- Генетический анализ

Содержание
- 2. Генетические задачи решаются легко только тогда, когда они предварительно уже решены другими. Поэтому необходимо предостеречь тех,
- 3. 1.Развитие представлений о гене как о единице мутации, рекомбинации и функции. Опыты С. Бензера по картированию
- 4. Т е о р и я г е н а
- 5. Ген – это структурная единица наследственной информации далее неделимая в функциональном отношении. Впервые термин «ген» ввел
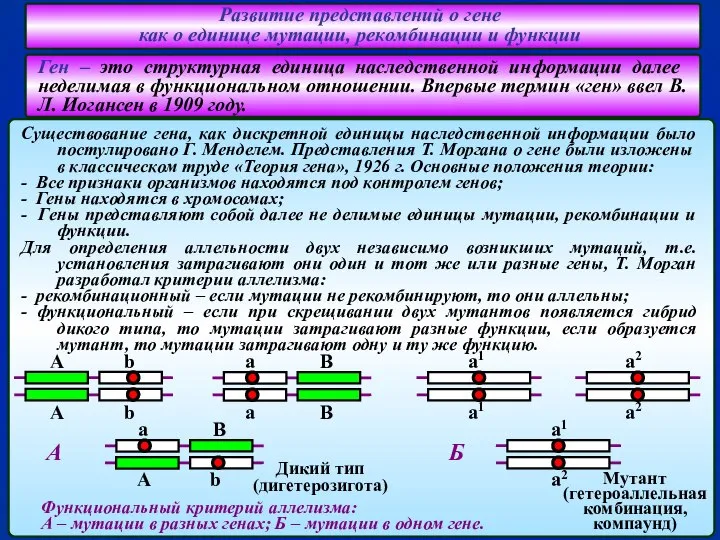
- 6. Развитие представлений о гене как о единице мутации, рекомбинации и функции А.С. Серебровский и его сотрудники
- 7. Развитие представлений о гене как о единице мутации, рекомбинации и функции В 40-х годах прошлого века
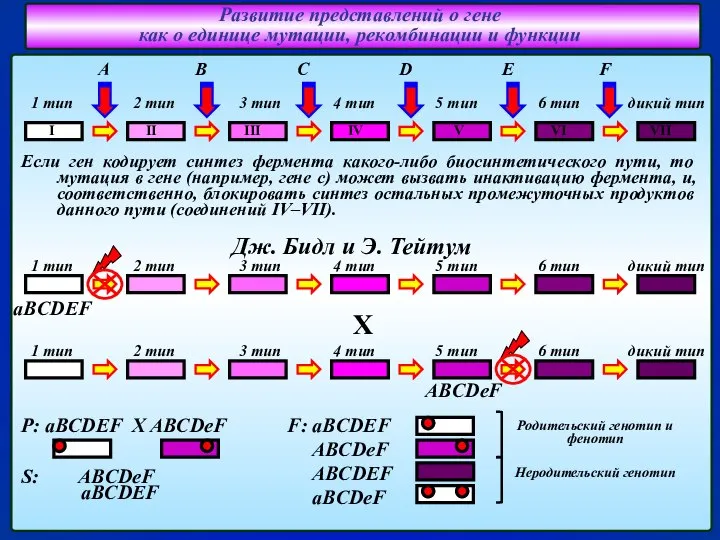
- 8. Развитие представлений о гене как о единице мутации, рекомбинации и функции Если ген кодирует синтез фермента
- 9. Развитие представлений о гене как о единице мутации, рекомбинации и функции Опыты С. Бензера по картированию
- 10. В таблице приведены результаты теста на комплементарность для десяти точковых мутаций. «+» – комплементация мутации; «-»
- 11. Для точной локализации мутации Бензер использовал не только скрещивание точковых мутантов между собой, но и метод
- 12. Опыты С. Бензера по картированию мутаций. Цис- транс- тест С. Бензер попытался ревизовать и понятие «ген».
- 13. Генные мутации
- 14. Генные мутации Генные мутации – это наследственно закрепленные изменения генетического материала на уровне отдельных генов. Используют:
- 15. Генные мутации Изучение генных мутаций может осуществляться на трех уровнях: 1 – на молекулярном уровне (на
- 16. Типы мутаци, вызываемых мутагенами При анализе генных мутаций на уровне ДНК необходимо помнить, что повреждение нуклеотидов
- 17. Молекулярные механизмы возникновения генных мутаций
- 18. Типы генных (точечных) мутаций Вставка, выпадение пары нуклеотидов – мутации, при которых происходит количественное изменение числа
- 19. Механизм мутантного действия 5-бромурацила Спаривание аденина (А) с 5-бромурацилом (BrU) Спаривание гуанина (G) с 5-бромурацилом (BrU)
- 20. Механизм мутантного действия 5-бромурацила ошибка включения (G/C→A/T) ошибка репликации (A/T→G/C) A || T A || BrU
- 21. Механизм мутантного действия 2-аминопурина Существуют два механизма, приводящие к транзициям: ошибка репликации (A/T→G/C) 2-аминопурин в аминоформе
- 22. Механизм мутантного действия 2-аминорурина ошибка включения (G/C→A/T) ошибка репликации (A/T→G/C) A || T AP || T
- 23. Механизм мутантного действия азотистой кислоты Спаривание гипоксантина (Hx) с цитозином (C) Спаривание урацила (U) с аденином
- 24. Причины спонтанных мутаций Апуринизация – потеря нуклеотидом пуринового основания вследствие разрыва фосфодиэфирной связи между основанием и
- 25. Характер проявления генных мутаций По характеру проявления на уровне белкового продукта генные мутации классифицируют как миссенс
- 26. Методы выявления генных мутаций Выявление мутационных изменений связано с рядом трудностей, обусловленных следующими факторами: большинство мутаций
- 27. Выявление рецессивнных летальных и видимых мутаций, возникающих в Х хромосоме зрелых сперматозоидов самцов дрозофилы Метод Меллер-5
- 28. Выявление рецессивнных летальных и видимых мутаций, возникающих в Х хромосоме зрелых сперматозоидов самцов дрозофилы Метод ClB
- 29. Учет генных мутаций При проведении количественного учета генных мутаций используют два основных приема: определение частоту появления
- 30. Геномные мутации
- 31. Типы геномных мутаций Геномные мутации – мутации, изменяющие структуру и состав генома в целом, т.е. изменение
- 32. Типы геномных мутаций Анеуплоиды у диплоидных видов бывают следующих типов: - Моносомики (2п-1), при этом отсутствует
- 33. Типы геномных мутаций. Автополиплоидия. У автополиплоидов каждая хромосома набора представлена гомологичными хромосомами в количестве, соответствующем уровню
- 34. Геномные мутаци: закономерности образования гамет У автополиплоидов и аллополиплоидов отмечены случаи предпочтительной (избирательной) бивалентной конъюгации хромосом
- 35. Геномные мутаци: закономерности образования гамет Возможно и другое схематическое изображение генотипа тетраплоида, образование у него гамет
- 36. Генетика популяций
- 37. Закон Г. Харди и В. Вайнберга Популяция – это совокупность особей одного вида свободно скрещивающихся между
- 38. Следствия, вытекающие из закона Харди-Вайнберга Частоты аллелей не изменяются от поколения к поколению. Докозательство: Частота аллеля
- 39. Следствия, вытекающие из закона Харди-Вайнберга Частоты аллелей не изменяются от поколения к поколению. Докозательство: Частота аллеля
- 40. Условия выполнения и применения закона Харди-Вайнберга Закон Харди-Вайнберга применим для популяции, на которые не действуют внешние
- 41. Структура популяции самоопылителей AA = 2n(2KAA + KAa) – KAa aa = 2n(2Kaa + KAa) –
- 42. Естественный отбор s (коэф. отбора)= 1 - w Относительная приспособленность указывает на направление отбора, но не
- 43. Отбор против гетерозигот (p=q) Δq = 0, если p = q (при w(AA) = w (aa)).
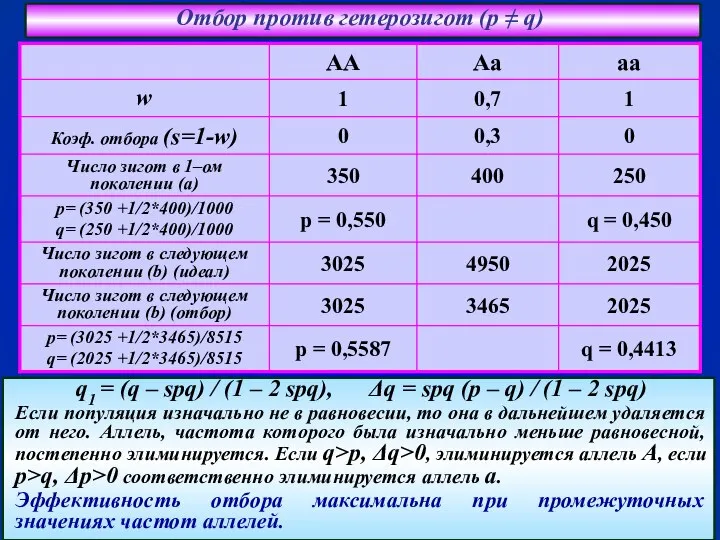
- 44. Отбор против гетерозигот (p ≠ q) q1 = (q – spq) / (1 – 2 spq),
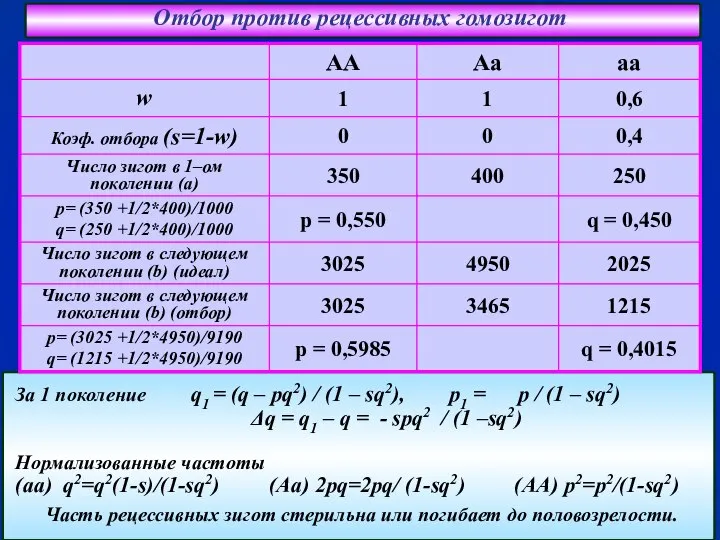
- 45. Отбор против рецессивных гомозигот За 1 поколение q1 = (q – pq2) / (1 – sq2),
- 47. Скачать презентацию

