Фізіологія м’язів. Фізіологічні основи рухової активності. Нейро-гуморальна регуляція м’язової діяльності
- Фізіологія м’язів. Фізіологічні основи рухової активності. Нейро-гуморальна регуляція м’язової діяльності

Содержание
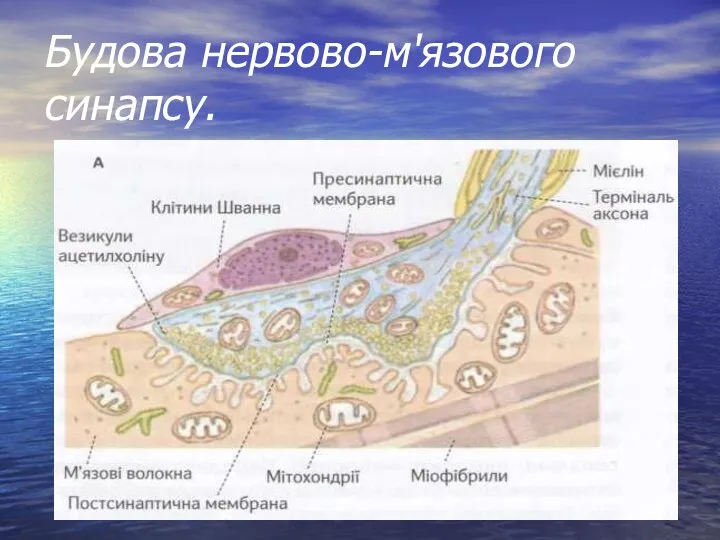
- 2. Будова нервово-м'язового синапсу.
- 3. Особливості проведення збудження через синапси: Однобічність проведення збудження. В аксоні збудження проходить в обох напрямках від
- 4. Сумація скорочень і тетанус м'яза У відповідь на одинокий імпульс відбувається скорочення, за яким спостерігається дещо
- 5. Електроміографія - це метод реєстрації потенціалів дії, що виникають у м’язах. При відведенні поверхневими електродами розрізняють
- 6. Одинокі скорочення (а), зубчатий (б) і гладкий (в) тетанус. Зубчастий тетанус виникає при такій частоті, коли
- 7. Ядра довгастого мозку. 1) рухові, 2) чутливі, 3) змішані та 4) ретикулярної формації. Рухові: ядро підязикового
- 8. Ядра моста. Вони поділяються на 1) рухові, 2) змішані та 3) ядра ретикулярної формації. Рухові: ядро
- 10. Середній мозок Середній мозок – це чотиригорбкове тіло і ніжки мозку. Ядра середнього мозку. Сіра речовина
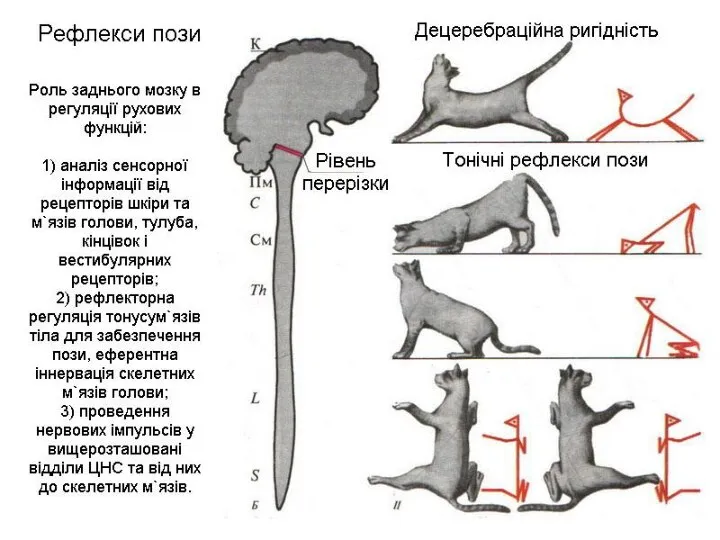
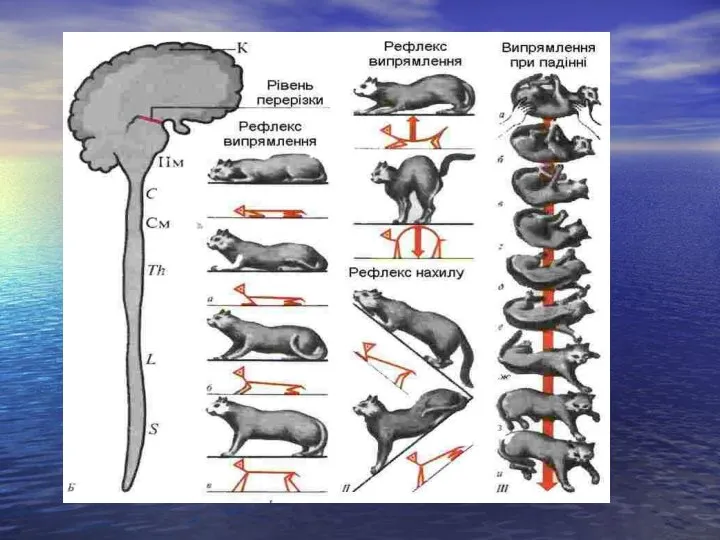
- 11. Рефлекторна функція середнього мозку. А. Середній мозок забезпечує здійснення орієнтувальних рефлексів: 1) зорових 2) слухових. Б.
- 13. ФУНКЦІЇ БАЗАЛЬНИХ ЯДЕР Основною функцією базальних ядер є планування та програмування рухових команд разом з корою
- 14. Мозочок за допомогою нейронних ланцюгів бере участь у регуляції і корекції рухових функцій, а саме: підтриманні
- 16. Роль спинного мозку в регуляції автономних функцій У перших п'яти грудних сегментах спинного мозку локалізовані прегангліонарні
