- Биофизика мембран

Содержание
- 8. Типичные мембранные белки в своей родной среде обитания — липидном бислое, за пределами которого они теряют
- 12. Мозаичная организация поверхности простейшей однокомпонентной мембраны. Слева представлена идеальная модель мембраны, справа — поверхность полноатомной мембраны
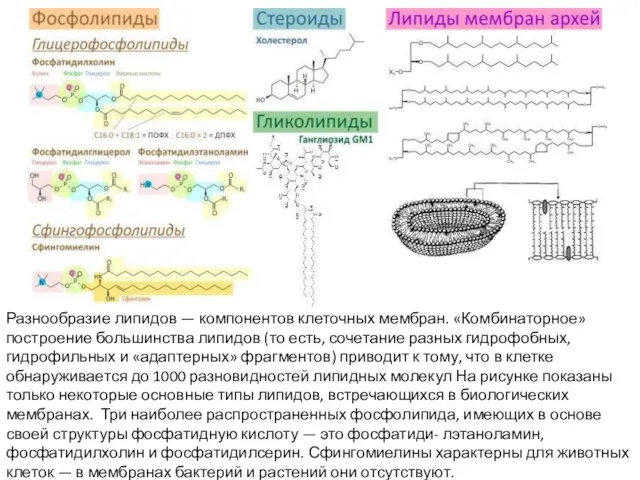
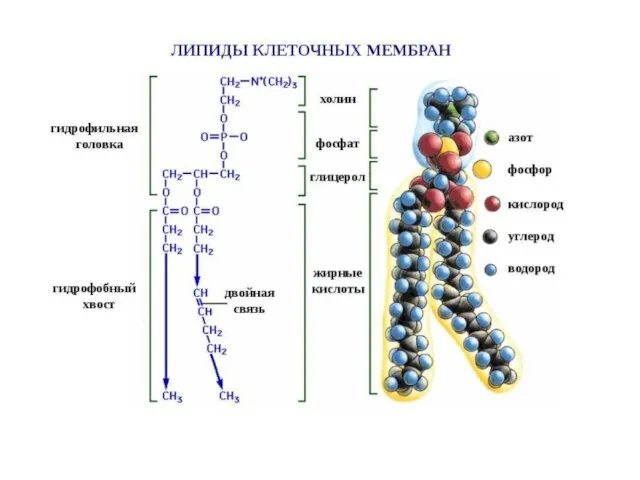
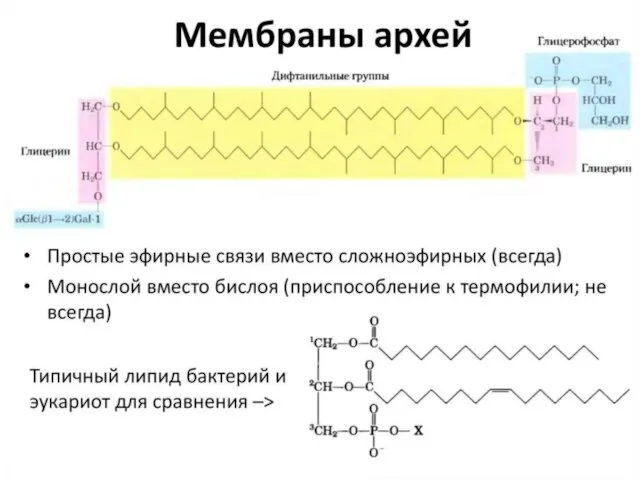
- 14. Разнообразие липидов — компонентов клеточных мембран. «Комбинаторное» построение большинства липидов (то есть, сочетание разных гидрофобных, гидрофильных
- 24. Текучесть липидной фазы мембраны обусловлена присутствием в углеводородных цепях большинства структурных фосфолипидов минимум одной ненасыщенной связи,
- 30. Распределение липидов и белков в плоскости мембраны не однородно, а, напротив, обладает характерной структурой, — это
