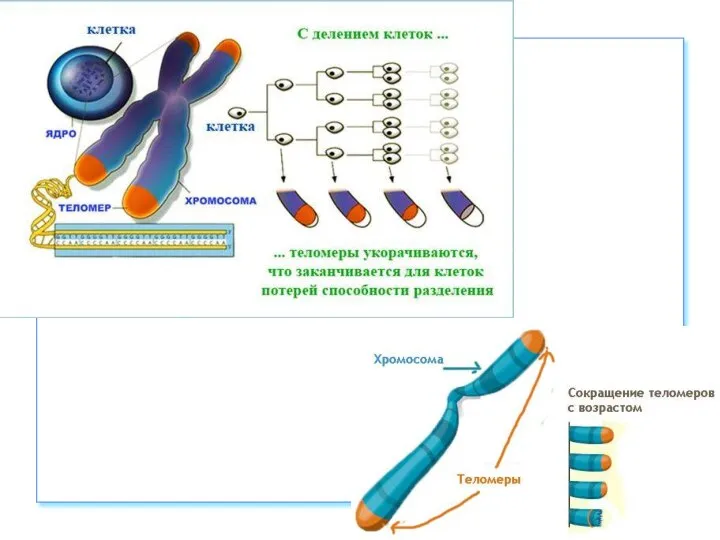
Слайд 56Теломеры
Участки на концах хромосом, которые не несут генетической информации
Каждая клетка человека
Теломеры
Участки на концах хромосом, которые не несут генетической информации
Каждая клетка человека

Слайд 57https://nauchforum.ru/archive/MNF_nature/5(23).pdf (дата обращения: 03.10.2018)
В 1930 году Герман Меллер и Барбара Мак-Клинток пришли к заключению, что
https://nauchforum.ru/archive/MNF_nature/5(23).pdf (дата обращения: 03.10.2018)
В 1930 году Герман Меллер и Барбара Мак-Клинток пришли к заключению, что

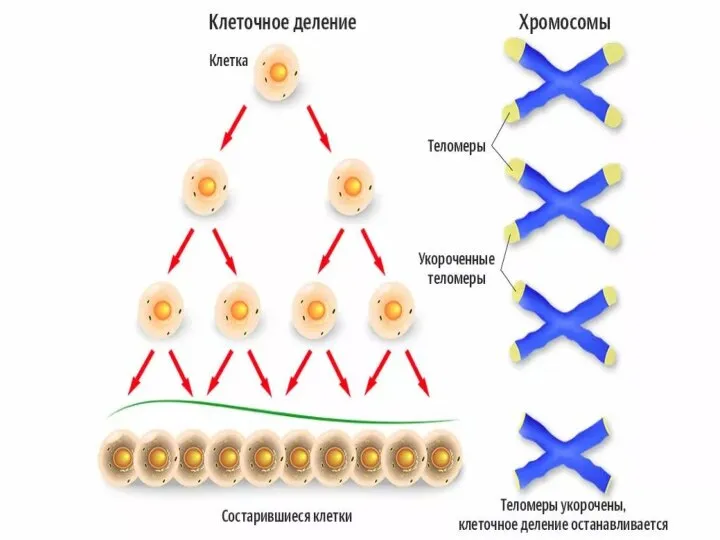
Слайд 60https://nauchforum.ru/archive/MNF_nature/5(23).pdf (дата обращения: 03.10.2018)
В 1971г. Алексей Оловников предложил теорию, которая объясняла механизм работы такого
https://nauchforum.ru/archive/MNF_nature/5(23).pdf (дата обращения: 03.10.2018)
В 1971г. Алексей Оловников предложил теорию, которая объясняла механизм работы такого

Слайд 61Теломераза – удивительный фермент
Способна достраивать короткие теломерные участки
Состоит из РНК-матрицы и белкового
Теломераза – удивительный фермент
Способна достраивать короткие теломерные участки
Состоит из РНК-матрицы и белкового

Слайд 62Молекулярная организация теломеров
Последовательности богатая G/C
Образует петли на концах хромосом
У человека теломеры содержат
Молекулярная организация теломеров
Последовательности богатая G/C
Образует петли на концах хромосом
У человека теломеры содержат

Слайд 63The Nobel Prize in Medicine 2009
"for the discovery of how chromosomes are
The Nobel Prize in Medicine 2009
"for the discovery of how chromosomes are

Слайд 64Функции теломер
Обеспечивают целостность хромосом и препятствуют слипанию их концов
Защищают концы хромосом от
Функции теломер
Обеспечивают целостность хромосом и препятствуют слипанию их концов
Защищают концы хромосом от

 Гусиный лук – символ города Бирюча
Гусиный лук – символ города Бирюча Кровообращение. 8 класс
Кровообращение. 8 класс Углеводы. Термин углеводы
Углеводы. Термин углеводы Галерея цветов
Галерея цветов Микронезия
Микронезия Сравнение свойств организма человека и животных
Сравнение свойств организма человека и животных Сердце – пламенный мотор
Сердце – пламенный мотор Атмосферний тиск
Атмосферний тиск Селекция
Селекция Тип Иглокожие 7 класс Учитель: Типикина Т.И.
Тип Иглокожие 7 класс Учитель: Типикина Т.И. Клеточный уровень. Клеточная теория. 9 класс
Клеточный уровень. Клеточная теория. 9 класс Основные критические фазы возрастного развития (большие и малые кризисы)
Основные критические фазы возрастного развития (большие и малые кризисы) Отъемно-доливной метод культивирования. Тубулярная культура. Хемостатный процесс непрерывного культивирования микроорганизмов
Отъемно-доливной метод культивирования. Тубулярная культура. Хемостатный процесс непрерывного культивирования микроорганизмов Виртуальный животный мир Омской области
Виртуальный животный мир Омской области Дыхание у растений
Дыхание у растений Борщевик – зеленый террорист
Борщевик – зеленый террорист Пищеварение. Функции пищеварительного тракта
Пищеварение. Функции пищеварительного тракта Крокодилы
Крокодилы Презентация на тему Животные космонавты
Презентация на тему Животные космонавты  Ожившие цветы. Часть 2
Ожившие цветы. Часть 2 Серое вещество спинного мозга
Серое вещество спинного мозга Зимующие птицы Ярославской области
Зимующие птицы Ярославской области Одува́нчик. Строение одуванчика
Одува́нчик. Строение одуванчика Антропологический состав населения мира
Антропологический состав населения мира Ара. Ара пайдалану
Ара. Ара пайдалану Ткани. Животные ткани
Ткани. Животные ткани Растения-хищники и растения-паразиты
Растения-хищники и растения-паразиты Мать - и - мачеха
Мать - и - мачеха