- Физиология возбудимых тканей

Содержание
- 2. Введение Если рассмотреть электронную микрофотографию ультратонкого среза живой ткани (после его фиксации и соответствующего прокрашивания), то
- 8. Последующие исследования, выполненные с помощью электронного микроскопа в 1959 г. Дж. Робертсоном, подтвердили правильность этой модели.
- 9. Функции биологических мембран Во всех живых клетках биологические мембраны выполняют функцию барьера, отделяющего клетку от окружающей
- 10. 4. Гибкая и эластичная пленка, которой по существу являются все мембраны, выполняет и определенную механическую функцию,
- 13. Липиды мембран В состав липидов мембран входят в основном фосфолипиды, сфингомиелины и холестерин. Основные фосфолипиды мембран
- 16. Полярные группы молекул глобулярных белков направлены наружу, в сторону водной фазы, а неполярные — в сторону
- 17. В физиологических условиях мембраны обладают динамическими свойствами. Липидный бислой по существу является вязкой жидкостью и характеризуется
- 18. БЕЛКИ, СВЯЗАННЫЕ С БИСЛОЕМ ЛИПИДОВ Все мембраны, помимо фосфолипидов, содержат белки, которые делят на две группы.
- 19. Схематическое изображение структуры цитоплазматической мембраны
- 20. Интегральные белки выступают в роли: гидролитических ферментов, рецепторов клеточной поверхности, окислительно-восстановительных компонентов транспортной системы электронов, в
- 21. Главные компоненты клеточных оболочек высших организмов представлены: углеводными компонентами гликолипидов, в частности цереброзидов и ганглиозидов; гликопротеидами;
- 22. РАЗДРАЖИМОСТЬ И РАЗДРАЖИТЕЛИ Живые организмы и все их клетки обладают свойством, называемым «раздражимостью», т.е. способностью отвечать
- 23. Все бесконечное разнообразие возможных раздра-жителей клеток и тканей можно классифицировать по трем категориям — по адекватности,
- 24. По природе раздражители можно разделить на множество групп. Это физические, физико-химические и химические. К числу физических
- 25. По силе раздражители можно разделить на подпороговые, пороговые и сверхпороговые. Применение подпороговых раздражителей не приводит к
- 26. Возбудимость — это свойство клеточных мембран отвечать на действие адекватных раздражителей специфическими изменениями ионной проницаемости и
- 27. Транспорт веществ через мембрану При всем многообразии строения и физико-химических свойств молекул проникающих веществ можно выделить
- 28. Транспорт веществ через мембрану
- 30. Посредством простой диффузии без помощи специального переносчика, Во-первых, осуществляется транспорт соединений непосредственно через липидный бислой. В
- 31. ТРАНСПОРТНЫЕ СИСТЕМЫ Определенные молекулы, а также ионы могут проходить через мембрану не путем диффузии, а посредством
- 32. Процесс облегченной диффузии подразумевает перемещение переносчиком вещества через мембрану по градиенту концентрации Активный транспорт опосредован переносчиком,
- 33. Облегченная диффузия Термин «облегченная диффузия» возник потому, что конечный результат диффузии и облегченной диффузии одинаков. В
- 34. Активный транспорт Для активного транспорта известно два варианта использования переносчиками энергии: прямое потребление АТФ в первично
- 35. Мембранный потенциал покоя является результатом разделения зарядов относительно клеточной мембраны В покое снаружи мембраны преобладают положительные
- 36. А- - органические анионы
- 37. 0 -30 -60 Введение электрода Мембранный потенциал покоя Время А Б Внутриклеточная микроэлектродная регистрация Величина МПП
- 38. В состоянии физиологического покоя мембрана нервных волокон в 25 раз более проницаема для ионов калия, чем
- 39. Расчет равновесного потенциала Равновесный потенциал для какого-либо иона Х можно рассчитать из уравнения, полученного в 1888
- 40. Мембранный потенциал покоя (-70 мВ) близок к его Еk (-75 мВ), но не точно равен ему,
- 41. Равновесные потенциалы(Е) Cl-каналы -89 - 47 -89 - 47 Cl-
- 42. Структуры мембраны, формирующие потенциал покоя
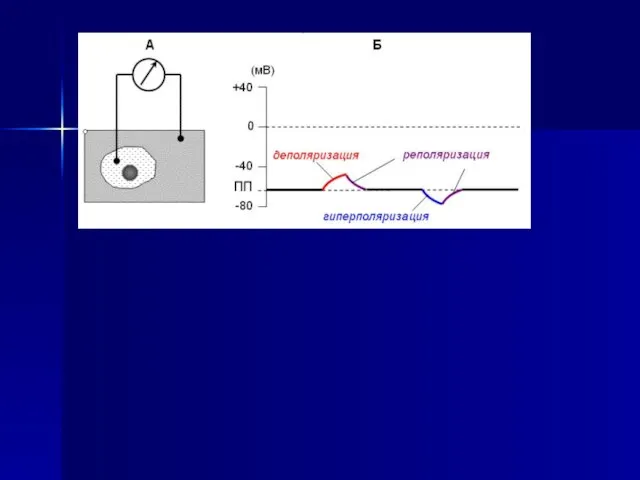
- 43. Быстрое колебание мембранного потенциала клетки в ответ на раздражение, сопровождающееся изменением знака заряда на мембране, возникающее
- 44. Фаза деполяризации Фаза реполяризации Раздражающий импульс
- 45. Потенциал действия зависит от внеклеточного Na
- 46. Блокирование потенциал-управляемых натриевых каналов нарушает генерацию потенциала действия Тетродотоксин – специфический блокатор натриевых каналов
- 47. Связь работы ионных каналов с фазами потенциала действия Раздражение Деполяризация (уменьшение мембранного потенциала) Быстрая активация натриевых
- 48. Временной ход ионных токов во время потенциала действия
- 49. Рефрактерность - снижение способности клетки отвечать на раздражение в результате временной инактивации натриевых каналов Абсолютная рефрактерность
- 50. Изменение мембранного потенциала, интенсивности трансмембранных токов и возбудимости клетки в разные фазы потенциала действия. Д –
- 51. Связь потенциала действия с ионными механизмами на качественном уровне, (а) Форма потенциала действия и его основные
- 52. ПД различных клеток: Нервной клетки Мышечной клетки Рабочего кардиомиоцита
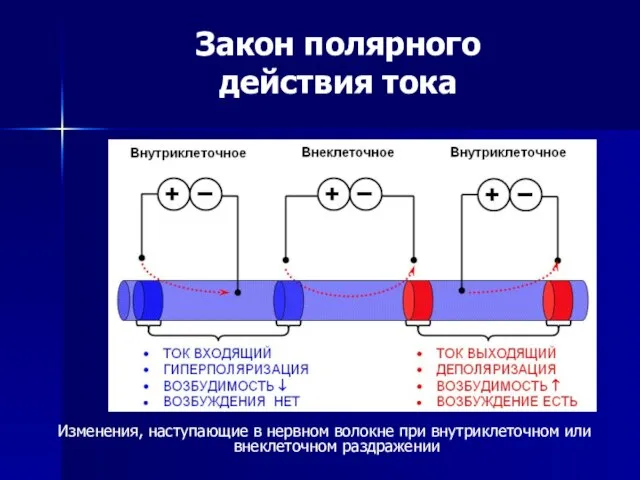
- 53. Законы электрического раздражения возбудимых тканей Закон полярного действия тока В случае приложения к объекту двух внешних
- 54. Закон полярного действия тока Изменения, наступающие в нервном волокне при внутриклеточном или внеклеточном раздражении
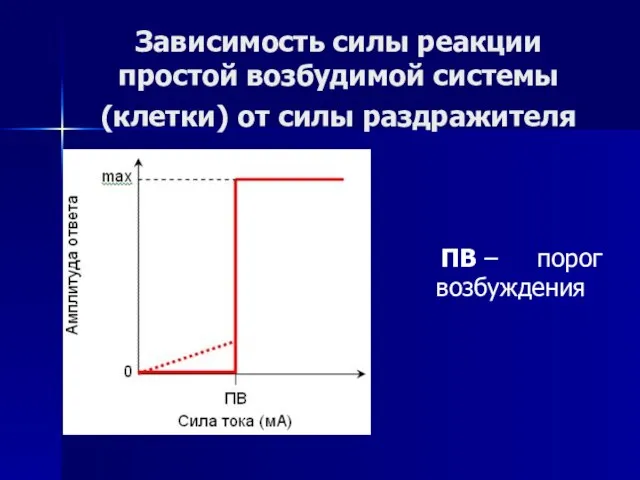
- 55. Зависимость силы реакции простой возбудимой системы (клетки) от силы раздражителя ПВ – порог возбуждения
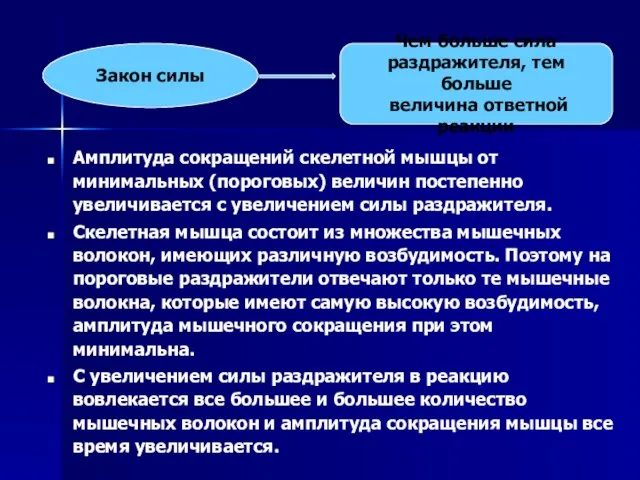
- 56. Амплитуда сокращений скелетной мышцы от минимальных (пороговых) величин постепенно увеличивается с увеличением силы раздражителя. Скелетная мышца
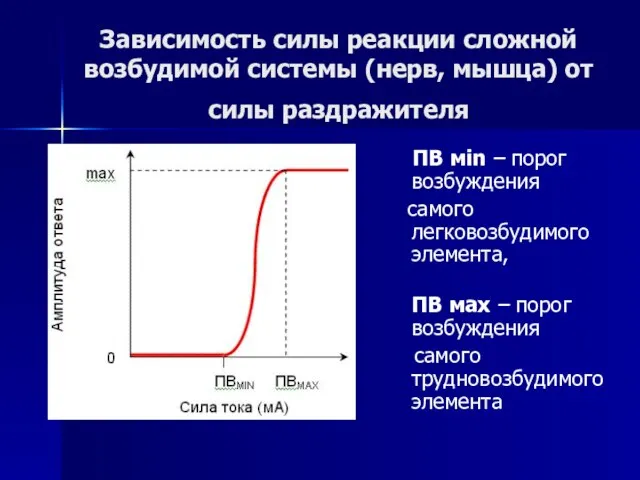
- 57. Зависимость силы реакции сложной возбудимой системы (нерв, мышца) от силы раздражителя ПВ мin – порог возбуждения
- 58. Законы электрического раздражения возбудимых тканей Закон крутизны раздражения Для раздражения деполяризующий ток должен нарастать достаточно круто.
- 59. Аккомодация - позитивное смещение КУД при длительной деполяризации Объясняется частичной инактивацией натриевых каналов и активацией калиевых.
- 60. Изменение мембранного потенциала и критического уровня деполяризации при медленном ( А ) и быстром ( Б
- 61. Закон силы-длительности раздражающее действие постоянного тока зависит не только от его величины, но и от времени,
- 62. Пороговая сила деполяризующего тока в известной мере зависит от длительности действия раздражающего тока. Эта зависимость выражается
- 63. Минимальная величина тока, способная вызвать возбуждение при неограниченно длительном его действии, называется реобазой. Время, в течение
- 64. Ток ниже некоторой минимальной величины не вызывает возбуждение, как бы длительно он не действовал, и чем
- 65. Изменение МП и КУД при действии длительного тока выходящего (деполяризующего) тока, ФКэт – физиологический катэлектротон, КД
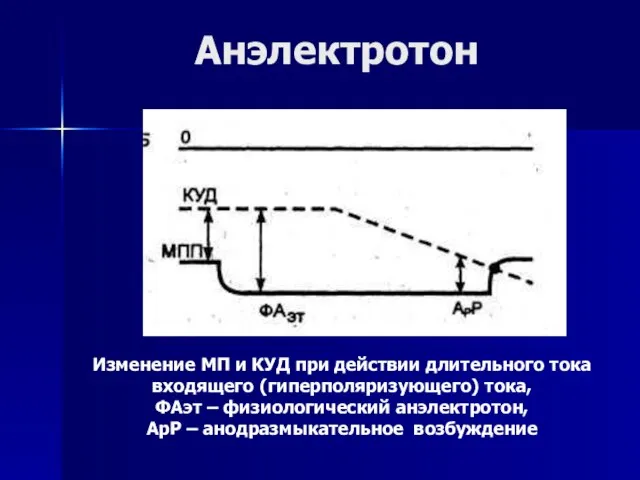
- 66. Анэлектротон Изменение МП и КУД при действии длительного тока входящего (гиперполяризующего) тока, ФАэт – физиологический анэлектротон,
- 69. Скачать презентацию
Слайд 2Введение
Если рассмотреть электронную микрофотографию ультратонкого среза живой ткани (после его фиксации и
Введение
Если рассмотреть электронную микрофотографию ультратонкого среза живой ткани (после его фиксации и



