- Физиология дыхания. Дыхательная система человека

Содержание
- 2. Основные этапы процесса дыхания Клетки нашего организма получают энергию с использованием кислорода. Из-за больших расстояний между
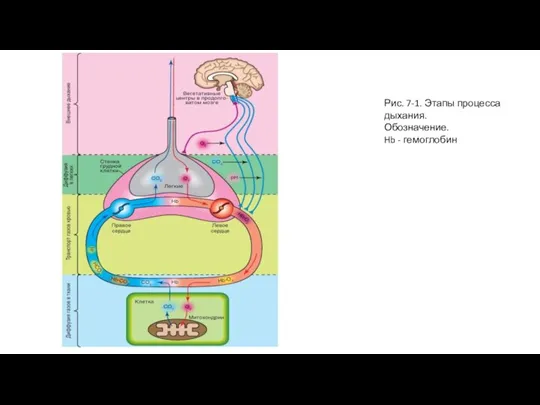
- 3. Рис. 7-1. Этапы процесса дыхания. Обозначение. Hb - гемоглобин
- 4. Основные физические характеристики компонентов газовой смеси Важнейшими составляющими транспортной системы для внешнего дыхания являются вентиляция, диффузия
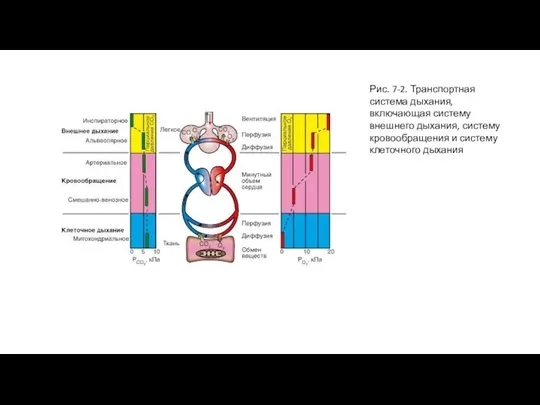
- 5. Рис. 7-2. Транспортная система дыхания, включающая систему внешнего дыхания, систему кровообращения и систему клеточного дыхания
- 6. Воздухоносные пути Воздухоносные пути ветвятся, как дерево, разделяясь на несколько уровней (рис. 7-3), причем проксимальные отделы
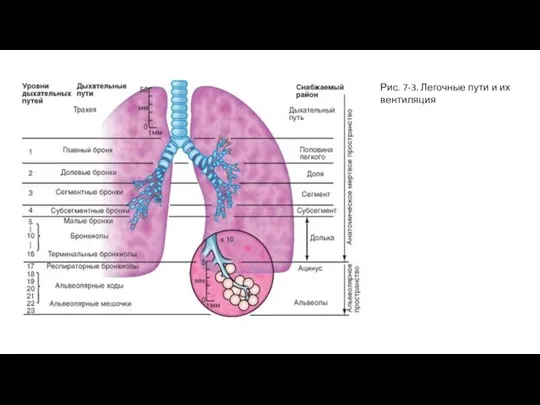
- 7. Рис. 7-3. Легочные пути и их вентиляция
- 8. Бронхиальное дерево Только около семи последних уровней разветвления бронхиального дерева, заканчивающихся дыхательными бронхиолами (bronchioli respiratorii) и
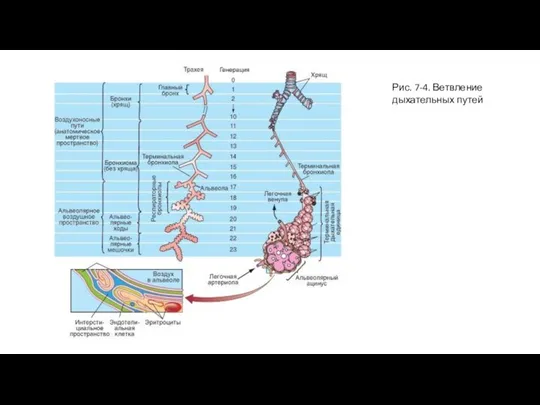
- 9. Рис. 7-4. Ветвление дыхательных путей
- 10. Защита организма от вредоносных компонентов вдыхаемого воздуха Слой ткани, который отделяет газы альвеолярного пространства от крови
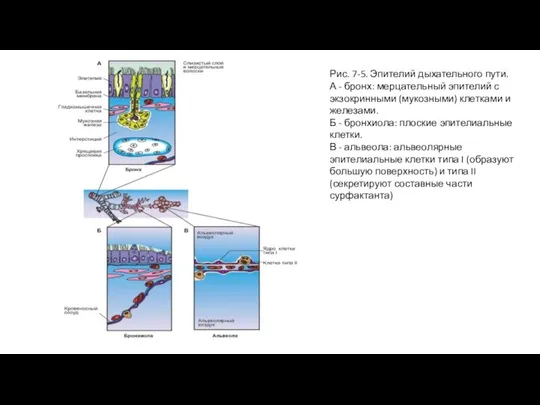
- 11. Рис. 7-5. Эпителий дыхательного пути. А - бронх: мерцательный эпителий с экзокринными (мукозными) клетками и железами.
- 12. Кровоснабжение дыхательных путей Ветви легочной артерии многократно разветвляются, причем они следуют разветвлениям воздухоносных дыхательных путей. Затем
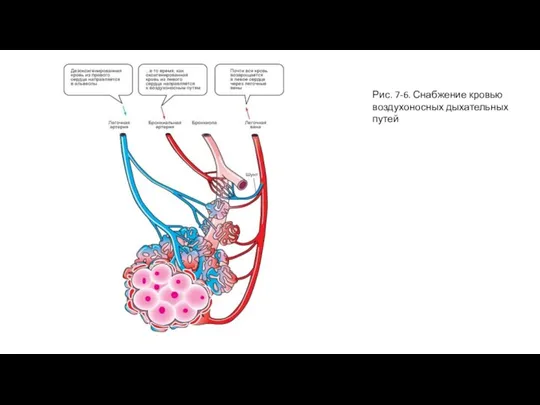
- 13. Рис. 7-6. Снабжение кровью воздухоносных дыхательных путей
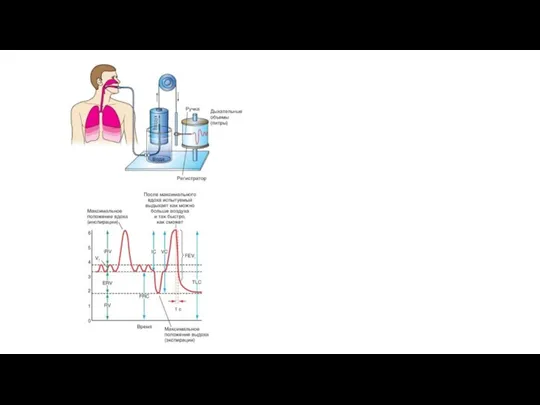
- 14. Спирография Спирометр (рис. 7-7) представляет собой прибор с ограниченным газовым пространством, из которого газ можно вдыхать
- 16. Плетизмография Общая плетизмография (body plethys mography) используется для исследования функций внешнего дыхания. Плетизмограф представляет собой герметичную
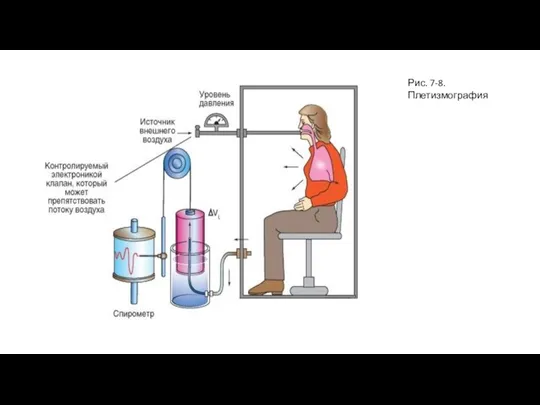
- 17. Рис. 7-8. Плетизмография
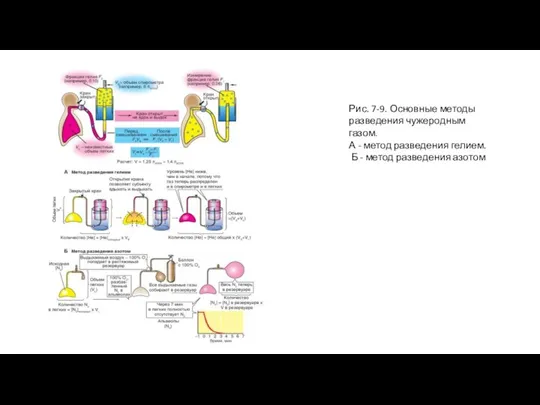
- 18. Определение общей и функциональной остаточных емкостей легких После максимального выдоха в легком остается еще около 1,5
- 19. Рис. 7-9. Основные методы разведения чужеродным газом. А - метод разведения гелием. Б - метод разведения
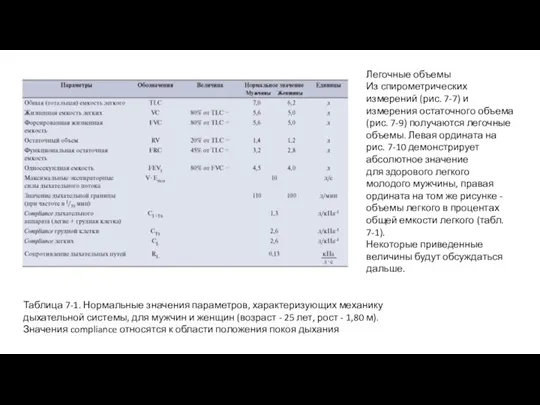
- 20. Легочные объемы Из спирометрических измерений (рис. 7-7) и измерения остаточного объема (рис. 7-9) получаются легочные объемы.
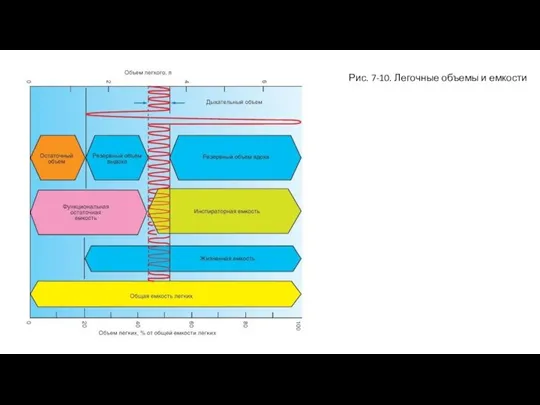
- 21. Рис. 7-10. Легочные объемы и емкости
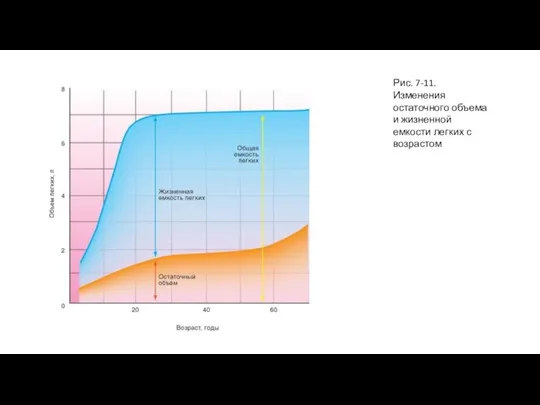
- 22. Возрастная динамика легочных объемов Периоду роста до его завершения соответствует увеличение жизненной емкости и остаточного объема
- 23. Рис. 7-11. Изменения остаточного объема и жизненной емкости легких с возрастом
- 24. Сompliance и elastance легких Если воздушный баллон, например воздушный шарик, постепенно заполняется воздухом, то внутри его
- 25. Рис. 7-12. Трансмуральная разность давления (Рtm). А - слева - внутреннее давление (Рi) повышается, в то
- 26. Кривые растяжения покоя Растяжимость легкого определяется его пассивными структурными элементами, т.е. изменяется не активно (например, с
- 27. Рис. 7-13. Кривые пассивного изменения давления от объема легкого, грудной клетки и дыхательного аппарата (кривые давление-объем).
- 28. Дыхательные мышцы Дыхательные мышцы являются двигателем вентиляции. При спокойном дыхании диафрагма выполняет функцию основной дыхательной мышцы,
- 29. Рис. 7-14. Работа дыхательных мышц. А - при движении вниз диафрагмы грудная клетка увеличивает свой объем.
- 30. Пневмоторакс При положении покоя дыхания альвеолярное давление (РА), которое представляет собой трансмуральную разность давления дыхательного аппарата,
- 31. Рис. 7-15. Пневмоторакс (коллапс легких)
- 32. Зависимость между давлением и объемом легких Эластические свойства присущи как отдельным элементам легких, так и легким
- 33. Рис. 7-16. Зависимость между давлением и объемом легких
- 34. Применение закона Лапласа к альвеолам Кривую гистерезиса и ее устранение при помощи заполнения легких водой трудно
- 35. Рис. 7-17. Закон Лапласа
- 36. Обнаружение сурфактанта В экстрактах легкого, в смывах и в жидкости, полученной из отечного легкого, удалось обнаружить
- 37. Рис. 7-18. Срез альвеолярной перегородки со скоплением сурфактанта, особенно в углах. Так устраняются сильные изгибы (маленькие
- 38. Сурфактант Существующее в легком поверхностное натяжение уменьшается при помощи пленки поверхностно активного вещества, названногосурфактантом, который действует
- 39. Рис. 7-19. Строение (А), механизм действия (Б) и метаболизм (В) сурфактанта
- 40. Эластическая тяга легких Легкое растянуто и стремиться сжаться, в то время как грудная клетка сжата и
- 41. Рис. 7-20. Эластическая тяга легких
- 42. Сопротивление воздухоносных путей Сопротивление воздухоносных путей локализуется в верхних дыхательных путях. Так как воздухоносные пути по
- 43. Рис. 7-21. Сопротивление потоку воздуха в дыхательном тракте. А - сопротивление потоку воздуха во всех (параллельно
- 44. Модель крепления альвеол Крепление альвеол друг другом является третьим фактором, определяющим эластичность легкого. Стремление к ретракции
- 45. Рис. 7-22. Ретракция легочной ткани. Эластичная альвеолярная ткань осуществляет растяжение интропульмональных бронхов и сосудов, которые моделируются
- 46. Измерение альвеолярного давления Мы вновь прибегаем к построению, причем на этот раз мы дополнительно измеряем с
- 47. Рис. 7-23. Объемная скорость вентиляции, альвеолярное давление и плевральное давление. При очень медленном дыхании (дыхательный объем
- 48. Типы потоков воздуха в дыхательных трубках Ламинарный поток (laminar flow) является таким типом движения, когда воздух
- 49. Рис. 7-24. Типы потоков воздуха в трубке. А - при ламинарном течении все мельчайшие частицы движутся
- 50. Изменение давлений во время дыхательного цикла Дыхательный аппарат упрощенно можно себе представить как два заключенных друг
- 51. Рис. 7-25. Связь альвеолярного (РА) и плеврального (Рр1) давлений во время дыхательного цикла. На рисунке введены
- 52. Изменение давлений в респираторной системе во время дыхательного цикла Во время вдоха инспираторные мышцы отклоняют дыхательный
