- Биопотенциал. Его физиологическое значение. Биофизика нервного импульса. Синаптическая передача

Содержание
- 2. Генерация и распространение электрических потенциалов лежит в основе: 1) регуляции внутриклеточных процессов; 2) генерации и распространению
- 3. Изучение клеточных потенциалов стало возможным благодаря созданию устройств, обладающих: 1) очень большим входным сопротивлением (до 1010
- 4. Мембранные потенциалы φs1 φs2 φм ∆φ φs1 и φs2 – разности потенциалов между мембраной и окружающей
- 5. Донановское равновесие и потенциал Донана. Устанавливается между клеткой и окружающей средой, при условии, что мембрана проницаема
- 6. Донановское равновесие и потенциал Донана. В основе представления о равновесном потенциале такой системы лежит представление о
- 7. Донановское равновесие и потенциал Донана. [Ko+] = [Ao-]=C0; [Ki+] = [Ai-]+ n[P-i]. По уравнению Нернста разность
- 8. Донановское равновесие и потенциал Донана. ψ= n[P-i]/2Co φ=RT/zF• n[P-i]/2Co Донановский потенциал прямо пропорционален концентрации белков в
- 9. Равновесный потенциал. [К+о] [К+in] Δφ Δφ = RT/zF●In([К+о]/ [К+in]) Согласно гипотезе, выдвинутой в 1902 году Бернштейном,
- 10. Стационарный потенциал. Ходжкин, Хаксли и Катц исходили из того, что потенциал покоя не равновесный, а стационарный.
- 11. Концентрация ионов (мМ/л) и разность потенциалов между двумя сторонами клеточной мембраны в состоянии покоя (из Волькенштейн
- 12. Равновесные Нернстовские потенциалы для ионов натрия, калия и хлора в состоянии покоя. ∆φ=RT/zF•In [Э out]/[Э in]
- 13. Потенциал действия -50 50 0 0 5 10 t,c При возбуждении нервных клеток, клеток мышц происходит
- 14. Потенциал действия В изучении ПД большой вклад внесли работы на гигантском аксоне кальмара. Он достаточно крупный,
- 15. Потенциал действия Величина ПД не зависит от величины стимулирующего тока и ПД не возникает, если электрический
- 16. Потенциал действия За счет каких процессов происходит изменение потенциала при прохождении ПД? Эксперименты с радиоактивными изотопами
- 17. Потенциал действия 2 4 t j 0 0,1 -0,1 K+ 4 2 0 0,1 -0,1 j
- 18. Распространение потенциала действия по нервному волокну. EK+ ENa+ EY C Наружный раствор аксоплазма Удельное сопротивление аксоплазмы
- 19. Распространение потенциала действия по нервному волокну. EK+ ENa+ EY C Наружный раствор аксоплазма Для скорейшего распространения
- 20. Синаптическая передача. Когда импульс достигает окончания нервного волокна, он либо переходит на другое волокно, либо на
- 21. Электрические синапсы - это прямые ионпроводящие соединения клеток посредством каналов (коннексонов) в области так называемых щелевых
- 22. Медиаторы высвобождаются путем регулируемого экзоцитоза так называемых синаптических пузырьков. Каждый пузырек содержит некоторое количество («квант») нейромедиатора.
- 23. Потенциал действия (1) повышает вероятность открывания потенциалзависимых Са2+-каналов в пресинаптической мембране, приводя к увеличению концентрации Са2+
- 24. АХЭ АХ АХ АХ АХ АХ Постсинаптическая мембрана Постсинаптическая мембрана содержит рецепторный белок (РБ), пронзающий мембрану
- 25. Механизм зрительного восприятия 1 2 3 4 5 6 7 1-сетчатка; 2-зрительный нерв; 3-хрусталик; 4-цилиарная мышца;
- 26. Механизм зрительного восприятия hν 1 2 3 4 5 6 7 8 9 Здесь приведена структура
- 27. Механизм зрительного восприятия hν 1 2 3 4 5 6 7 8 9 Горизонтальные (4) и
- 28. Молекулярный механизм фоторецепции Юнг, а позднее Максвелл и Гельмгольц предложили существование трех видов светочувствительных рецепторов в
- 29. Молекулярный механизм фоторецепции В фоторецепторах ретинол существует в альдегидной форме-ретиналь. Он связан с липопротеином опсином и
- 31. Скачать презентацию
Слайд 2Генерация и распространение электрических потенциалов лежит в основе:
1) регуляции внутриклеточных процессов;
2)
Генерация и распространение электрических потенциалов лежит в основе: 1) регуляции внутриклеточных процессов; 2)