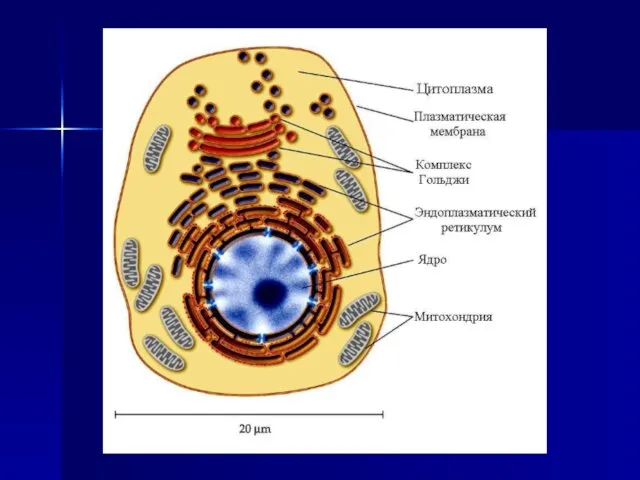
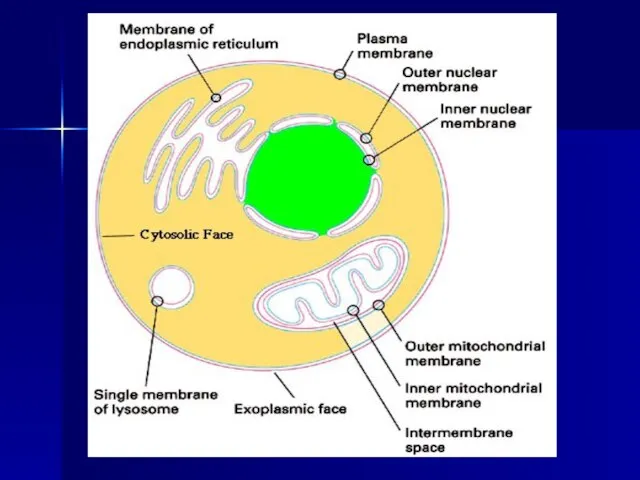