- 21. Щитоподібна залоза Йодовмісні гормони: тироксин або тетрайодтиронін і трийодтиронін та тиреокальцитонін. Метаболічний ефект. Гормони викликають індукцію
- 22. Паращитоподібні залози Паратгормон. 1. Стимулює активність остеобластів, що приводить до звільнення іонів кальцію і фосфору з
- 23. Підшлункова залоза
- 24. Підшлункова залоза Острівцеві клітини, синтезують гормони інсулін, глюкагон і соматостатин. Клітини, що синтезують інсулін, називають β-клітинами,
- 25. Метаболічна дія інсуліну Вплив інсуліну на вуглеводний обмін. 1. Збільшує проникність клітинних мембран для глюкози. 2.
- 26. Ефекти глюкагону. Діє на печінку, де: 1. Підсилює розщеплення глікогену. 2. Сприяє глюконеогенезу. 3. Стимулює ліполіз.
- 27. Гормони статевих залоз Чоловічі статеві гормони – андрогени (тестостерон), Жіночі – естрогени, прогестини (прогестерон). У період
- 28. Епіфіз Містить велику кількість біологічно активних речовин. Найбільше серед них серотоніну, меланіну і адреногломерулотропіну. Виділення мелатоніну
- 29. Загрудинна залоза.
- 30. Загрудинна залоза. Протягом останніх років з загрудинної залози були виділені тимозин, тимопоетин і інші поліпептиди, що
- 32. Скачать презентацию
Слайд 3Особливості проведення збудження через синапси:
Однобічність проведення збудження. В аксоні збудження проходить в
Особливості проведення збудження через синапси:
Однобічність проведення збудження. В аксоні збудження проходить в

Слайд 4Сумація скорочень і тетанус м'яза
У відповідь на одинокий імпульс відбувається скорочення, за
Сумація скорочень і тетанус м'яза
У відповідь на одинокий імпульс відбувається скорочення, за