- 31. липидная компонента, будучи жидкой, тем не менее, способна образовывать частично изолированные области бислоя, обладающие особыми структурными
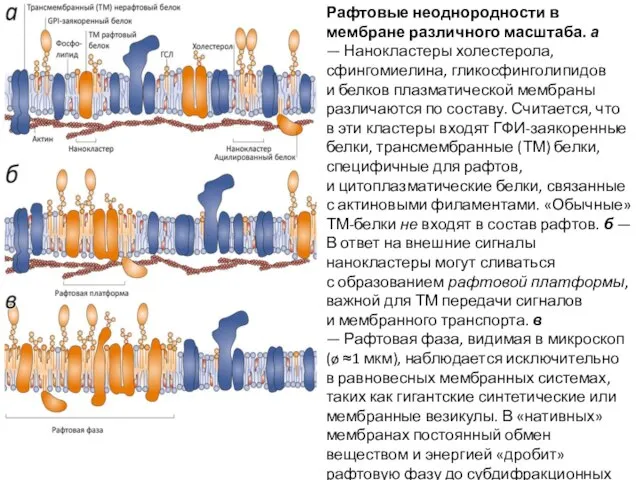
- 33. Рафтовые неоднородности в мембране различного масштаба. а — Нанокластеры холестерола, сфингомиелина, гликосфинголипидов и белков плазматической мембраны
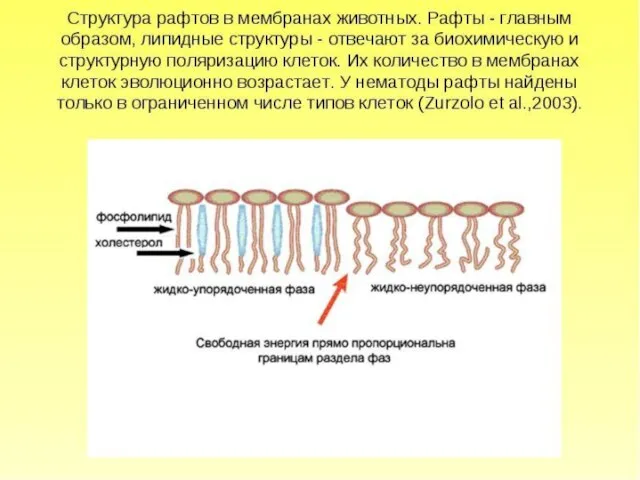
- 37. Состояние мембраны неравновесно. Оно может быть стационарным (когда концентрации разных липидов сохраняются примерно на том же
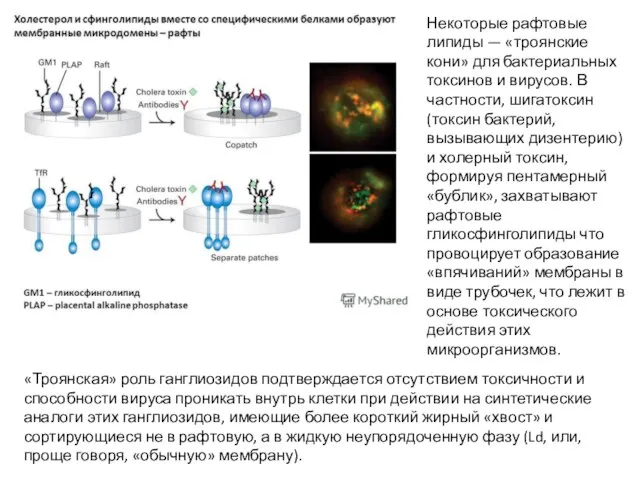
- 39. Некоторые рафтовые липиды — «троянские кони» для бактериальных токсинов и вирусов. В частности, шигатоксин (токсин бактерий,
- 40. Многие вирусы, покидая зараженную ранее клетку, облачаются в липидную оболочку — часть мембраны клетки-хозяина. Некоторые из
- 42. Скачать презентацию