Слайд 8Последующие исследования, выполненные с помощью электронного микроскопа в 1959 г. Дж. Робертсоном,
Последующие исследования, выполненные с помощью электронного микроскопа в 1959 г. Дж. Робертсоном,

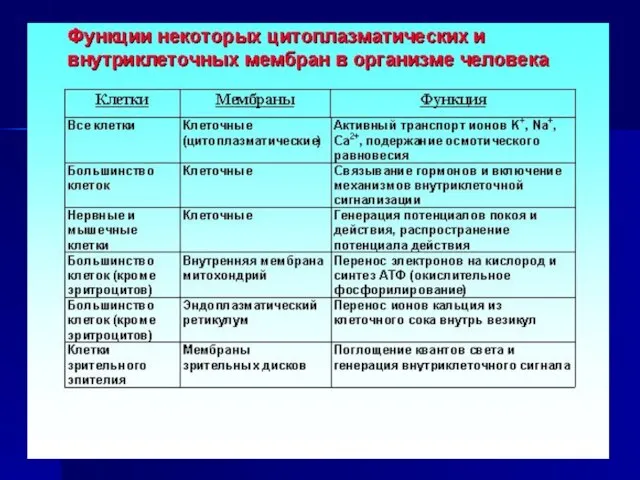
Слайд 9Функции биологических мембран
Во всех живых клетках биологические мембраны выполняют функцию барьера,
Функции биологических мембран
Во всех живых клетках биологические мембраны выполняют функцию барьера,

Слайд 104. Гибкая и эластичная пленка, которой по существу являются все мембраны, выполняет
4. Гибкая и эластичная пленка, которой по существу являются все мембраны, выполняет


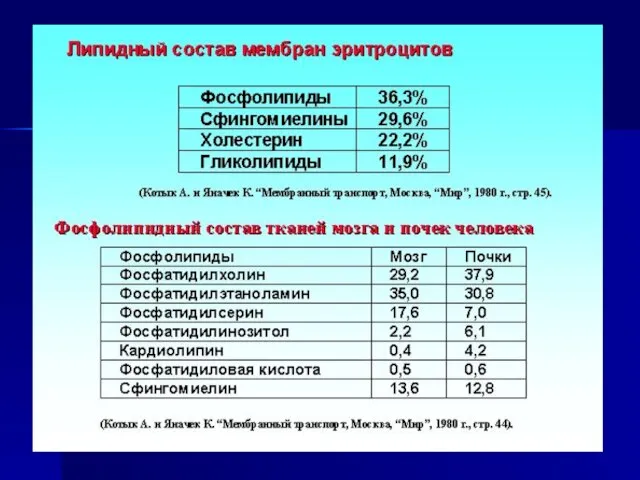
Слайд 13Липиды мембран
В состав липидов мембран входят в основном фосфолипиды, сфингомиелины и холестерин.
Липиды мембран
В состав липидов мембран входят в основном фосфолипиды, сфингомиелины и холестерин.


Слайд 16Полярные группы молекул глобулярных белков направлены наружу, в сторону водной фазы, а
Полярные группы молекул глобулярных белков направлены наружу, в сторону водной фазы, а

Слайд 17В физиологических условиях мембраны обладают динамическими свойствами. Липидный бислой по существу является
В физиологических условиях мембраны обладают динамическими свойствами. Липидный бислой по существу является

Слайд 18БЕЛКИ, СВЯЗАННЫЕ С БИСЛОЕМ ЛИПИДОВ
Все мембраны, помимо фосфолипидов, содержат белки, которые делят
БЕЛКИ, СВЯЗАННЫЕ С БИСЛОЕМ ЛИПИДОВ
Все мембраны, помимо фосфолипидов, содержат белки, которые делят

Слайд 19Схематическое изображение структуры
цитоплазматической мембраны
Схематическое изображение структуры
цитоплазматической мембраны
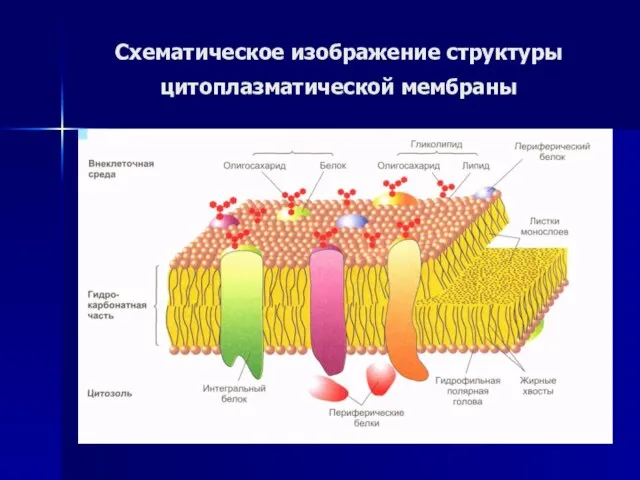
Слайд 20Интегральные белки выступают в роли:
гидролитических ферментов,
рецепторов клеточной поверхности,
окислительно-восстановительных компонентов
Интегральные белки выступают в роли:
гидролитических ферментов,
рецепторов клеточной поверхности,
окислительно-восстановительных компонентов

Слайд 21Главные компоненты клеточных оболочек высших организмов представлены:
углеводными компонентами гликолипидов, в частности
Главные компоненты клеточных оболочек высших организмов представлены:
углеводными компонентами гликолипидов, в частности

Слайд 22РАЗДРАЖИМОСТЬ И РАЗДРАЖИТЕЛИ
Живые организмы и все их клетки обладают свойством, называемым «раздражимостью»,
РАЗДРАЖИМОСТЬ И РАЗДРАЖИТЕЛИ
Живые организмы и все их клетки обладают свойством, называемым «раздражимостью»,