Слайд 31.Развитие представлений о гене как о единице мутации, рекомбинации и функции.
Опыты С.
1.Развитие представлений о гене как о единице мутации, рекомбинации и функции.
Опыты С.

Слайд 4Т е о р и я
г е н а
Т е о р и я
г е н а

Слайд 5Ген – это структурная единица наследственной информации далее неделимая в функциональном отношении.
Ген – это структурная единица наследственной информации далее неделимая в функциональном отношении.
Слайд 6Развитие представлений о гене
как о единице мутации, рекомбинации и функции
А.С. Серебровский
Развитие представлений о гене
как о единице мутации, рекомбинации и функции
А.С. Серебровский

Слайд 7Развитие представлений о гене
как о единице мутации, рекомбинации и функции
В 40-х
Развитие представлений о гене
как о единице мутации, рекомбинации и функции
В 40-х

Слайд 8Развитие представлений о гене
как о единице мутации, рекомбинации и функции
Если ген
Развитие представлений о гене
как о единице мутации, рекомбинации и функции
Если ген
Слайд 9Развитие представлений о гене
как о единице мутации, рекомбинации и функции
Опыты С.
Развитие представлений о гене
как о единице мутации, рекомбинации и функции
Опыты С.

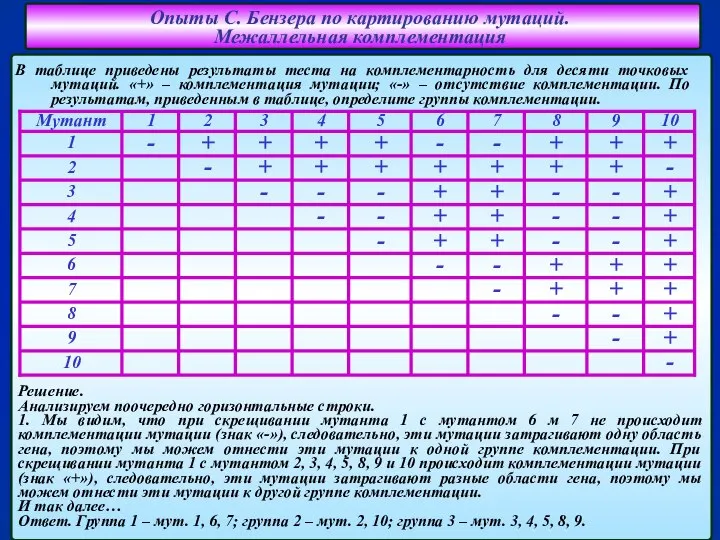
Слайд 10В таблице приведены результаты теста на комплементарность для десяти точковых мутаций. «+»
В таблице приведены результаты теста на комплементарность для десяти точковых мутаций. «+»
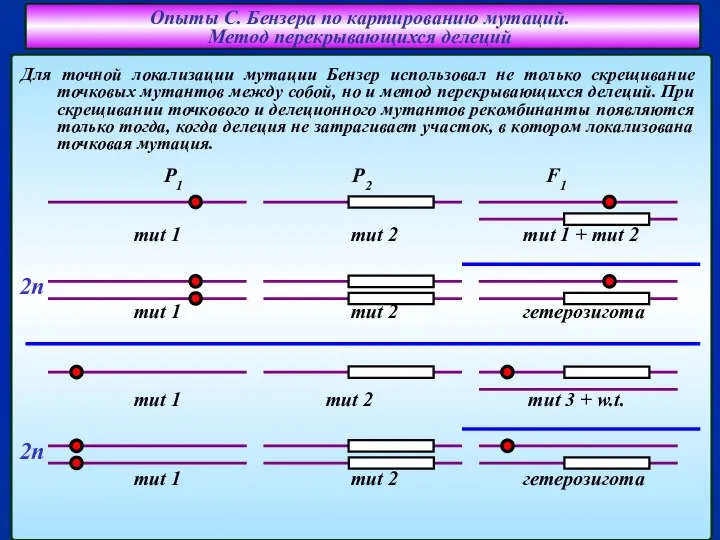
Слайд 11Для точной локализации мутации Бензер использовал не только скрещивание точковых мутантов между
Для точной локализации мутации Бензер использовал не только скрещивание точковых мутантов между
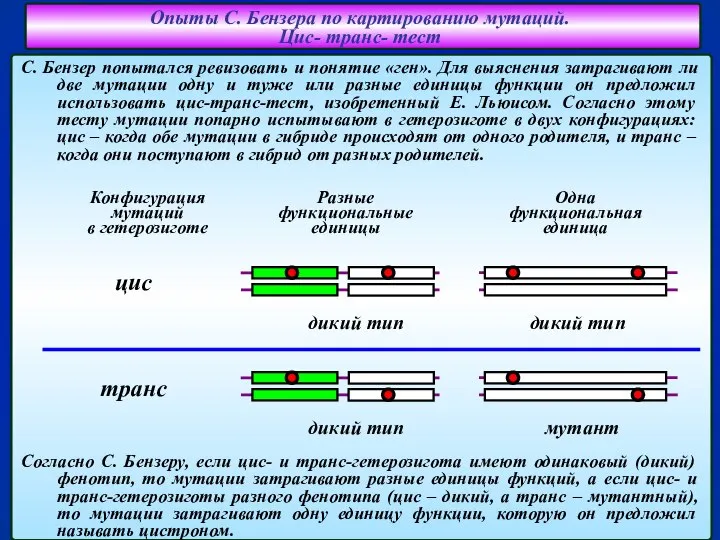
Слайд 12Опыты С. Бензера по картированию мутаций.
Цис- транс- тест
С. Бензер попытался ревизовать
Опыты С. Бензера по картированию мутаций.
Цис- транс- тест
С. Бензер попытался ревизовать
Слайд 13Генные
мутации
Генные
мутации