Слайд 5Електроміографія - це метод реєстрації потенціалів дії, що виникають у м’язах.
При відведенні
Електроміографія - це метод реєстрації потенціалів дії, що виникають у м’язах.
При відведенні

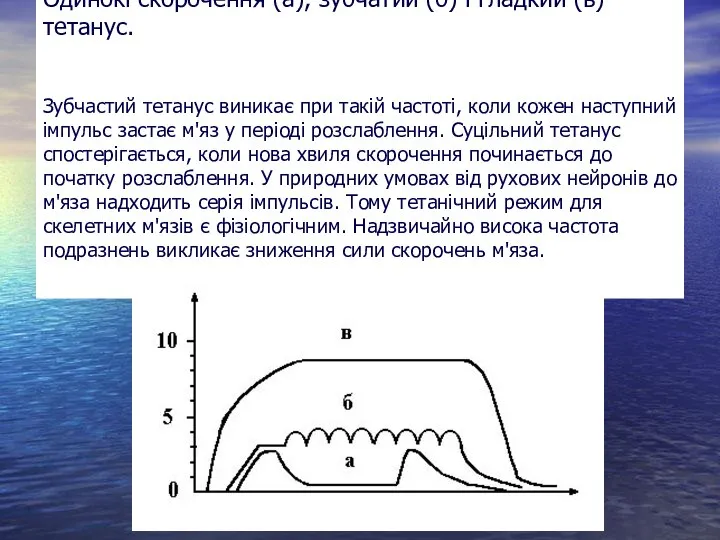
Слайд 6Одинокі скорочення (а), зубчатий (б) і гладкий (в) тетанус.
Зубчастий тетанус виникає при
Одинокі скорочення (а), зубчатий (б) і гладкий (в) тетанус. Зубчастий тетанус виникає при
Слайд 7Ядра довгастого мозку.
1) рухові, 2) чутливі, 3) змішані та 4) ретикулярної
Ядра довгастого мозку.
1) рухові, 2) чутливі, 3) змішані та 4) ретикулярної

Слайд 8Ядра моста.
Вони поділяються на 1) рухові, 2) змішані та 3) ядра ретикулярної
Ядра моста.
Вони поділяються на 1) рухові, 2) змішані та 3) ядра ретикулярної

Слайд 10Середній мозок
Середній мозок – це чотиригорбкове тіло і ніжки мозку.
Ядра середнього
Середній мозок
Середній мозок – це чотиригорбкове тіло і ніжки мозку.
Ядра середнього

Слайд 11Рефлекторна функція середнього мозку.
А. Середній мозок забезпечує здійснення орієнтувальних рефлексів:
1)
Рефлекторна функція середнього мозку.
А. Середній мозок забезпечує здійснення орієнтувальних рефлексів:
1)

Слайд 13ФУНКЦІЇ БАЗАЛЬНИХ ЯДЕР
Основною функцією базальних ядер є планування та програмування рухових
ФУНКЦІЇ БАЗАЛЬНИХ ЯДЕР
Основною функцією базальних ядер є планування та програмування рухових

Слайд 14Мозочок за допомогою нейронних ланцюгів бере участь у регуляції і корекції рухових
Мозочок за допомогою нейронних ланцюгів бере участь у регуляції і корекції рухових

