- Физиология нейронов и глии

Содержание
- 2. 2. Эфферентные нейроны передают информацию от ЦНС к рабочим органам. Мультиполярные. Тела в сером веществе ЦНС
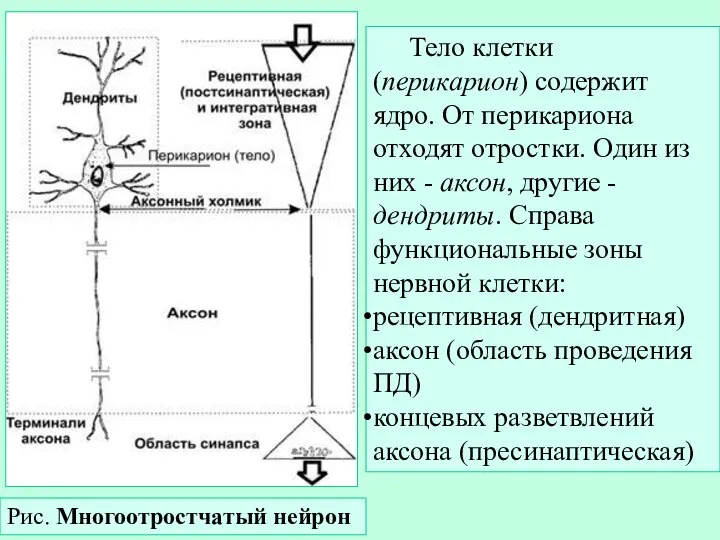
- 3. Рис. Многоотростчатый нейрон Тело клетки (перикарион) содержит ядро. От перикариона отходят отростки. Один из них -
- 4. Нейрон и его компоненты А - нервная клетка, аксон, мышца; Б - строение нервного волокна
- 5. II. Функции нейрона: прием, кодирование, обработка, хранение и передача информации. Нейроны формируют управляющие команды для внутренних
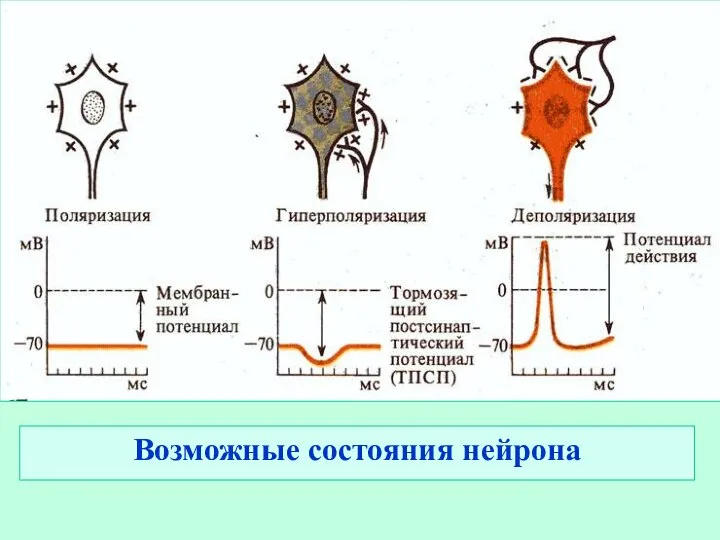
- 6. Три состояния нейрона: покой, активность и торможение. Покой - стабильный уровень мембранного потенциала и в любой
- 7. Возможные состояния нейрона
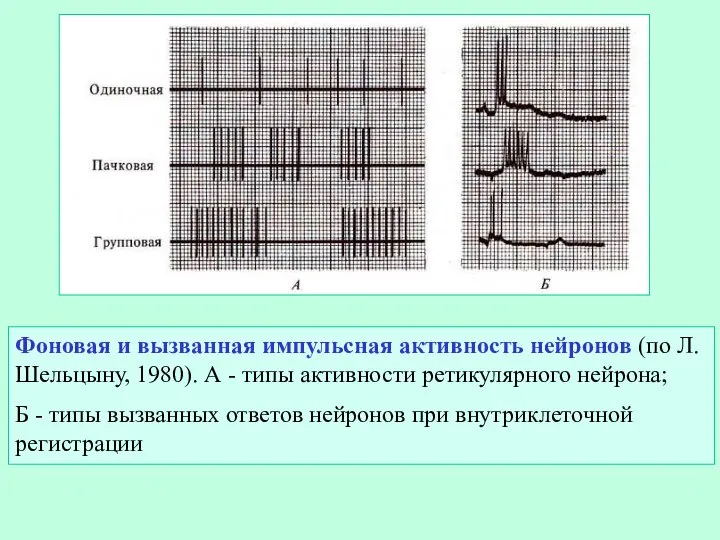
- 8. Фоновая и вызванная импульсная активность нейронов (по Л. Шельцыну, 1980). А - типы активности ретикулярного нейрона;
- 9. III. Нейроглия (от гр. neuron – нерв, и glia – клей) – это разнородная группа клеток
- 10. Олигодендроциты окружают тела нейронов, входят в состав нервных волокон и нервных окончаний и, благодаря выработке миелина,