Слайд 14Генные мутации
Генные мутации – это наследственно закрепленные изменения генетического материала на уровне
Генные мутации
Генные мутации – это наследственно закрепленные изменения генетического материала на уровне

Слайд 15Генные мутации
Изучение генных мутаций может осуществляться на трех уровнях:
1 – на
Генные мутации
Изучение генных мутаций может осуществляться на трех уровнях:
1 – на

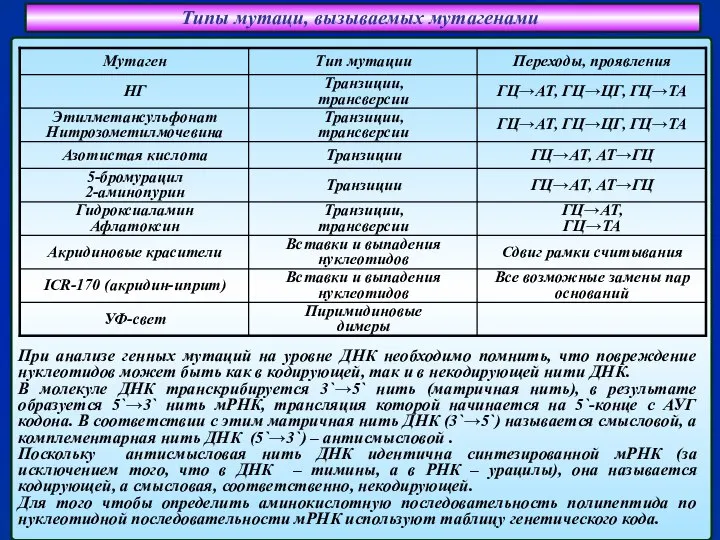
Слайд 16Типы мутаци, вызываемых мутагенами
При анализе генных мутаций на уровне ДНК необходимо помнить,
Типы мутаци, вызываемых мутагенами
При анализе генных мутаций на уровне ДНК необходимо помнить,
Слайд 17Молекулярные механизмы
возникновения
генных мутаций
Молекулярные механизмы
возникновения
генных мутаций

