- Биоэлектрогенез

Содержание
- 2. Сокращение ноги лягушки в ответ на электрический разряд положило начало поискам электричества в организме человека и
- 3. Эмиль Дюбуа-Реймон (1818-1896) Основатель научной школы электрофизиологии, заведующий кафедрой физиологии Берлинского университета (1858 г), секретарь Берлинской
- 4. Головоногий моллюск кальмар (схема). Английский зоолог Джон Юнг впервые описал у кальмаров и каракатиц необычайно толстые
- 5. Юлиус Бернштейн (1902): У живых клеток в покое между внутренним содержимым клетки и наружным раствором существует
- 6. Плотность потока вещества jm – это величина, численно равная количеству вещества, перенесенного за единицу времени через
- 7. Уравнение, описывающее электрохимический потенциал: электрохимический потенциал где R — газовая постоянная, Т — абсолютная температура, F
- 8. электрохимический потенциал Na+ Cl- Na+Cl- K1=1.0 Na+Cl- K2=0.1 Уравнение Нернста:
- 9. цитоплазматическая мембрана S. J. Singer and G. L. Nicolson. The Fluid Mosaic Model of the Structure
- 10. Мембрана как конденсатор. Электрические параметры биологических мембран : Удельное электрическое сопротивление (R=107 Ом*м2) Удельная электроемкость (C=0,5*10-2
- 11. Осмос – регуляция водного обмена клетки Эндоцитоз (экзоцитоз) – перенос крупных молекул и небольших частиц Диффузия
- 12. Избирательная проницаемость двойного слоя липидов (полупроницаемость). Пассивный транспорт через мембрану: диффузия без затраты клеткой энергии. пассивный
- 13. Показаны типичные концентрации ионов внутри и вне клетки, и коэффициенты проницаемости ионов через мембрану. Неравномерное распределение
- 14. Внутриклеточный потенциал, при котором суммарный пассивный поток ионов калия равен нулю, называется потенциалом равновесия ионов калия
- 15. где РК, РNa, РCl - коэффициенты проницаемости мембраны для ионов, [K] , [Na], [Cl] - концентрация
- 16. Имеет место в том случае, когда перенос осуществляется против градиента концентрации. Такой перенос требует затраты энергии
- 17. Схема, иллюстрирующая вклад пассивных ионных токов и активного ионного транспорта для поддержания нормального потенциала покоя Трансмембранные
- 18. ХОДЖКИН Алан и ХАКСЛИ Эндрю Нобелевская премия по медицине 1963 за открытие механизма возникновения потенциала действия.
- 19. Схематичное изображение идеализированного потенциала действия В покое внутренняя сторона мембраны аксона отрицательно заряжена по сравнению с
- 20. В основе возникновения ПД лежит свойство мембраны возбудимой клетки быстро менять свою проницаемость для определённого вида
- 21. Потенциалозависимость инактивации Na+ системы. Абсцисса (E-ER): отклонение мембранного потенциала от потенциала покоя. Ордината: отношение возникающих максимальных
- 22. A. Ток ионов калия, протекающий через калиевые каналы, чувствителен к изменениям мембранного потенциала. Для сравнения показана
- 23. максимальная частота генерации ПД, где Тп –длительность стимула Тр – длительность рефрактерного периода рефрактерность нервного волокна
- 24. Потенциалы действия, зарегистрированные с помощью внутриклеточных микроэлектродов: а — гигантского аксона кальмара; б — скелетного мышечного
- 25. Невозбудимые клетки способные к поддержанию потенциала покоя, но не способные к генерированию потенциала действия Возбудимые клетки
- 27. Скачать презентацию
Слайд 2Сокращение ноги лягушки в ответ на электрический разряд положило начало поискам электричества
Сокращение ноги лягушки в ответ на электрический разряд положило начало поискам электричества

Слайд 3Эмиль Дюбуа-Реймон
(1818-1896)
Основатель научной школы электрофизиологии, заведующий кафедрой физиологии Берлинского университета (1858 г),
Эмиль Дюбуа-Реймон
(1818-1896)
Основатель научной школы электрофизиологии, заведующий кафедрой физиологии Берлинского университета (1858 г),

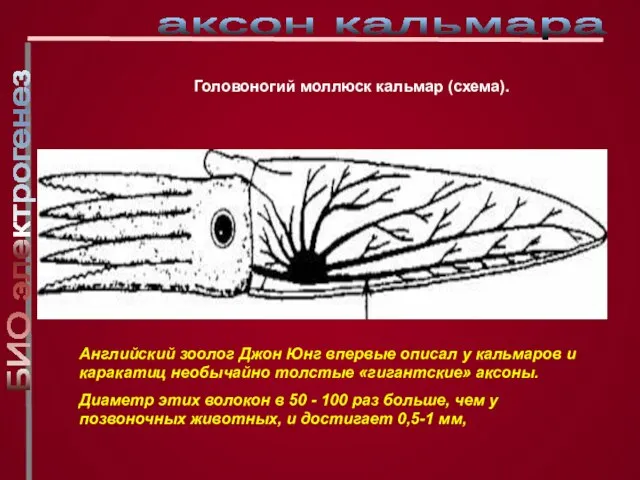
Слайд 4Головоногий моллюск кальмар (схема).
