- Биоенергетика

Содержание
- 2. Общи закони на термодинамиката I. Първи общ закон за запазване на енергията: При всяка физична или
- 3. За затворени системи: ΔG = ΔH - T ΔS или в условията на биохимичните реакции: ΔG
- 4. Изчисляване на ΔG промяна в свободната енергия на Гибс, т.е тази част от общата енергетична промяна
- 5. Изчисляване на ΔG ΔG ΔG > 0, реакцията е ендергонична – протича само, ако в системата
- 6. ИЗПОЛЗВАЕМА ЕНЕРГИЯ Източник на енергия за човека и животните - само химическата енергия, отделяна при разграждане
- 7. Стационарно състояние и термодинамично равновесие Химичните реакции в отворените системи: рядко до равновесно състояние еднопосочно продуктите
- 8. Спрягане на екзергонични и ендергонични реакции Доставящи енергия - окислителни, катаболитни и екзергонични пр. гликолиза, β-окисление
- 9. Спрягане на процесите, доставящи и консумиращи енергия чрез макроергични съединения
- 10. Едновременно или последователно протичане с общ метаболит – енергия за протичането на ендергоничната реакция: Ендергонична реакция
- 11. Макроергични съединения Макроергични връзки: при хидролитно разграждане ΔG°' ≥ 30 кJ/mol (до 70 кJ/mol) обикновени ковалентни
- 12. Пирофосфатни макроергични връзки: Нуклеозидтрифосфати - три фосфатни връзки: α, β и γ - α - обикновена
- 13. Примери за други макроергични съединения:
- 14. Различни енергетични нива на фосфорната киселина в организма нулево енергетично ниво - свободната фосфорна киселина нормално
- 15. ЦЕНТРАЛНА РОЛЯ НА АДЕНИЛОВАТА СИСТЕМА АТФ не е най-богатото на енергия съединение междинна стойност за ΔG°'
- 16. Биологично окисление Термини: Окисление – процес, при който от веществата се отделят електрони Редукция – процес,
- 17. СТАДИИ В КАТАБОЛИЗМА аеробен метаболизъм - кислород като електронен акцептор за генериране на енергия анаеробен метаболизъм
- 18. ОСОБЕНОСТИ НА БИОЛОГИЧНОТО ОКИСЛЕНИЕ 1. Ензими - от група I (оксидоредуктази) Отделените от субстрата водород или
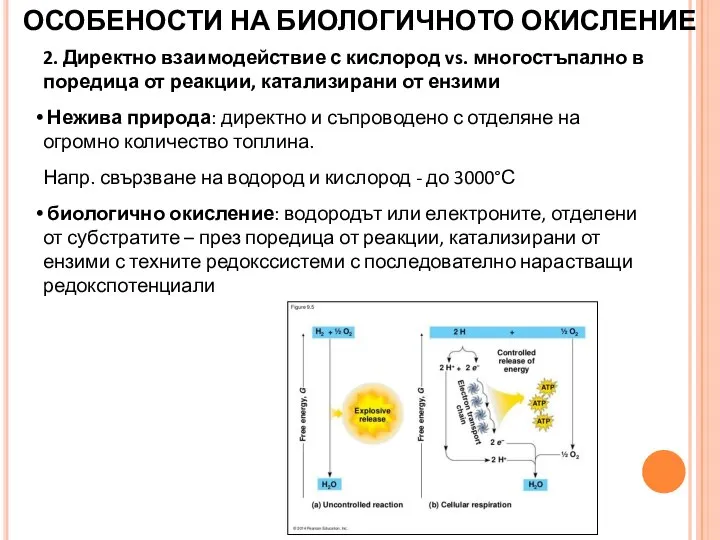
- 19. 2. Директно взаимодействие с кислород vs. многостъпално в поредица от реакции, катализирани от ензими Нежива природа:
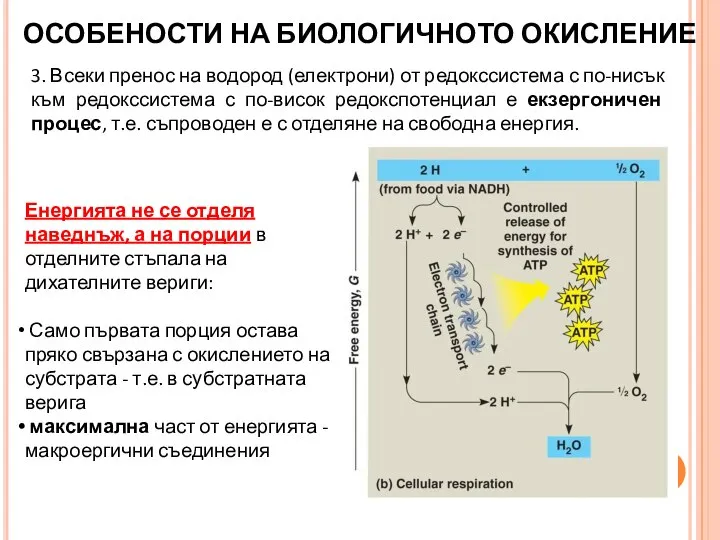
- 20. 3. Всеки пренос на водород (електрони) от редокссистема с по-нисък към редокссистема с по-висок редокспотенциал е
