- Ферменты 1.pptxпрезент

Содержание
- 2. Ферменты (от лат. fermentum - закваска), или энзимы (от греч. en - внутри, zym - закваска)
- 3. Отличие ферментов от небиологических катализаторов 1. Высокая эффективность действия (скорость ферментативных реакций в 106-1012 раз выше,
- 4. Подобно всем белкам ферменты избирательно присоединяют определенные вещества – лиганды . Лиганд, присоединяющийся к ферменту и
- 5. Активный центр фермента: 1. Участок молекулы фермента, сформированный на уровне третичной структуры, ответственный за связывание с
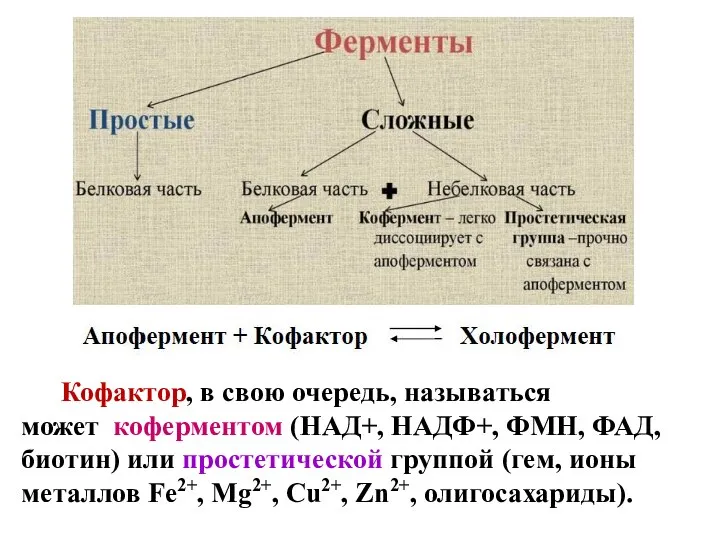
- 6. Кофактор, в свою очередь, называться может коферментом (НАД+, НАДФ+, ФМН, ФАД, биотин) или простетической группой (гем,
- 7. Деление на коферменты и простетические группы не всегда однозначно: - если связь кофактора с белком прочная,
- 8. Апоферменты синтезируются в организме, термолабильны и определяют специфичность ферментов. Кофактор термостабилен, не всегда может образоваться в
- 9. Схема строения ферментов Схема формирования сложного фермента
- 10. Механизм действия ферментов В химическую реакцию могут вступить лишь те молекулы, которые имеют определенный запас энергии.
- 11. При участии фермента реакция идет в обход энергетического барьера. Это обусловлено тем, что в ходе реакции
- 12. Этапы ферментативного катализа I – образование фермент-субстратного комплекса: На этом этапе: а) субстрат приближается к активному
- 13. Типы ферментативных реакций 1. Тип "пинг-понг" – фермент сначала взаимодействует с субстратом А, отбирая у него
- 14. 2. Тип последовательных реакций – к ферменту последовательно присоединяются субстраты А и В, образуя "тройной комплекс",
- 15. 3. Тип случайных взаимодействий – субстраты А и В присоединяются к ферменту в любом порядке, неупорядоченно,
- 16. Абсолютная специфичность Групповая специфичность Стереоспецифичность Активный центр фермента Участок связывания Обеспечивает субстратную специфичность (выбор субстрата) Обеспечивает
- 17. Абсолютная субстратная специфичность – фермент катализирует превращение только одного конкретного субстрата.
- 18. Относительная (групповая) специфичность – фермент катализирует однотипные превращения схожих по строению веществ. Например, липаза гидролизует жиры:
- 19. Стереоспецифичность – фермент катализирует превращение только одного из стереоизомеров для данного вещества. Например, фермент фумараза оказывает
- 20. Каталитическая специфичность (специфичность пути превращения) – фермент катализирует только одно превращение субстрата из всех возможных:
- 21. Теории о специфичности действия ферментов 1. Теория Фишера (1890г.,модель «ключ – замок», модель "жесткой матрицы«) -
- 22. 2.Теория Кошланда (1958г., модель "индуцированного соответствия", "рука-перчатка") – подразумевает гибкость активного центра. Присоединение субстрата к якорному
- 23. Кинетика ферментативных реакций Зависимость скорости ферментативной реакции от температуры среды Скорость химической реакции, катализируемая ферментами, имеет
- 24. Графмк зависимости скорости реакции от температуры
- 25. Зависимость скорости реакции от pH среды Оптимум pH – это такое значение pH, при котором фермент
- 26. При изменении pH среды в сторону от оптимума активность фермента уменьшается, т.к. происходит изменение ионизации функциональных
- 27. Зависимость скорости реакции от концентрации субстрата Если концентрацию ферментов оставить постоянной, изменяя только количество субстрата, то
- 28. Vmax – это такая скорость ферментативной реакции, при которой достигается полное насыщение фермента субстратом, т.е. когда
- 29. Зависимость скорости реакции от концентрации фермента В условиях избытка субстрата скорость ферментативной реакции прямо пропорциональна концентрации
- 30. Для того, чтобы оценить количество молекул фермента среди других белков данной ткани, определяют удельную активность. Удельная
- 31. Классификация ферментов В 1961 г в Москве V Международный биохимический союз принял современную классификацию ферментов. В
- 32. Классификация ферментов
- 33. Шифр фермента Каждому ферменту присвоен четырехзначный классификационный номер(шифр - КФ), включающий класс, подкласс, подподкласс и порядковый
- 34. Название фермента Систематическое – согласно современной классификации, применяемое для однозначной идентификации фермента. Рабочее - добавляется окончание
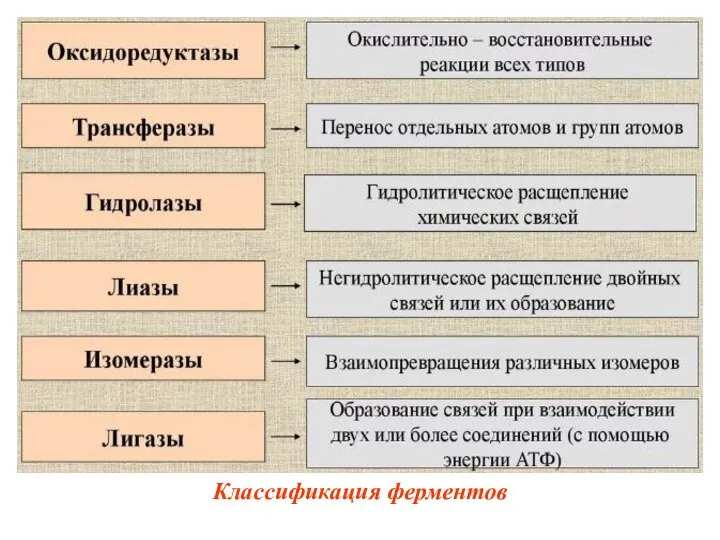
- 35. 1. Оксидоредуктазы Катализирует различные окислительно-восстановительные реакции с участием двух субстратов (перенос электронов или атомов водорода с
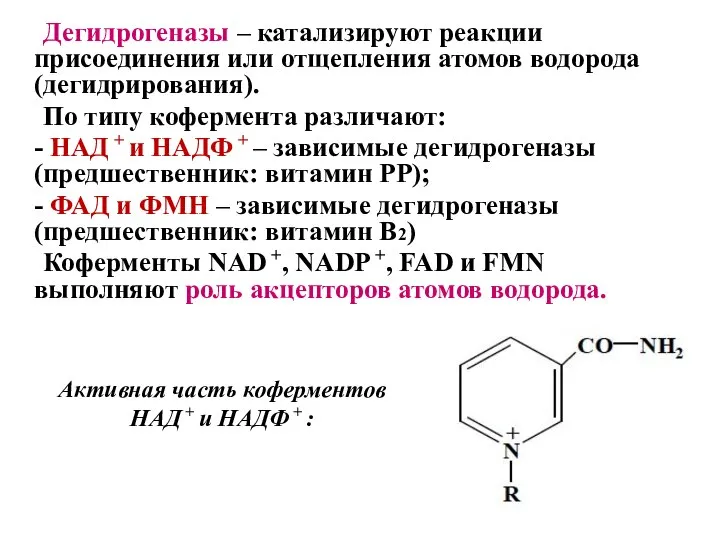
- 36. Дегидрогеназы – катализируют реакции присоединения или отщепления атомов водорода (дегидрирования). По типу кофермента различают: - НАД
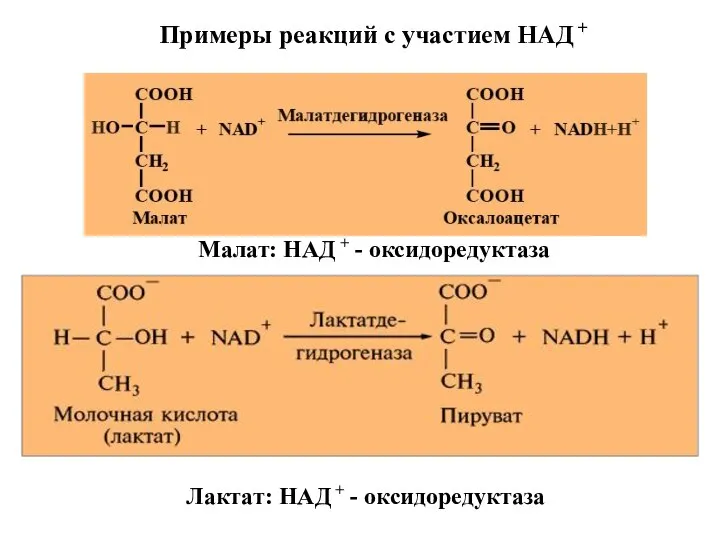
- 37. Примеры реакций с участием НАД + Малат: НАД + - оксидоредуктаза Лактат: НАД + - оксидоредуктаза
- 38. Примеры реакций с участием ФАД:
- 39. Оксидазы. Акцептором электрона служит молекулярный кислород (н., цитохромоксидаза). Оксигеназы - это ферменты, катализирующие активирование О2 и
- 40. 2. Трансферазы. Катализируют перенос функциональных групп от одного соединения к другому. Подразделяют в зависимости от переносимой
- 41. Аминотрансферазы – катализируют перенос аминогруппы с аминокислоты на кетокислоту. Пример: Фермент – Аланин:α-кетоглутарат-аминотрансфераза Кофермент: Пиридоксальфосфат(витамин B6).
- 42. Фосфотрансферазы (киназы) Киназы– катализируют перенос фосфатной группы, донором которой является АТФ (реже ГТФ). Реакции, катализируемые киназами
- 43. Метилтрансферазы – перенос метильной группы.
- 44. Ацилтрансферазы – перенос ацильной группы. Коферментную функцию у ацилтрансфераз выполняет коэнзим А. Фермент: холинацетилтрансфераза
- 45. 3.Гидролазы– катализируют реакции разрыва связей в молекуле с участием воды, т.е. путем гидролиза. Примеры: Фосфатазы– катализируют
- 46. Липазы– гидролиз связей в жирах: Пептидазы – гидролизуют пептидную связь:
- 47. 4. Лиазы К лиазам относят ферменты, отщепляющие от субстратов негидролитическим путем определенную группу (при этом могут
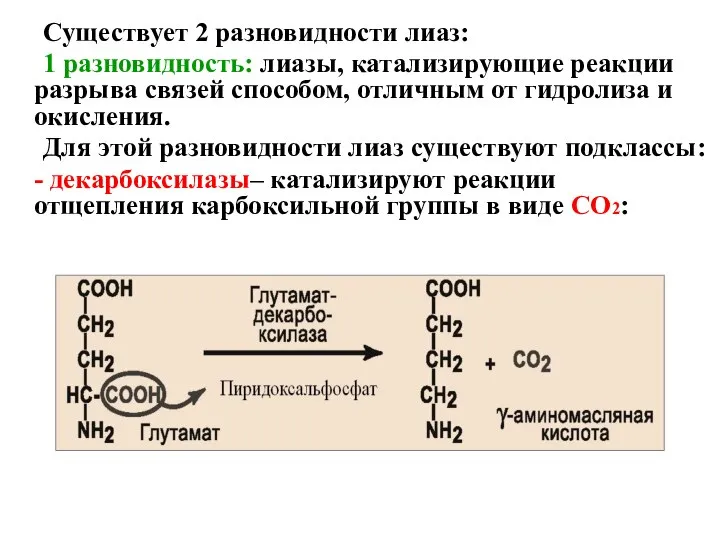
- 48. Существует 2 разновидности лиаз: 1 разновидность: лиазы, катализирующие реакции разрыва связей способом, отличным от гидролиза и
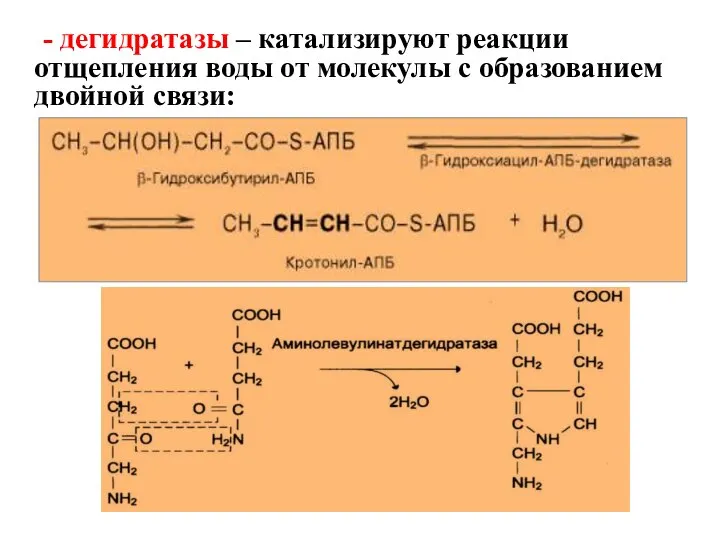
- 49. - дегидратазы – катализируют реакции отщепления воды от молекулы с образованием двойной связи:
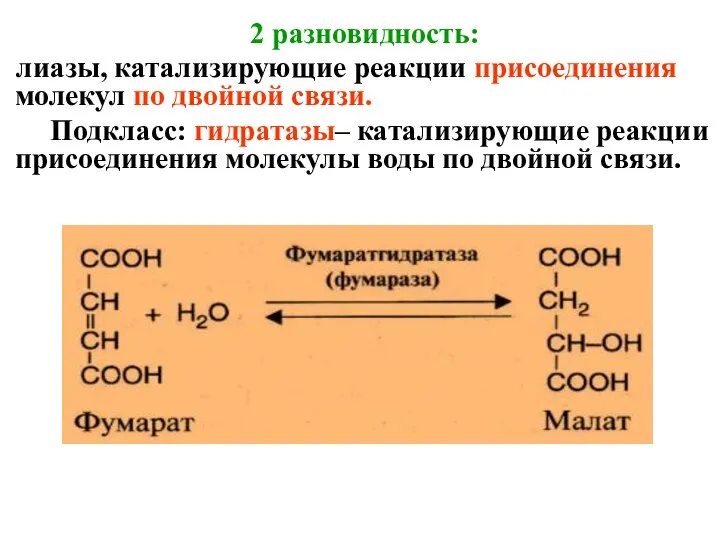
- 50. 2 разновидность: лиазы, катализирующие реакции присоединения молекул по двойной связи. Подкласс: гидратазы– катализирующие реакции присоединения молекулы
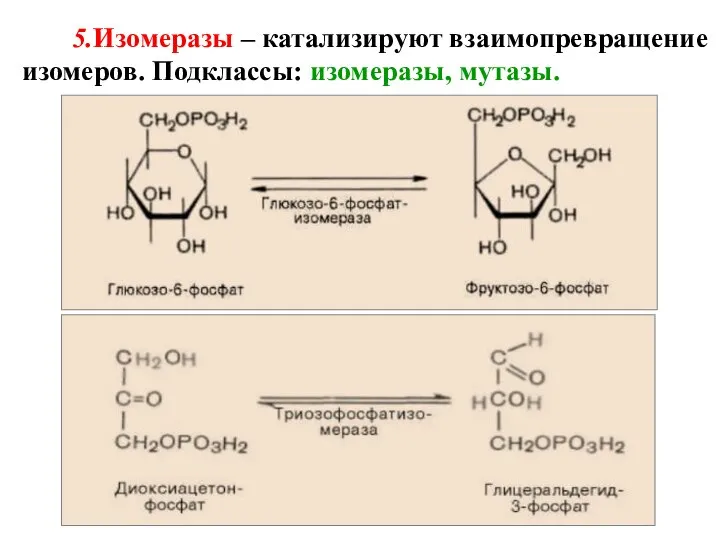
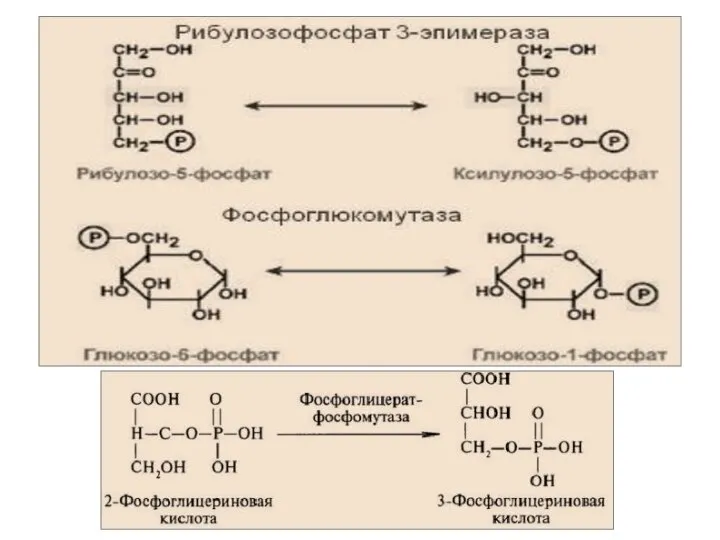
- 51. 5.Изомеразы – катализируют взаимопревращение изомеров. Подклассы: изомеразы, мутазы.
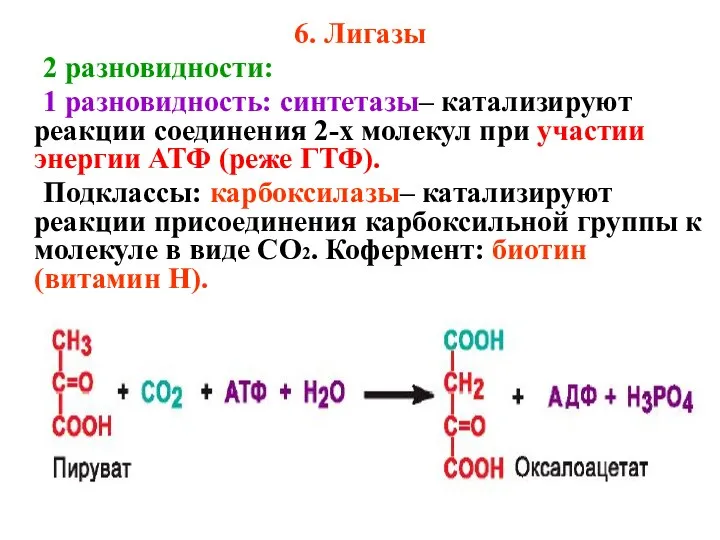
- 53. 6. Лигазы 2 разновидности: 1 разновидность: синтетазы– катализируют реакции соединения 2-х молекул при участии энергии АТФ
- 55. Скачать презентацию