- 11. IV. Нервные волокна Главная функция нервных волокон - проведение нервных импульсов. Механизм распространения нервного импульса -
- 12. Проведение возбуждения в нервных волокнах (по Дж. Бендоллу, 1970): I - немиелинизированное волокно II - миелинизированное
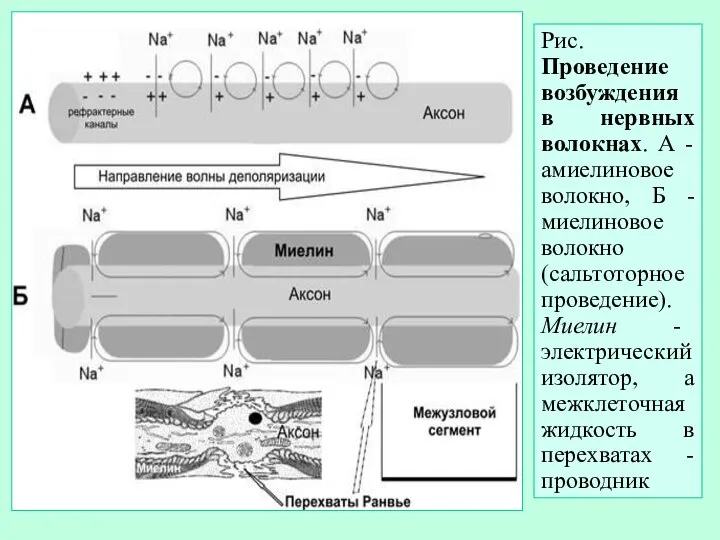
- 13. Рис. Проведение возбуждения в нервных волокнах. А - амиелиновое волокно, Б - миелиновое волокно (сальтоторное проведение).
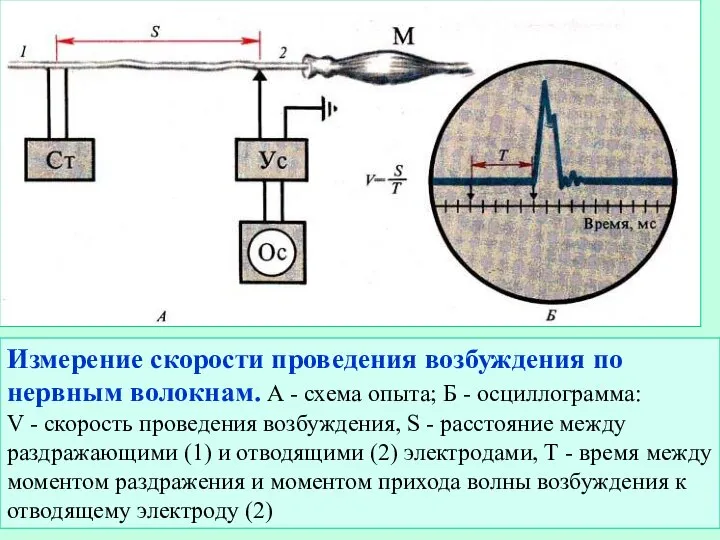
- 14. Измерение скорости проведения возбуждения по нервным волокнам. А - схема опыта; Б - осциллограмма: V -
- 15. Законы проведения возбуждения по нервным волокнам 1. З. изолированного проведения возбуждения: по отдельным нервным волокнам, проходящим
- 16. 4. З. двустороннего проведения возбуждения по нервному волокну: любое нервное волокно способно проводить возбуждение в обоих
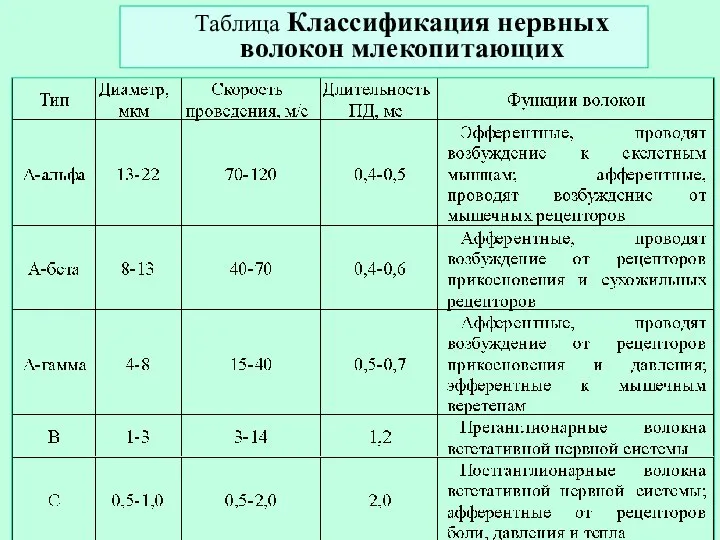
- 17. Таблица Классификация нервных волокон млекопитающих
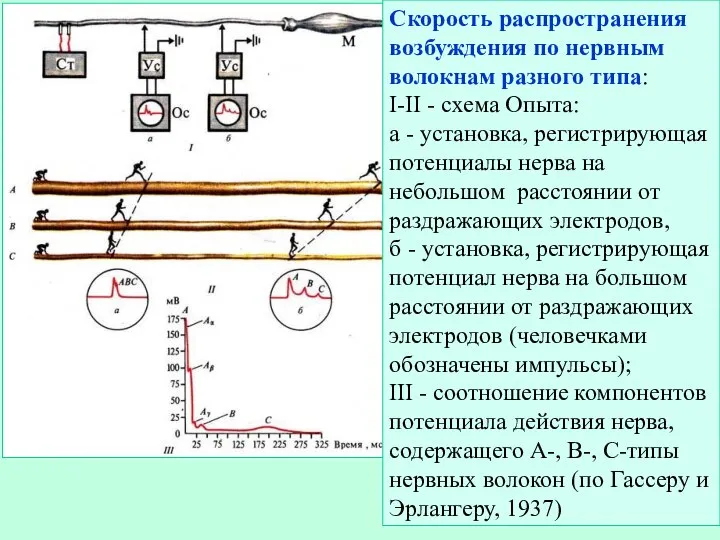
- 18. Скорость распространения возбуждения по нервным волокнам разного типа: I-II - схема Опыта: а - установка, регистрирующая
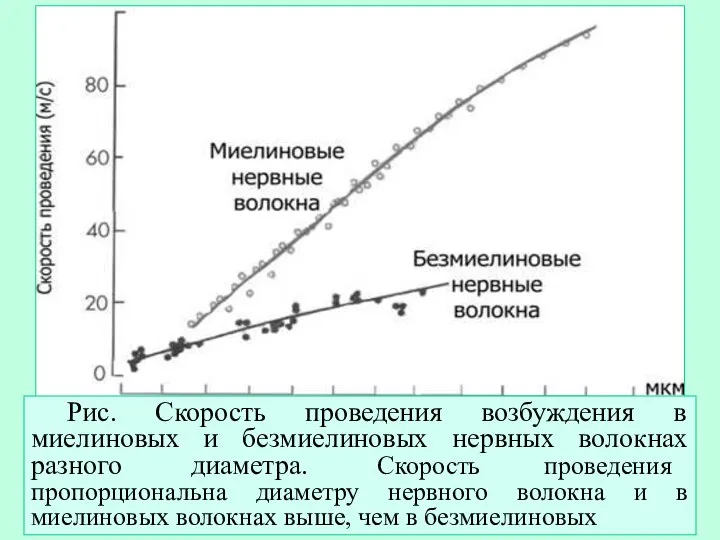