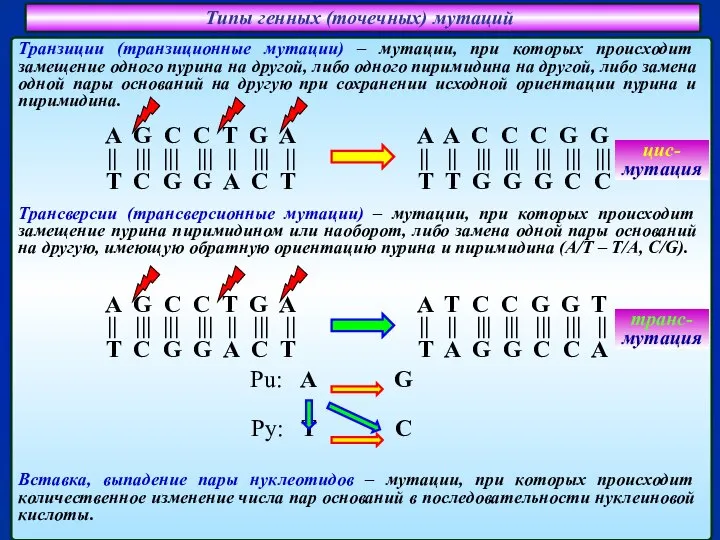
Слайд 18Типы генных (точечных) мутаций
Вставка, выпадение пары нуклеотидов – мутации, при которых происходит
Типы генных (точечных) мутаций
Вставка, выпадение пары нуклеотидов – мутации, при которых происходит
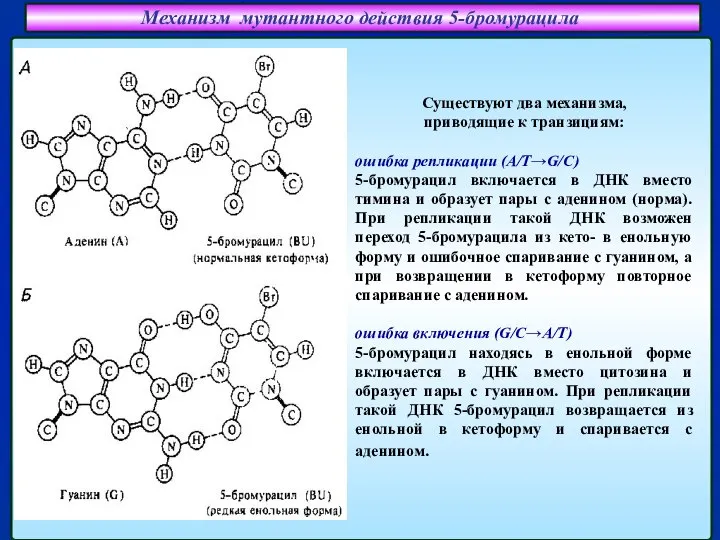
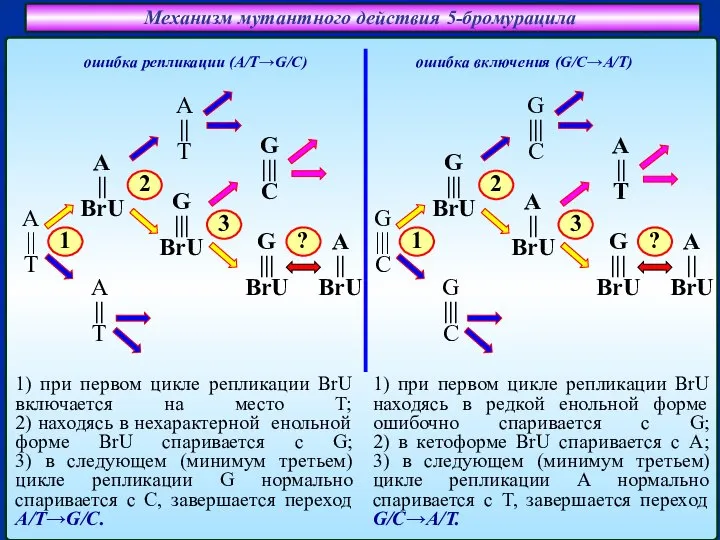
Слайд 19Механизм мутантного действия 5-бромурацила
Спаривание аденина (А) с 5-бромурацилом (BrU)
Спаривание гуанина (G) с
Механизм мутантного действия 5-бромурацила
Спаривание аденина (А) с 5-бромурацилом (BrU)
Спаривание гуанина (G) с
Слайд 20Механизм мутантного действия 5-бромурацила
ошибка включения (G/C→A/T)
ошибка репликации (A/T→G/C)
A
||
T
A
||
BrU
A
||
T
A
||
T
G
|||
BrU
G
|||
C
G
|||
BrU
A
||
BrU
G
|||
C
G
|||
BrU
G
|||
C
G
|||
C
A
||
BrU
A
||
T
G
|||
BrU
A
||
BrU
1) при первом цикле репликации
Механизм мутантного действия 5-бромурацила
ошибка включения (G/C→A/T)
ошибка репликации (A/T→G/C)
A
||
T
A
||
BrU
A
||
T
A
||
T
G
|||
BrU
G
|||
C
G
|||
BrU
A
||
BrU
G
|||
C
G
|||
BrU
G
|||
C
G
|||
C
A
||
BrU
A
||
T
G
|||
BrU
A
||
BrU
1) при первом цикле репликации
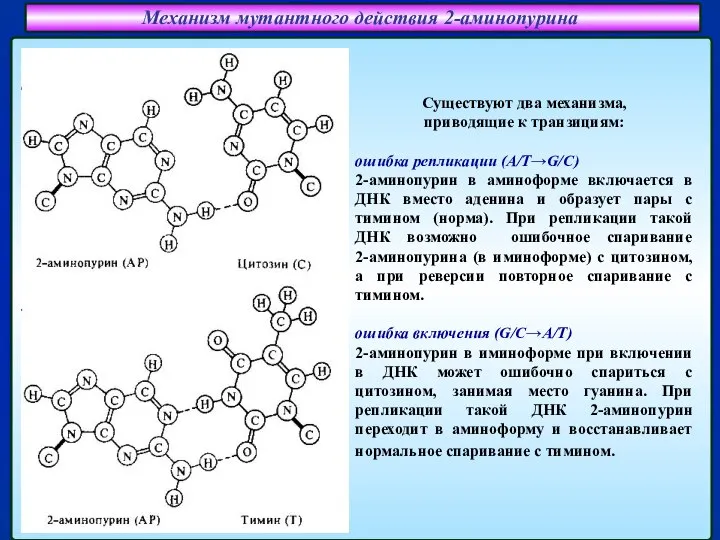
Слайд 21Механизм мутантного действия 2-аминопурина
Существуют два механизма,
приводящие к транзициям:
ошибка репликации (A/T→G/C)
2-аминопурин
Механизм мутантного действия 2-аминопурина