Английский зоолог Джон Юнг впервые описал у кальмаров
Головоногий моллюск кальмар (схема).
Английский зоолог Джон Юнг впервые описал у кальмаров
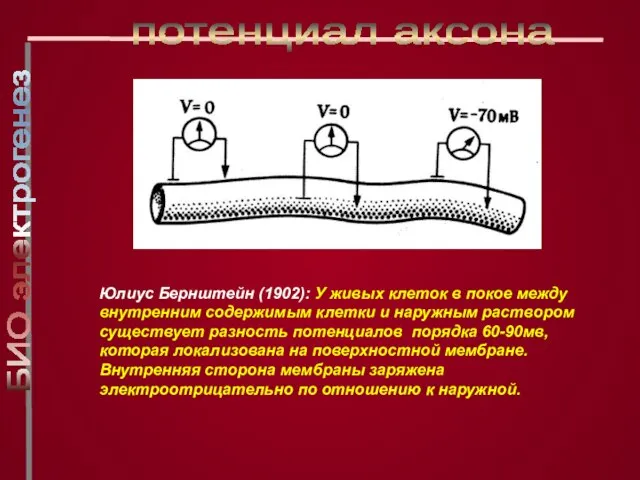
Слайд 5Юлиус Бернштейн (1902): У живых клеток в покое между внутренним содержимым клетки
Юлиус Бернштейн (1902): У живых клеток в покое между внутренним содержимым клетки
Слайд 6Плотность потока вещества jm – это величина, численно равная количеству вещества, перенесенного
Плотность потока вещества jm – это величина, численно равная количеству вещества, перенесенного

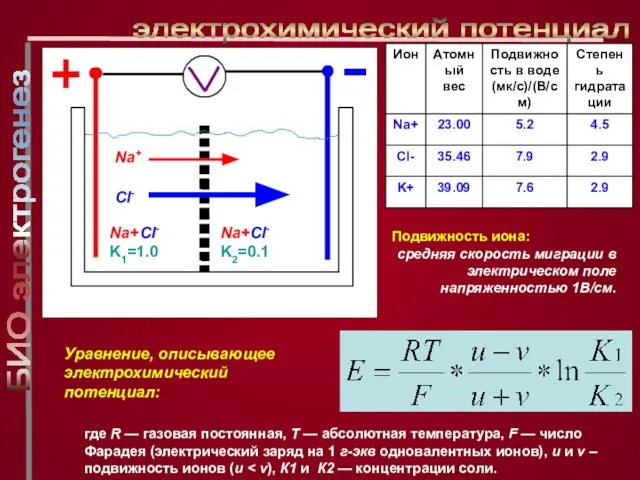
Слайд 7Уравнение, описывающее электрохимический потенциал:
электрохимический потенциал
где R — газовая постоянная, Т — абсолютная
Уравнение, описывающее электрохимический потенциал:
электрохимический потенциал
где R — газовая постоянная, Т — абсолютная
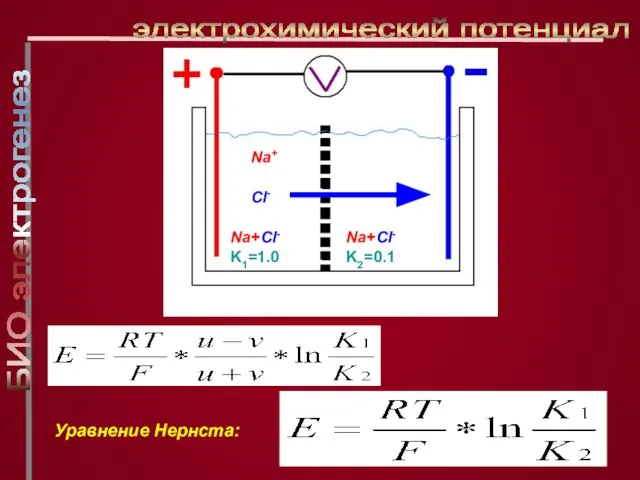
Слайд 8электрохимический потенциал