Слайд 3Отличие ферментов от небиологических катализаторов
1. Высокая эффективность действия (скорость ферментативных реакций в
Отличие ферментов от небиологических катализаторов
1. Высокая эффективность действия (скорость ферментативных реакций в

Слайд 4 Подобно всем белкам ферменты избирательно присоединяют определенные вещества – лиганды .
Лиганд,
Подобно всем белкам ферменты избирательно присоединяют определенные вещества – лиганды .
Лиганд,

Слайд 5Активный центр фермента:
1. Участок молекулы фермента, сформированный на уровне третичной структуры, ответственный
Активный центр фермента:
1. Участок молекулы фермента, сформированный на уровне третичной структуры, ответственный

Слайд 6
Кофактор, в свою очередь, называться
может коферментом (НАД+, НАДФ+, ФМН, ФАД, биотин)
Кофактор, в свою очередь, называться
может коферментом (НАД+, НАДФ+, ФМН, ФАД, биотин)
Слайд 7 Деление на коферменты и простетические группы не всегда однозначно:
- если связь кофактора
Деление на коферменты и простетические группы не всегда однозначно: - если связь кофактора

Слайд 8 Апоферменты синтезируются в организме, термолабильны и определяют специфичность ферментов.
Кофактор термостабилен,
Апоферменты синтезируются в организме, термолабильны и определяют специфичность ферментов.
Кофактор термостабилен,

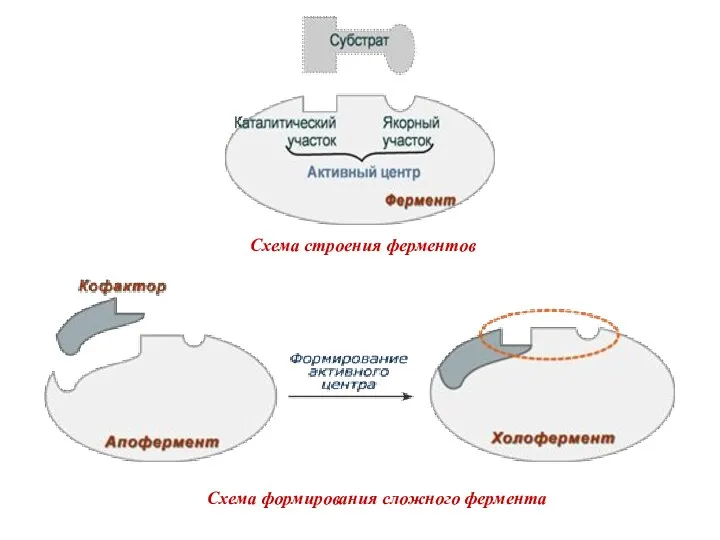
Слайд 9Схема строения ферментов
Схема формирования сложного фермента
Схема строения ферментов
Схема формирования сложного фермента
Слайд 10 Механизм действия ферментов
В химическую реакцию могут вступить лишь те молекулы, которые имеют
Механизм действия ферментов
В химическую реакцию могут вступить лишь те молекулы, которые имеют