Существуют два механизма,
приводящие к транзициям:
ошибка репликации (A/T→G/C)
2-аминопурин
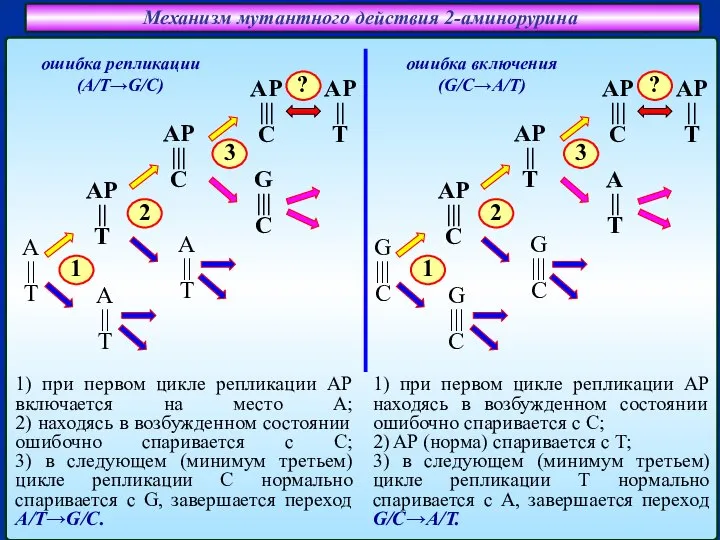
Слайд 22Механизм мутантного действия 2-аминорурина
ошибка включения
(G/C→A/T)
ошибка репликации
(A/T→G/C)
A
||
T
AP
||
T
A
||
T
AP
|||
C
A
||
T
G
|||
C
AP
|||
C
AP
||
T
1) при первом цикле репликации
Механизм мутантного действия 2-аминорурина
ошибка включения
(G/C→A/T)
ошибка репликации
(A/T→G/C)
A
||
T
AP
||
T
A
||
T
AP
|||
C
A
||
T
G
|||
C
AP
|||
C
AP
||
T
1) при первом цикле репликации
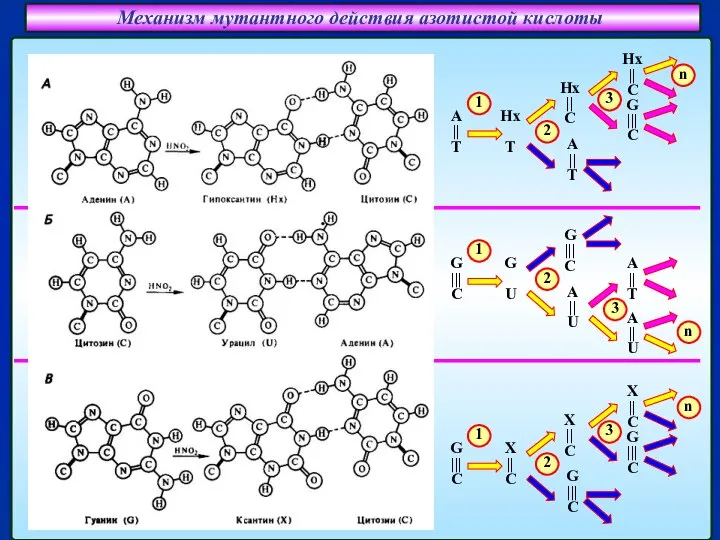
Слайд 23Механизм мутантного действия азотистой кислоты
Спаривание гипоксантина (Hx)
с цитозином (C)
Спаривание урацила (U)
Механизм мутантного действия азотистой кислоты
Спаривание гипоксантина (Hx)
с цитозином (C)
Спаривание урацила (U)
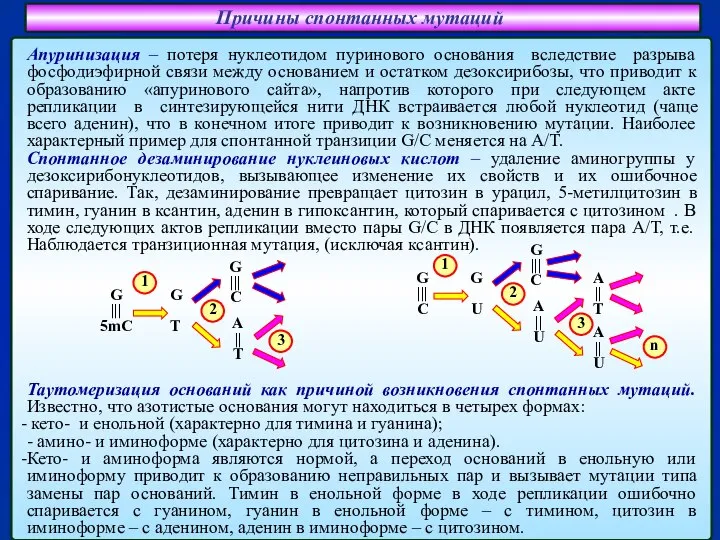
Слайд 24Причины спонтанных мутаций
Апуринизация – потеря нуклеотидом пуринового основания вследствие разрыва фосфодиэфирной связи
Причины спонтанных мутаций
Апуринизация – потеря нуклеотидом пуринового основания вследствие разрыва фосфодиэфирной связи
Слайд 25Характер проявления генных мутаций
По характеру проявления на уровне белкового продукта генные мутации
Характер проявления генных мутаций
По характеру проявления на уровне белкового продукта генные мутации