Na+
Cl-
Na+Cl- K1=1.0
Na+Cl- K2=0.1
Уравнение Нернста:
электрохимический потенциал
Na+
Cl-
Na+Cl- K1=1.0
Na+Cl- K2=0.1
Уравнение Нернста:
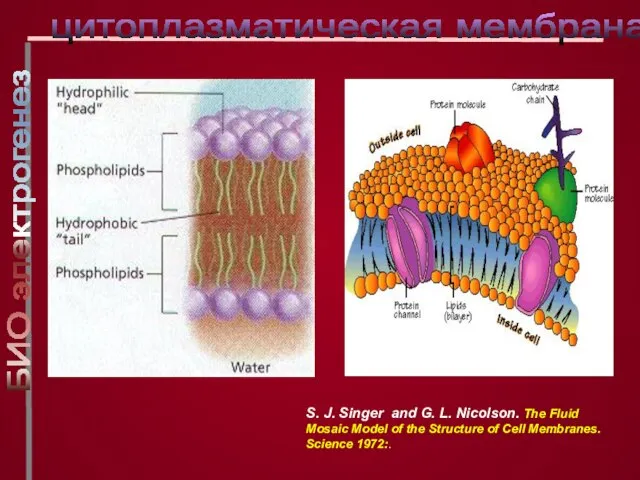
Слайд 9цитоплазматическая мембрана
S. J. Singer and G. L. Nicolson. The Fluid Mosaic Model
цитоплазматическая мембрана
S. J. Singer and G. L. Nicolson. The Fluid Mosaic Model
Слайд 10Мембрана как конденсатор.
Электрические параметры биологических мембран :
Удельное электрическое сопротивление (R=107 Ом*м2)
Мембрана как конденсатор.
Электрические параметры биологических мембран :
Удельное электрическое сопротивление (R=107 Ом*м2)

Слайд 11Осмос – регуляция водного обмена клетки
Эндоцитоз (экзоцитоз) – перенос крупных молекул и
Осмос – регуляция водного обмена клетки
Эндоцитоз (экзоцитоз) – перенос крупных молекул и

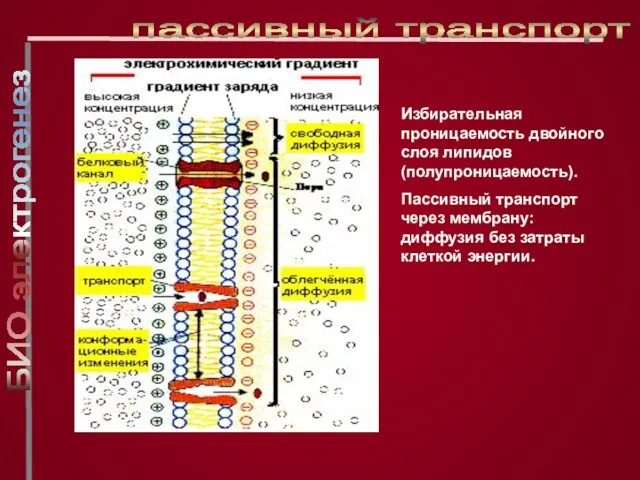
Слайд 12Избирательная проницаемость двойного слоя липидов (полупроницаемость).