Слайд 23Все бесконечное разнообразие возможных раздра-жителей клеток и тканей можно классифицировать по трем
Все бесконечное разнообразие возможных раздра-жителей клеток и тканей можно классифицировать по трем

Слайд 24По природе раздражители можно разделить на множество групп.
Это физические, физико-химические
По природе раздражители можно разделить на множество групп.
Это физические, физико-химические

Слайд 25По силе раздражители можно разделить на подпороговые, пороговые и сверхпороговые.
Применение подпороговых раздражителей
По силе раздражители можно разделить на подпороговые, пороговые и сверхпороговые.
Применение подпороговых раздражителей

Слайд 26Возбудимость — это свойство клеточных мембран отвечать на действие адекватных раздражителей специфическими
Возбудимость — это свойство клеточных мембран отвечать на действие адекватных раздражителей специфическими

Слайд 27Транспорт веществ через мембрану
При всем многообразии строения и физико-химических свойств молекул проникающих
Транспорт веществ через мембрану
При всем многообразии строения и физико-химических свойств молекул проникающих

Слайд 28Транспорт веществ через мембрану
Транспорт веществ через мембрану
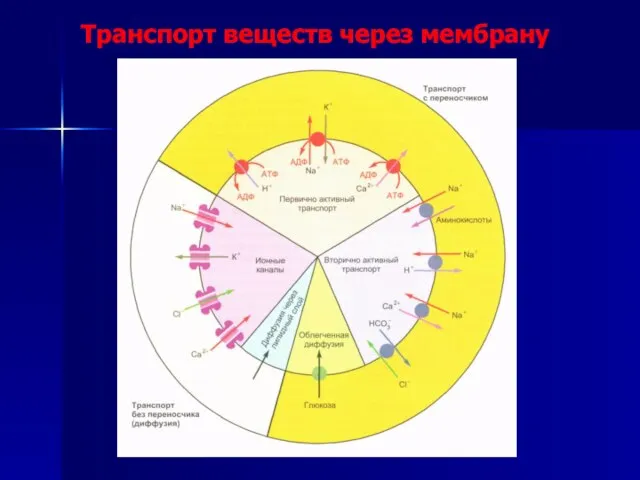
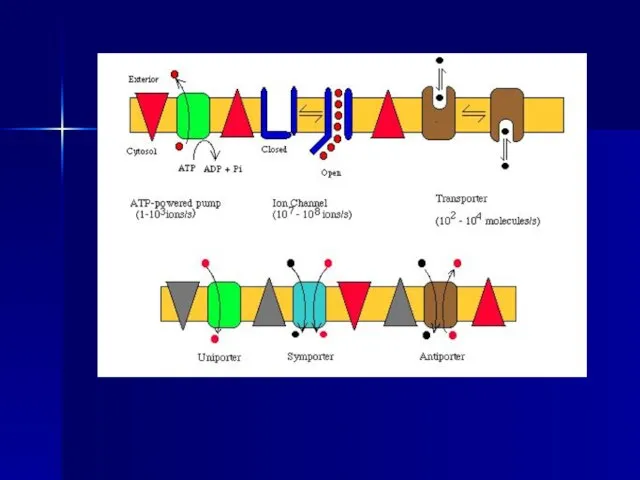
Слайд 30Посредством простой диффузии без помощи специального переносчика,
Во-первых, осуществляется транспорт соединений непосредственно
Посредством простой диффузии без помощи специального переносчика,
Во-первых, осуществляется транспорт соединений непосредственно

Слайд 31ТРАНСПОРТНЫЕ СИСТЕМЫ
Определенные молекулы, а также ионы могут проходить через мембрану не путем
ТРАНСПОРТНЫЕ СИСТЕМЫ
Определенные молекулы, а также ионы могут проходить через мембрану не путем

Слайд 32Процесс
облегченной диффузии подразумевает
перемещение переносчиком вещества через
мембрану по градиенту концентрации
Активный
Процесс
облегченной диффузии подразумевает
перемещение переносчиком вещества через
мембрану по градиенту концентрации
Активный
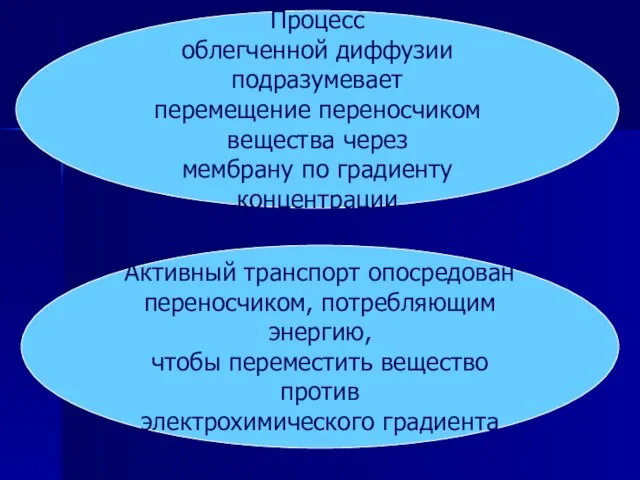
Слайд 33Облегченная диффузия
Термин «облегченная диффузия» возник потому, что конечный результат диффузии и облегченной
Облегченная диффузия
Термин «облегченная диффузия» возник потому, что конечный результат диффузии и облегченной