- 21. ОКСИДОРЕДУКТАЗИ (ЕНЗИМИ, ОСЪЩЕСТВЯВАЩИ БИОЛОГИЧНО ОКИСЛЕНИЕ) Дехидрогенази анаеробни, т. е. не могат да използват кислород като акцептор
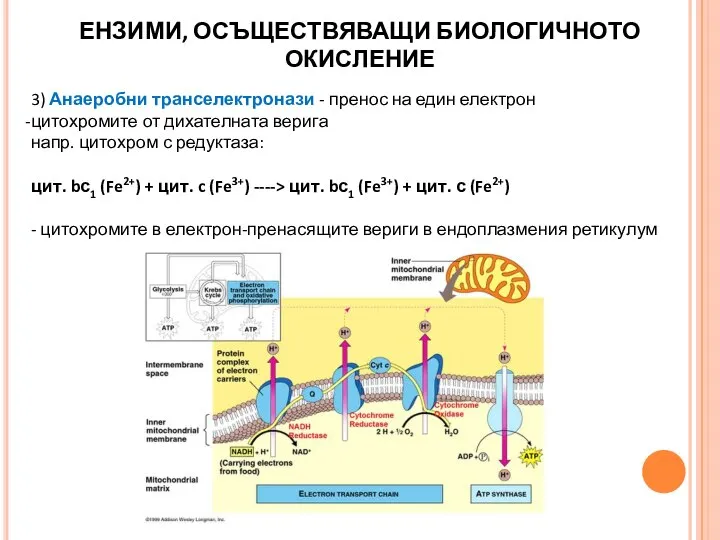
- 22. 3) Анаеробни транселектронази - пренос на един електрон цитохромите от дихателната верига напр. цитохром с редуктаза:
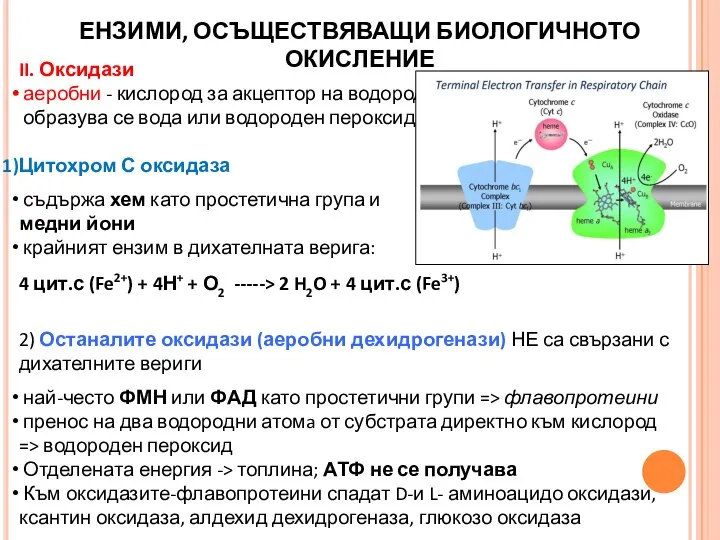
- 23. II. Оксидази аеробни - кислород за акцептор на водород, образува се вода или водороден пероксид Цитохром
- 24. III. Оксигенази Монооксигенази (хидроксилази): катализират вмъкването на един атом кислород в субстрата, при което се получава
- 25. 2) Диоксигенази вмъкват два атома кислород в ароматни пръстени, последвано от окислително разтваряне на пръстена. Участват
- 26. IV. Хидроксипероксидази разграждане на вредните за организма пероксиди и получаващите се от тях свободни радикали 1)
- 27. РЕДОКС-СИСТЕМИ С БИОЛОГИЧНО ЗНАЧЕНИЕ Никотинамидни редокссистеми никотинамидаденин динуклеотид (НАД+/НАДН + Н+) никотинамидаденин динуклеотидфосфат (НАДФ+/НАДФН + Н+)
- 28. Тези редокссистеми пренасят един Н атом и един електрон, т. е. хидриден йон Н- Окислени форми:
- 29. Никотинамидните редокссистеми: близка структура, много близък, при това нисък нормален редокспотенциал (по-нисък от на флавиновите) =>
- 30. 2.Флавинови редокссистеми: флавинмононуклеотид (ФМН) флавинадениндинуклеотид (ФАД) производни на витамин В2 (рибофлавин) пренасят два Н атома (електрони),
- 31. 3. Редокс-системи с хинонова структура КоQ – убихинон РЕДОКС-СИСТЕМИ С БИОЛОГИЧНО ЗНАЧЕНИЕ KоQ.H2 и KoQ -
- 32. 4. Метал-съдържащи редокссистеми Железни или медни йони, обикновено здраво свързани с белтъчен компонент, могат да пренасят
- 33. 5. Тиолови редокс-системи 1) липоева киселина 2) глутатион Липоева киселина Тиооктанова киселина в редуцирано състояние -
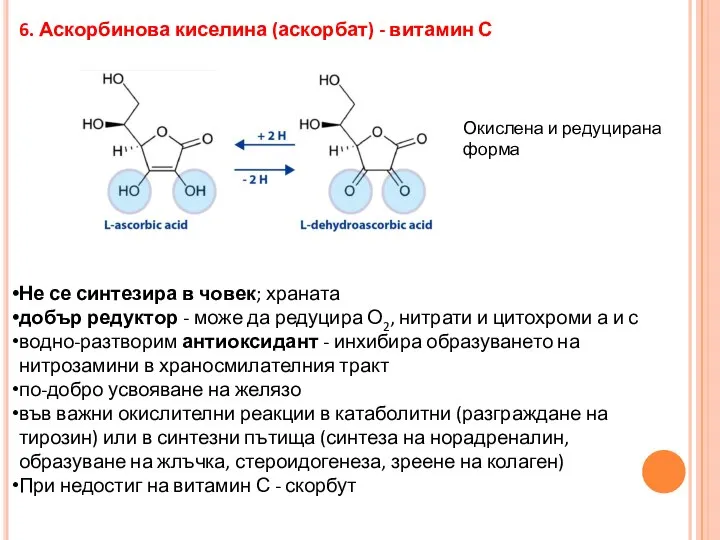
- 34. 6. Аскорбинова киселина (аскорбат) - витамин С Не се синтезира в човек; храната добър редуктор -
- 35. СУБСТРАТНО ОКИСЛЕНИЕ едностъпално анаеробно дехидрогениране на стотици различни субстрати под действие на специфични дехидрогенази, кооперирани най-често
- 36. ОКИСЛИТЕЛНО ФОСФОРИЛИРАНЕ НА СУБСТРАТНО НИВО Синтезата на АТФ или други макроергични съединения за сметка на енергия,
- 37. Скромен количествен принос, но има значение, тъй като: В условията на кислородна недостатъчност - единствен източник
- 39. Скачать презентацию