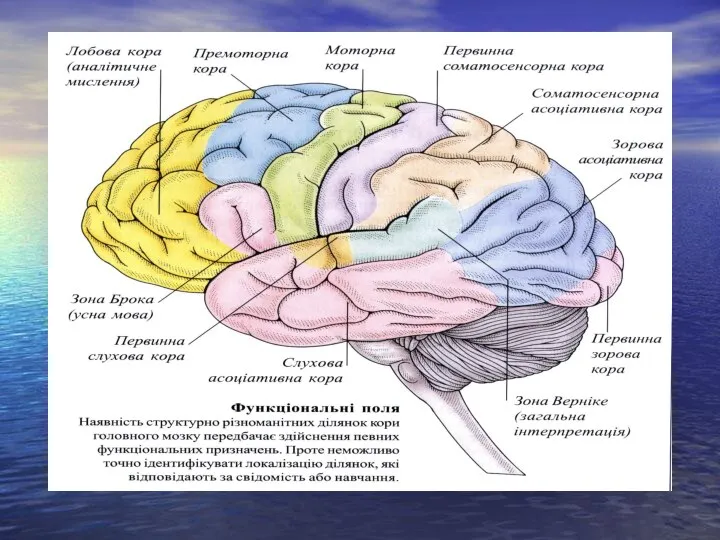
Слайд 16Роль спинного мозку в регуляції автономних функцій
У перших п'яти грудних сегментах спинного
Роль спинного мозку в регуляції автономних функцій
У перших п'яти грудних сегментах спинного


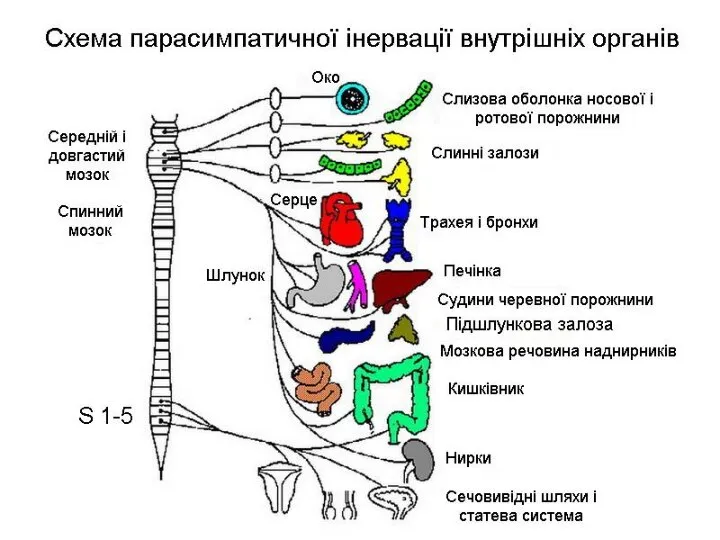

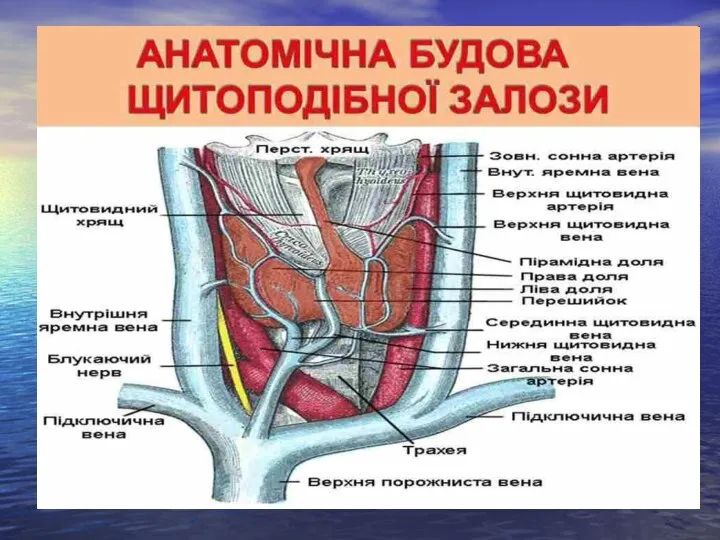
Слайд 21Щитоподібна залоза
Йодовмісні гормони: тироксин або тетрайодтиронін і трийодтиронін та тиреокальцитонін.
Метаболічний ефект.
Щитоподібна залоза
Йодовмісні гормони: тироксин або тетрайодтиронін і трийодтиронін та тиреокальцитонін.
Метаболічний ефект.