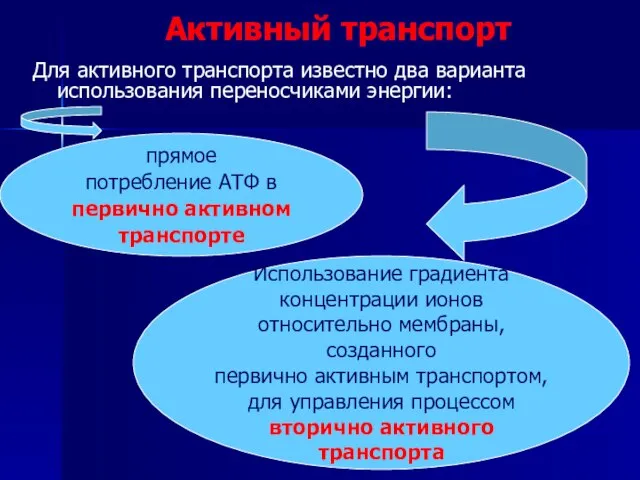
Слайд 34Активный транспорт
Для активного транспорта известно два варианта использования переносчиками энергии:
прямое
потребление
Активный транспорт
Для активного транспорта известно два варианта использования переносчиками энергии:
прямое
потребление
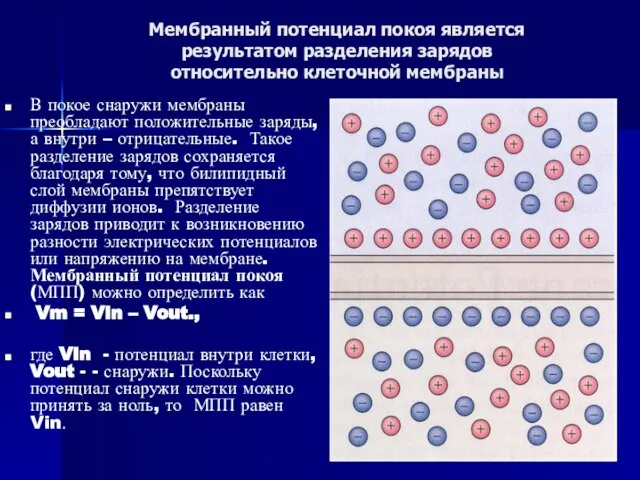
Слайд 35Мембранный потенциал покоя является
результатом разделения зарядов
относительно клеточной мембраны
В покое снаружи мембраны
Мембранный потенциал покоя является
результатом разделения зарядов
относительно клеточной мембраны
В покое снаружи мембраны
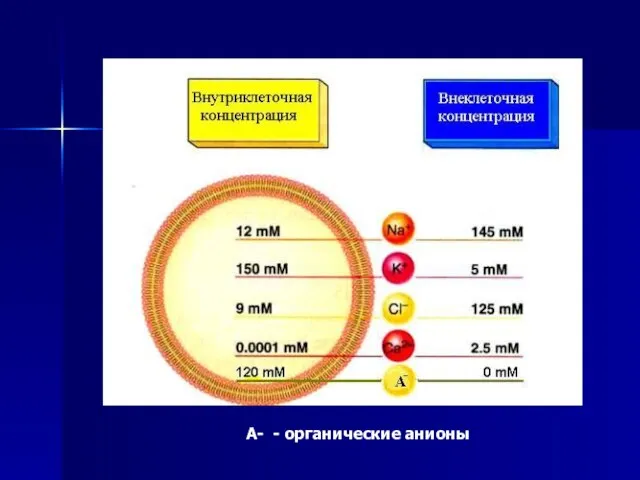
Слайд 36А- - органические анионы
А- - органические анионы
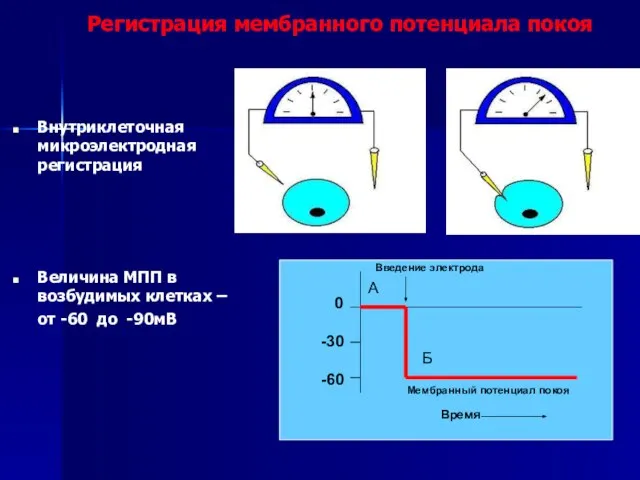
Слайд 37 0
-30
-60
Введение электрода
Мембранный потенциал покоя
Время
А
Б
Внутриклеточная микроэлектродная регистрация
Величина МПП в возбудимых клетках
0
-30
-60
Введение электрода
Мембранный потенциал покоя
Время
А
Б
Внутриклеточная микроэлектродная регистрация
Величина МПП в возбудимых клетках
Слайд 38 В состоянии физиологического покоя мембрана нервных волокон в 25 раз более
В состоянии физиологического покоя мембрана нервных волокон в 25 раз более

Слайд 39Расчет равновесного потенциала
Равновесный потенциал для какого-либо иона Х можно рассчитать из уравнения,
Расчет равновесного потенциала
Равновесный потенциал для какого-либо иона Х можно рассчитать из уравнения,

Слайд 40Мембранный потенциал покоя (-70 мВ) близок к его Еk (-75 мВ), но
Мембранный потенциал покоя (-70 мВ) близок к его Еk (-75 мВ), но

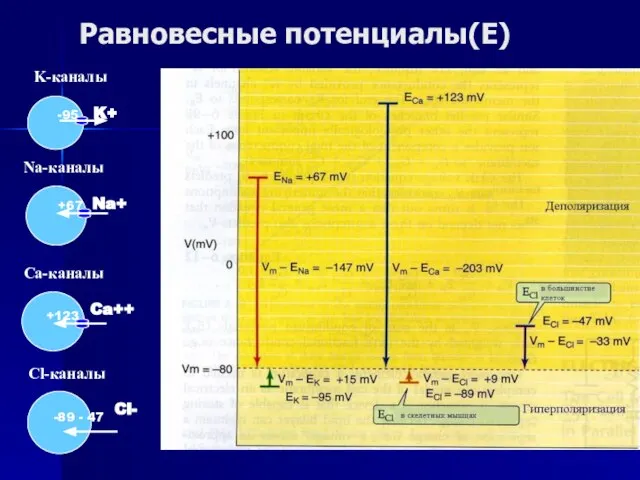
Слайд 41Равновесные потенциалы(Е)
Cl-каналы
-89 - 47
-89 - 47
Cl-
Равновесные потенциалы(Е)
Cl-каналы
-89 - 47
-89 - 47
Cl-
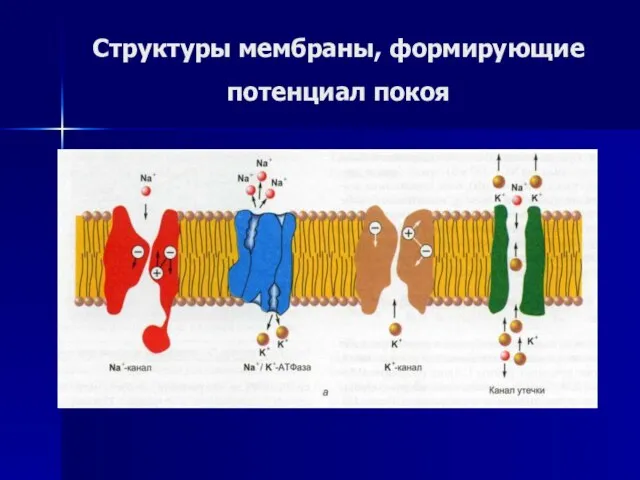
Слайд 42Структуры мембраны, формирующие потенциал покоя
Структуры мембраны, формирующие потенциал покоя
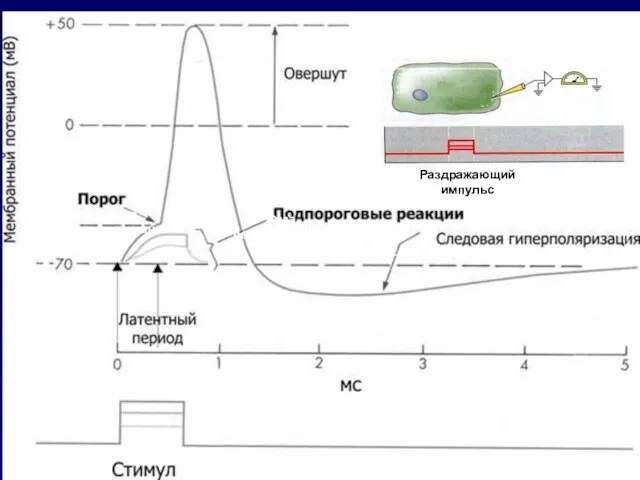
Слайд 43Быстрое колебание мембранного потенциала клетки в ответ на раздражение, сопровождающееся изменением знака
Быстрое колебание мембранного потенциала клетки в ответ на раздражение, сопровождающееся изменением знака