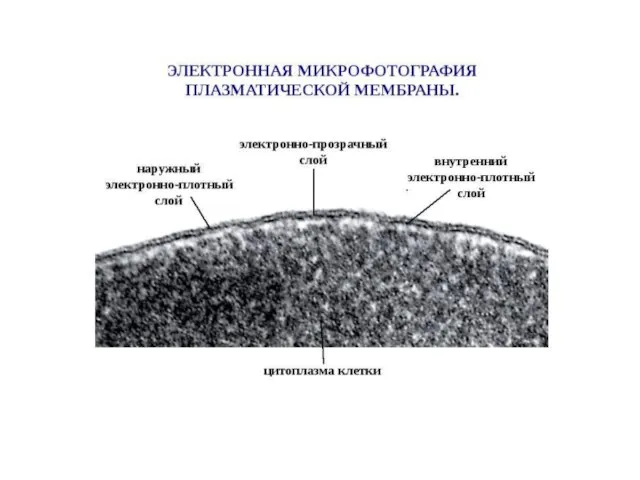

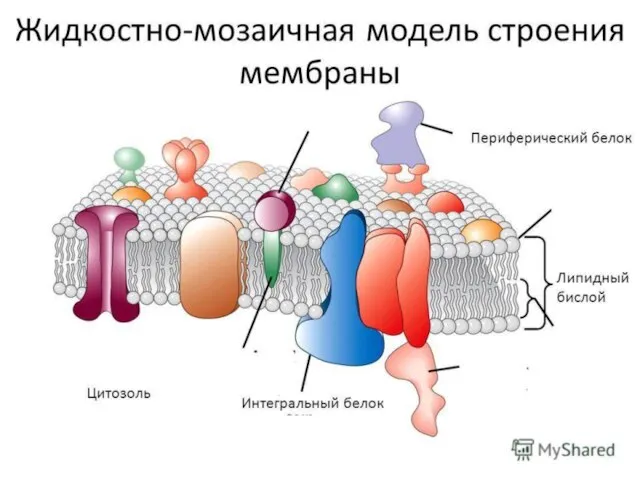
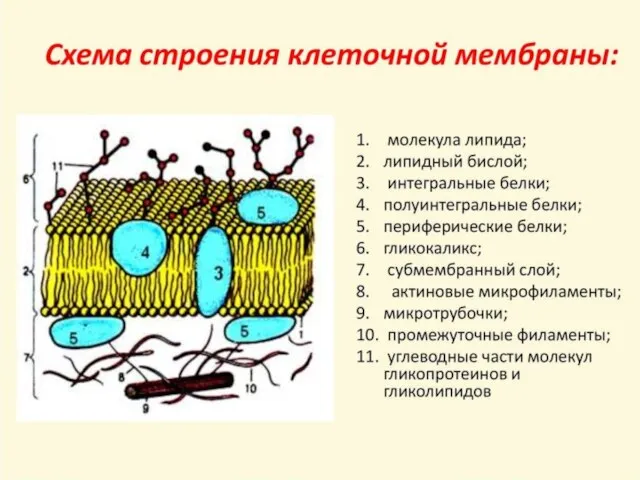
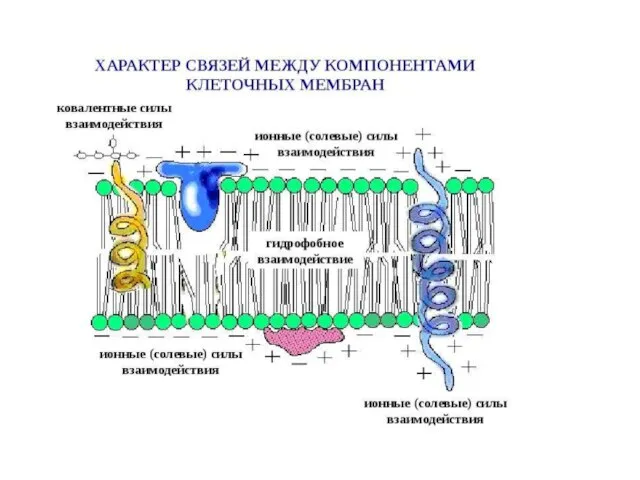
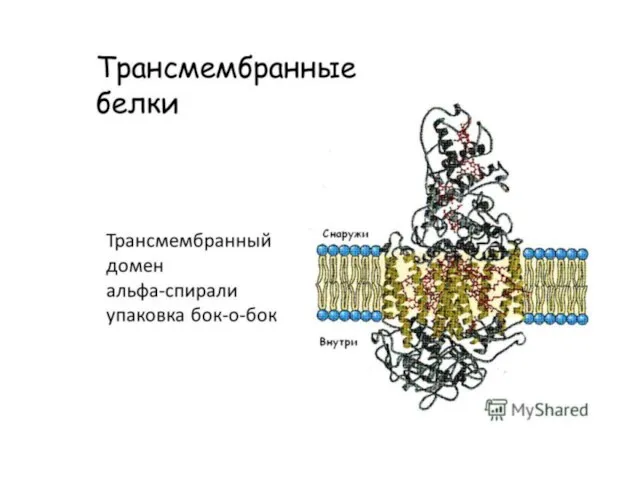
Слайд 8Типичные мембранные белки в своей родной среде обитания — липидном бислое, за пределами которого они
Типичные мембранные белки в своей родной среде обитания — липидном бислое, за пределами которого они