Пассивный транспорт через мембрану: диффузия без
Избирательная проницаемость двойного слоя липидов (полупроницаемость).
Пассивный транспорт через мембрану: диффузия без
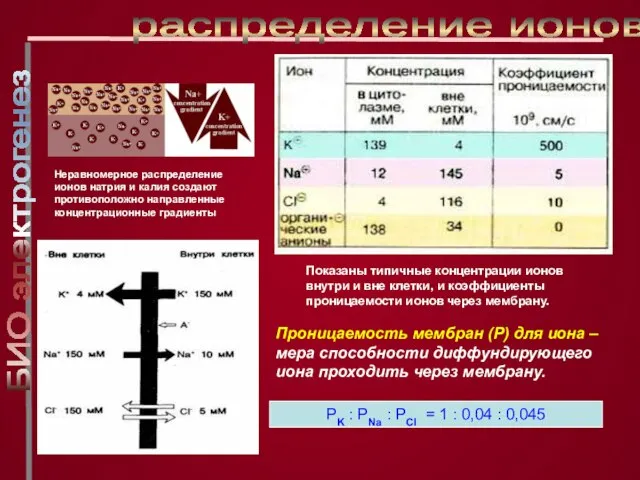
Слайд 13Показаны типичные концентрации ионов внутри и вне клетки, и коэффициенты проницаемости ионов
Показаны типичные концентрации ионов внутри и вне клетки, и коэффициенты проницаемости ионов
Слайд 14Внутриклеточный потенциал, при котором суммарный пассивный поток ионов калия равен нулю, называется
Внутриклеточный потенциал, при котором суммарный пассивный поток ионов калия равен нулю, называется
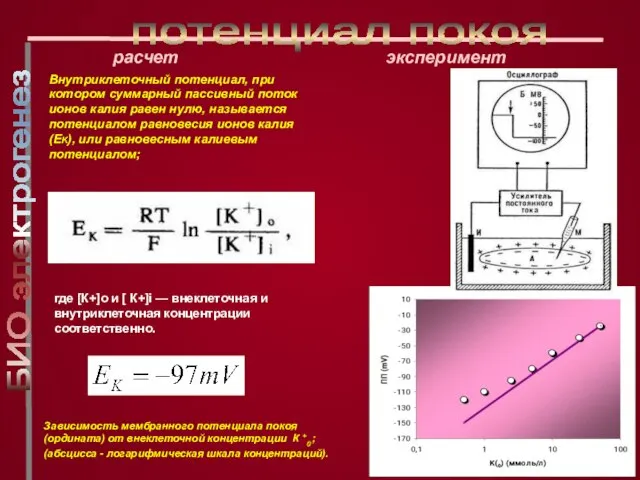
Слайд 15где РК, РNa, РCl - коэффициенты проницаемости мембраны для ионов,
[K] ,
где РК, РNa, РCl - коэффициенты проницаемости мембраны для ионов,
[K] ,
![где РК, РNa, РCl - коэффициенты проницаемости мембраны для ионов, [K] ,](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/896116/slide-14.jpg)
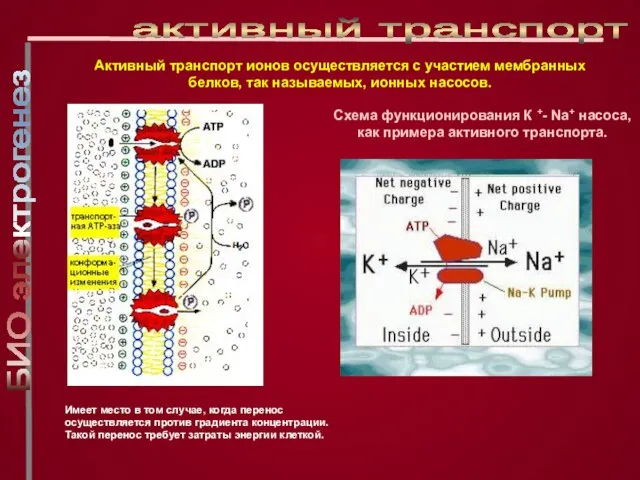
Слайд 16Имеет место в том случае, когда перенос осуществляется против градиента концентрации. Такой