Слайд 3Изучение клеточных потенциалов стало возможным благодаря созданию устройств, обладающих:
1) очень большим входным
Изучение клеточных потенциалов стало возможным благодаря созданию устройств, обладающих: 1) очень большим входным

Слайд 4Мембранные потенциалы
φs1
φs2
φм
∆φ
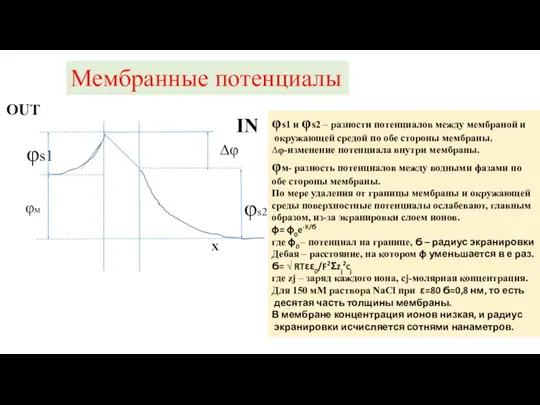
φs1 и φs2 – разности потенциалов между мембраной и
окружающей
Мембранные потенциалы
φs1
φs2
φм
∆φ
φs1 и φs2 – разности потенциалов между мембраной и
окружающей
Слайд 5Донановское равновесие и потенциал Донана.
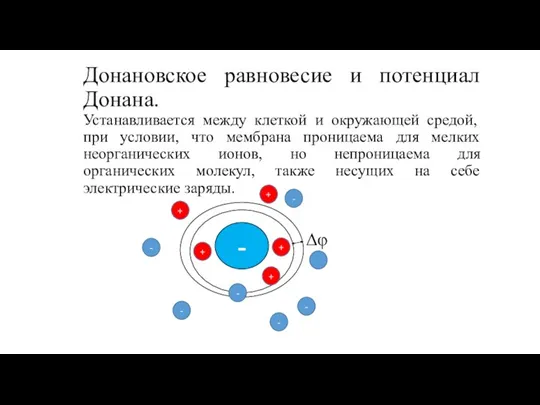
Устанавливается между клеткой и окружающей средой, при условии,
Донановское равновесие и потенциал Донана. Устанавливается между клеткой и окружающей средой, при условии,
Слайд 6Донановское равновесие и потенциал Донана.
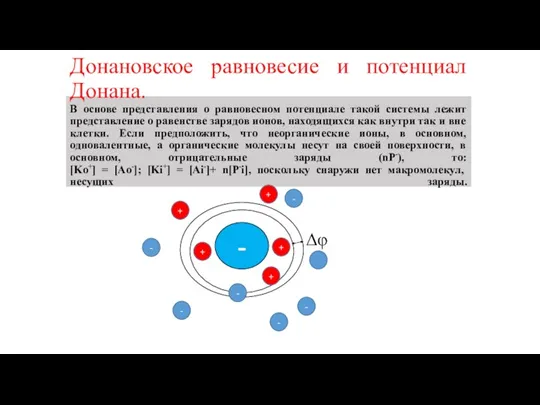
В основе представления о равновесном потенциале такой системы
Донановское равновесие и потенциал Донана. В основе представления о равновесном потенциале такой системы
Слайд 7Донановское равновесие и потенциал Донана.
[Ko+] = [Ao-]=C0; [Ki+] = [Ai-]+ n[P-i].
По уравнению
Донановское равновесие и потенциал Донана. [Ko+] = [Ao-]=C0; [Ki+] = [Ai-]+ n[P-i]. По уравнению
![Донановское равновесие и потенциал Донана. [Ko+] = [Ao-]=C0; [Ki+] = [Ai-]+ n[P-i].](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1075745/slide-6.jpg)
Слайд 8Донановское равновесие и потенциал Донана.
ψ= n[P-i]/2Co
φ=RT/zF• n[P-i]/2Co
Донановский потенциал прямо пропорционален
Донановское равновесие и потенциал Донана.
ψ= n[P-i]/2Co
φ=RT/zF• n[P-i]/2Co
Донановский потенциал прямо пропорционален
![Донановское равновесие и потенциал Донана. ψ= n[P-i]/2Co φ=RT/zF• n[P-i]/2Co Донановский потенциал прямо](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1075745/slide-7.jpg)
Слайд 9Равновесный потенциал.
[К+о]
[К+in]
Δφ
Δφ = RT/zF●In([К+о]/ [К+in])
Согласно гипотезе, выдвинутой в 1902 году Бернштейном,
Потенциал покоя
Равновесный потенциал.
[К+о]
[К+in]
Δφ
Δφ = RT/zF●In([К+о]/ [К+in])
Согласно гипотезе, выдвинутой в 1902 году Бернштейном,
Потенциал покоя
![Равновесный потенциал. [К+о] [К+in] Δφ Δφ = RT/zF●In([К+о]/ [К+in]) Согласно гипотезе, выдвинутой](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1075745/slide-8.jpg)
Слайд 10Стационарный потенциал.
Ходжкин, Хаксли и Катц исходили из того, что потенциал покоя не
Стационарный потенциал.
Ходжкин, Хаксли и Катц исходили из того, что потенциал покоя не