- 53. Рис. 7-26. Дилятация и спадение воздухоносных путей при прохождении потока воздуха. Представлена схема, иллюстрирующая сдавливание дыхательных
- 54. Изменение давлений в респираторной системе во время форсированного выдоха Максимальный форсированный выдох или выдох, осуществленный с
- 55. Рис. 7-27. Форсированный выдох. А - отсутствие потока. Б - максимальный форсированный выдох. В - кривые
- 56. Альвеолярная вентиляция С каждым литром воздуха мы вдыхаем около 170 мл О2. Часть кислорода попадает в
- 57. Рис. 7-28. Поступление О2 в легкие и выделение СО2. А - балансы поступления О2 с дыханием
- 58. Структура и функции гемоглобина Кислород и углекислый газ частично переносятся в крови в физически растворенном виде.
- 59. Рис. 7-29. Гемоглобин. А - гемоглобиновый тетрамер. Б - молекула гена. В - конформационные изменения при
- 60. Чтобы лучше представить свойства реакции связывания О2 гемоглобином, принимают во внимание только концентрацию О2, связанного с
- 61. Рис. 7-31. Сатурационная кривая О2. Кривая связывания О2 крови (оранжевая кривая). Показана зависимость между насыщением гемоглобина
- 62. Изменение сродства гемоглобина к кислороду Ряд факторов влияет на аффинность гемоглобина к О2, т.е. на насыщение
- 63. Рис. 7-32. Факторы влияющие на аффинность гемоглобина к О2. Уменьшение аффинности означает, что при одинаковом РО2
- 64. Карбоксигемоглобин Способность к транспорту кислорода кровью нарушается, если гемоглобин не способен присоединять О2. При этом, практически
- 65. Рис. 7-33. Влияние угарного газа на связывание гемоглобином О2 крови. Если построить кривую, отражающую половину кислородной
- 66. Факторы, влияющие на связывание кислорода гемоглобином Напомним, что существует ряд факторов, которые влияют на аффинность гемоглобина
- 67. Рис. 7-34. Сдвиги сатурационной кривой О2 при ряде воздействий. А - эффекты респираторных нарушений, связанные с
- 68. Связывание углекислого газа По аналогии с кривой связывания О2, кривая связывания СО2 описывает связь между концентрацией
- 69. Рис. 7-35. Связывание СО2. А - кривая связывания СО2 в крови человека. Общая концентрация СО2 составляется
- 70. Обмен СО2 Рисунок 7-36 демонстрирует процессы, которые происходят в крови, когда в тканях образуется СО2, который
- 71. Рис. 7-36. Процессы обмена СО2 в большом круге кровообращения
- 72. Эффекты Бора и Холдейна Эффект Холдейна, основывается на одинаковом механизме с эффектом Бора, а именно на
- 73. Рис. 7-37. Принципы эффекта Бора (А) и эффекта Холдейна (Б). A - эффект Бора описывает влияние
- 74. Газообмен в большом круге кровообращения На рисунке представлен эритроцит, пришедший в большой круг кровообращения, плазма, в
- 75. Рис. 7-38. Обмен газов в большом круге кровообращения
- 76. Газообмен в малом круге кровообращения На рис. 7-39 представлен эритроцит, пришедший в малый круг кровообращения, плазма,
- 77. Диффузионная способность легких Между газовой средой в альвеолярном пространстве и кровью, протекающей в легочных капиллярах, находится
- 78. Рис. 7-40. Закон Фика и сложность его использования. А - изменение площади и толщины. Б -
- 79. Диффузия кислорода в легких На рис. 7-41 А продемонстрировано поступление О2 из альвеолярного воздуха в кровь
- 80. Рис. 7-41. Диффузия кислорода. На панели А показаны гипотетическая альвеола и капилляр. Продемонстрировано, как эритроциты входят
- 81. Диффузия углекислого газа в легких Углекислый газ диффундирует из легочных капилляров в альвеолы и выводится из
- 82. Рис. 7-42. Диффузия углекислого газа. На панели А показана гипотетическая альвеола и капилляр. Продемонстрировано, как эритроциты
- 83. Диффузия угарного газа (СО) в легких На рис. 7-43 представлена диффузия оксида углерода (II) (угарный газ,
- 84. Рис. 7-43. Диффузия монооксида углерода (угарного газа) из альвеол в кровь
- 85. Диффузия оксида азота (N2O) в легких Предположим, что в некой альвеоле содержится другой чужеродный газ, например
- 86. Рис. 7-44. Диффузия оксида азота (веселящего газа) из альвеол в кровь
- 87. Неравномерность вентиляционноперфузионного отношения в легких Под влиянием силы тяжести объемная скорость кровотока или (применительно к анатомическофункциональной
- 88. Рис. 7-45. Распределение вентиляции - перфузии в легком. Под действием силы тяжести увеличивается перфузия (Q слева)
- 89. Вентиляционно-перфузионное отношение Региональная неравномерность VA / Q означает региональные различия альвеолярных парциальных давлений. Альвеолярная вентиляция (VA)
- 90. Рис. 7-46. Отношение вентиляции и перфузии (VA/Q) альвеолы определяет ее газовый состав (показан только О2). Альвеолярная
- 91. Нормальное перфузионновентиляционное отношение в легких Как было показано на рис. 7-45 перфузия (Q) распределена во всех
- 92. Рис. 7-47. Нормальное распределение VA/Q . Приведенный на рисунке пример является идеализированным
- 93. Перфузионно-вентиляционное отношение в области мертвого пространства Рассмотрим случай, при котором вентиляционноперфузионное отношение постепенно увеличивается в результате
- 94. Рис. 7-48. Вентиляция области альвеолярного мертвого пространства
- 95. Перфузионно-вентиляционное отношение в области шунта Предположим теперь, что вентиляционно-перфузионное отношение постепенно уменьшается за счет снижения вентиляции
- 96. Рис. 7-49. Шунт
- 97. Регуляция дыхания Под регуляцией дыхания понимают способность легочной вентиляции приспосабливаться к потребностям обмена веществ организма и
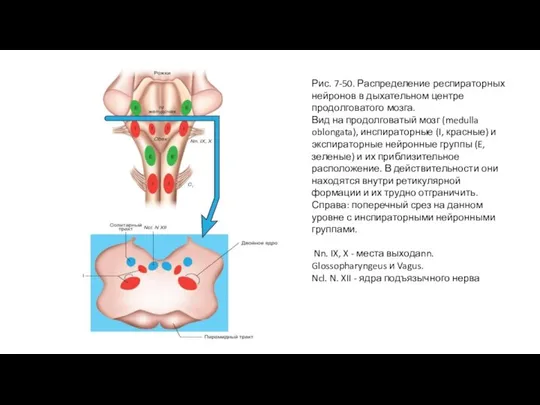
- 98. Рис. 7-50. Распределение респираторных нейронов в дыхательном центре продолговатого мозга. Вид на продолговатый мозг (medulla oblongata),
- 99. Регуляция дыхания под влиянием сигналов хеморецепторов Изменение дыхания под влиянием химических факторов осуществляется через хеморецепторы, которые,
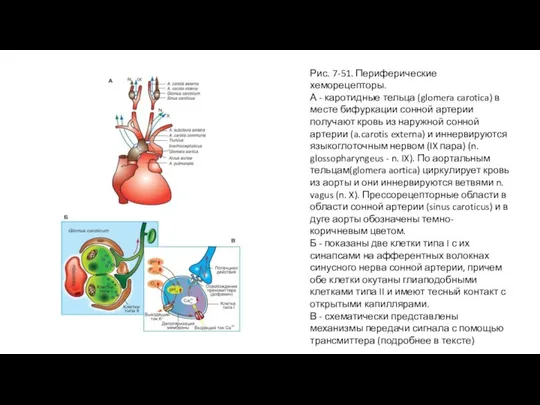
- 100. Рис. 7-51. Периферические хеморецепторы. А - каротидные тельца (glomera carotica) в месте бифуркации сонной артерии получают
- 101. Типы дыхания При нормальном дыхании дыхательный объем и частота дыхания могут значительно изменяться (рис. 7-52). При
- 103. Скачать презентацию
Слайд 2Основные этапы процесса дыхания
Клетки нашего организма получают энергию с использованием кислорода. Из-за
Основные этапы процесса дыхания
Клетки нашего организма получают энергию с использованием кислорода. Из-за