- 19. Рис. Скорость проведения возбуждения в миелиновых и безмиелиновых нервных волокнах разного диаметра. Скорость проведения пропорциональна диаметру
- 20. V. Синапсы Синапс – это морфофункциональное образование нервной системы, которое обеспечивает передачу сигнала с нейрона на
- 21. 2. По развитию в онтогенезе – стабильные (синапсы дуг безусловного рефлекса) и динамичные, появляющиеся в процессе
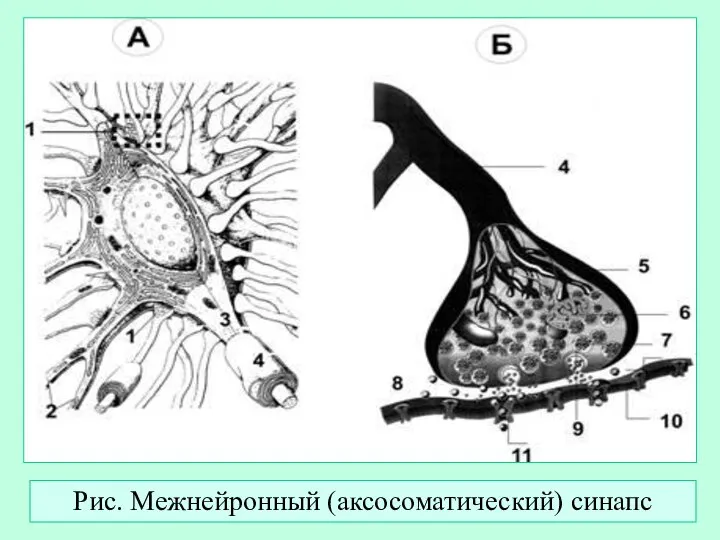
- 22. Рис. Межнейронный (аксосоматический) синапс
- 23. Механизм синаптической передачи 1. Под влиянием потенциала действия происходит деполяризация пресинаптической мембраны, 2. Повышается проницаемость кальциевых
- 24. 6. Деполяризация постсинаптической мембраны – возбуждающий постсинаптический потенциал (ВПСП). 7. Если его величина достигает критического уровня
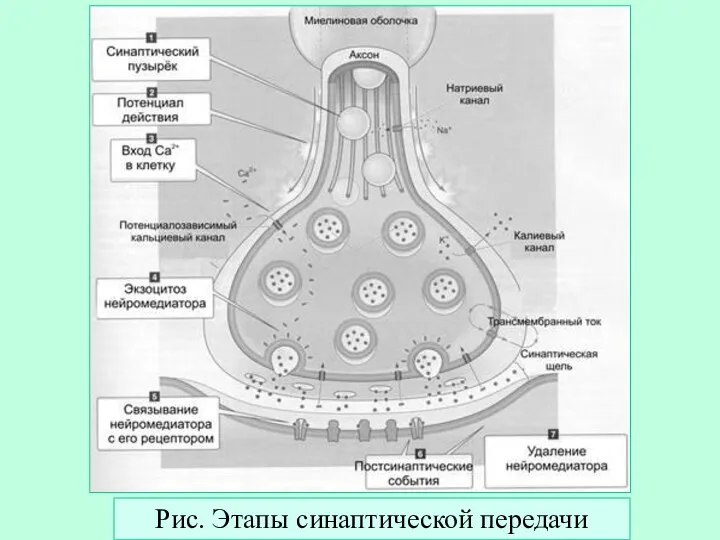
- 25. Рис. Этапы синаптической передачи
- 26. Свойства химических синапсов 1. Односторонняя проводимость – морфологическая и функциональная асимметрия синапса. 2. Синаптическая задержка: время
- 27. 5. Облегчение или потенциация - возрастание ответа постсинаптической мембраны при учащении подачи импульсов по аксону (накопление
- 29. Скачать презентацию