Слайд 11Концентрация ионов (мМ/л) и разность потенциалов между двумя сторонами клеточной мембраны в
Концентрация ионов (мМ/л) и разность потенциалов между двумя сторонами клеточной мембраны в

Слайд 12Равновесные Нернстовские потенциалы для ионов натрия, калия и хлора в состоянии покоя.
Равновесные Нернстовские потенциалы для ионов натрия, калия и хлора в состоянии покоя.

Слайд 13Потенциал действия
-50
50
0
0
5
10
t,c
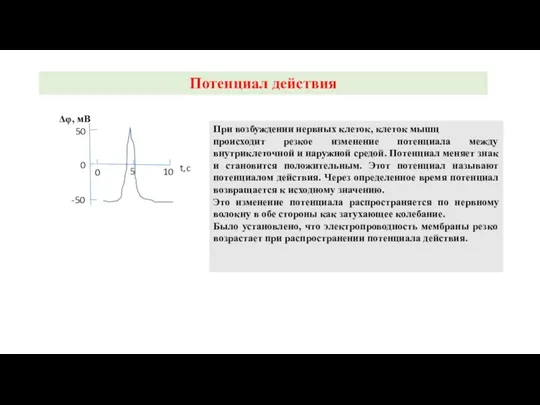
При возбуждении нервных клеток, клеток мышц
происходит резкое изменение потенциала между
Потенциал действия
-50
50
0
0
5
10
t,c
При возбуждении нервных клеток, клеток мышц
происходит резкое изменение потенциала между
Слайд 14Потенциал действия
В изучении ПД большой вклад внесли работы на гигантском аксоне кальмара.
Потенциал действия
В изучении ПД большой вклад внесли работы на гигантском аксоне кальмара.

Слайд 15Потенциал действия
Величина ПД не зависит от величины стимулирующего тока и ПД не
Потенциал действия
Величина ПД не зависит от величины стимулирующего тока и ПД не

Слайд 16Потенциал действия
За счет каких процессов происходит изменение потенциала при прохождении ПД? Эксперименты
Потенциал действия
За счет каких процессов происходит изменение потенциала при прохождении ПД? Эксперименты

Слайд 17Потенциал действия
2
4
t
j
0
0,1
-0,1
K+
4
2
0
0,1
-0,1
j
t
Na+
Плотность тока через мембрану измеряли при постоянном мембранном
потенциале. За + принимался
Потенциал действия
2
4
t
j
0
0,1
-0,1
K+
4
2
0
0,1
-0,1
j
t
Na+
Плотность тока через мембрану измеряли при постоянном мембранном
потенциале. За + принимался

Слайд 18Распространение потенциала действия по нервному волокну.
EK+
ENa+
EY
C
Наружный раствор
аксоплазма
Удельное сопротивление аксоплазмы равно 10-100 Ом●см.
Распространение потенциала действия по нервному волокну.
EK+
ENa+
EY
C
Наружный раствор
аксоплазма
Удельное сопротивление аксоплазмы равно 10-100 Ом●см.