Слайд 3Рис. 7-1. Этапы процесса дыхания.
Обозначение.
Hb - гемоглобин
Рис. 7-1. Этапы процесса дыхания.
Обозначение.
Hb - гемоглобин
Слайд 4Основные физические характеристики компонентов газовой смеси
Важнейшими составляющими транспортной системы для внешнего дыхания
Основные физические характеристики компонентов газовой смеси
Важнейшими составляющими транспортной системы для внешнего дыхания

Слайд 5Рис. 7-2. Транспортная система дыхания, включающая систему внешнего дыхания, систему кровообращения и
Рис. 7-2. Транспортная система дыхания, включающая систему внешнего дыхания, систему кровообращения и
Слайд 6Воздухоносные пути
Воздухоносные пути ветвятся, как дерево, разделяясь на несколько уровней (рис. 7-3),
Воздухоносные пути
Воздухоносные пути ветвятся, как дерево, разделяясь на несколько уровней (рис. 7-3),

Слайд 7Рис. 7-3. Легочные пути и их вентиляция
Рис. 7-3. Легочные пути и их вентиляция
Слайд 8Бронхиальное дерево
Только около семи последних уровней разветвления бронхиального дерева, заканчивающихся дыхательными бронхиолами
Бронхиальное дерево
Только около семи последних уровней разветвления бронхиального дерева, заканчивающихся дыхательными бронхиолами

Слайд 9Рис. 7-4. Ветвление дыхательных путей
Рис. 7-4. Ветвление дыхательных путей
Слайд 10Защита организма от вредоносных компонентов вдыхаемого воздуха
Слой ткани, который отделяет газы альвеолярного
Защита организма от вредоносных компонентов вдыхаемого воздуха
Слой ткани, который отделяет газы альвеолярного