Слайд 22Паращитоподібні залози
Паратгормон.
1. Стимулює активність остеобластів, що приводить до звільнення іонів кальцію
Паращитоподібні залози
Паратгормон.
1. Стимулює активність остеобластів, що приводить до звільнення іонів кальцію

Слайд 23 Підшлункова залоза
Підшлункова залоза
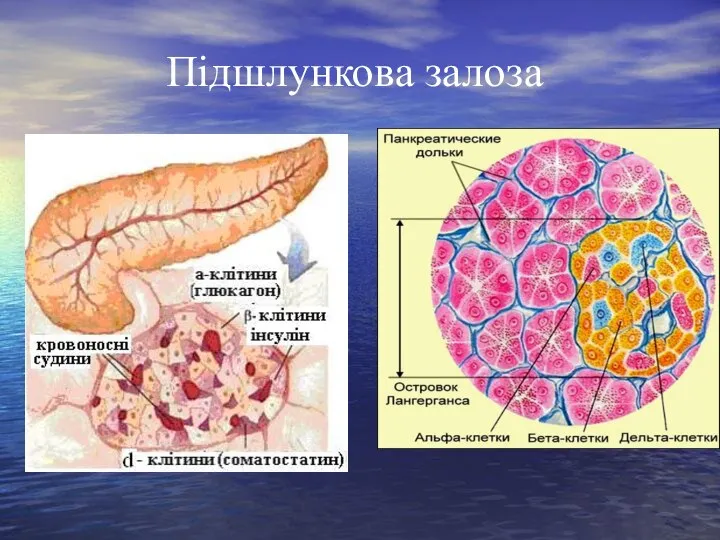
Слайд 24Підшлункова залоза
Острівцеві клітини, синтезують гормони інсулін, глюкагон і соматостатин.
Клітини, що синтезують
Підшлункова залоза
Острівцеві клітини, синтезують гормони інсулін, глюкагон і соматостатин.
Клітини, що синтезують

Слайд 25Метаболічна дія інсуліну
Вплив інсуліну на вуглеводний обмін.
1. Збільшує проникність клітинних мембран для
Метаболічна дія інсуліну
Вплив інсуліну на вуглеводний обмін.
1. Збільшує проникність клітинних мембран для

Слайд 26Ефекти глюкагону.
Діє на печінку, де:
1. Підсилює розщеплення глікогену.
2. Сприяє глюконеогенезу.
3. Стимулює ліполіз.
4.
Ефекти глюкагону.
Діє на печінку, де:
1. Підсилює розщеплення глікогену.
2. Сприяє глюконеогенезу.
3. Стимулює ліполіз.
4.

Слайд 27Гормони статевих залоз
Чоловічі статеві гормони – андрогени (тестостерон),
Жіночі – естрогени, прогестини
Гормони статевих залоз
Чоловічі статеві гормони – андрогени (тестостерон),
Жіночі – естрогени, прогестини

Слайд 28Епіфіз
Містить велику кількість біологічно активних речовин. Найбільше серед них серотоніну, меланіну і
Епіфіз
Містить велику кількість біологічно активних речовин. Найбільше серед них серотоніну, меланіну і