Слайд 3За затворени системи:
ΔG = ΔH - T ΔS
или в условията на
За затворени системи:
ΔG = ΔH - T ΔS
или в условията на

Слайд 4Изчисляване на ΔG
промяна в свободната енергия на Гибс, т.е тази част от
Изчисляване на ΔG
промяна в свободната енергия на Гибс, т.е тази част от

Слайд 5Изчисляване на ΔG
ΔG < 0, реакцията е екзергонична – протича спонтанно с
Изчисляване на ΔG
ΔG < 0, реакцията е екзергонична – протича спонтанно с
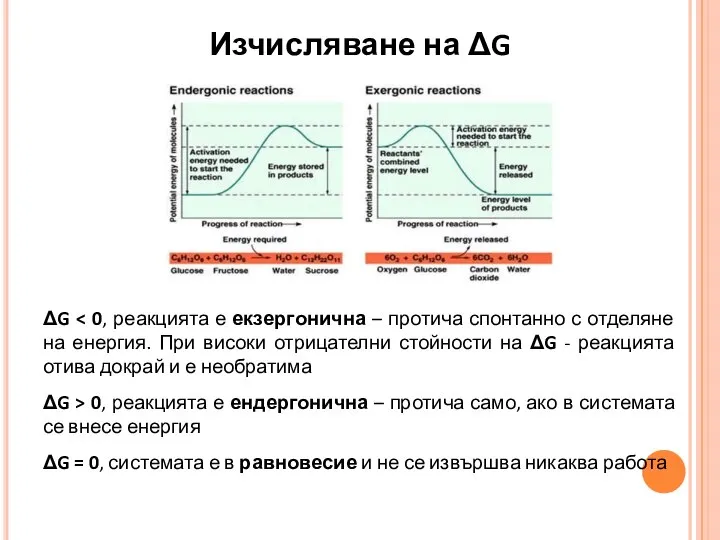
Слайд 6ИЗПОЛЗВАЕМА ЕНЕРГИЯ
Източник на енергия за човека и животните - само химическата енергия,
ИЗПОЛЗВАЕМА ЕНЕРГИЯ
Източник на енергия за човека и животните - само химическата енергия,

Слайд 7Стационарно състояние и термодинамично равновесие
Химичните реакции в отворените системи:
рядко до равновесно
Стационарно състояние и термодинамично равновесие
Химичните реакции в отворените системи:
рядко до равновесно

Слайд 8Спрягане на екзергонични и ендергонични реакции
Доставящи енергия - окислителни, катаболитни и екзергонични
Спрягане на екзергонични и ендергонични реакции
Доставящи енергия - окислителни, катаболитни и екзергонични

Слайд 9Спрягане на процесите, доставящи и консумиращи енергия чрез макроергични съединения
Спрягане на процесите, доставящи и консумиращи енергия чрез макроергични съединения
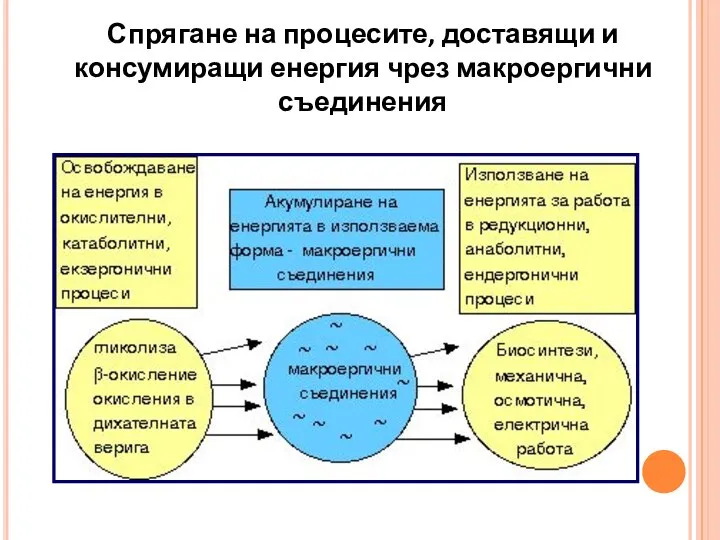
Слайд 10Едновременно или последователно протичане с общ метаболит – енергия за протичането на
Едновременно или последователно протичане с общ метаболит – енергия за протичането на

Слайд 11Макроергични съединения
Макроергични връзки: при хидролитно разграждане
ΔG°' ≥ 30 кJ/mol (до
Макроергични съединения
Макроергични връзки: при хидролитно разграждане
ΔG°' ≥ 30 кJ/mol (до