Слайд 11Рис. 7-5. Эпителий дыхательного пути.
А - бронх: мерцательный эпителий с экзокринными (мукозными)
Рис. 7-5. Эпителий дыхательного пути.
А - бронх: мерцательный эпителий с экзокринными (мукозными)
Слайд 12Кровоснабжение дыхательных путей
Ветви легочной артерии многократно разветвляются, причем они следуют разветвлениям воздухоносных
Кровоснабжение дыхательных путей
Ветви легочной артерии многократно разветвляются, причем они следуют разветвлениям воздухоносных

Слайд 13Рис. 7-6. Снабжение кровью воздухоносных дыхательных путей
Рис. 7-6. Снабжение кровью воздухоносных дыхательных путей
Слайд 14Спирография
Спирометр (рис. 7-7) представляет собой прибор с ограниченным газовым пространством, из которого
Спирография
Спирометр (рис. 7-7) представляет собой прибор с ограниченным газовым пространством, из которого

Слайд 16Плетизмография
Общая плетизмография (body plethys mography) используется для исследования функций внешнего дыхания. Плетизмограф
Плетизмография
Общая плетизмография (body plethys mography) используется для исследования функций внешнего дыхания. Плетизмограф

Слайд 17Рис. 7-8. Плетизмография
Рис. 7-8. Плетизмография
Слайд 18Определение общей и функциональной остаточных емкостей легких
После максимального выдоха в легком остается
Определение общей и функциональной остаточных емкостей легких
После максимального выдоха в легком остается

Слайд 19Рис. 7-9. Основные методы разведения чужеродным газом.
А - метод разведения гелием.
Б
Рис. 7-9. Основные методы разведения чужеродным газом.
А - метод разведения гелием.
Б
Слайд 20Легочные объемы
Из спирометрических измерений (рис. 7-7) и измерения остаточного объема (рис. 7-9)
Легочные объемы
Из спирометрических измерений (рис. 7-7) и измерения остаточного объема (рис. 7-9)
Слайд 21Рис. 7-10. Легочные объемы и емкости
Рис. 7-10. Легочные объемы и емкости
Слайд 22Возрастная динамика легочных объемов
Периоду роста до его завершения соответствует увеличение жизненной емкости
Возрастная динамика легочных объемов
Периоду роста до его завершения соответствует увеличение жизненной емкости

Слайд 23Рис. 7-11. Изменения остаточного объема и жизненной емкости легких с возрастом
Рис. 7-11. Изменения остаточного объема и жизненной емкости легких с возрастом
Слайд 24Сompliance и elastance легких
Если воздушный баллон, например воздушный шарик, постепенно заполняется воздухом,
Сompliance и elastance легких
Если воздушный баллон, например воздушный шарик, постепенно заполняется воздухом,

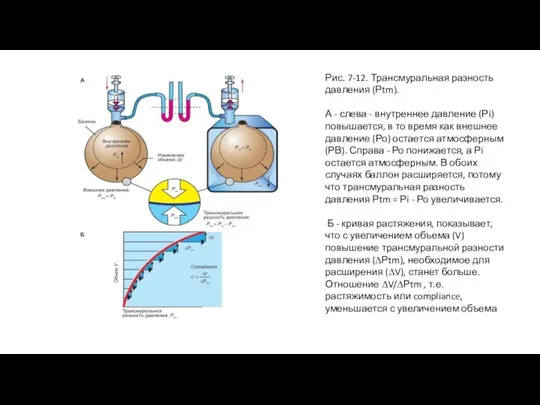
Слайд 25Рис. 7-12. Трансмуральная разность давления (Рtm).
А - слева - внутреннее давление (Рi)
Рис. 7-12. Трансмуральная разность давления (Рtm).
А - слева - внутреннее давление (Рi)
Слайд 26Кривые растяжения покоя
Растяжимость легкого определяется его пассивными структурными элементами, т.е. изменяется не
Кривые растяжения покоя
Растяжимость легкого определяется его пассивными структурными элементами, т.е. изменяется не

Слайд 27Рис. 7-13. Кривые пассивного изменения давления от объема легкого, грудной клетки и
Рис. 7-13. Кривые пассивного изменения давления от объема легкого, грудной клетки и

Слайд 28Дыхательные мышцы
Дыхательные мышцы являются двигателем вентиляции. При спокойном дыхании диафрагма выполняет функцию
Дыхательные мышцы
Дыхательные мышцы являются двигателем вентиляции. При спокойном дыхании диафрагма выполняет функцию

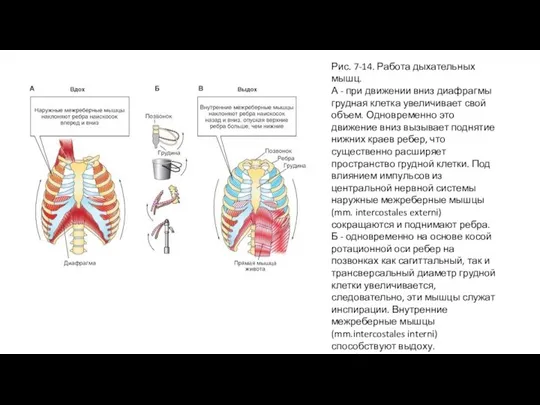
Слайд 29Рис. 7-14. Работа дыхательных мышц.
А - при движении вниз диафрагмы грудная клетка
Рис. 7-14. Работа дыхательных мышц.
А - при движении вниз диафрагмы грудная клетка
Слайд 30Пневмоторакс
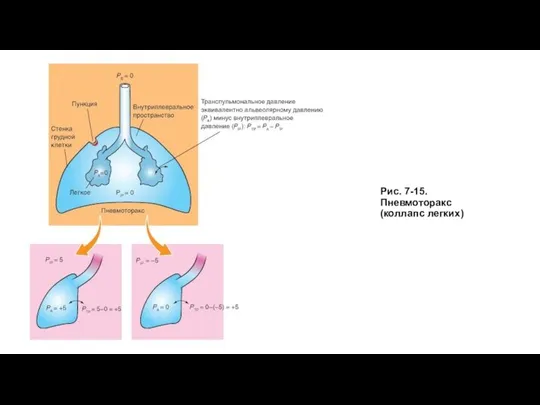
При положении покоя дыхания альвеолярное давление (РА), которое представляет собой трансмуральную разность
Пневмоторакс
При положении покоя дыхания альвеолярное давление (РА), которое представляет собой трансмуральную разность

Слайд 31Рис. 7-15. Пневмоторакс (коллапс легких)
Рис. 7-15. Пневмоторакс (коллапс легких)
Слайд 32Зависимость между давлением и объемом легких
Эластические свойства присущи как отдельным элементам легких,
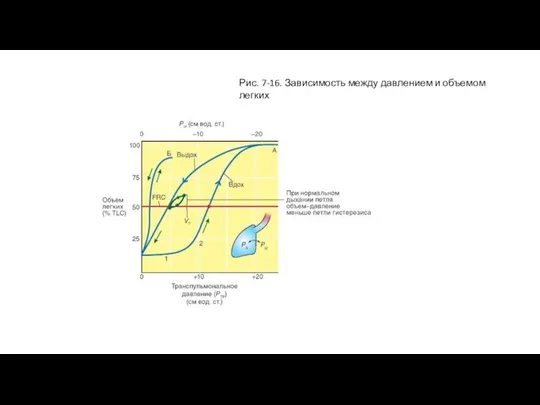
Зависимость между давлением и объемом легких
Эластические свойства присущи как отдельным элементам легких,

Слайд 33Рис. 7-16. Зависимость между давлением и объемом легких
Рис. 7-16. Зависимость между давлением и объемом легких
Слайд 34Применение закона Лапласа к альвеолам
Кривую гистерезиса и ее устранение при помощи заполнения
Применение закона Лапласа к альвеолам
Кривую гистерезиса и ее устранение при помощи заполнения