Слайд 3Рис. Многоотростчатый нейрон
Тело клетки (перикарион) содержит ядро. От перикариона отходят отростки. Один
Рис. Многоотростчатый нейрон
Тело клетки (перикарион) содержит ядро. От перикариона отходят отростки. Один
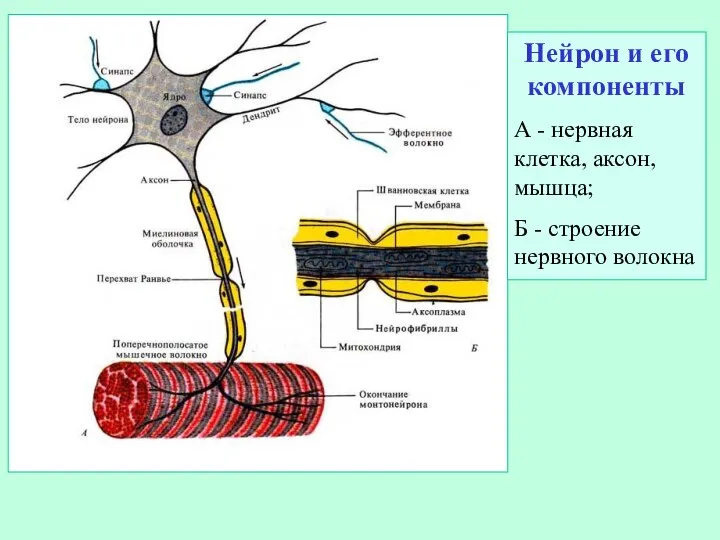
Слайд 4Нейрон и его компоненты
А - нервная клетка, аксон, мышца;
Б -
Нейрон и его компоненты
А - нервная клетка, аксон, мышца;
Б -
Слайд 5II. Функции нейрона: прием, кодирование, обработка, хранение и передача информации.
Нейроны
II. Функции нейрона: прием, кодирование, обработка, хранение и передача информации.
Нейроны