Слайд 12Пирофосфатни макроергични връзки:
Нуклеозидтрифосфати - три фосфатни връзки: α, β и γ
- α
Пирофосфатни макроергични връзки:
Нуклеозидтрифосфати - три фосфатни връзки: α, β и γ
- α
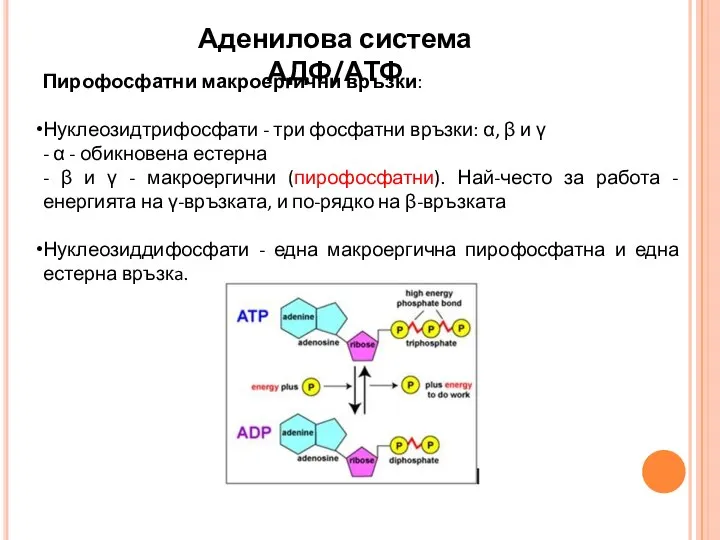
Слайд 13Примери за други макроергични съединения:
Примери за други макроергични съединения:
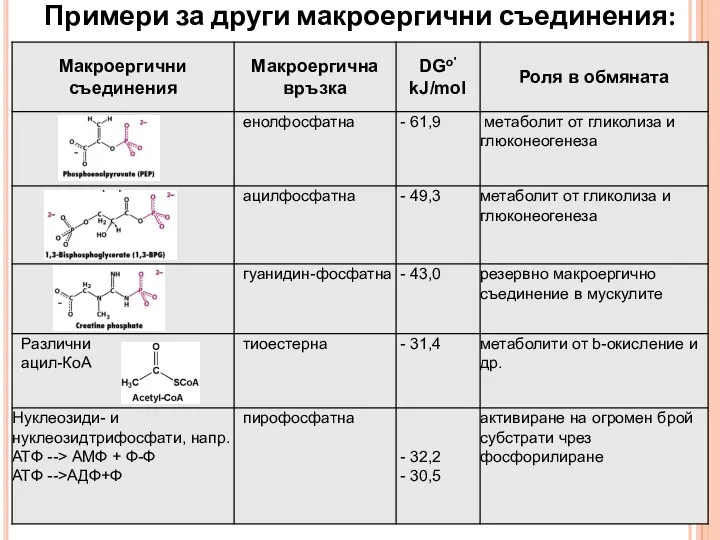
Слайд 14Различни енергетични нива на фосфорната киселина в организма
нулево енергетично ниво - свободната
Различни енергетични нива на фосфорната киселина в организма
нулево енергетично ниво - свободната