Слайд 35Рис. 7-17. Закон Лапласа
Рис. 7-17. Закон Лапласа
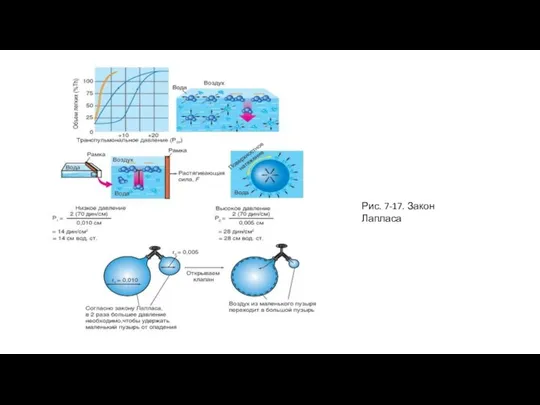
Слайд 36Обнаружение сурфактанта
В экстрактах легкого, в смывах и в жидкости, полученной из отечного
Обнаружение сурфактанта
В экстрактах легкого, в смывах и в жидкости, полученной из отечного

Слайд 37Рис. 7-18. Срез альвеолярной перегородки со скоплением сурфактанта, особенно в углах. Так
Рис. 7-18. Срез альвеолярной перегородки со скоплением сурфактанта, особенно в углах. Так
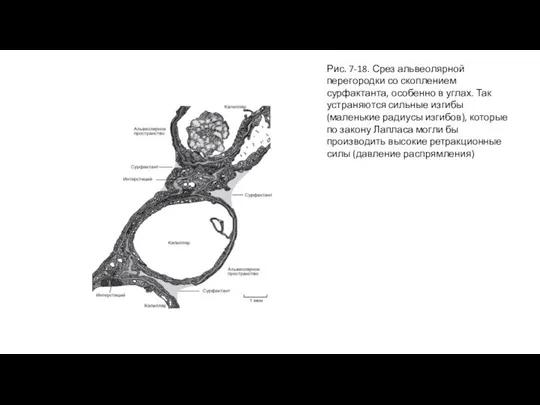
Слайд 38Сурфактант
Существующее в легком поверхностное натяжение уменьшается при помощи пленки поверхностно активного вещества,
Сурфактант
Существующее в легком поверхностное натяжение уменьшается при помощи пленки поверхностно активного вещества,

Слайд 39Рис. 7-19. Строение (А), механизм действия (Б) и метаболизм
(В) сурфактанта
Рис. 7-19. Строение (А), механизм действия (Б) и метаболизм
(В) сурфактанта
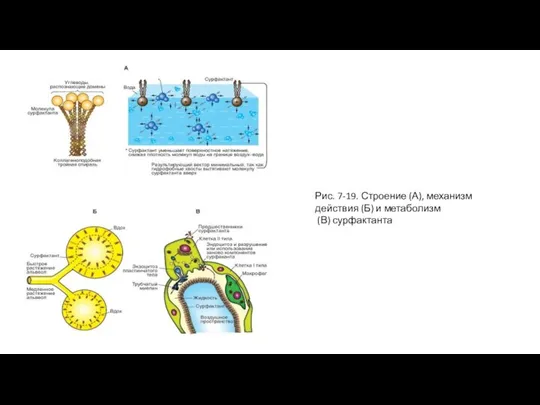
Слайд 40Эластическая тяга легких
Легкое растянуто и стремиться сжаться, в то время как грудная
Эластическая тяга легких
Легкое растянуто и стремиться сжаться, в то время как грудная

Слайд 41Рис. 7-20. Эластическая тяга легких
Рис. 7-20. Эластическая тяга легких
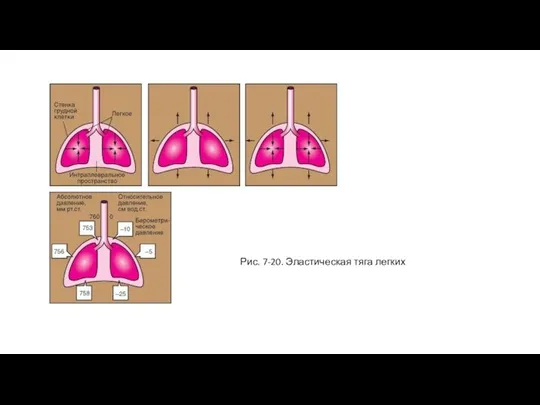
Слайд 42Сопротивление воздухоносных путей
Сопротивление воздухоносных путей локализуется в верхних дыхательных путях. Так как
Сопротивление воздухоносных путей
Сопротивление воздухоносных путей локализуется в верхних дыхательных путях. Так как

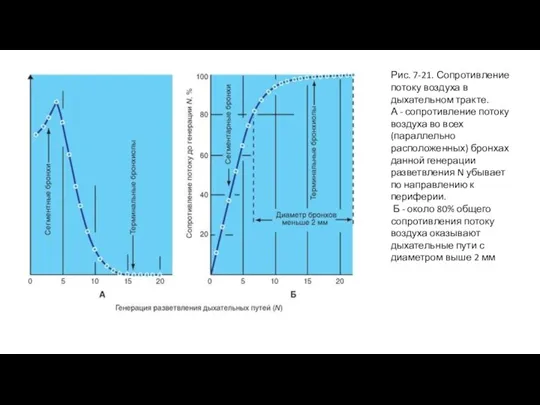
Слайд 43Рис. 7-21. Сопротивление потоку воздуха в дыхательном тракте.
А - сопротивление потоку воздуха
Рис. 7-21. Сопротивление потоку воздуха в дыхательном тракте.
А - сопротивление потоку воздуха
Слайд 44Модель крепления альвеол
Крепление альвеол друг другом является третьим фактором, определяющим эластичность легкого.
Модель крепления альвеол
Крепление альвеол друг другом является третьим фактором, определяющим эластичность легкого.

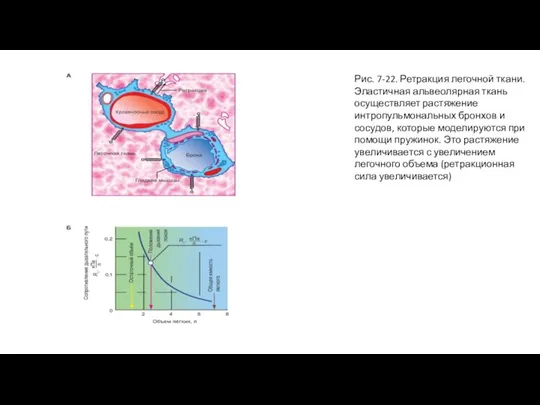
Слайд 45Рис. 7-22. Ретракция легочной ткани. Эластичная альвеолярная ткань осуществляет растяжение интропульмональных бронхов
Рис. 7-22. Ретракция легочной ткани. Эластичная альвеолярная ткань осуществляет растяжение интропульмональных бронхов
Слайд 46Измерение альвеолярного давления
Мы вновь прибегаем к построению, причем на этот раз мы
Измерение альвеолярного давления
Мы вновь прибегаем к построению, причем на этот раз мы

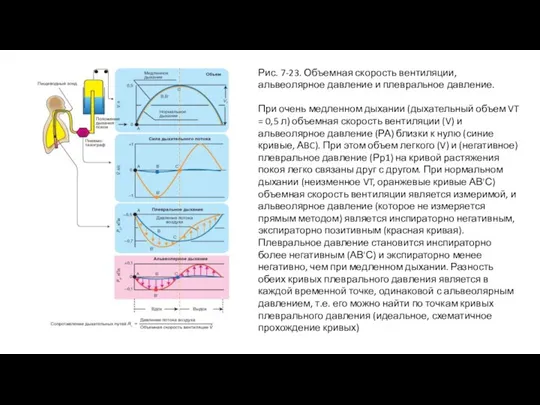
Слайд 47Рис. 7-23. Объемная скорость вентиляции, альвеолярное давление и плевральное давление.
При очень медленном
Рис. 7-23. Объемная скорость вентиляции, альвеолярное давление и плевральное давление.
При очень медленном
Слайд 48Типы потоков воздуха в дыхательных трубках
Ламинарный поток (laminar flow) является таким типом
Типы потоков воздуха в дыхательных трубках
Ламинарный поток (laminar flow) является таким типом

Слайд 49Рис. 7-24. Типы потоков воздуха в трубке.
А - при ламинарном течении все
Рис. 7-24. Типы потоков воздуха в трубке.
А - при ламинарном течении все

Слайд 50Изменение давлений во время дыхательного цикла
Дыхательный аппарат упрощенно можно себе представить как
Изменение давлений во время дыхательного цикла
Дыхательный аппарат упрощенно можно себе представить как

Слайд 51Рис. 7-25. Связь альвеолярного (РА) и плеврального (Рр1) давлений во время дыхательного
Рис. 7-25. Связь альвеолярного (РА) и плеврального (Рр1) давлений во время дыхательного