Слайд 12Мозаичная организация поверхности простейшей однокомпонентной мембраны. Слева представлена идеальная модель мембраны, справа
Мозаичная организация поверхности простейшей однокомпонентной мембраны. Слева представлена идеальная модель мембраны, справа

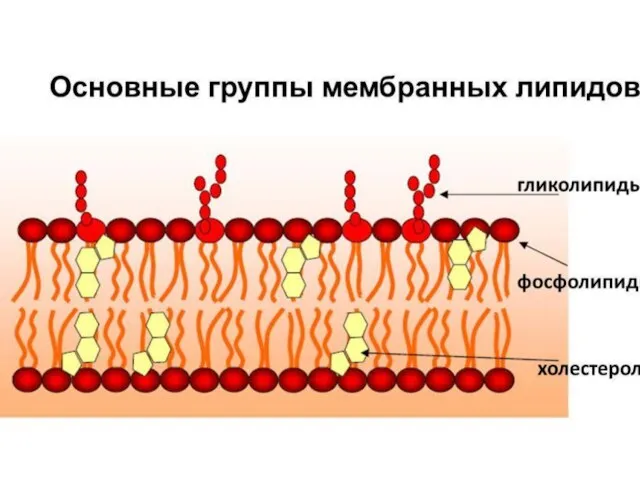
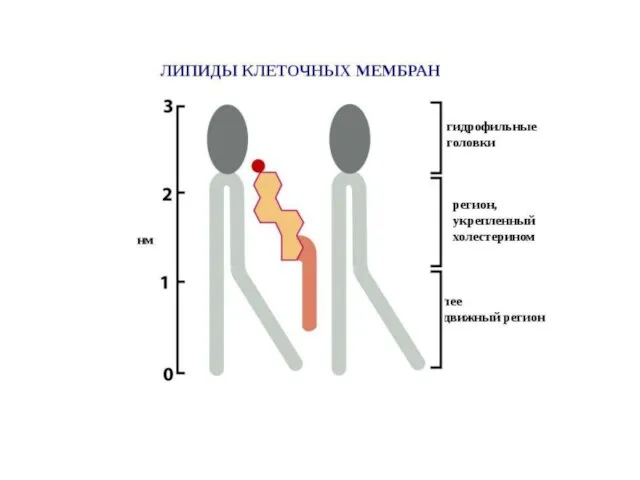
Слайд 14Разнообразие липидов — компонентов клеточных мембран. «Комбинаторное» построение большинства липидов (то есть,
Разнообразие липидов — компонентов клеточных мембран. «Комбинаторное» построение большинства липидов (то есть,