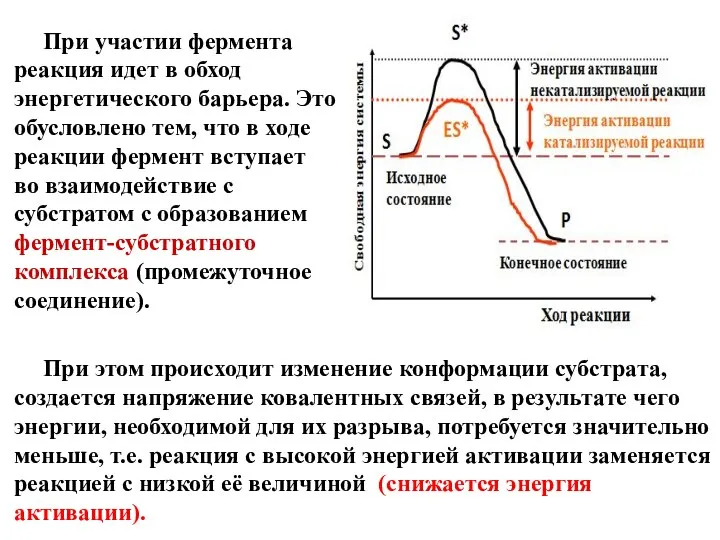
Слайд 11 При участии фермента реакция идет в обход энергетического барьера. Это обусловлено тем,
При участии фермента реакция идет в обход энергетического барьера. Это обусловлено тем,
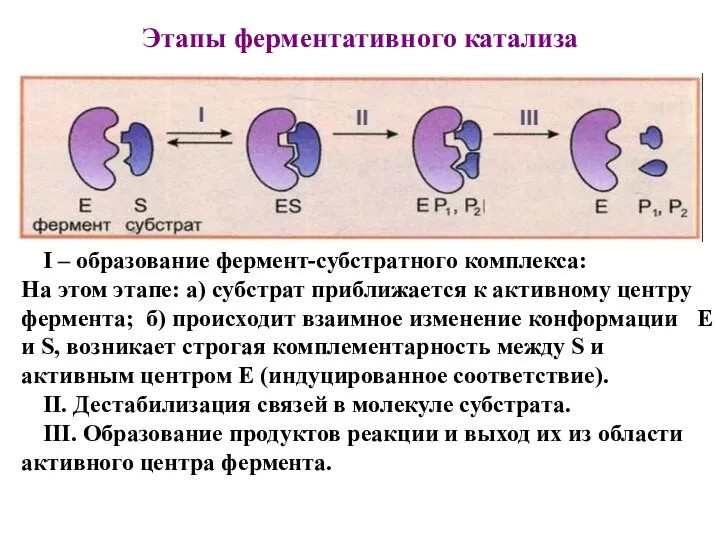
Слайд 12Этапы ферментативного катализа
I – образование фермент-субстратного комплекса:
На этом этапе: а) субстрат
Этапы ферментативного катализа
I – образование фермент-субстратного комплекса:
На этом этапе: а) субстрат
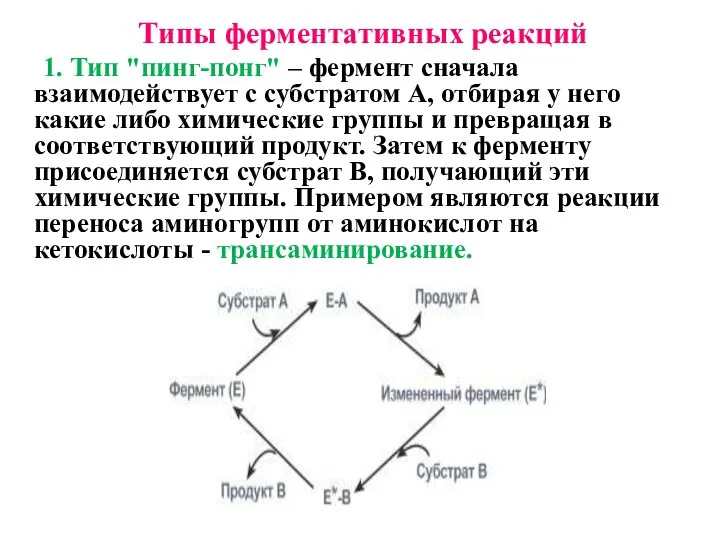
Слайд 13Типы ферментативных реакций
1. Тип "пинг-понг" – фермент сначала взаимодействует с субстратом А, отбирая у
Типы ферментативных реакций
1. Тип "пинг-понг" – фермент сначала взаимодействует с субстратом А, отбирая у
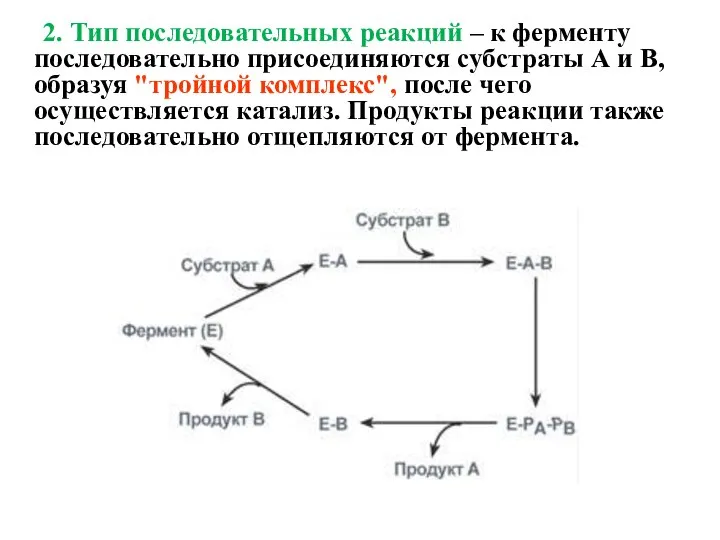
Слайд 14 2. Тип последовательных реакций – к ферменту последовательно присоединяются субстраты А и В, образуя
2. Тип последовательных реакций – к ферменту последовательно присоединяются субстраты А и В, образуя
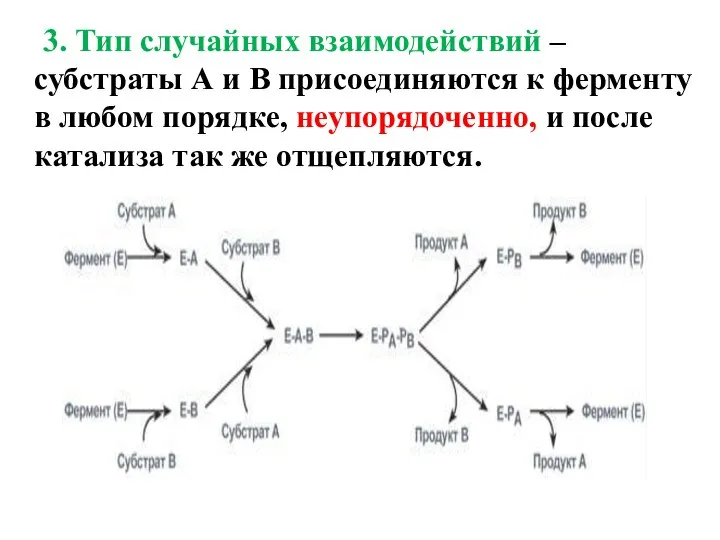
Слайд 15 3. Тип случайных взаимодействий – субстраты А и В присоединяются к ферменту в любом
3. Тип случайных взаимодействий – субстраты А и В присоединяются к ферменту в любом
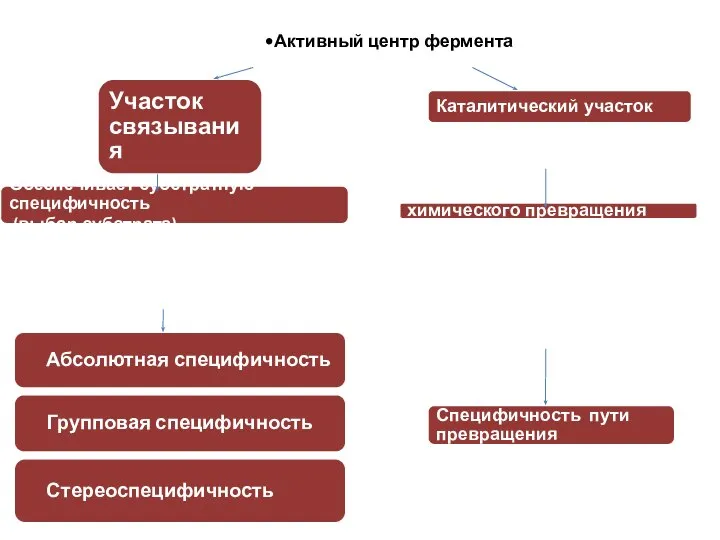
Слайд 16 Абсолютная специфичность
Групповая специфичность
Стереоспецифичность
Активный центр фермента
Участок связывания
Обеспечивает субстратную специфичность
(выбор
Абсолютная специфичность
Групповая специфичность
Стереоспецифичность
Активный центр фермента
Участок связывания
Обеспечивает субстратную специфичность
(выбор
Слайд 17 Абсолютная субстратная специфичность – фермент катализирует превращение только одного конкретного субстрата.
Абсолютная субстратная специфичность – фермент катализирует превращение только одного конкретного субстрата.