Слайд 52Изменение давлений в респираторной системе во время дыхательного цикла
Во время вдоха инспираторные
Изменение давлений в респираторной системе во время дыхательного цикла
Во время вдоха инспираторные

Слайд 53Рис. 7-26. Дилятация и спадение воздухоносных путей при прохождении потока воздуха.
Представлена схема,
Рис. 7-26. Дилятация и спадение воздухоносных путей при прохождении потока воздуха.
Представлена схема,

Слайд 54Изменение давлений в респираторной системе во время форсированного выдоха
Максимальный форсированный выдох или
Изменение давлений в респираторной системе во время форсированного выдоха
Максимальный форсированный выдох или

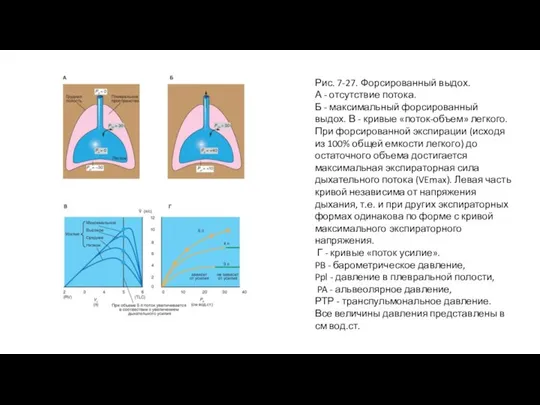
Слайд 55Рис. 7-27. Форсированный выдох.
А - отсутствие потока.
Б - максимальный форсированный выдох.
Рис. 7-27. Форсированный выдох.
А - отсутствие потока.
Б - максимальный форсированный выдох.
Слайд 56Альвеолярная вентиляция
С каждым литром воздуха мы вдыхаем около 170 мл О2. Часть
Альвеолярная вентиляция
С каждым литром воздуха мы вдыхаем около 170 мл О2. Часть

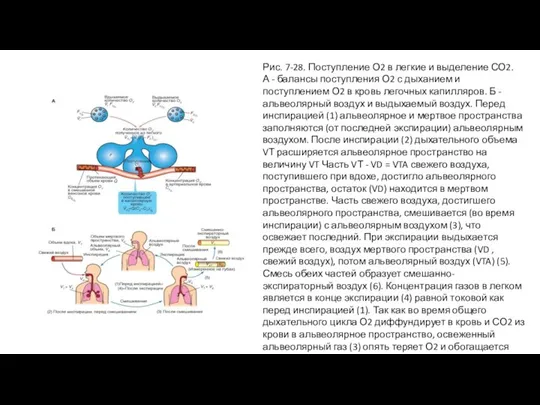
Слайд 57Рис. 7-28. Поступление О2 в легкие и выделение СО2.
А - балансы поступления
Рис. 7-28. Поступление О2 в легкие и выделение СО2.
А - балансы поступления
Слайд 58Структура и функции гемоглобина
Кислород и углекислый газ частично переносятся в крови в
Структура и функции гемоглобина
Кислород и углекислый газ частично переносятся в крови в

Слайд 59Рис. 7-29. Гемоглобин.
А - гемоглобиновый тетрамер.
Б - молекула гена.
В -
Рис. 7-29. Гемоглобин.
А - гемоглобиновый тетрамер.
Б - молекула гена.
В -

Слайд 60Чтобы лучше представить свойства реакции связывания О2 гемоглобином, принимают во внимание только
Чтобы лучше представить свойства реакции связывания О2 гемоглобином, принимают во внимание только

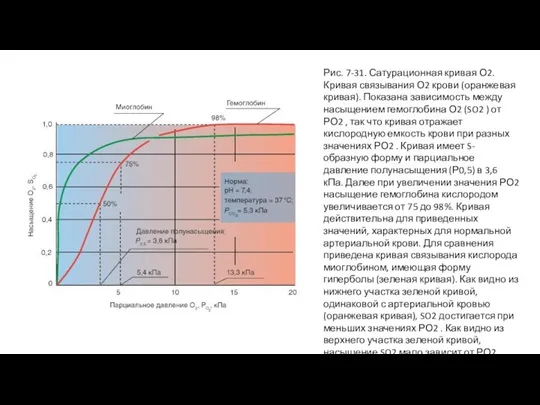
Слайд 61Рис. 7-31. Сатурационная кривая О2.
Кривая связывания О2 крови (оранжевая кривая). Показана зависимость
Рис. 7-31. Сатурационная кривая О2.
Кривая связывания О2 крови (оранжевая кривая). Показана зависимость
Слайд 62Изменение сродства гемоглобина к кислороду
Ряд факторов влияет на аффинность гемоглобина к О2,
Изменение сродства гемоглобина к кислороду
Ряд факторов влияет на аффинность гемоглобина к О2,

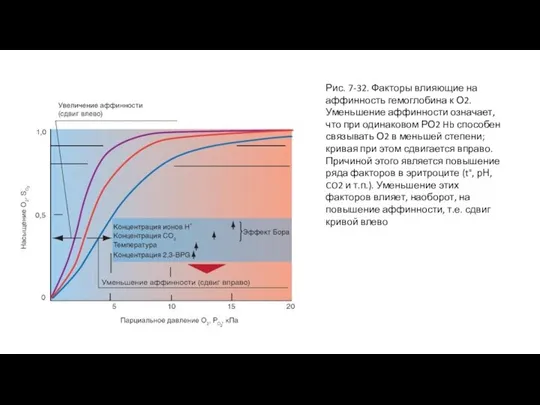
Слайд 63Рис. 7-32. Факторы влияющие на аффинность гемоглобина к О2.
Уменьшение аффинности означает, что
Рис. 7-32. Факторы влияющие на аффинность гемоглобина к О2.
Уменьшение аффинности означает, что
Слайд 64Карбоксигемоглобин
Способность к транспорту кислорода кровью нарушается, если гемоглобин не способен присоединять О2.
Карбоксигемоглобин
Способность к транспорту кислорода кровью нарушается, если гемоглобин не способен присоединять О2.

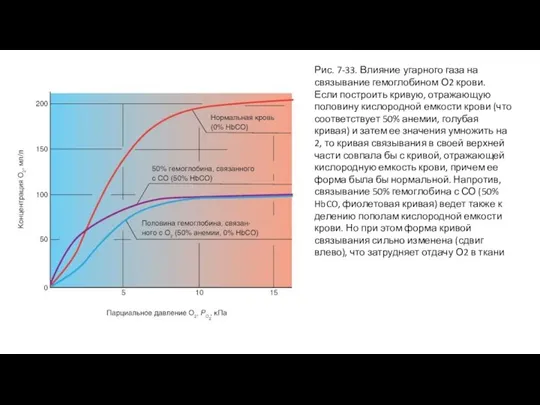
Слайд 65Рис. 7-33. Влияние угарного газа на связывание гемоглобином О2 крови.
Если построить кривую,
Рис. 7-33. Влияние угарного газа на связывание гемоглобином О2 крови.
Если построить кривую,
Слайд 66Факторы, влияющие на связывание кислорода гемоглобином
Напомним, что существует ряд факторов, которые влияют
Факторы, влияющие на связывание кислорода гемоглобином
Напомним, что существует ряд факторов, которые влияют

Слайд 67Рис. 7-34. Сдвиги сатурационной кривой О2 при ряде воздействий.
А - эффекты респираторных
Рис. 7-34. Сдвиги сатурационной кривой О2 при ряде воздействий.
А - эффекты респираторных

Слайд 68Связывание углекислого газа
По аналогии с кривой связывания О2, кривая связывания СО2 описывает
Связывание углекислого газа
По аналогии с кривой связывания О2, кривая связывания СО2 описывает