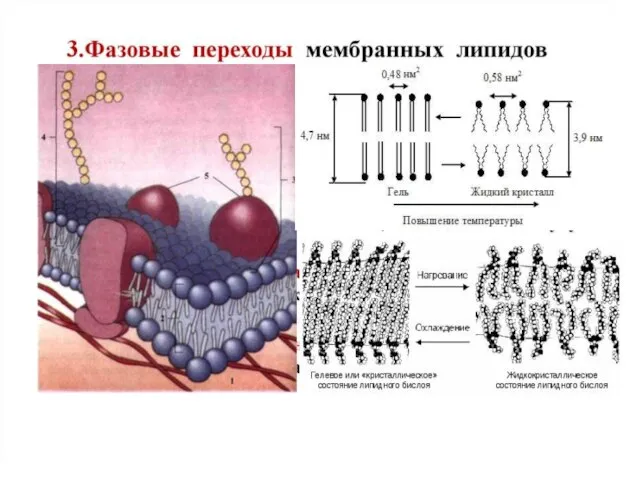



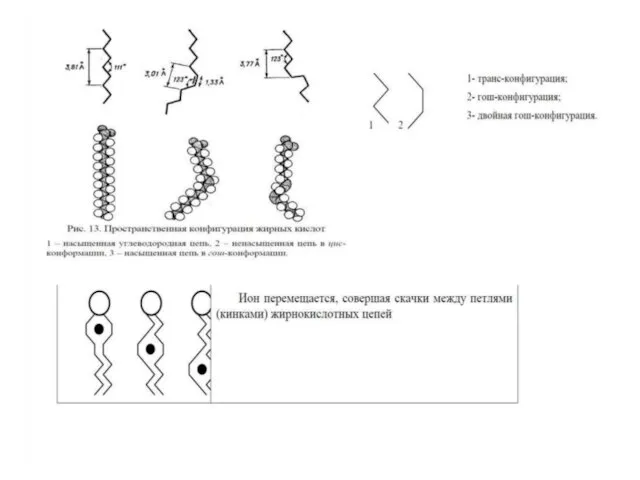
Слайд 24Текучесть липидной фазы мембраны обусловлена присутствием в углеводородных цепях большинства структурных фосфолипидов минимум
Текучесть липидной фазы мембраны обусловлена присутствием в углеводородных цепях большинства структурных фосфолипидов минимум


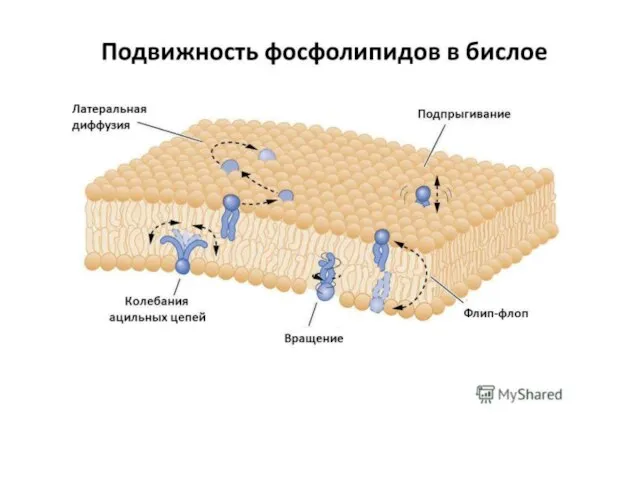
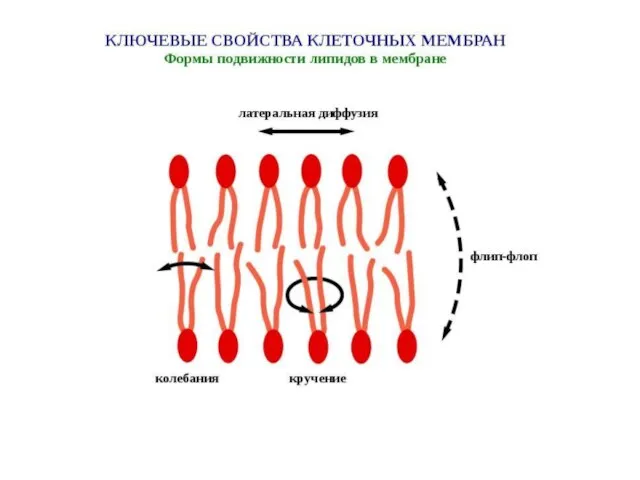
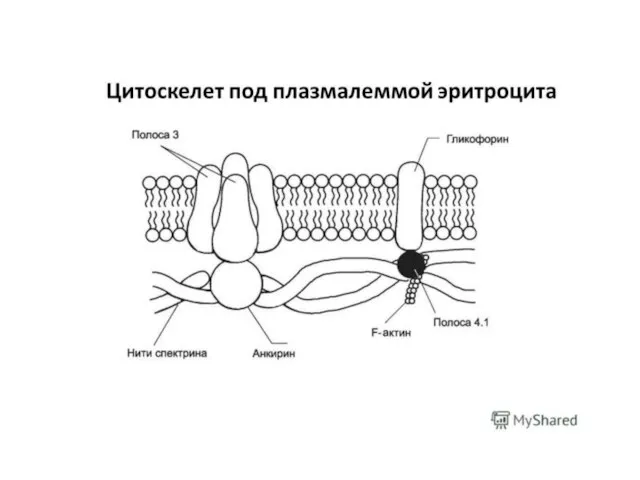
Слайд 30Распределение липидов и белков в плоскости мембраны не однородно, а, напротив, обладает
Распределение липидов и белков в плоскости мембраны не однородно, а, напротив, обладает