Слайд 26Методы выявления генных мутаций
Выявление мутационных изменений связано с рядом трудностей, обусловленных следующими
Методы выявления генных мутаций
Выявление мутационных изменений связано с рядом трудностей, обусловленных следующими

Слайд 27Выявление рецессивнных летальных и видимых мутаций,
возникающих в Х хромосоме зрелых сперматозоидов
Выявление рецессивнных летальных и видимых мутаций, возникающих в Х хромосоме зрелых сперматозоидов

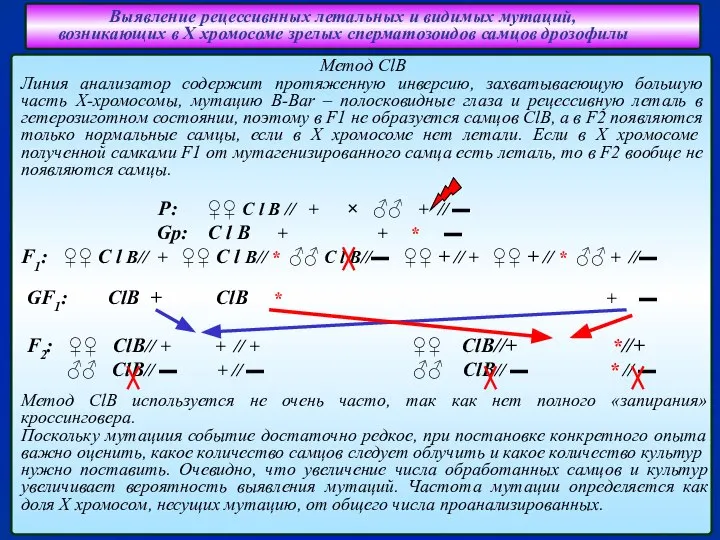
Слайд 28Выявление рецессивнных летальных и видимых мутаций,
возникающих в Х хромосоме зрелых сперматозоидов
Выявление рецессивнных летальных и видимых мутаций, возникающих в Х хромосоме зрелых сперматозоидов
Слайд 29Учет генных мутаций
При проведении количественного учета генных мутаций используют два основных приема:
Учет генных мутаций
При проведении количественного учета генных мутаций используют два основных приема:

Слайд 30Геномные
мутации
Геномные
мутации

Слайд 31Типы геномных мутаций
Геномные мутации – мутации, изменяющие структуру и состав генома в
Типы геномных мутаций
Геномные мутации – мутации, изменяющие структуру и состав генома в

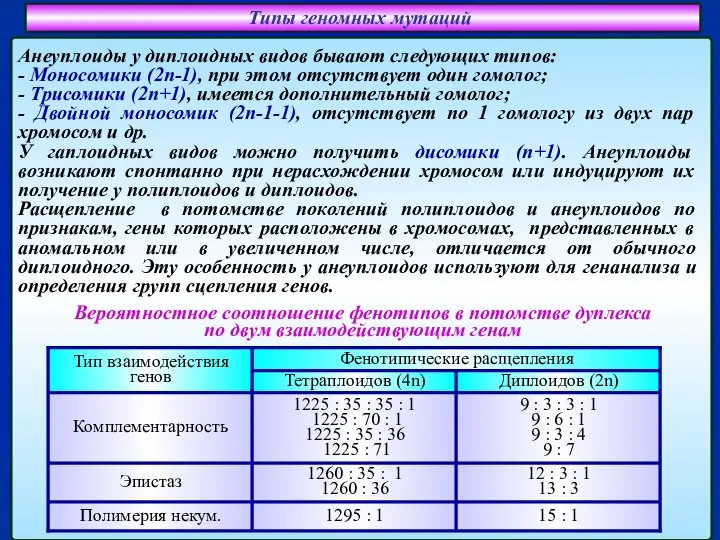
Слайд 32Типы геномных мутаций
Анеуплоиды у диплоидных видов бывают следующих типов:
- Моносомики (2п-1), при
Типы геномных мутаций
Анеуплоиды у диплоидных видов бывают следующих типов:
- Моносомики (2п-1), при
Слайд 33Типы геномных мутаций. Автополиплоидия.
У автополиплоидов каждая хромосома набора представлена гомологичными хромосомами в
Типы геномных мутаций. Автополиплоидия.
У автополиплоидов каждая хромосома набора представлена гомологичными хромосомами в

Слайд 34Геномные мутаци: закономерности образования гамет
У автополиплоидов и аллополиплоидов отмечены случаи предпочтительной (избирательной)
Геномные мутаци: закономерности образования гамет
У автополиплоидов и аллополиплоидов отмечены случаи предпочтительной (избирательной)

Слайд 35Геномные мутаци: закономерности образования гамет
Возможно и другое схематическое изображение генотипа тетраплоида, образование
Геномные мутаци: закономерности образования гамет
Возможно и другое схематическое изображение генотипа тетраплоида, образование

Слайд 36Генетика
популяций
Генетика
популяций

Слайд 37Закон Г. Харди и В. Вайнберга
Популяция – это совокупность особей одного вида
Закон Г. Харди и В. Вайнберга
Популяция – это совокупность особей одного вида

Слайд 38Следствия, вытекающие из закона Харди-Вайнберга
Частоты аллелей не изменяются от поколения к
Следствия, вытекающие из закона Харди-Вайнберга
Частоты аллелей не изменяются от поколения к

Слайд 39Следствия, вытекающие из закона Харди-Вайнберга
Частоты аллелей не изменяются от поколения к
Следствия, вытекающие из закона Харди-Вайнберга
Частоты аллелей не изменяются от поколения к

Слайд 40Условия выполнения и применения
закона Харди-Вайнберга
Закон Харди-Вайнберга применим для популяции, на
Условия выполнения и применения
закона Харди-Вайнберга
Закон Харди-Вайнберга применим для популяции, на

Слайд 41Структура популяции самоопылителей
AA = 2n(2KAA + KAa) – KAa
aa = 2n(2Kaa +
Структура популяции самоопылителей
AA = 2n(2KAA + KAa) – KAa
aa = 2n(2Kaa +

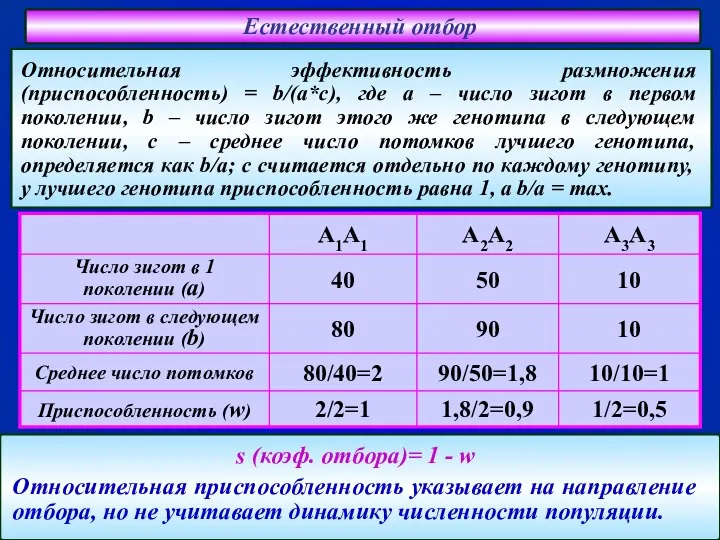
Слайд 42Естественный отбор
s (коэф. отбора)= 1 - w
Относительная приспособленность указывает на направление
Естественный отбор
s (коэф. отбора)= 1 - w
Относительная приспособленность указывает на направление
Слайд 43Отбор против гетерозигот (p=q)
Δq = 0,
если p = q (при w(AA)
Отбор против гетерозигот (p=q)
Δq = 0,
если p = q (при w(AA)