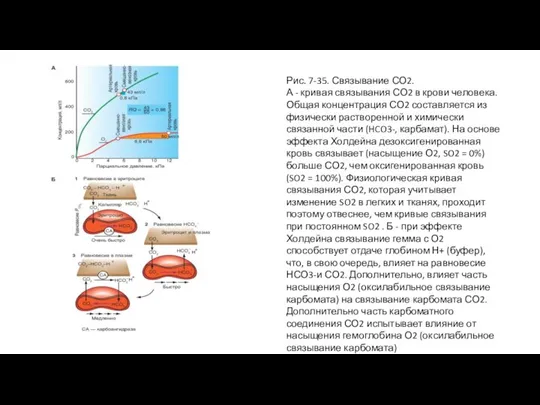
Слайд 69Рис. 7-35. Связывание СО2.
А - кривая связывания СО2 в крови человека. Общая
Рис. 7-35. Связывание СО2.
А - кривая связывания СО2 в крови человека. Общая
Слайд 70Обмен СО2
Рисунок 7-36 демонстрирует процессы, которые происходят в крови, когда в тканях
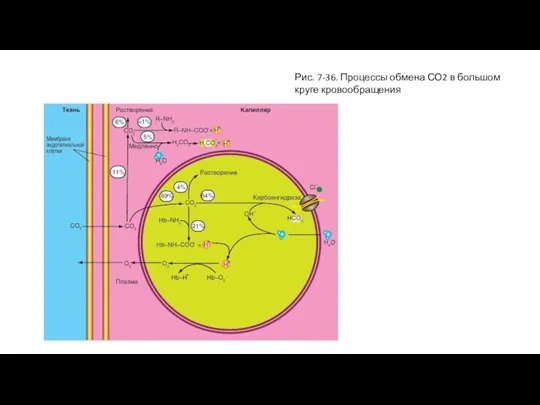
Обмен СО2
Рисунок 7-36 демонстрирует процессы, которые происходят в крови, когда в тканях

Слайд 71Рис. 7-36. Процессы обмена СО2 в большом круге кровообращения
Рис. 7-36. Процессы обмена СО2 в большом круге кровообращения
Слайд 72Эффекты Бора и Холдейна
Эффект Холдейна, основывается на одинаковом механизме с эффектом Бора,
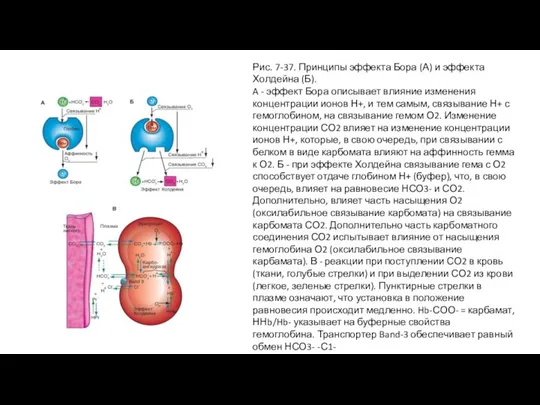
Эффекты Бора и Холдейна
Эффект Холдейна, основывается на одинаковом механизме с эффектом Бора,

Слайд 73Рис. 7-37. Принципы эффекта Бора (А) и эффекта Холдейна (Б).
A - эффект
Рис. 7-37. Принципы эффекта Бора (А) и эффекта Холдейна (Б).
A - эффект
Слайд 74Газообмен в большом круге кровообращения
На рисунке представлен эритроцит, пришедший в большой круг
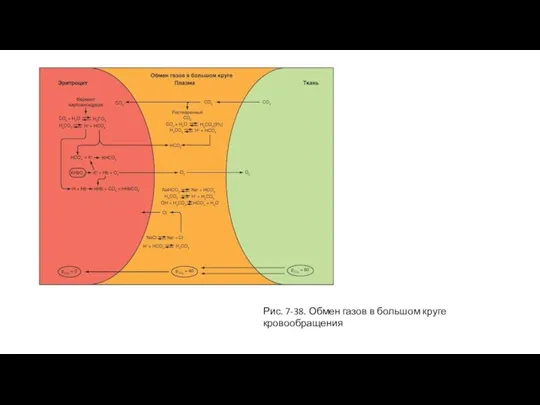
Газообмен в большом круге кровообращения
На рисунке представлен эритроцит, пришедший в большой круг

Слайд 75Рис. 7-38. Обмен газов в большом круге кровообращения
Рис. 7-38. Обмен газов в большом круге кровообращения
Слайд 76Газообмен в малом круге кровообращения
На рис. 7-39 представлен эритроцит, пришедший в малый
Газообмен в малом круге кровообращения
На рис. 7-39 представлен эритроцит, пришедший в малый

Слайд 77Диффузионная способность легких
Между газовой средой в альвеолярном пространстве и кровью, протекающей в
Диффузионная способность легких
Между газовой средой в альвеолярном пространстве и кровью, протекающей в

Слайд 78Рис. 7-40. Закон Фика и сложность его использования.
А - изменение площади и
Рис. 7-40. Закон Фика и сложность его использования.
А - изменение площади и
Слайд 79Диффузия кислорода в легких
На рис. 7-41 А продемонстрировано поступление О2 из альвеолярного
Диффузия кислорода в легких
На рис. 7-41 А продемонстрировано поступление О2 из альвеолярного

Слайд 80Рис. 7-41. Диффузия кислорода.
На панели А показаны гипотетическая альвеола и капилляр. Продемонстрировано,
Рис. 7-41. Диффузия кислорода.
На панели А показаны гипотетическая альвеола и капилляр. Продемонстрировано,

Слайд 81Диффузия углекислого газа в легких
Углекислый газ диффундирует из легочных капилляров в альвеолы
Диффузия углекислого газа в легких
Углекислый газ диффундирует из легочных капилляров в альвеолы

Слайд 82Рис. 7-42. Диффузия углекислого газа.
На панели А показана гипотетическая альвеола и капилляр.
Рис. 7-42. Диффузия углекислого газа.
На панели А показана гипотетическая альвеола и капилляр.

Слайд 83Диффузия угарного газа (СО) в легких
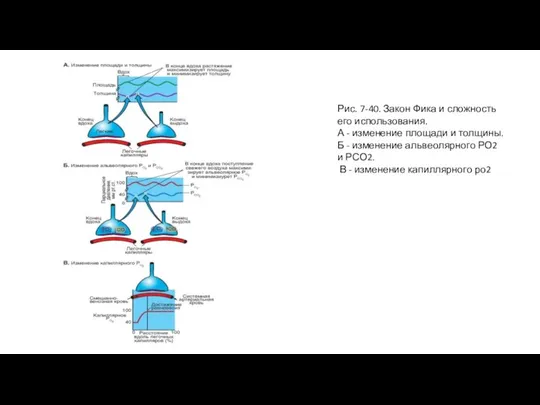
На рис. 7-43 представлена диффузия оксида углерода
Диффузия угарного газа (СО) в легких
На рис. 7-43 представлена диффузия оксида углерода

Слайд 84Рис. 7-43. Диффузия монооксида углерода (угарного газа) из альвеол в кровь
Рис. 7-43. Диффузия монооксида углерода (угарного газа) из альвеол в кровь
Слайд 85Диффузия оксида азота (N2O) в легких
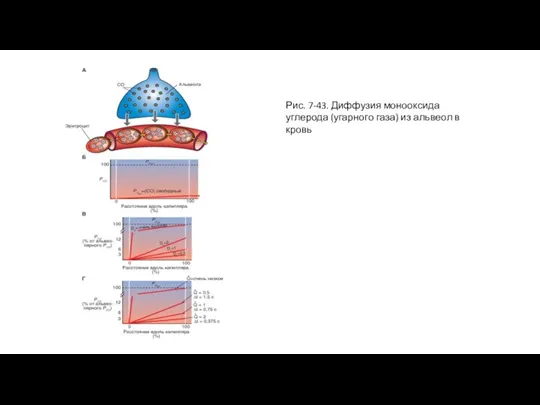
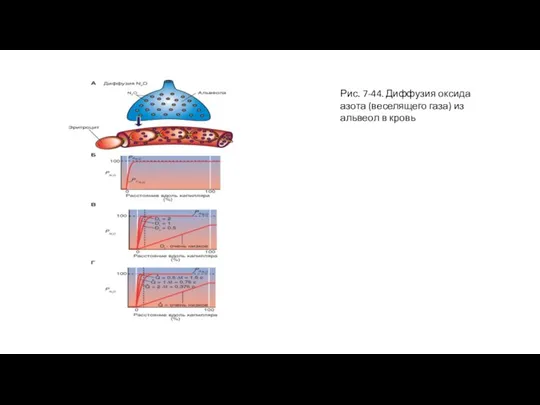
Предположим, что в некой альвеоле содержится другой
Диффузия оксида азота (N2O) в легких
Предположим, что в некой альвеоле содержится другой