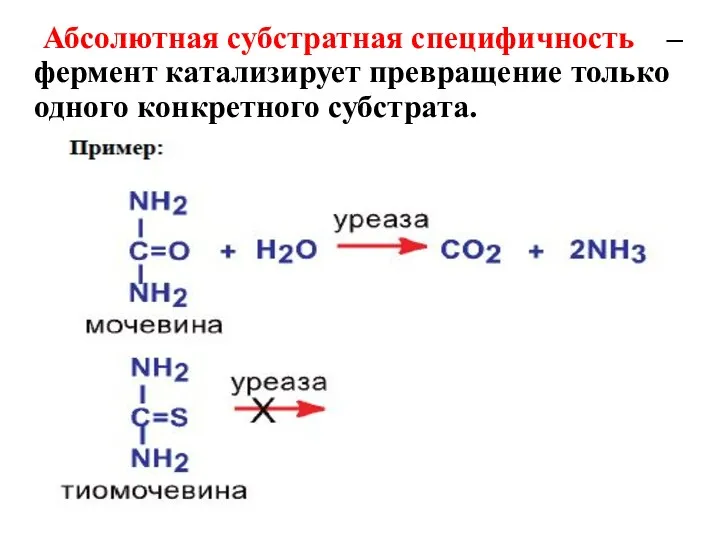
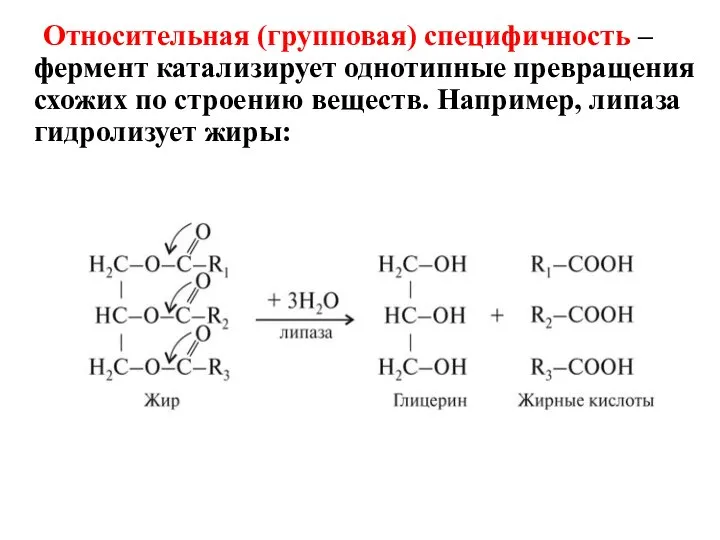
Слайд 18 Относительная (групповая) специфичность – фермент катализирует однотипные превращения схожих по строению веществ.
Относительная (групповая) специфичность – фермент катализирует однотипные превращения схожих по строению веществ.
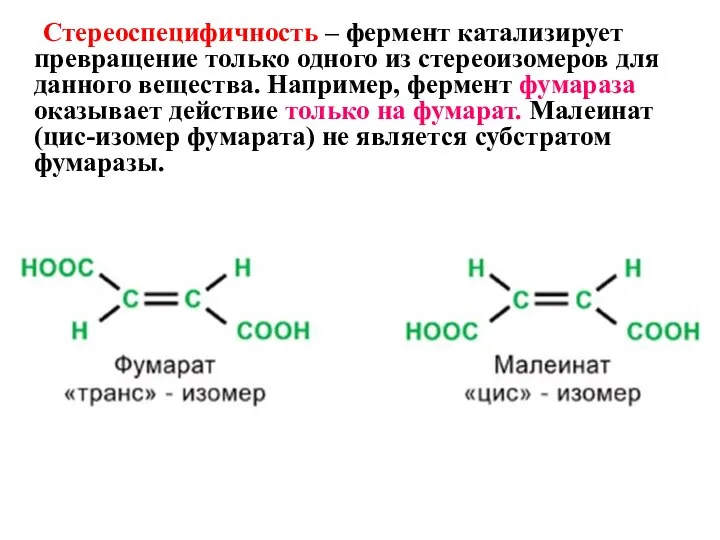
Слайд 19 Стереоспецифичность – фермент катализирует превращение только одного из стереоизомеров для данного вещества.
Стереоспецифичность – фермент катализирует превращение только одного из стереоизомеров для данного вещества.
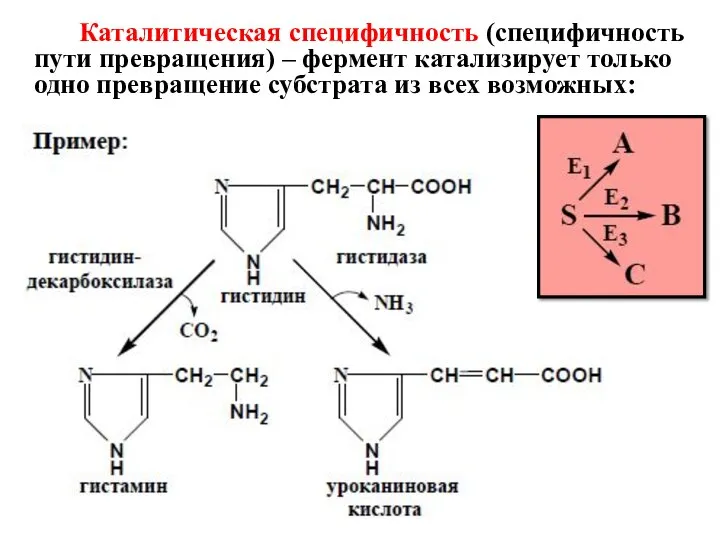
Слайд 20 Каталитическая специфичность (специфичность пути превращения) – фермент катализирует только одно превращение субстрата
Каталитическая специфичность (специфичность пути превращения) – фермент катализирует только одно превращение субстрата
Слайд 21 Теории о специфичности действия ферментов
1. Теория Фишера (1890г.,модель «ключ – замок», модель
Теории о специфичности действия ферментов
1. Теория Фишера (1890г.,модель «ключ – замок», модель
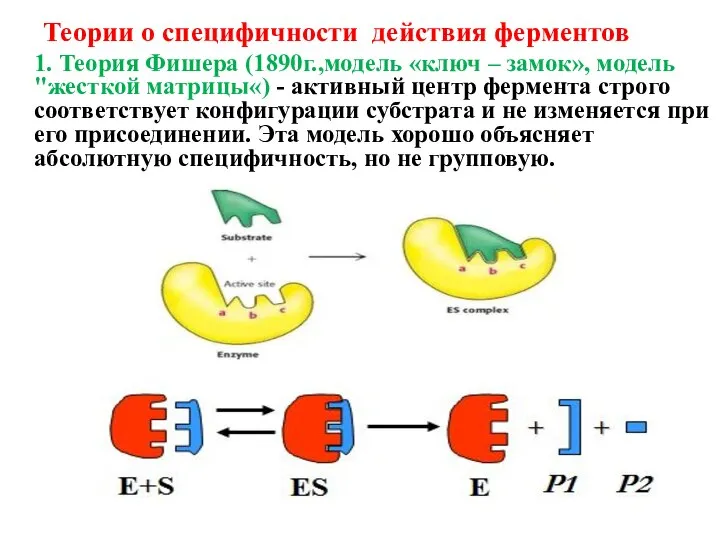
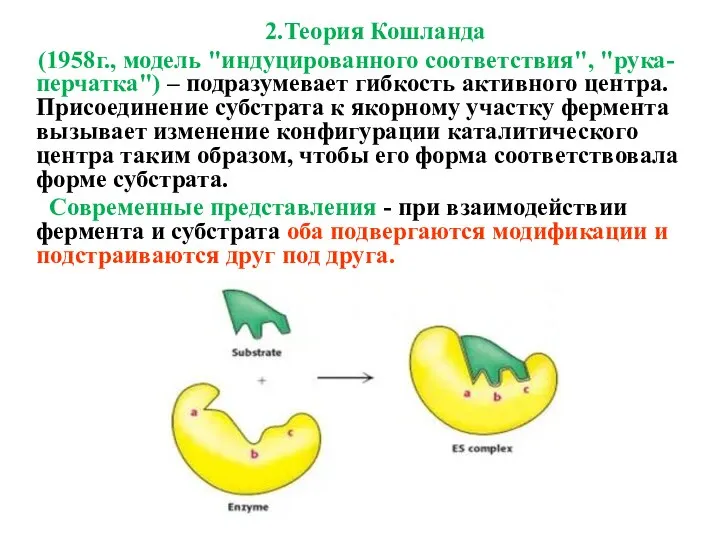
Слайд 222.Теория Кошланда
(1958г., модель "индуцированного соответствия", "рука-перчатка") – подразумевает гибкость активного центра. Присоединение
2.Теория Кошланда
(1958г., модель "индуцированного соответствия", "рука-перчатка") – подразумевает гибкость активного центра. Присоединение
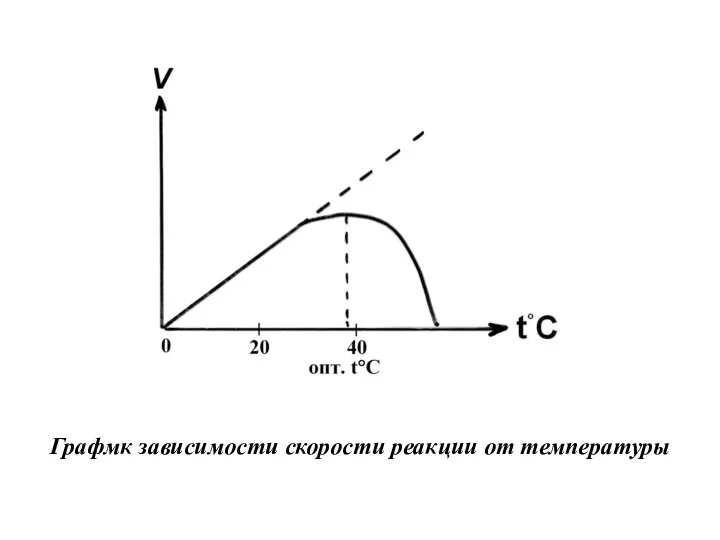
Слайд 23 Кинетика ферментативных реакций
Зависимость скорости ферментативной реакции от температуры среды
Скорость химической
Кинетика ферментативных реакций
Зависимость скорости ферментативной реакции от температуры среды
Скорость химической