Слайд 44Фаза
деполяризации
Фаза
реполяризации
Раздражающий
импульс
Фаза
деполяризации
Фаза
реполяризации
Раздражающий
импульс
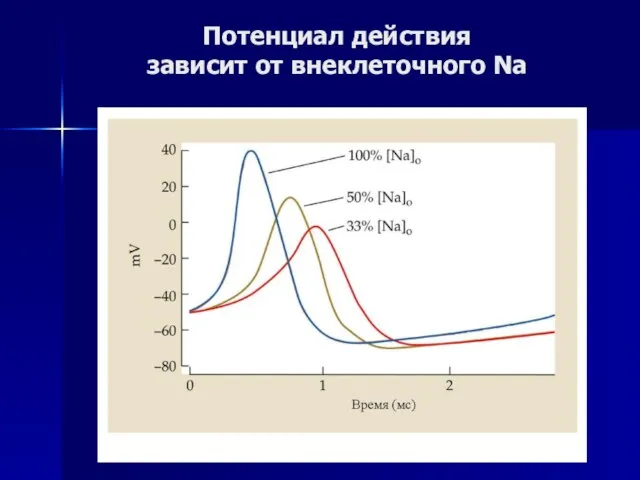
Слайд 45Потенциал действия
зависит от внеклеточного Na
Потенциал действия
зависит от внеклеточного Na
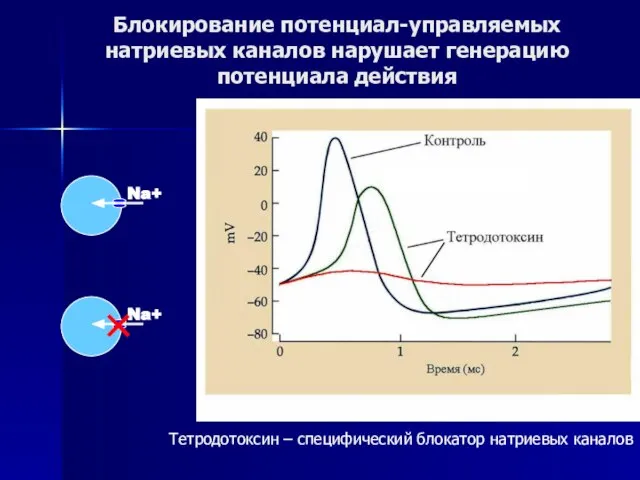
Слайд 46Блокирование потенциал-управляемых натриевых каналов нарушает генерацию потенциала действия
Тетродотоксин – специфический блокатор натриевых
Блокирование потенциал-управляемых натриевых каналов нарушает генерацию потенциала действия
Тетродотоксин – специфический блокатор натриевых
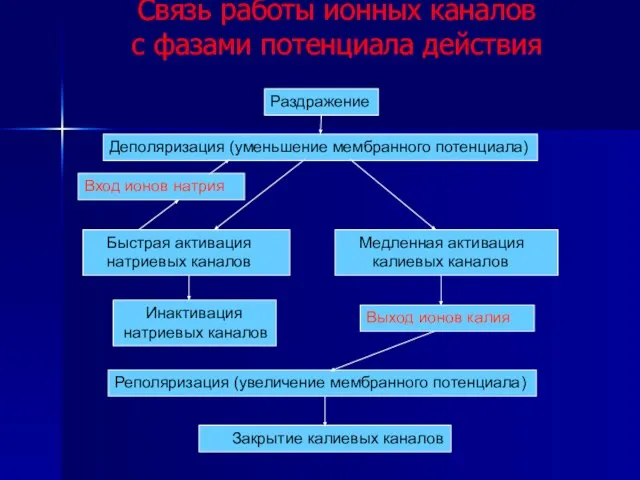
Слайд 47Связь работы ионных каналов
с фазами потенциала действия
Раздражение
Деполяризация (уменьшение мембранного потенциала)
Быстрая
Связь работы ионных каналов
с фазами потенциала действия
Раздражение
Деполяризация (уменьшение мембранного потенциала)
Быстрая
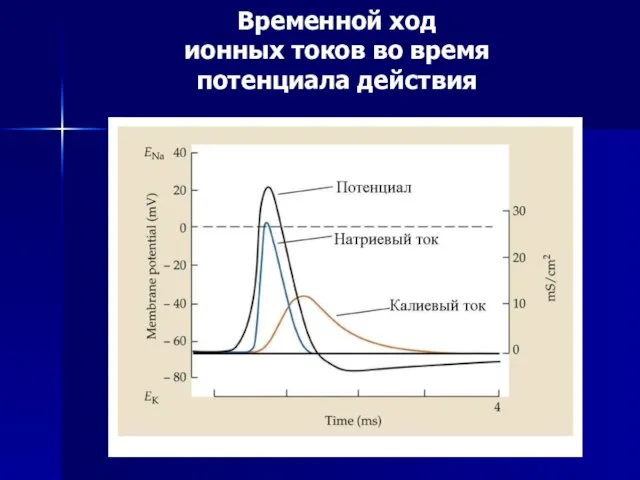
Слайд 48Временной ход
ионных токов во время
потенциала действия
Временной ход
ионных токов во время
потенциала действия
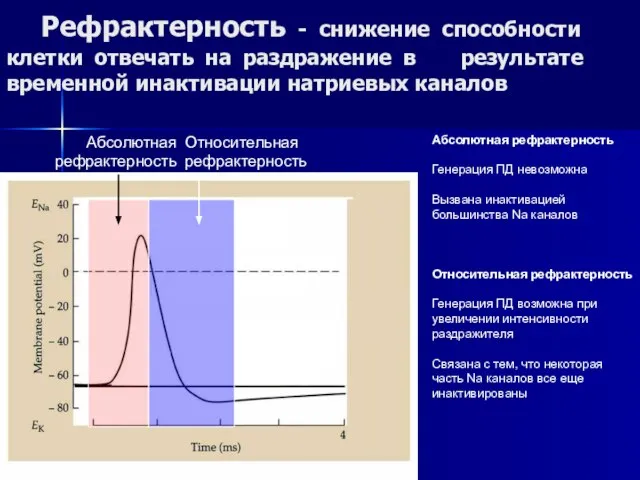
Слайд 49 Рефрактерность - снижение способности клетки отвечать на раздражение в результате временной
Рефрактерность - снижение способности клетки отвечать на раздражение в результате временной
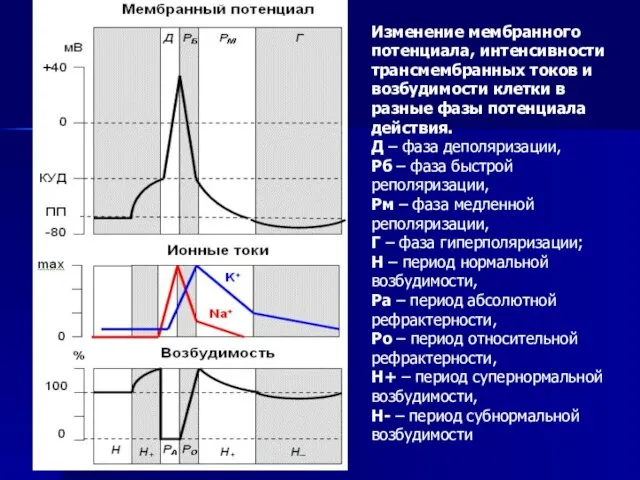
Слайд 50Изменение мембранного потенциала, интенсивности трансмембранных токов и возбудимости клетки в разные фазы
Изменение мембранного потенциала, интенсивности трансмембранных токов и возбудимости клетки в разные фазы
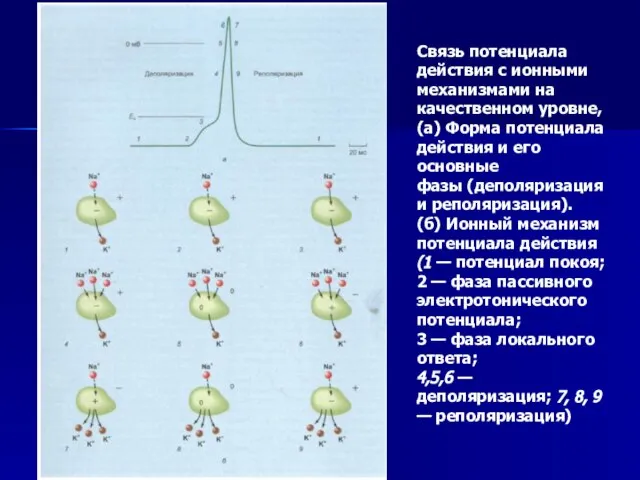
Слайд 51Связь потенциала действия с ионными механизмами на качественном уровне,
(а) Форма потенциала
Связь потенциала действия с ионными механизмами на качественном уровне,
(а) Форма потенциала
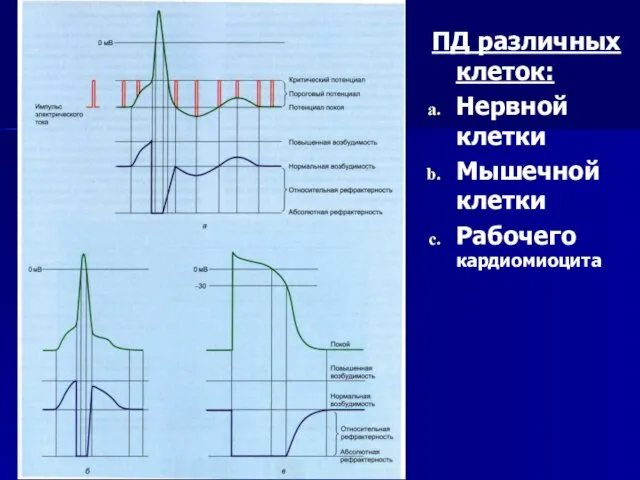
Слайд 52ПД различных клеток:
Нервной клетки
Мышечной клетки
Рабочего кардиомиоцита
ПД различных клеток:
Нервной клетки
Мышечной клетки
Рабочего кардиомиоцита
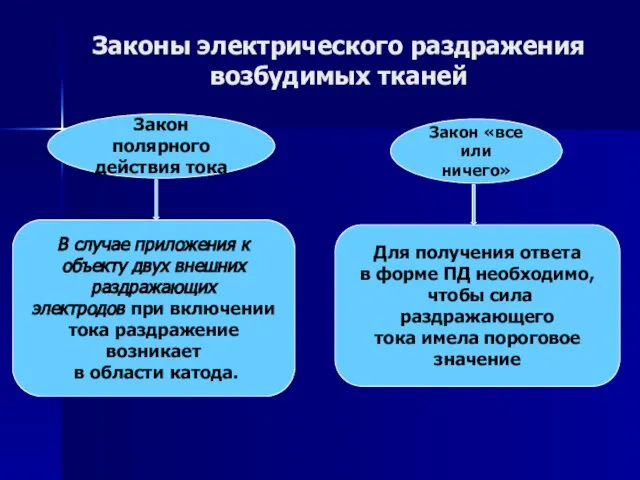
Слайд 53Законы электрического раздражения
возбудимых тканей
Закон полярного
действия тока
В случае приложения к
объекту двух
Законы электрического раздражения
возбудимых тканей
Закон полярного
действия тока
В случае приложения к
объекту двух
Слайд 54Закон полярного
действия тока
Изменения, наступающие в нервном волокне при внутриклеточном или внеклеточном
Закон полярного
действия тока
Изменения, наступающие в нервном волокне при внутриклеточном или внеклеточном
Слайд 55Зависимость силы реакции простой возбудимой системы (клетки) от силы раздражителя
ПВ
Зависимость силы реакции простой возбудимой системы (клетки) от силы раздражителя
ПВ
Слайд 56Амплитуда сокращений скелетной мышцы от минимальных (пороговых) величин постепенно увеличивается с увеличением
Амплитуда сокращений скелетной мышцы от минимальных (пороговых) величин постепенно увеличивается с увеличением
Слайд 57Зависимость силы реакции сложной возбудимой системы (нерв, мышца) от силы раздражителя
Зависимость силы реакции сложной возбудимой системы (нерв, мышца) от силы раздражителя
Слайд 58Законы электрического раздражения
возбудимых тканей
Закон крутизны
раздражения
Для раздражения
деполяризующий ток
должен нарастать
достаточно круто.
В
Законы электрического раздражения
возбудимых тканей
Закон крутизны
раздражения
Для раздражения
деполяризующий ток
должен нарастать
достаточно круто.
В
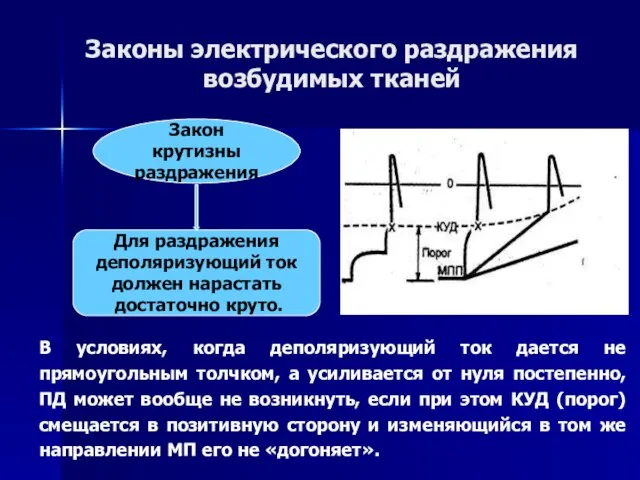
Слайд 59Аккомодация - позитивное смещение КУД при длительной деполяризации
Объясняется частичной инактивацией натриевых
Аккомодация - позитивное смещение КУД при длительной деполяризации
Объясняется частичной инактивацией натриевых