Слайд 19Распространение потенциала действия по нервному волокну.
EK+
ENa+
EY
C
Наружный раствор
аксоплазма
Для скорейшего распространения импульса по волокну
Распространение потенциала действия по нервному волокну.
EK+
ENa+
EY
C
Наружный раствор
аксоплазма
Для скорейшего распространения импульса по волокну

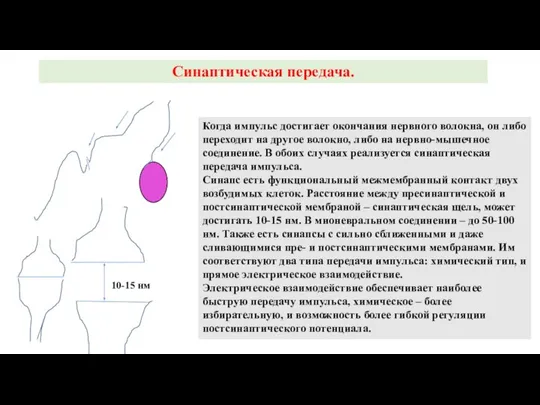
Слайд 20Синаптическая передача.
Когда импульс достигает окончания нервного волокна, он либо переходит на другое
Синаптическая передача.
Когда импульс достигает окончания нервного волокна, он либо переходит на другое
Слайд 21Электрические синапсы - это прямые ионпроводящие соединения клеток посредством каналов (коннексонов) в
Электрические синапсы - это прямые ионпроводящие соединения клеток посредством каналов (коннексонов) в

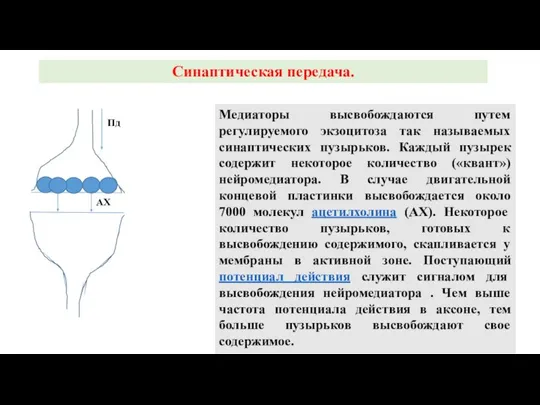
Слайд 22Медиаторы высвобождаются путем регулируемого экзоцитоза так называемых синаптических пузырьков. Каждый пузырек содержит
Медиаторы высвобождаются путем регулируемого экзоцитоза так называемых синаптических пузырьков. Каждый пузырек содержит
Слайд 23Потенциал действия (1) повышает вероятность открывания потенциалзависимых Са2+-каналов в пресинаптической мембране, приводя
Потенциал действия (1) повышает вероятность открывания потенциалзависимых Са2+-каналов в пресинаптической мембране, приводя

Слайд 24АХЭ
АХ
АХ
АХ
АХ
АХ
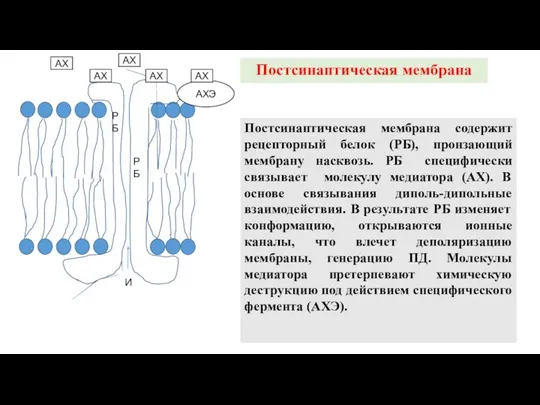
Постсинаптическая мембрана
Постсинаптическая мембрана содержит рецепторный белок (РБ), пронзающий мембрану насквозь. РБ специфически
АХЭ
АХ
АХ
АХ
АХ
АХ
Постсинаптическая мембрана
Постсинаптическая мембрана содержит рецепторный белок (РБ), пронзающий мембрану насквозь. РБ специфически
Слайд 25Механизм зрительного восприятия
1
2
3
4
5
6
7
1-сетчатка;
2-зрительный нерв;
3-хрусталик;
4-цилиарная мышца;
5-радужная оболочка;
6-роговица;
7-зрачок
Механизм зрительного восприятия
1
2
3
4
5
6
7
1-сетчатка;
2-зрительный нерв;
3-хрусталик;
4-цилиарная мышца;
5-радужная оболочка;
6-роговица;
7-зрачок