Слайд 24
Графмк зависимости скорости реакции от температуры
Графмк зависимости скорости реакции от температуры
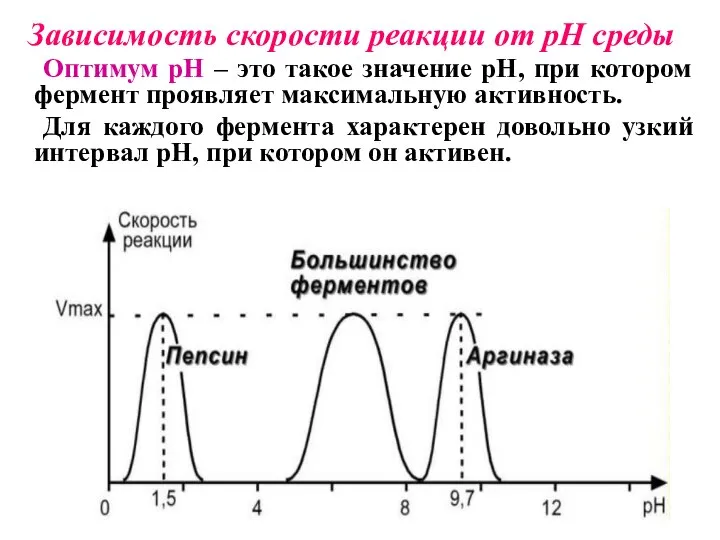
Слайд 25Зависимость скорости реакции от pH среды
Оптимум pH – это такое значение pH,
Зависимость скорости реакции от pH среды
Оптимум pH – это такое значение pH,
Слайд 26 При изменении pH среды в сторону от оптимума активность фермента уменьшается,
При изменении pH среды в сторону от оптимума активность фермента уменьшается,