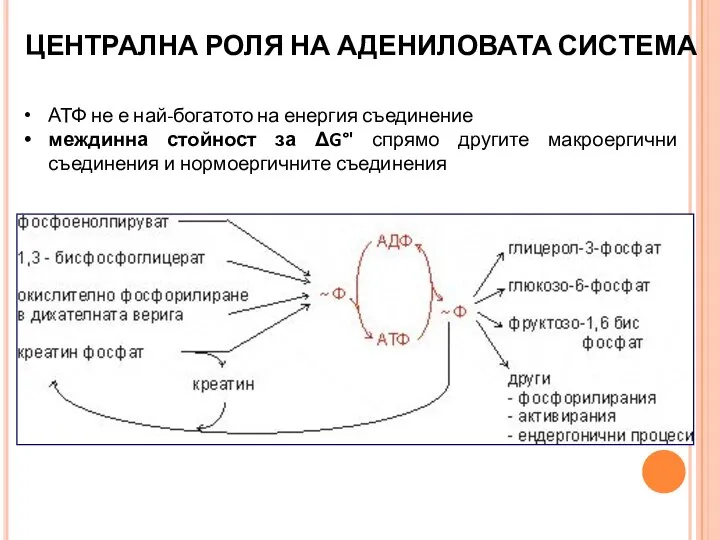
Слайд 15ЦЕНТРАЛНА РОЛЯ НА АДЕНИЛОВАТА СИСТЕМА
АТФ не е най-богатото на енергия съединение
междинна
ЦЕНТРАЛНА РОЛЯ НА АДЕНИЛОВАТА СИСТЕМА
АТФ не е най-богатото на енергия съединение
междинна
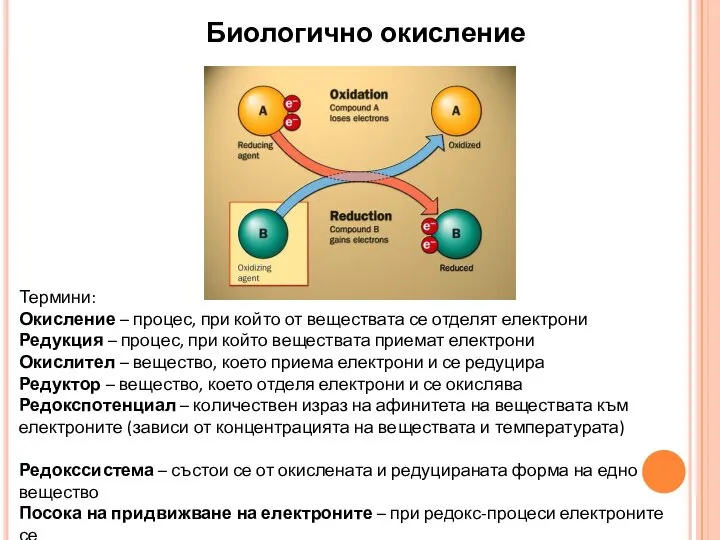
Слайд 16Биологично окисление
Термини:
Окисление – процес, при който от веществата се отделят електрони
Редукция –
Биологично окисление
Термини:
Окисление – процес, при който от веществата се отделят електрони
Редукция –
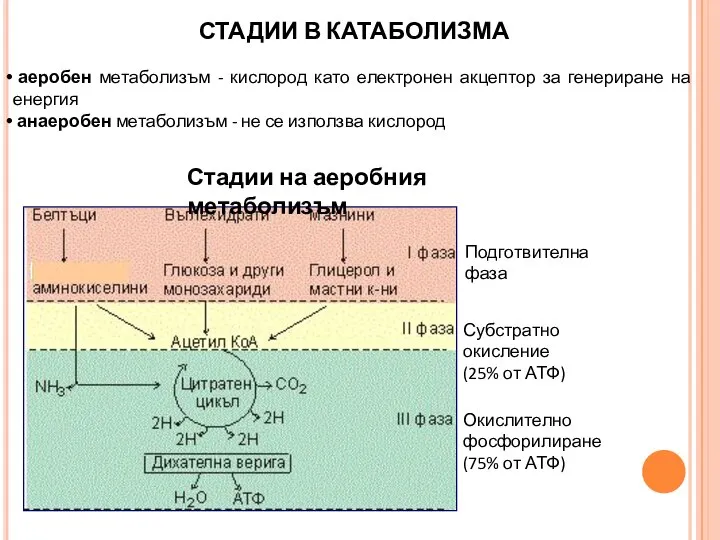
Слайд 17СТАДИИ В КАТАБОЛИЗМА
аеробен метаболизъм - кислород като електронен акцептор за генериране на
СТАДИИ В КАТАБОЛИЗМА
аеробен метаболизъм - кислород като електронен акцептор за генериране на
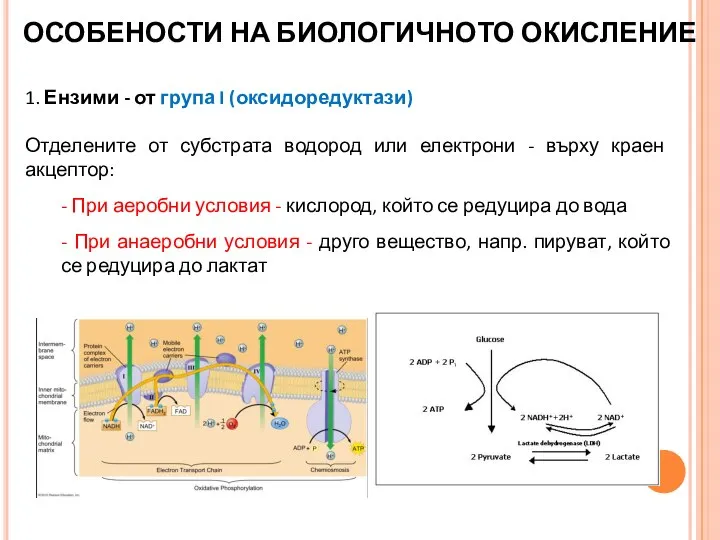
Слайд 18ОСОБЕНОСТИ НА БИОЛОГИЧНОТО ОКИСЛЕНИЕ
1. Ензими - от група I (оксидоредуктази)
Отделените от
ОСОБЕНОСТИ НА БИОЛОГИЧНОТО ОКИСЛЕНИЕ
1. Ензими - от група I (оксидоредуктази)
Отделените от
Слайд 192. Директно взаимодействие с кислород vs. многостъпално в поредица от реакции, катализирани
2. Директно взаимодействие с кислород vs. многостъпално в поредица от реакции, катализирани
Слайд 203. Всеки пренос на водород (електрони) от редокссистема с по-нисък към редокссистема
3. Всеки пренос на водород (електрони) от редокссистема с по-нисък към редокссистема
Слайд 21ОКСИДОРЕДУКТАЗИ (ЕНЗИМИ, ОСЪЩЕСТВЯВАЩИ БИОЛОГИЧНО ОКИСЛЕНИЕ)
Дехидрогенази
анаеробни, т. е. не могат да използват кислород
ОКСИДОРЕДУКТАЗИ (ЕНЗИМИ, ОСЪЩЕСТВЯВАЩИ БИОЛОГИЧНО ОКИСЛЕНИЕ)
Дехидрогенази
анаеробни, т. е. не могат да използват кислород