Слайд 26Механизм зрительного восприятия
hν
1
2
3
4
5
6
7
8
9
Здесь приведена структура сетчатки. Свет, прежде чем достичь рецепторных клеток
Механизм зрительного восприятия
hν
1
2
3
4
5
6
7
8
9
Здесь приведена структура сетчатки. Свет, прежде чем достичь рецепторных клеток

Слайд 27Механизм зрительного восприятия
hν
1
2
3
4
5
6
7
8
9
Горизонтальные (4) и амакриновые (6) клетки соединяют соседние фоторецепторы, обеспечивая
Механизм зрительного восприятия
hν
1
2
3
4
5
6
7
8
9
Горизонтальные (4) и амакриновые (6) клетки соединяют соседние фоторецепторы, обеспечивая

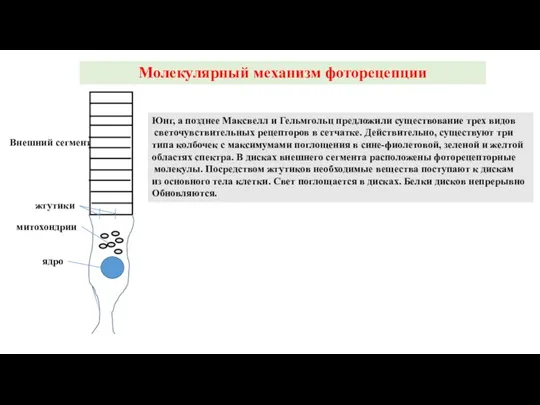
Слайд 28Молекулярный механизм фоторецепции
Юнг, а позднее Максвелл и Гельмгольц предложили существование трех видов
Молекулярный механизм фоторецепции
Юнг, а позднее Максвелл и Гельмгольц предложили существование трех видов
Слайд 29Молекулярный механизм фоторецепции
В фоторецепторах ретинол существует в альдегидной форме-ретиналь. Он связан с
Молекулярный механизм фоторецепции
В фоторецепторах ретинол существует в альдегидной форме-ретиналь. Он связан с

 Презентация на тему Мхи, папоротники, хвощи и плауны
Презентация на тему Мхи, папоротники, хвощи и плауны  День птиц
День птиц Презентация на тему Тип Членистоногие
Презентация на тему Тип Членистоногие  Арахнология
Арахнология Презентация на тему Белки и аминокислоты
Презентация на тему Белки и аминокислоты  Опорно двигательная система
Опорно двигательная система Презентация на тему Покрытосеменные растения (6 класс)
Презентация на тему Покрытосеменные растения (6 класс)  Определение хронобиотипа учащихся
Определение хронобиотипа учащихся Элиминация - гибель организмов вследствие различных абиотических и биотических факторов внешней среды
Элиминация - гибель организмов вследствие различных абиотических и биотических факторов внешней среды Презентация на тему Многообразие водорослей и их значение
Презентация на тему Многообразие водорослей и их значение  Условно-рефлекторное поведение и научение. Концепция И.П. Павлова
Условно-рефлекторное поведение и научение. Концепция И.П. Павлова Эндокринная система и половые гормоны. Взаимосвязь гипофиза и семенника
Эндокринная система и половые гормоны. Взаимосвязь гипофиза и семенника Механизмы проведения возбуждения
Механизмы проведения возбуждения Обмен углеводов. Гликолиз
Обмен углеводов. Гликолиз Бактериофаги. Роль бактериофагов в биосфере
Бактериофаги. Роль бактериофагов в биосфере Жизненный цикл клетки. Митоз
Жизненный цикл клетки. Митоз Презентация на тему РАСТЕНИЯ СЕМЕЙСТВ КРЕСТОЦВЕТНЫХ И ПАСЛЕНОВЫХ
Презентация на тему РАСТЕНИЯ СЕМЕЙСТВ КРЕСТОЦВЕТНЫХ И ПАСЛЕНОВЫХ  Жить здорово
Жить здорово Презентация на тему Словарная работа
Презентация на тему Словарная работа  Мир собак
Мир собак Экологический мониторинг рассады
Экологический мониторинг рассады Обмен нуклеопротеинов
Обмен нуклеопротеинов Сайты для подготовки к ВПР
Сайты для подготовки к ВПР Полесские робинзоны. Республиканская квест-игра
Полесские робинзоны. Республиканская квест-игра Sos and cor
Sos and cor Дегелек. Дегелектің тұқымдастары
Дегелек. Дегелектің тұқымдастары Времена года в Крыму
Времена года в Крыму Памятники природы
Памятники природы