Слайд 6 Три состояния нейрона: покой, активность и торможение.
Покой - стабильный уровень мембранного
Три состояния нейрона: покой, активность и торможение.
Покой - стабильный уровень мембранного

Слайд 7Возможные состояния нейрона
Возможные состояния нейрона
Слайд 8Фоновая и вызванная импульсная активность нейронов (по Л. Шельцыну, 1980). А -
Фоновая и вызванная импульсная активность нейронов (по Л. Шельцыну, 1980). А -
Слайд 9 III. Нейроглия (от гр. neuron – нерв, и glia – клей) –
III. Нейроглия (от гр. neuron – нерв, и glia – клей) –

Слайд 10 Олигодендроциты окружают тела нейронов, входят в состав нервных волокон и нервных окончаний
Олигодендроциты окружают тела нейронов, входят в состав нервных волокон и нервных окончаний

Слайд 11IV. Нервные волокна
Главная функция нервных волокон - проведение нервных импульсов.
Механизм распространения нервного
IV. Нервные волокна
Главная функция нервных волокон - проведение нервных импульсов.
Механизм распространения нервного

Слайд 12Проведение возбуждения в нервных волокнах
(по Дж. Бендоллу, 1970):
I - немиелинизированное волокно
II
Проведение возбуждения в нервных волокнах
(по Дж. Бендоллу, 1970):
I - немиелинизированное волокно
II
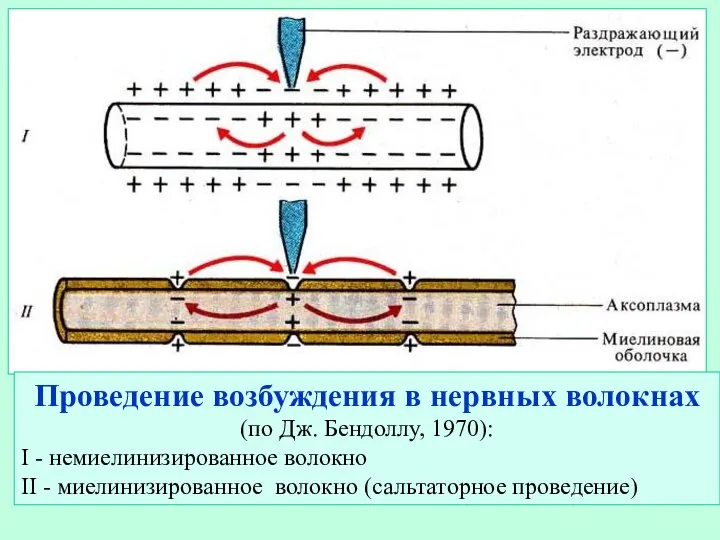
Слайд 13Рис. Проведение возбуждения в нервных волокнах. А - амиелиновое волокно, Б -
Рис. Проведение возбуждения в нервных волокнах. А - амиелиновое волокно, Б -
Слайд 14Измерение скорости проведения возбуждения по нервным волокнам. А - схема опыта; Б
Измерение скорости проведения возбуждения по нервным волокнам. А - схема опыта; Б
Слайд 15Законы проведения возбуждения
по нервным волокнам
1. З. изолированного проведения возбуждения: по
Законы проведения возбуждения
по нервным волокнам
1. З. изолированного проведения возбуждения: по

Слайд 164. З. двустороннего проведения возбуждения по нервному волокну: любое нервное волокно способно
4. З. двустороннего проведения возбуждения по нервному волокну: любое нервное волокно способно