Имеет место в том случае, когда перенос осуществляется против градиента концентрации. Такой
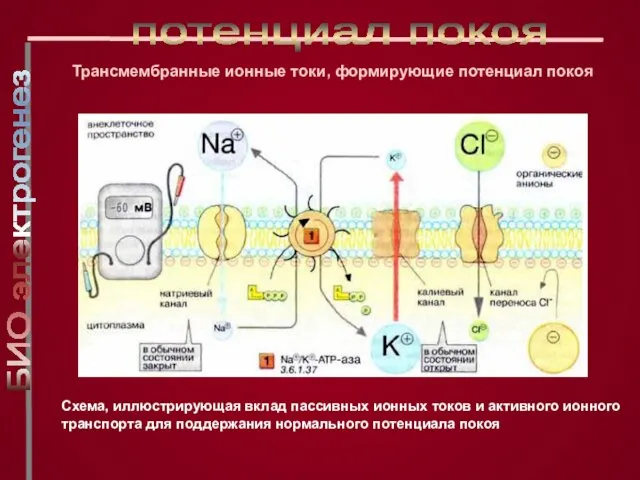
Слайд 17Схема, иллюстрирующая вклад пассивных ионных токов и активного ионного транспорта для поддержания
Схема, иллюстрирующая вклад пассивных ионных токов и активного ионного транспорта для поддержания
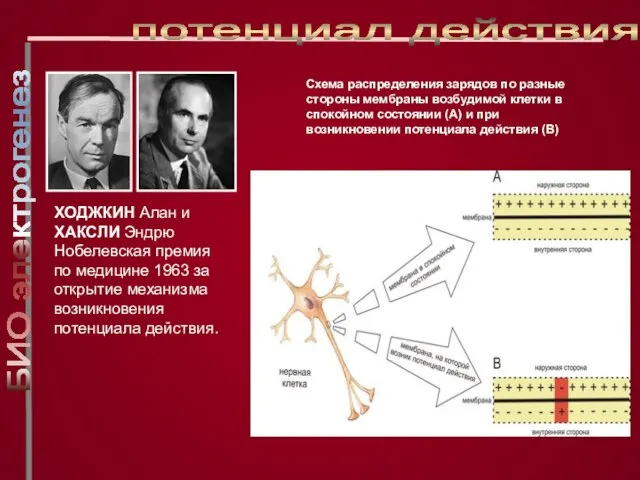
Слайд 18ХОДЖКИН Алан и ХАКСЛИ Эндрю
Нобелевская премия по медицине 1963 за открытие
ХОДЖКИН Алан и ХАКСЛИ Эндрю
Нобелевская премия по медицине 1963 за открытие
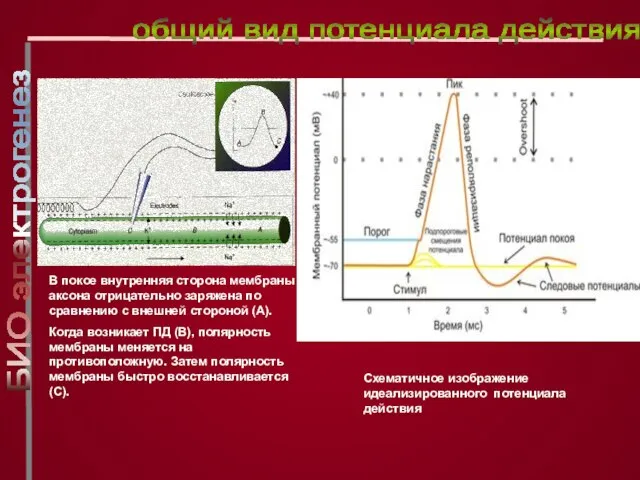
Слайд 19Схематичное изображение идеализированного потенциала действия
В покое внутренняя сторона мембраны аксона отрицательно заряжена
Схематичное изображение идеализированного потенциала действия
В покое внутренняя сторона мембраны аксона отрицательно заряжена
Слайд 20В основе возникновения ПД лежит свойство мембраны возбудимой клетки быстро менять свою
В основе возникновения ПД лежит свойство мембраны возбудимой клетки быстро менять свою

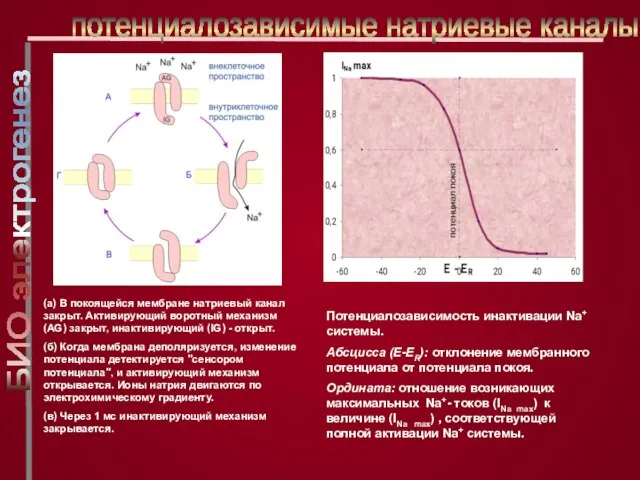
Слайд 21Потенциалозависимость инактивации Na+ системы.
Абсцисса (E-ER): отклонение мембранного потенциала от потенциала покоя.