Слайд 223) Анаеробни транселектронази - пренос на един електрон
цитохромите от дихателната верига
напр.
3) Анаеробни транселектронази - пренос на един електрон
цитохромите от дихателната верига
напр.
Слайд 23II. Оксидази
аеробни - кислород за акцептор на водород,
образува се вода
II. Оксидази
аеробни - кислород за акцептор на водород,
образува се вода
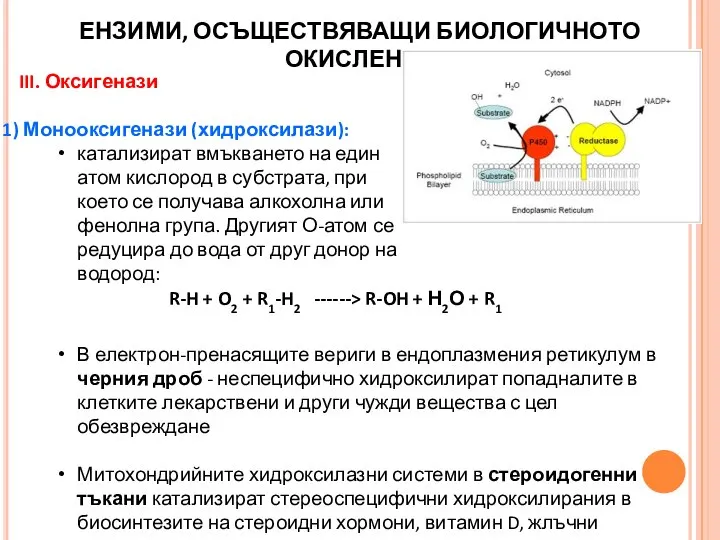
Слайд 24III. Оксигенази
Монооксигенази (хидроксилази):
катализират вмъкването на един атом кислород в субстрата, при
III. Оксигенази
Монооксигенази (хидроксилази):
катализират вмъкването на един атом кислород в субстрата, при
Слайд 252) Диоксигенази
вмъкват два атома кислород в ароматни пръстени, последвано от окислително
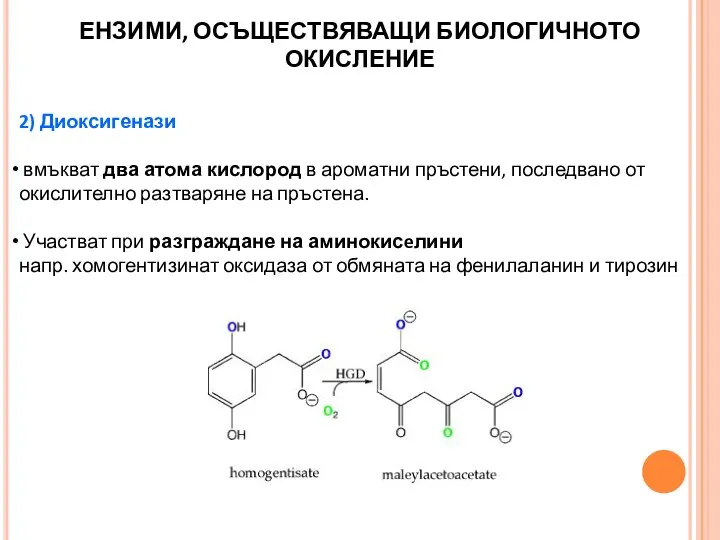
2) Диоксигенази
вмъкват два атома кислород в ароматни пръстени, последвано от окислително
Слайд 26IV. Хидроксипероксидази
разграждане на вредните за организма пероксиди и получаващите се от
IV. Хидроксипероксидази
разграждане на вредните за организма пероксиди и получаващите се от

Слайд 27РЕДОКС-СИСТЕМИ С БИОЛОГИЧНО ЗНАЧЕНИЕ
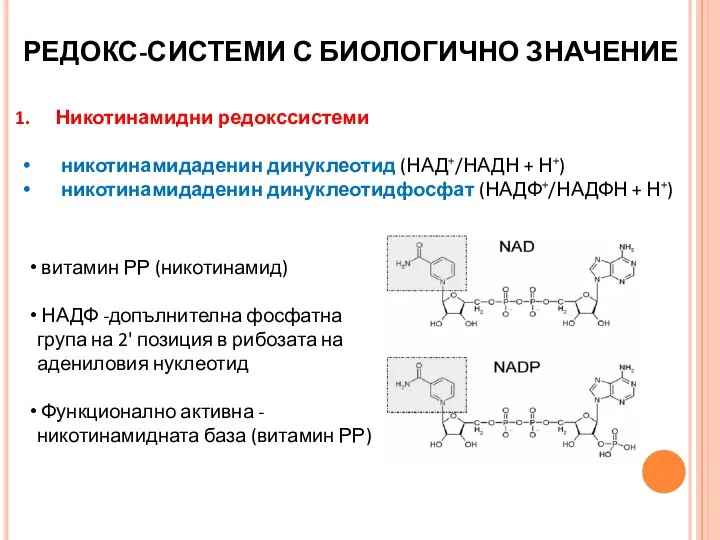
Никотинамидни редокссистеми
никотинамидаденин динуклеотид (НАД+/НАДН + Н+)
никотинамидаденин динуклеотидфосфат
РЕДОКС-СИСТЕМИ С БИОЛОГИЧНО ЗНАЧЕНИЕ
Никотинамидни редокссистеми
никотинамидаденин динуклеотид (НАД+/НАДН + Н+)
никотинамидаденин динуклеотидфосфат
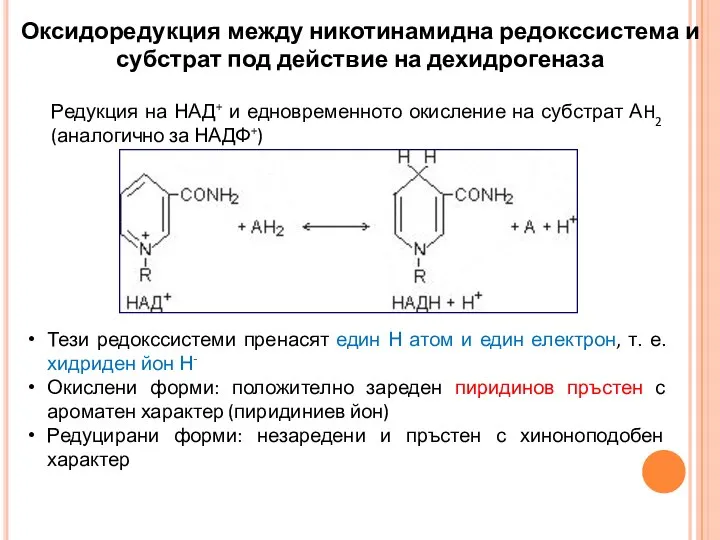
Слайд 28Тези редокссистеми пренасят един Н атом и един електрон, т. е. хидриден
Тези редокссистеми пренасят един Н атом и един електрон, т. е. хидриден
Слайд 29Никотинамидните редокссистеми: близка структура, много близък, при това нисък нормален редокспотенциал (по-нисък
Никотинамидните редокссистеми: близка структура, много близък, при това нисък нормален редокспотенциал (по-нисък