Слайд 27 Зависимость скорости реакции от концентрации субстрата
Если концентрацию ферментов оставить постоянной, изменяя
Зависимость скорости реакции от концентрации субстрата
Если концентрацию ферментов оставить постоянной, изменяя
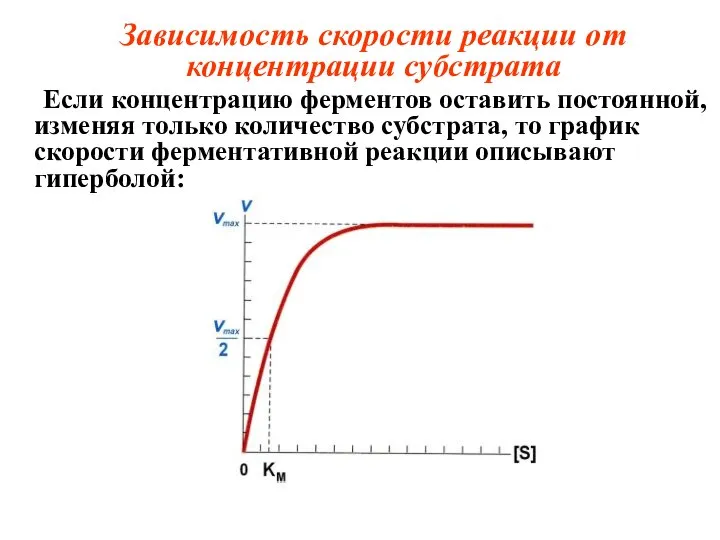
Слайд 28 Vmax – это такая скорость ферментативной реакции, при которой достигается полное насыщение
Vmax – это такая скорость ферментативной реакции, при которой достигается полное насыщение

Слайд 29 Зависимость скорости реакции от концентрации фермента
В условиях избытка субстрата скорость ферментативной
Зависимость скорости реакции от концентрации фермента
В условиях избытка субстрата скорость ферментативной

Слайд 30 Для того, чтобы оценить количество молекул фермента среди других белков данной ткани,
Для того, чтобы оценить количество молекул фермента среди других белков данной ткани,

Слайд 31 Классификация ферментов
В 1961 г в Москве V Международный биохимический союз принял современную
Классификация ферментов
В 1961 г в Москве V Международный биохимический союз принял современную

Слайд 32Классификация ферментов
Классификация ферментов
Слайд 33 Шифр фермента
Каждому ферменту присвоен четырехзначный классификационный номер(шифр - КФ), включающий класс, подкласс,
Шифр фермента
Каждому ферменту присвоен четырехзначный классификационный номер(шифр - КФ), включающий класс, подкласс,

Слайд 34 Название фермента
Систематическое – согласно современной классификации, применяемое для однозначной идентификации фермента.
Рабочее
Название фермента
Систематическое – согласно современной классификации, применяемое для однозначной идентификации фермента.
Рабочее

Слайд 35 1. Оксидоредуктазы
Катализирует различные окислительно-восстановительные реакции с участием двух субстратов (перенос электронов или
1. Оксидоредуктазы
Катализирует различные окислительно-восстановительные реакции с участием двух субстратов (перенос электронов или