Потенциалозависимость инактивации Na+ системы.
Абсцисса (E-ER): отклонение мембранного потенциала от потенциала покоя.
Слайд 22A. Ток ионов калия, протекающий через калиевые каналы, чувствителен к изменениям мембранного
A. Ток ионов калия, протекающий через калиевые каналы, чувствителен к изменениям мембранного
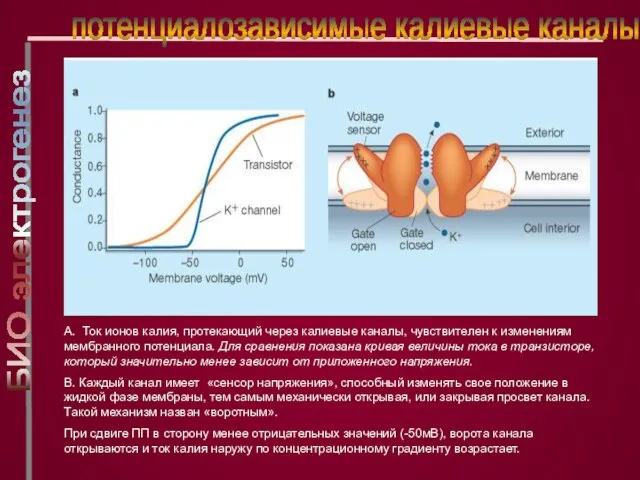
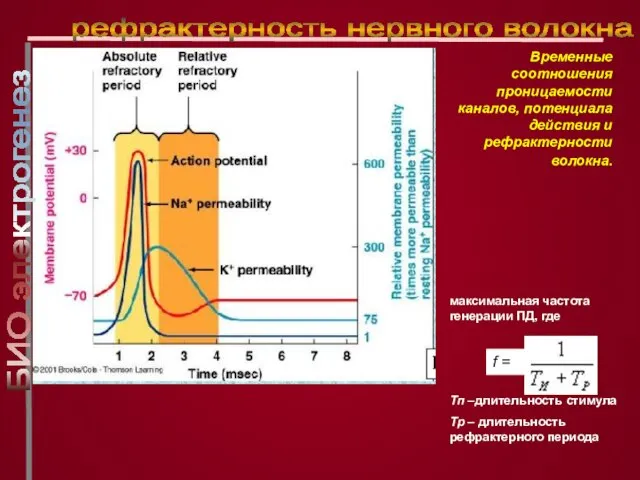
Слайд 23максимальная частота генерации ПД, где
Тп –длительность стимула
Тр – длительность рефрактерного периода
рефрактерность нервного
максимальная частота генерации ПД, где
Тп –длительность стимула
Тр – длительность рефрактерного периода
рефрактерность нервного
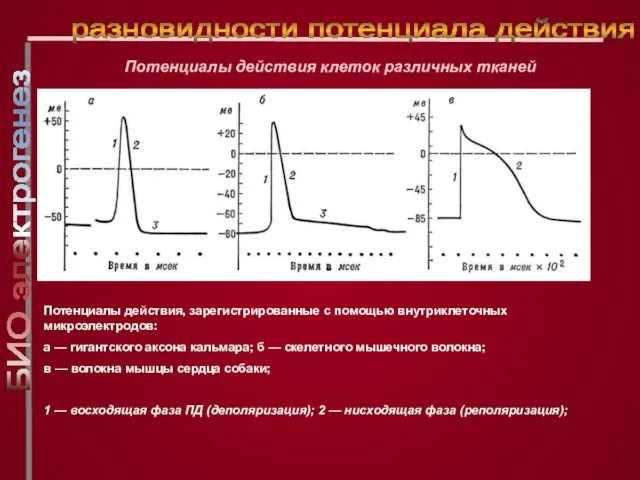
Слайд 24Потенциалы действия, зарегистрированные с помощью внутриклеточных микроэлектродов:
а — гигантского аксона кальмара; б
Потенциалы действия, зарегистрированные с помощью внутриклеточных микроэлектродов:
а — гигантского аксона кальмара; б
Слайд 25Невозбудимые клетки способные к поддержанию потенциала покоя, но не способные к генерированию
Невозбудимые клетки способные к поддержанию потенциала покоя, но не способные к генерированию