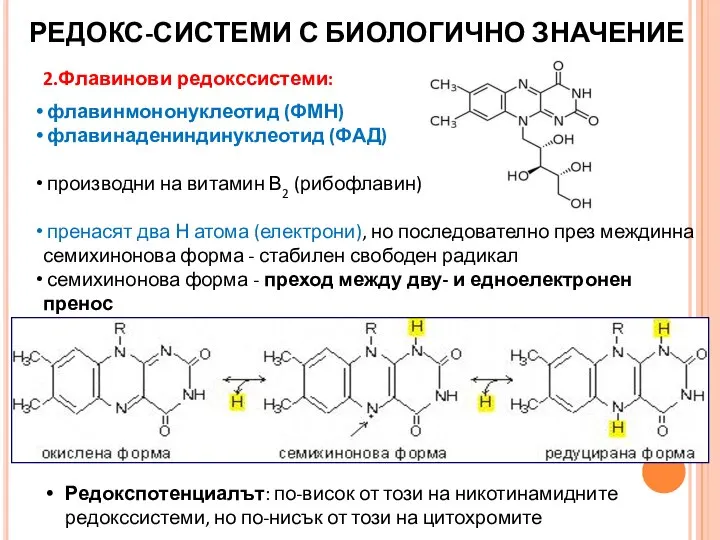
Слайд 302.Флавинови редокссистеми:
флавинмононуклеотид (ФМН)
флавинадениндинуклеотид (ФАД)
производни на витамин В2 (рибофлавин)
пренасят
2.Флавинови редокссистеми:
флавинмононуклеотид (ФМН)
флавинадениндинуклеотид (ФАД)
производни на витамин В2 (рибофлавин)
пренасят
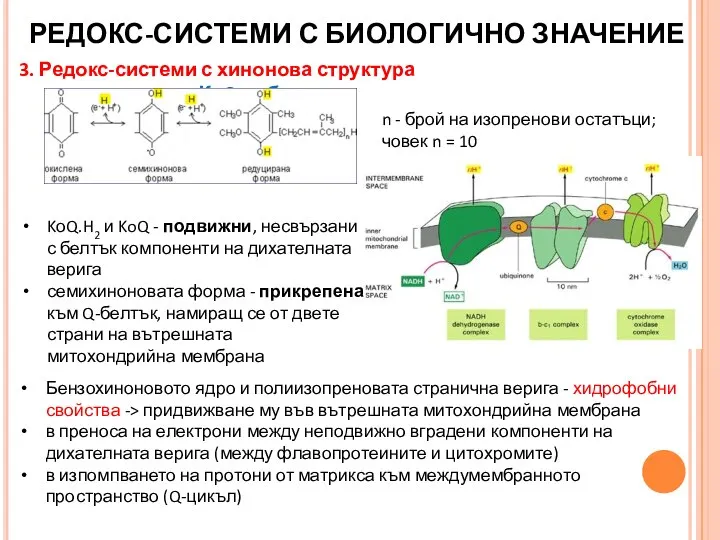
Слайд 313. Редокс-системи с хинонова структура
КоQ – убихинон
РЕДОКС-СИСТЕМИ С БИОЛОГИЧНО ЗНАЧЕНИЕ
KоQ.H2 и
3. Редокс-системи с хинонова структура
КоQ – убихинон
РЕДОКС-СИСТЕМИ С БИОЛОГИЧНО ЗНАЧЕНИЕ
KоQ.H2 и
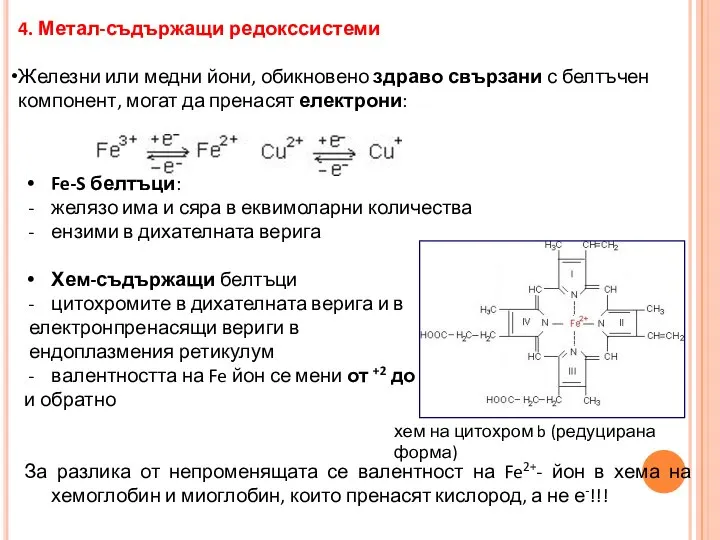
Слайд 324. Метал-съдържащи редокссистеми
Железни или медни йони, обикновено здраво свързани с белтъчен компонент,
4. Метал-съдържащи редокссистеми
Железни или медни йони, обикновено здраво свързани с белтъчен компонент,
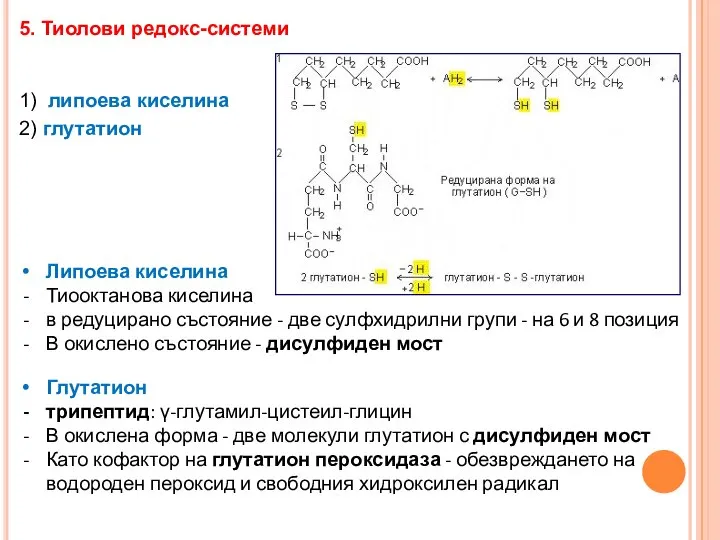
Слайд 335. Тиолови редокс-системи
1) липоева киселина
2) глутатион
Липоева киселина
Тиооктанова киселина
в редуцирано състояние
5. Тиолови редокс-системи
1) липоева киселина
2) глутатион
Липоева киселина
Тиооктанова киселина
в редуцирано състояние
Слайд 346. Аскорбинова киселина (аскорбат) - витамин С
Не се синтезира в човек; храната
добър
6. Аскорбинова киселина (аскорбат) - витамин С
Не се синтезира в човек; храната
добър
Слайд 35СУБСТРАТНО ОКИСЛЕНИЕ
едностъпално анаеробно дехидрогениране на стотици различни субстрати под действие на специфични
СУБСТРАТНО ОКИСЛЕНИЕ
едностъпално анаеробно дехидрогениране на стотици различни субстрати под действие на специфични