Слайд 36 Дегидрогеназы – катализируют реакции присоединения или отщепления атомов водорода (дегидрирования).
По типу кофермента
Дегидрогеназы – катализируют реакции присоединения или отщепления атомов водорода (дегидрирования).
По типу кофермента
Слайд 37 Примеры реакций с участием НАД +
Малат: НАД + - оксидоредуктаза
Лактат: НАД +
Примеры реакций с участием НАД +
Малат: НАД + - оксидоредуктаза
Лактат: НАД +
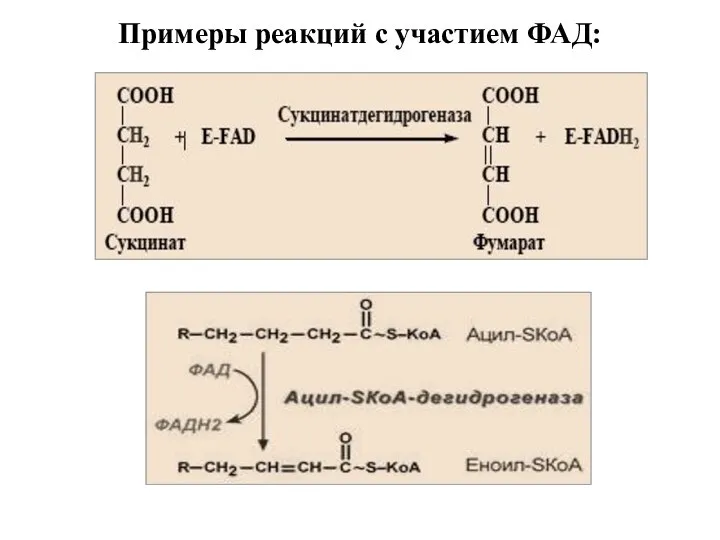
Слайд 38Примеры реакций с участием ФАД:
Примеры реакций с участием ФАД:
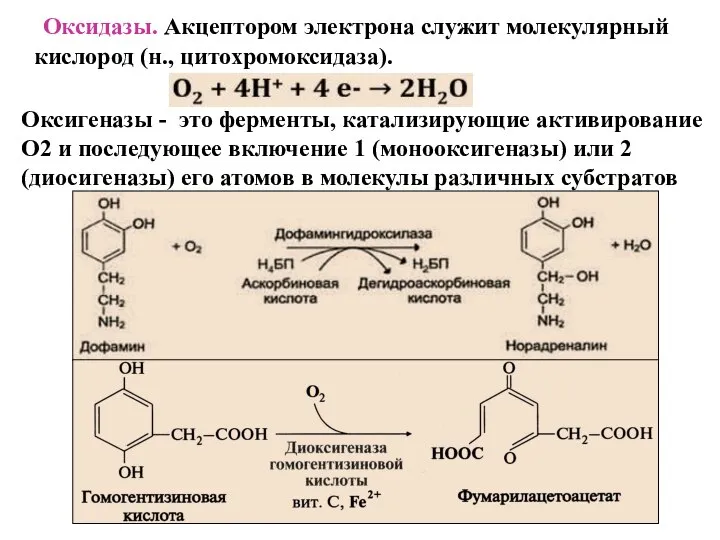
Слайд 39 Оксидазы. Акцептором электрона служит молекулярный кислород (н., цитохромоксидаза).
Оксигеназы - это ферменты, катализирующие
Оксидазы. Акцептором электрона служит молекулярный кислород (н., цитохромоксидаза).
Оксигеназы - это ферменты, катализирующие
Слайд 40 2. Трансферазы. Катализируют перенос функциональных групп от одного соединения к другому.
Подразделяют
2. Трансферазы. Катализируют перенос функциональных групп от одного соединения к другому.
Подразделяют

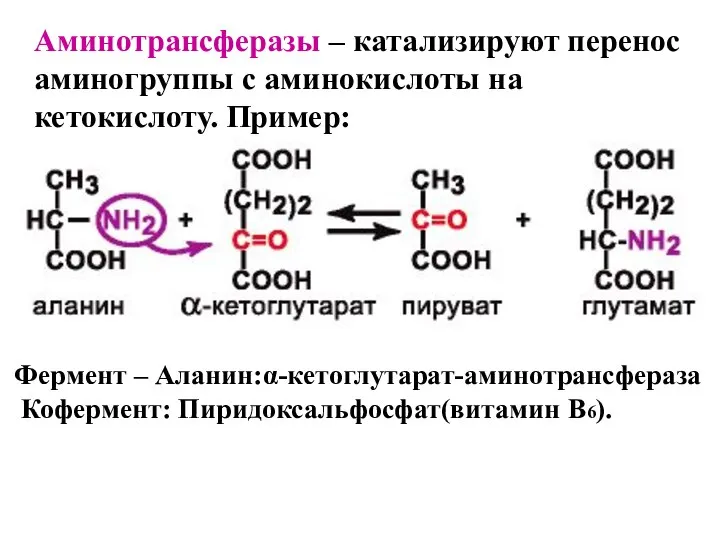
Слайд 41 Аминотрансферазы – катализируют перенос аминогруппы с аминокислоты на кетокислоту. Пример:
Фермент – Аланин:α-кетоглутарат-аминотрансфераза
Аминотрансферазы – катализируют перенос аминогруппы с аминокислоты на кетокислоту. Пример:
Фермент – Аланин:α-кетоглутарат-аминотрансфераза
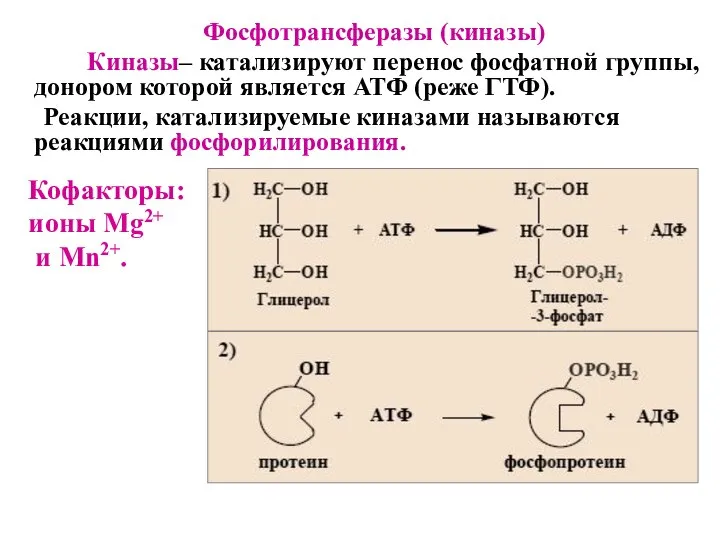
Слайд 42Фосфотрансферазы (киназы)
Киназы– катализируют перенос фосфатной группы, донором которой является АТФ
Фосфотрансферазы (киназы)
Киназы– катализируют перенос фосфатной группы, донором которой является АТФ
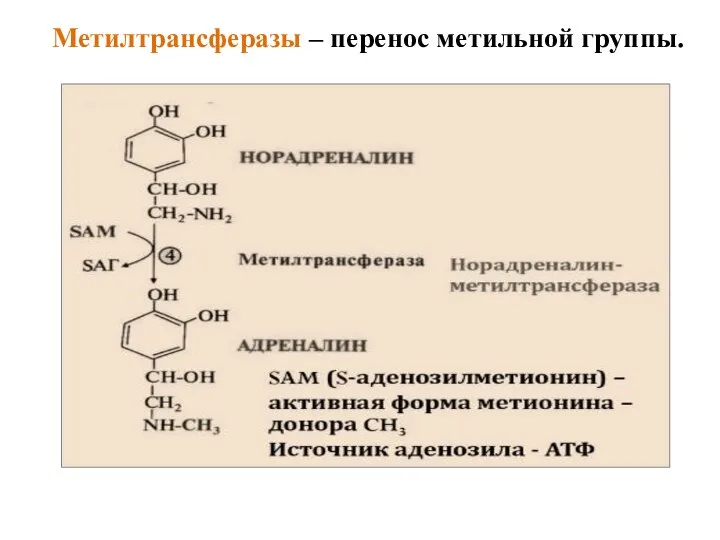
Слайд 43Метилтрансферазы – перенос метильной группы.
Метилтрансферазы – перенос метильной группы.
Слайд 44Ацилтрансферазы – перенос ацильной группы. Коферментную функцию у ацилтрансфераз выполняет коэнзим А.
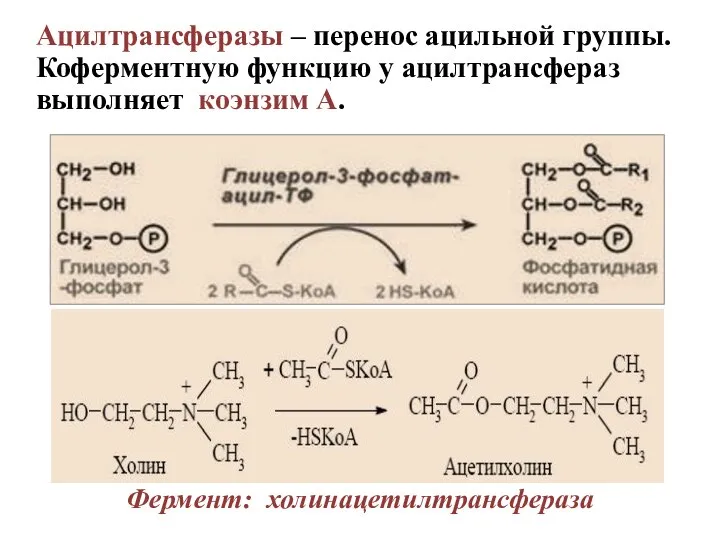
Ацилтрансферазы – перенос ацильной группы. Коферментную функцию у ацилтрансфераз выполняет коэнзим А.
Слайд 45 3.Гидролазы– катализируют реакции разрыва связей в молекуле с участием воды, т.е. путем
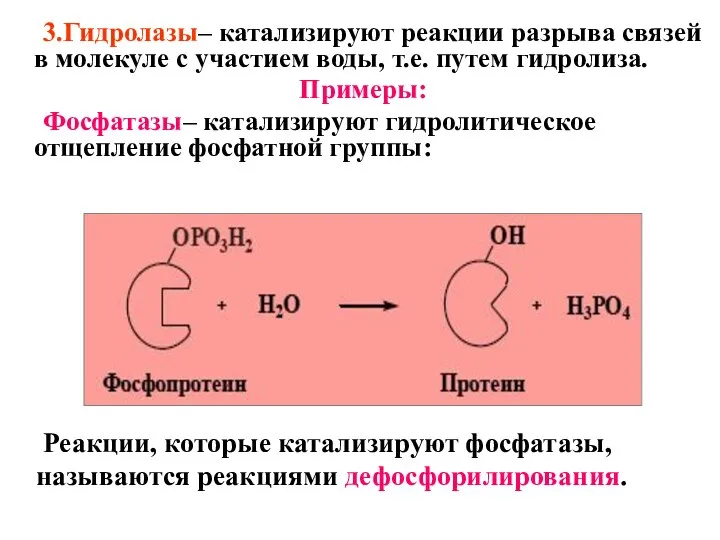
3.Гидролазы– катализируют реакции разрыва связей в молекуле с участием воды, т.е. путем
Слайд 46Липазы– гидролиз связей в жирах:
Пептидазы – гидролизуют пептидную связь:
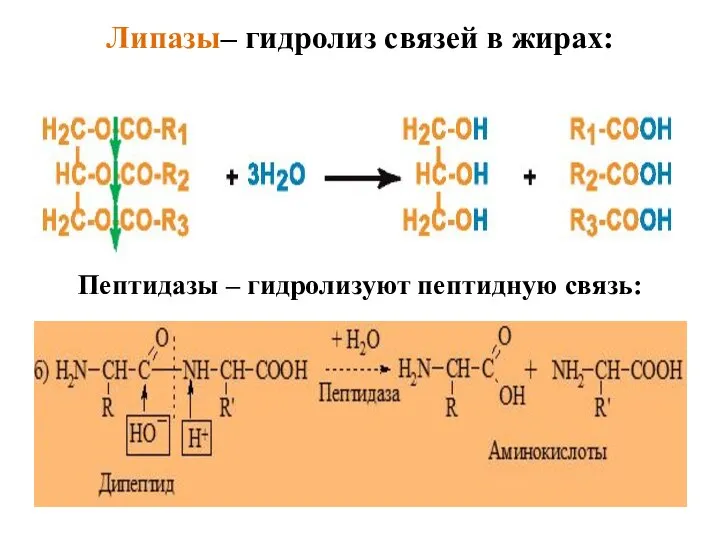
Липазы– гидролиз связей в жирах:
Пептидазы – гидролизуют пептидную связь:
Слайд 474. Лиазы
К лиазам относят ферменты, отщепляющие от субстратов негидролитическим путем определенную группу
4. Лиазы
К лиазам относят ферменты, отщепляющие от субстратов негидролитическим путем определенную группу