 Офиуры или змеехвостки. Тип иглокожие
Офиуры или змеехвостки. Тип иглокожие Угадай животное по фрагменту (ответы)
Угадай животное по фрагменту (ответы) Конечный мозг: полушария, борозды, извилины, базальные ядра. Локализация функций в коре полушарий
Конечный мозг: полушария, борозды, извилины, базальные ядра. Локализация функций в коре полушарий Регуляторы роста и развития растений
Регуляторы роста и развития растений Нерная система. Инстинкт. Органы чувств. Регуляция деятельности организма
Нерная система. Инстинкт. Органы чувств. Регуляция деятельности организма Основи культури in vitro рослин
Основи культури in vitro рослин Метаболическая биохимия
Метаболическая биохимия Класс Амфибии, 7 класс
Класс Амфибии, 7 класс Путешествия с культурными растениями. Викторина
Путешествия с культурными растениями. Викторина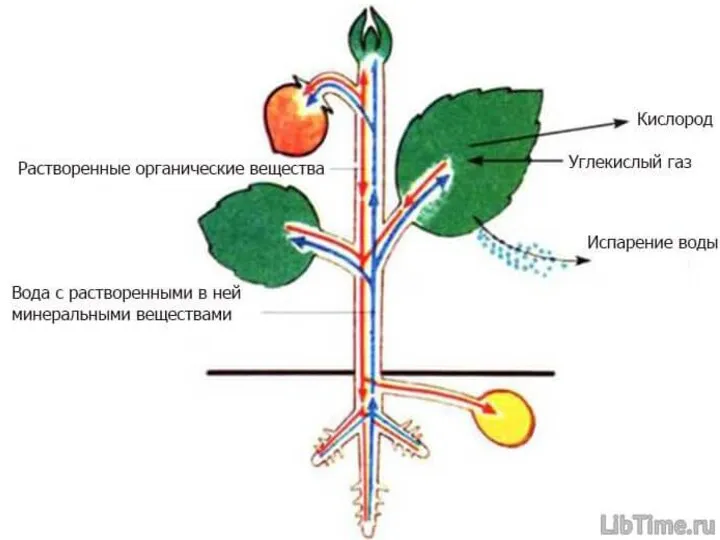 Проводящие ткани
Проводящие ткани Загальна характеристика класу Ракоподібні
Загальна характеристика класу Ракоподібні Центральная нервная система. Brainstem
Центральная нервная система. Brainstem Зрение. Стоение зрительной системы