Слайд 44Отбор против гетерозигот (p ≠ q)
q1 = (q – spq) / (1
Отбор против гетерозигот (p ≠ q)
q1 = (q – spq) / (1
Слайд 45Отбор против рецессивных гомозигот
За 1 поколение q1 = (q – pq2) /
Отбор против рецессивных гомозигот
За 1 поколение q1 = (q – pq2) /
 Промежуточный мозг (diencephalon)
Промежуточный мозг (diencephalon) Предмет патофизиологии
Предмет патофизиологии Американский стаффордширский терьер
Американский стаффордширский терьер Строение и функции зубов
Строение и функции зубов Дигибридное скрещивание
Дигибридное скрещивание Коза – это звучит гордо!
Коза – это звучит гордо! Размножение и развитие организмов
Размножение и развитие организмов Рыбы. Характеристика класс рыб
Рыбы. Характеристика класс рыб Практикум по решению генетических задач
Практикум по решению генетических задач Гуморальная регуляция
Гуморальная регуляция Теория разумного замысла или дарвинизм. За и против
Теория разумного замысла или дарвинизм. За и против Пищеварительная система
Пищеварительная система Витамины и их роль в организме человека
Витамины и их роль в организме человека Genetik
Genetik Строение организма человека
Строение организма человека Строение мышц
Строение мышц Викторина. 10 класс
Викторина. 10 класс Презентация на тему ОРГАНЫ ЧУВСТВ, ИХ ЗНАЧЕНИЕ И ГИГИЕНА
Презентация на тему ОРГАНЫ ЧУВСТВ, ИХ ЗНАЧЕНИЕ И ГИГИЕНА  Будьмо знайомі. Мурахоїд
Будьмо знайомі. Мурахоїд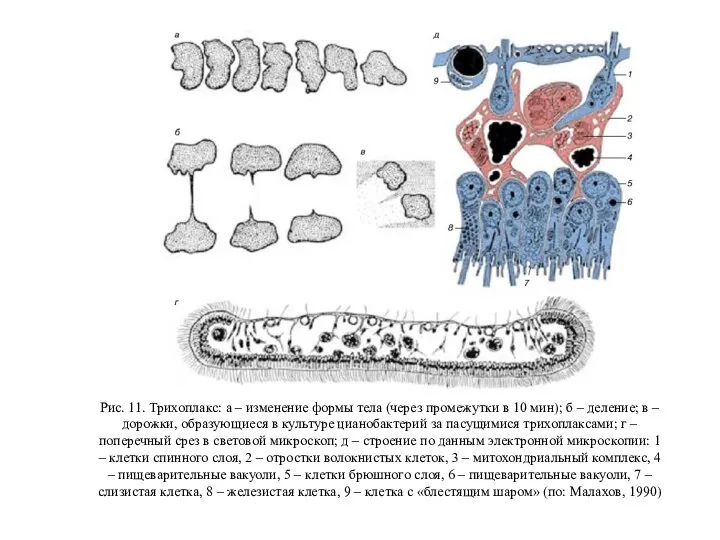 Трихоплакс. Лекция 2
Трихоплакс. Лекция 2 Голубая кровь среди живых существ
Голубая кровь среди живых существ Свекла. Полезно или нет
Свекла. Полезно или нет Значение и строение выделительной системы человека
Значение и строение выделительной системы человека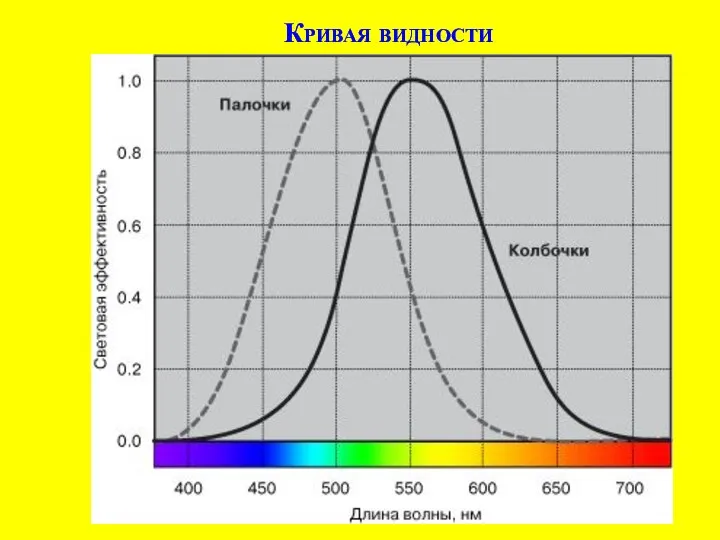 Г. фон Гельмгольц (1821 – 1894) - цветовое зрение
Г. фон Гельмгольц (1821 – 1894) - цветовое зрение Тип Моллюски
Тип Моллюски Тип Плоские черви. Класс Сосальщики
Тип Плоские черви. Класс Сосальщики Парамфистоматозы и другие широко распространенного трематодозы
Парамфистоматозы и другие широко распространенного трематодозы Круглые черви
Круглые черви