Слайд 48 Существует 2 разновидности лиаз:
1 разновидность: лиазы, катализирующие реакции разрыва связей способом,
Существует 2 разновидности лиаз:
1 разновидность: лиазы, катализирующие реакции разрыва связей способом,
Слайд 49 - дегидратазы – катализируют реакции отщепления воды от молекулы с образованием двойной
- дегидратазы – катализируют реакции отщепления воды от молекулы с образованием двойной
Слайд 502 разновидность:
лиазы, катализирующие реакции присоединения молекул по двойной связи.
Подкласс:
2 разновидность:
лиазы, катализирующие реакции присоединения молекул по двойной связи.
Подкласс:
Слайд 51 5.Изомеразы – катализируют взаимопревращение изомеров. Подклассы: изомеразы, мутазы.
5.Изомеразы – катализируют взаимопревращение изомеров. Подклассы: изомеразы, мутазы.
Слайд 536. Лигазы
2 разновидности:
1 разновидность: синтетазы– катализируют реакции соединения 2-х молекул
6. Лигазы
2 разновидности:
1 разновидность: синтетазы– катализируют реакции соединения 2-х молекул
 Комнатные растения
Комнатные растения Дендрофлора Приморского края
Дендрофлора Приморского края Особенности организации плоских червей. Важнейшие ароморфозы
Особенности организации плоских червей. Важнейшие ароморфозы Митоз. Репликация (удвоение) ДНК - основа митоза
Митоз. Репликация (удвоение) ДНК - основа митоза Исследование воздействия запахов
Исследование воздействия запахов Компостирование пищевых отходов в домашних условиях
Компостирование пищевых отходов в домашних условиях Видовой состав растений горы Магнитной
Видовой состав растений горы Магнитной Строение листа. Жилкование листа. Листорасположение
Строение листа. Жилкование листа. Листорасположение Звери и птицы весной
Звери и птицы весной Проведение вирусологических методов исследования
Проведение вирусологических методов исследования Органы чувств и их значение. Строение органа зрения
Органы чувств и их значение. Строение органа зрения Катаболизм. Общая схема цикла Кребса
Катаболизм. Общая схема цикла Кребса Классификация моллюсков
Классификация моллюсков Органы цветковых растений. Типы корневых систем
Органы цветковых растений. Типы корневых систем Хамелеон
Хамелеон Класс ракообразные
Класс ракообразные Ваши права и обязанности
Ваши права и обязанности Процесс желчеобразования. Свойства желчи
Процесс желчеобразования. Свойства желчи ATLAS by Kurahma. Медицинский атлас
ATLAS by Kurahma. Медицинский атлас Морфофункциональная характеристика скелета туловища, головы и аппарата их движения
Морфофункциональная характеристика скелета туловища, головы и аппарата их движения lektsia_4_Molekulyarnaya_genetika (1)
lektsia_4_Molekulyarnaya_genetika (1) Функции жиров
Функции жиров Фізіологія ендокринної системи
Фізіологія ендокринної системи Класс Пресмыкающиеся, или Рептилии
Класс Пресмыкающиеся, или Рептилии Галофобы
Галофобы Презентация на тему Винегрет: устный журнал о правильном питании школьников
Презентация на тему Винегрет: устный журнал о правильном питании школьников  Поєднання знімків Sentinel-2 і даних повидільної таксації для дешифрування деревного запасу лісових насаджень
Поєднання знімків Sentinel-2 і даних повидільної таксації для дешифрування деревного запасу лісових насаджень Чёрный аист
Чёрный аист