Слайд 36ОКИСЛИТЕЛНО ФОСФОРИЛИРАНЕ НА СУБСТРАТНО НИВО
Синтезата на АТФ или други макроергични съединения за
ОКИСЛИТЕЛНО ФОСФОРИЛИРАНЕ НА СУБСТРАТНО НИВО
Синтезата на АТФ или други макроергични съединения за

Слайд 37Скромен количествен принос, но има значение, тъй като:
В условията на кислородна недостатъчност
Скромен количествен принос, но има значение, тъй като:
В условията на кислородна недостатъчност

 Презентация на тему Многообразие птиц
Презентация на тему Многообразие птиц  Бактерии
Бактерии Методы стимуляции функций полового аппарата самок и самцов
Методы стимуляции функций полового аппарата самок и самцов Мышцы нижних конечностей
Мышцы нижних конечностей Нуклеиновые кислоты
Нуклеиновые кислоты Индукция толерогенных свойств дендритных клеток мышей линий BALB/C и C57BL/6
Индукция толерогенных свойств дендритных клеток мышей линий BALB/C и C57BL/6 Системный принцип организации природы
Системный принцип организации природы Презентация на тему Зоология наука о животных
Презентация на тему Зоология наука о животных  Телята на ферме
Телята на ферме Рождение истории биологии. (Лекция 1а)
Рождение истории биологии. (Лекция 1а) Сурок Байбак
Сурок Байбак Рефлекторная регуляция
Рефлекторная регуляция Углеводы. Термин углеводы
Углеводы. Термин углеводы Человек и биосфера
Человек и биосфера Animales varios
Animales varios The major schools of psychology
The major schools of psychology Грибы. Тело гриба
Грибы. Тело гриба Эволюция органического мира
Эволюция органического мира Адаптация к ультрафиолетовому излучению Солнца
Адаптация к ультрафиолетовому излучению Солнца Моллюски Байкала
Моллюски Байкала Презентация. Биотехнология. 10 класс
Презентация. Биотехнология. 10 класс Семейство луковые
Семейство луковые Анализ заболеваемости и смертности экзотических и диких животных, содержащихся в неволе
Анализ заболеваемости и смертности экзотических и диких животных, содержащихся в неволе Сердечно-сосудистая система
Сердечно-сосудистая система 1-2-3 урок
1-2-3 урок Як утворився торф і кам`яне вугілля
Як утворився торф і кам`яне вугілля Biological factors
Biological factors Зачем подкармливать птиц зимой?
Зачем подкармливать птиц зимой?