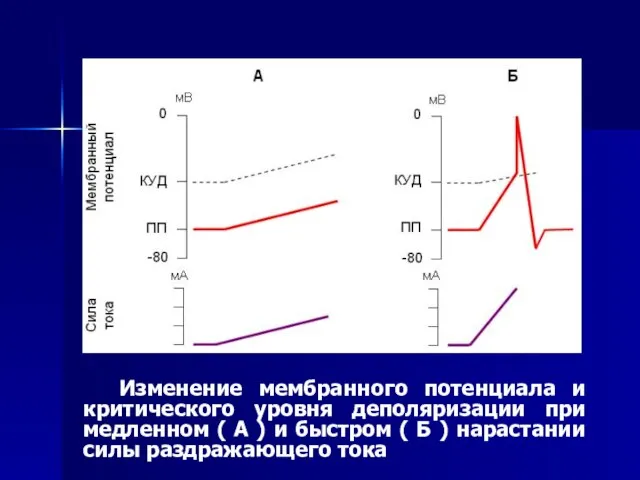
Слайд 60 Изменение мембранного потенциала и критического уровня деполяризации при медленном ( А
Изменение мембранного потенциала и критического уровня деполяризации при медленном ( А
Слайд 61Закон силы-длительности
раздражающее действие постоянного тока зависит не только от его величины,
Закон силы-длительности
раздражающее действие постоянного тока зависит не только от его величины,