Слайд 29Загрудинна залоза.
Загрудинна залоза.
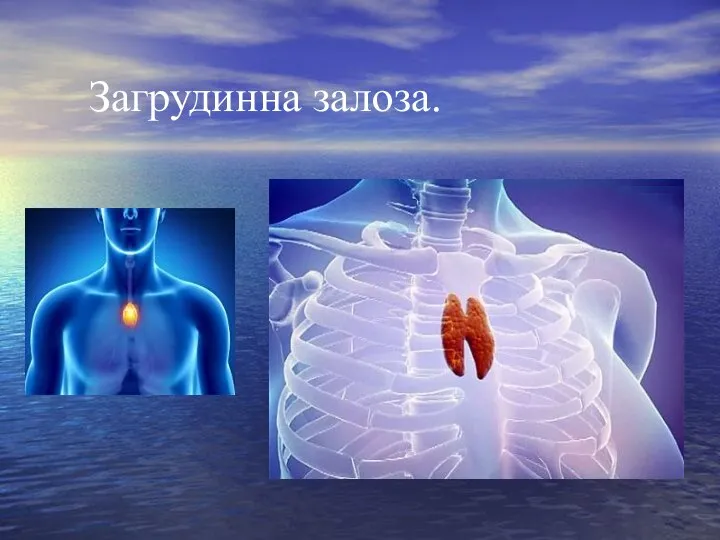
Слайд 30Загрудинна залоза.
Протягом останніх років з загрудинної залози були виділені тимозин, тимопоетин і
Загрудинна залоза.
Протягом останніх років з загрудинної залози були виділені тимозин, тимопоетин і

 Пищеварительная система
Пищеварительная система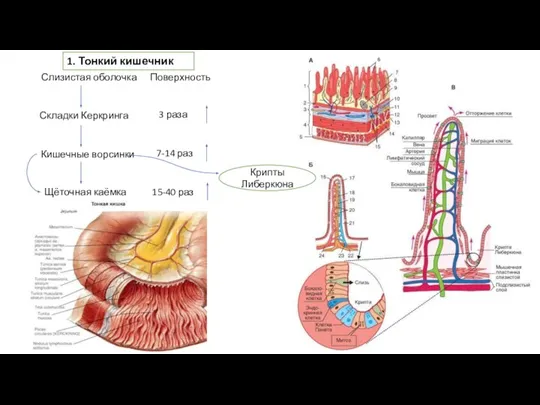 Тонкий кишечник
Тонкий кишечник Ткани человека. 1
Ткани человека. 1 Нервная система человека
Нервная система человека Мариукультура. Культивирование тихоокеанских устриц
Мариукультура. Культивирование тихоокеанских устриц Особенности выращивания в домашних условиях фазанов ( из опыта работы)
Особенности выращивания в домашних условиях фазанов ( из опыта работы) Презентация на тему Мечников Илья Ильич
Презентация на тему Мечников Илья Ильич  Таламус (горизонтальный срез)
Таламус (горизонтальный срез) Голосовой аппарат человека. Занятие 6
Голосовой аппарат человека. Занятие 6 Чарлз Дарвин и его эволюционное учение
Чарлз Дарвин и его эволюционное учение Презентация на тему Гаметогенез у растений
Презентация на тему Гаметогенез у растений  Углеводы. Дисахариды. Важнейшие представители
Углеводы. Дисахариды. Важнейшие представители Химическая организация клетки
Химическая организация клетки Презентация на тему Влияние школьного портфеля на осанку
Презентация на тему Влияние школьного портфеля на осанку  Тип кольчатые черви. (Часть 2)
Тип кольчатые черви. (Часть 2) Биосфера. Тест
Биосфера. Тест Репарационные процессы ДНК. (Лекция 3)
Репарационные процессы ДНК. (Лекция 3) Оксид азота и его физиологическая роль
Оксид азота и его физиологическая роль Классификация пород собак
Классификация пород собак Причины и последствия нарушений липидного обмена, гиперхолестеринемии
Причины и последствия нарушений липидного обмена, гиперхолестеринемии Осьминоги. Размеры осьминогов
Осьминоги. Размеры осьминогов Как живут растения. Окружающий мир (1 класс)
Как живут растения. Окружающий мир (1 класс) Презентация на тему Лекарственные растения Викторина
Презентация на тему Лекарственные растения Викторина  Особенности внешнего строения птиц
Особенности внешнего строения птиц Фенотипик үзгәрүчәнлек
Фенотипик үзгәрүчәнлек Алекс Флемінг
Алекс Флемінг Внешнее строение птиц
Внешнее строение птиц Intention. Precuneus
Intention. Precuneus