Зрение. Стоение зрительной системы Лепторумора Микеля
Лепторумора Микеля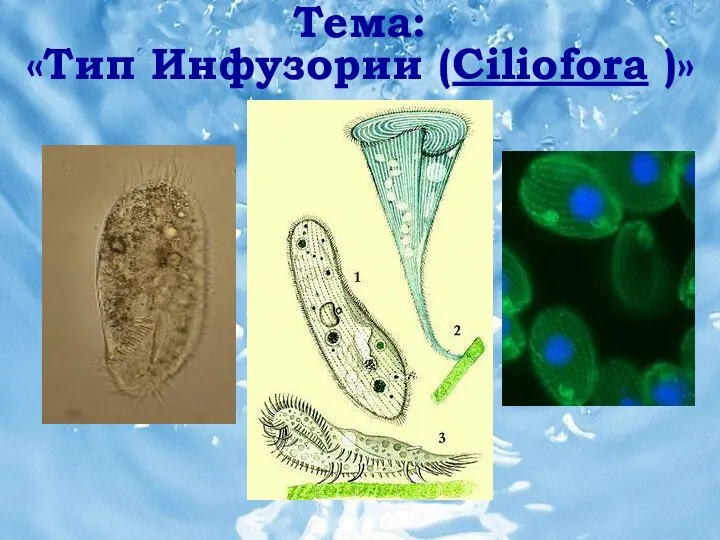 Тип инфузории (ciliofora)
Тип инфузории (ciliofora) Росповсюдження мікроорганізмів у природі
Росповсюдження мікроорганізмів у природі Кролики. Занятие с дошкольниками
Кролики. Занятие с дошкольниками Популяция как единица эволюции
Популяция как единица эволюции Mochepolovaya
Mochepolovaya Деревья России
Деревья России Строение черепа
Строение черепа Выращивание саженцев сосны в гидропонике
Выращивание саженцев сосны в гидропонике Строение, функции и работа мышц
Строение, функции и работа мышц Корень. Виды корней и типы корневых систем. Видоизменения корня
Корень. Виды корней и типы корневых систем. Видоизменения корня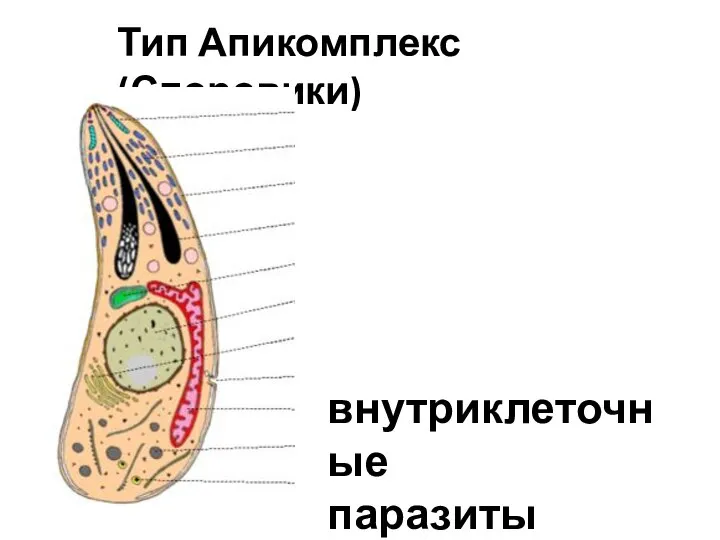 Тип Апикомплекс (Споровики). Внутриклеточные паразиты
Тип Апикомплекс (Споровики). Внутриклеточные паразиты Грибы - паразиты
Грибы - паразиты Железы внешней, внутренней и смешанной секреции. Расположение ЖВС, их строение. Гормоны
Железы внешней, внутренней и смешанной секреции. Расположение ЖВС, их строение. Гормоны Функции и виды тканей
Функции и виды тканей