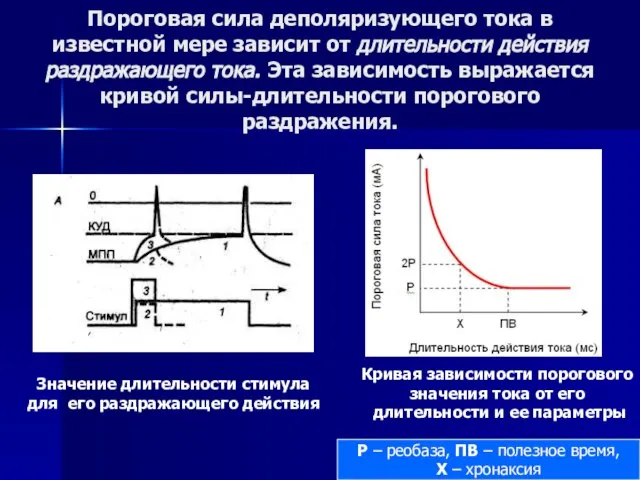
Слайд 62Пороговая сила деполяризующего тока в известной мере зависит от длительности действия раздражающего
Пороговая сила деполяризующего тока в известной мере зависит от длительности действия раздражающего
Слайд 63Минимальная величина тока, способная вызвать возбуждение при неограниченно длительном его действии, называется
Минимальная величина тока, способная вызвать возбуждение при неограниченно длительном его действии, называется

Слайд 64Ток ниже некоторой минимальной величины не вызывает возбуждение, как бы длительно он
Ток ниже некоторой минимальной величины не вызывает возбуждение, как бы длительно он

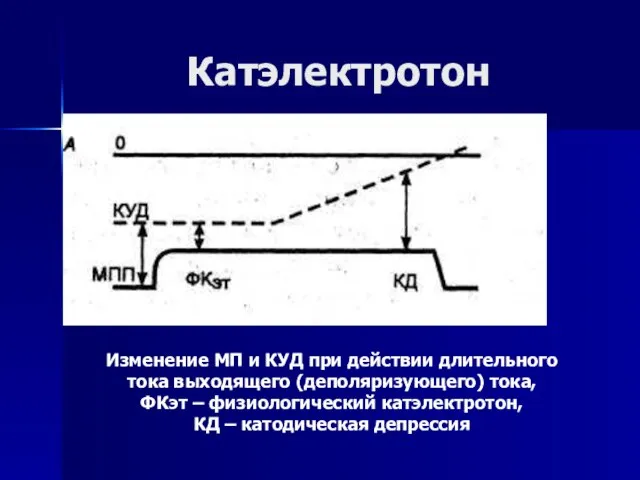
Слайд 65Изменение МП и КУД при действии длительного тока выходящего (деполяризующего) тока,
ФКэт
Изменение МП и КУД при действии длительного тока выходящего (деполяризующего) тока,
ФКэт
Слайд 66Анэлектротон
Изменение МП и КУД при действии длительного тока
входящего (гиперполяризующего) тока,
ФАэт
Анэлектротон
Изменение МП и КУД при действии длительного тока
входящего (гиперполяризующего) тока,
ФАэт
 Позвоночные животные
Позвоночные животные Методы биологических исследований. Таблица ОГЭ -> ЕГЭ с примерами
Методы биологических исследований. Таблица ОГЭ -> ЕГЭ с примерами Презентация на тему Вода в жизни растений и животных
Презентация на тему Вода в жизни растений и животных  Дополнительная литература по биологии за 6-8 классы
Дополнительная литература по биологии за 6-8 классы Препараты витаминов,гормонов,кровезаменителей и антигистаминных средств. Лекция 8
Препараты витаминов,гормонов,кровезаменителей и антигистаминных средств. Лекция 8 Учёт численности сообщества настоящих тюленей семейства Phocidae в заливе Пильтун (остров Сахалин) c применением БПЛА в 2020 г
Учёт численности сообщества настоящих тюленей семейства Phocidae в заливе Пильтун (остров Сахалин) c применением БПЛА в 2020 г ТОРМОЖЕНИЕ (1)
ТОРМОЖЕНИЕ (1) Мейоз
Мейоз Надкласс Рыбы. Общая характеристика и внешнее строение
Надкласс Рыбы. Общая характеристика и внешнее строение Класс Птицы
Класс Птицы 1_2_Metody_tsitologii_Kletochnaya_teoria_Pro_i_Eukarioty_Sravnenie_Tsarstv_RZhG (1)
1_2_Metody_tsitologii_Kletochnaya_teoria_Pro_i_Eukarioty_Sravnenie_Tsarstv_RZhG (1) Анатомия и физиология головного мозга
Анатомия и физиология головного мозга Опытническая работа Методика проращивания кофе аравийского (cofféa arábica) в условиях учебной теплицы
Опытническая работа Методика проращивания кофе аравийского (cofféa arábica) в условиях учебной теплицы Разнообразие условий существования для жизни растений
Разнообразие условий существования для жизни растений Самые быстрые птицы
Самые быстрые птицы Презентация на тему Постэмбриональное развитие
Презентация на тему Постэмбриональное развитие  Липиды
Липиды Дыхательная система человека
Дыхательная система человека Предпосылки развития теории Ч.Дарвина
Предпосылки развития теории Ч.Дарвина Эволюционная физиология
Эволюционная физиология Зачет Анатомия и морфология растений, особенности жизнедеятельности
Зачет Анатомия и морфология растений, особенности жизнедеятельности Особенности питания и системы удобрения земляники садовой
Особенности питания и системы удобрения земляники садовой Органические вещества клетки
Органические вещества клетки Страус
Страус Красная книга
Красная книга Птицы. Отличительные признаки птиц
Птицы. Отличительные признаки птиц Тектология А.А. Богданова
Тектология А.А. Богданова Экстерьер задних конечностей
Экстерьер задних конечностей