Слайд 31липидная компонента, будучи жидкой, тем не менее, способна образовывать частично изолированные области
липидная компонента, будучи жидкой, тем не менее, способна образовывать частично изолированные области


Слайд 33Рафтовые неоднородности в мембране различного масштаба. а — Нанокластеры холестерола, сфингомиелина, гликосфинголипидов и белков
Рафтовые неоднородности в мембране различного масштаба. а — Нанокластеры холестерола, сфингомиелина, гликосфинголипидов и белков


Слайд 37Состояние мембраны неравновесно.
Оно может быть стационарным (когда концентрации разных липидов сохраняются
Состояние мембраны неравновесно.
Оно может быть стационарным (когда концентрации разных липидов сохраняются


Слайд 39Некоторые рафтовые липиды — «троянские кони» для бактериальных токсинов и вирусов. В
Некоторые рафтовые липиды — «троянские кони» для бактериальных токсинов и вирусов. В
Слайд 40Многие вирусы, покидая зараженную ранее клетку, облачаются в липидную оболочку — часть
Многие вирусы, покидая зараженную ранее клетку, облачаются в липидную оболочку — часть

 Технологии животноводства. Заболевания животных и их предупреждение
Технологии животноводства. Заболевания животных и их предупреждение Строение головного мозга. Функции продолговатого мозга, моста, мозжечка и среднего мозга
Строение головного мозга. Функции продолговатого мозга, моста, мозжечка и среднего мозга Агамы
Агамы Животный мир Ленинградской области
Животный мир Ленинградской области Плауновидные растения
Плауновидные растения Вегетативное размножение растений черенками
Вегетативное размножение растений черенками Посадка дерева, кустарника
Посадка дерева, кустарника Лесная аптека на службе человека
Лесная аптека на службе человека Видоизменение побегов. Луковица
Видоизменение побегов. Луковица Семейство Тироглифоидные. Представитель: Домашний клещ
Семейство Тироглифоидные. Представитель: Домашний клещ Рослини
Рослини Поверхностные мышцы спины (первый слой)
Поверхностные мышцы спины (первый слой) Предостерегающая окраска
Предостерегающая окраска Влияние биоритмов на умственную работоспособность учащихся
Влияние биоритмов на умственную работоспособность учащихся Животные леса: земноводные и пресмыкающиеся
Животные леса: земноводные и пресмыкающиеся Биология – наука о живой природе
Биология – наука о живой природе Презентация на тему Мутации (11 класс)
Презентация на тему Мутации (11 класс)  Презентация на тему ФАКТОРЫ ЗДОРОВЬЯ
Презентация на тему ФАКТОРЫ ЗДОРОВЬЯ  Покрытосеменные или цветковые растения
Покрытосеменные или цветковые растения Строение клетки
Строение клетки Высшая нервная деятельность детей и подростков
Высшая нервная деятельность детей и подростков Однодольные и двудольные растения. Бинарная номенклатура
Однодольные и двудольные растения. Бинарная номенклатура Алоэ - комнатные растения
Алоэ - комнатные растения Класс земноводные или амфибии
Класс земноводные или амфибии 3. Общая характеритика царства растений_ (6 класс)
3. Общая характеритика царства растений_ (6 класс) Бактерии
Бактерии Ткани растений
Ткани растений Нарушения теплового баланса
Нарушения теплового баланса