Слайд 17Таблица Классификация нервных волокон млекопитающих
Таблица Классификация нервных волокон млекопитающих
Слайд 18Скорость распространения возбуждения по нервным волокнам разного типа:
I-II - схема Опыта:
Скорость распространения возбуждения по нервным волокнам разного типа:
I-II - схема Опыта:
Слайд 19 Рис. Скорость проведения возбуждения в миелиновых и безмиелиновых нервных волокнах разного диаметра.
Рис. Скорость проведения возбуждения в миелиновых и безмиелиновых нервных волокнах разного диаметра.
Слайд 20V. Синапсы
Синапс – это морфофункциональное образование нервной системы, которое обеспечивает передачу
V. Синапсы
Синапс – это морфофункциональное образование нервной системы, которое обеспечивает передачу

Слайд 212. По развитию в онтогенезе – стабильные (синапсы дуг безусловного рефлекса) и
2. По развитию в онтогенезе – стабильные (синапсы дуг безусловного рефлекса) и

Слайд 22Рис. Межнейронный (аксосоматический) синапс
Рис. Межнейронный (аксосоматический) синапс
Слайд 23Механизм синаптической передачи
1. Под влиянием потенциала действия происходит деполяризация пресинаптической мембраны,
Механизм синаптической передачи
1. Под влиянием потенциала действия происходит деполяризация пресинаптической мембраны,

Слайд 24 6. Деполяризация постсинаптической мембраны – возбуждающий постсинаптический потенциал (ВПСП).
7.
6. Деполяризация постсинаптической мембраны – возбуждающий постсинаптический потенциал (ВПСП).
7.

Слайд 25Рис. Этапы синаптической передачи
Рис. Этапы синаптической передачи
Слайд 26Свойства химических синапсов
1. Односторонняя проводимость – морфологическая и функциональная асимметрия синапса.
Свойства химических синапсов
1. Односторонняя проводимость – морфологическая и функциональная асимметрия синапса.

Слайд 275. Облегчение или потенциация - возрастание ответа постсинаптической мембраны при учащении подачи
5. Облегчение или потенциация - возрастание ответа постсинаптической мембраны при учащении подачи

 Найди рыбу. Игра
Найди рыбу. Игра Деление клетки. Митоз и мейоз
Деление клетки. Митоз и мейоз Защитные системы эукариот
Защитные системы эукариот Проверочная работа Органоиды клетки
Проверочная работа Органоиды клетки Подготовка к контрольному тестированию по теме «Дыхательная система»
Подготовка к контрольному тестированию по теме «Дыхательная система» Во что одеты растения
Во что одеты растения Общая характеристика класса Пресмыкающиеся (Рептилии)
Общая характеристика класса Пресмыкающиеся (Рептилии) 848856
848856 Презентация на тему Плоды
Презентация на тему Плоды  Зимующие птицы
Зимующие птицы Стебель – осевая часть побега растения
Стебель – осевая часть побега растения Презентация на тему Дыхание растений 6 класс
Презентация на тему Дыхание растений 6 класс  Тико-моделирование. Удивительное животное жираф
Тико-моделирование. Удивительное животное жираф Плоды и семена. 6 класс
Плоды и семена. 6 класс Муниципальное образовательное учреждение средняя общеобразовательная школа № 36 имени Героя Советского Союза В.Г. Миловатского г
Муниципальное образовательное учреждение средняя общеобразовательная школа № 36 имени Героя Советского Союза В.Г. Миловатского г Органы растений
Органы растений Углеводы или сахариды
Углеводы или сахариды Презентация на тему КТО ЖИВЕТ В ПОЧВЕ
Презентация на тему КТО ЖИВЕТ В ПОЧВЕ  Великий круговорот жизни
Великий круговорот жизни Дикие утки
Дикие утки Кишечнополостные
Кишечнополостные Вегетативное развитие различных растений в зависимости от норм фосфорных удобрений
Вегетативное развитие различных растений в зависимости от норм фосфорных удобрений Минеральное питание растений
Минеральное питание растений Одномембранные органеллы
Одномембранные органеллы Дикие животные
Дикие животные Усики, лапки, крылышки
Усики, лапки, крылышки Экто- и эндотермная регуляция температуры тела
Экто- и эндотермная регуляция температуры тела Цветочная посуда и инструменты
Цветочная посуда и инструменты