Слайд 86Рис. 7-44. Диффузия оксида азота (веселящего газа) из альвеол в кровь
Рис. 7-44. Диффузия оксида азота (веселящего газа) из альвеол в кровь
Слайд 87Неравномерность вентиляционноперфузионного отношения в легких
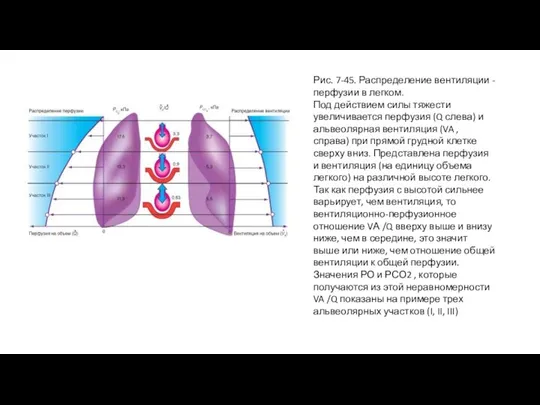
Под влиянием силы тяжести объемная скорость кровотока или
Неравномерность вентиляционноперфузионного отношения в легких
Под влиянием силы тяжести объемная скорость кровотока или

Слайд 88Рис. 7-45. Распределение вентиляции - перфузии в легком.
Под действием силы тяжести увеличивается
Рис. 7-45. Распределение вентиляции - перфузии в легком.
Под действием силы тяжести увеличивается
Слайд 89Вентиляционно-перфузионное отношение
Региональная неравномерность VA / Q означает региональные различия альвеолярных парциальных давлений.
Вентиляционно-перфузионное отношение
Региональная неравномерность VA / Q означает региональные различия альвеолярных парциальных давлений.

Слайд 90Рис. 7-46. Отношение вентиляции и перфузии (VA/Q) альвеолы определяет ее газовый состав
Рис. 7-46. Отношение вентиляции и перфузии (VA/Q) альвеолы определяет ее газовый состав
Слайд 91Нормальное перфузионновентиляционное отношение в легких
Как было показано на рис. 7-45 перфузия (Q)
Нормальное перфузионновентиляционное отношение в легких
Как было показано на рис. 7-45 перфузия (Q)

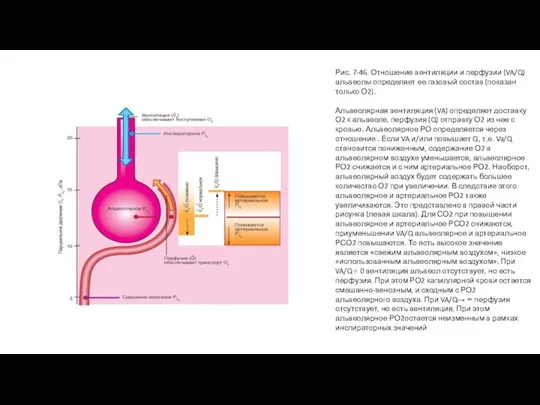
Слайд 92Рис. 7-47. Нормальное распределение VA/Q . Приведенный на рисунке пример является идеализированным
Рис. 7-47. Нормальное распределение VA/Q . Приведенный на рисунке пример является идеализированным
Слайд 93Перфузионно-вентиляционное отношение в области мертвого пространства
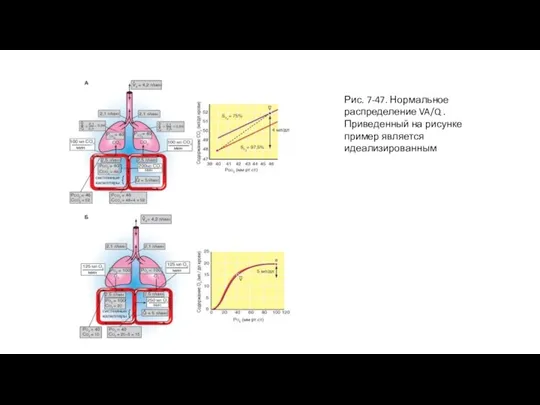
Рассмотрим случай, при котором вентиляционноперфузионное отношение постепенно
Перфузионно-вентиляционное отношение в области мертвого пространства
Рассмотрим случай, при котором вентиляционноперфузионное отношение постепенно

Слайд 94Рис. 7-48. Вентиляция области альвеолярного мертвого пространства
Рис. 7-48. Вентиляция области альвеолярного мертвого пространства
Слайд 95Перфузионно-вентиляционное отношение в области шунта
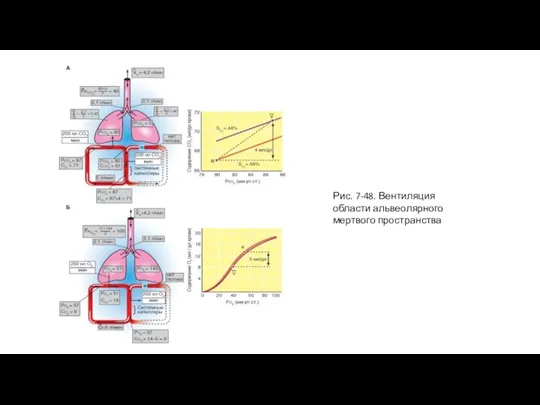
Предположим теперь, что вентиляционно-перфузионное отношение постепенно уменьшается за
Перфузионно-вентиляционное отношение в области шунта
Предположим теперь, что вентиляционно-перфузионное отношение постепенно уменьшается за

Слайд 96Рис. 7-49. Шунт
Рис. 7-49. Шунт
Слайд 97Регуляция дыхания
Под регуляцией дыхания понимают способность легочной вентиляции приспосабливаться к потребностям обмена
Регуляция дыхания
Под регуляцией дыхания понимают способность легочной вентиляции приспосабливаться к потребностям обмена

Слайд 98Рис. 7-50. Распределение респираторных нейронов в дыхательном центре продолговатого мозга.
Вид на продолговатый
Рис. 7-50. Распределение респираторных нейронов в дыхательном центре продолговатого мозга.
Вид на продолговатый
Слайд 99Регуляция дыхания под влиянием сигналов хеморецепторов
Изменение дыхания под влиянием химических факторов осуществляется
Регуляция дыхания под влиянием сигналов хеморецепторов
Изменение дыхания под влиянием химических факторов осуществляется

Слайд 100Рис. 7-51. Периферические хеморецепторы.
А - каротидные тельца (glomera carotica) в месте бифуркации
Рис. 7-51. Периферические хеморецепторы.
А - каротидные тельца (glomera carotica) в месте бифуркации
Слайд 101Типы дыхания
При нормальном дыхании дыхательный объем и частота дыхания могут значительно изменяться
Типы дыхания
При нормальном дыхании дыхательный объем и частота дыхания могут значительно изменяться

 Эмбриональное развитие. Урок № 17
Эмбриональное развитие. Урок № 17 Презентация на тему ИВАН ПЕТРОВИЧ ПАВЛОВ
Презентация на тему ИВАН ПЕТРОВИЧ ПАВЛОВ  Молекулярная биология
Молекулярная биология Решение заданий части С. Цитологические задачи (С5)
Решение заданий части С. Цитологические задачи (С5) Собака - наш четвероногий друг
Собака - наш четвероногий друг Современные биотехнологии для восстановления и поддержания формы спортсменов во время тренировок и соревнований
Современные биотехнологии для восстановления и поддержания формы спортсменов во время тренировок и соревнований Деревья
Деревья Современные концепции биологической эволюции человека
Современные концепции биологической эволюции человека Значение насекомых для человека
Значение насекомых для человека Цветочная посуда и инструменты
Цветочная посуда и инструменты Викторина по биологии
Викторина по биологии Характеристика животных организмов с позиции токсичности
Характеристика животных организмов с позиции токсичности Нейрофизиология
Нейрофизиология 0883b59bade141dc824d9745db128548
0883b59bade141dc824d9745db128548 Голосеменные растения
Голосеменные растения Строение нервной системы
Строение нервной системы Пернатые друзья. Интересные факты о птицах
Пернатые друзья. Интересные факты о птицах Отдел Голосеменные, особенности строения
Отдел Голосеменные, особенности строения Все выше, выше и выше
Все выше, выше и выше Заповедники Ростовской области
Заповедники Ростовской области Медуница лекарственная
Медуница лекарственная 20171026_bakterii
20171026_bakterii Типы соединения костей. Опора и движение урок 4
Типы соединения костей. Опора и движение урок 4 Интересные растения и плоды
Интересные растения и плоды Хордовые: строение, происхождение и родственные связи
Хордовые: строение, происхождение и родственные связи Яичник. Строение
Яичник. Строение Фотосинтез
Фотосинтез Дикие животные и их детёныши
Дикие животные и их детёныши