- Транскрипция - биосинтез РНК на матрице ДНК

Содержание
- 2. Транскрипция - биосинтез РНК на матрице ДНК Последовательность рибонуклеотидов в молекуле РНК комплементарна последовательности дезоксирибонуклеотидов одной
- 3. Считывание информации с ДНК-матрицы на РНК, синтез идет с помощью РНК-полимераз. У эукариот три РНК-полимеразы: (для
- 4. Механизм РНК-полимеразной реакции тот же, что и ДНК-полимеразной, направление синтеза 5’?3’, (субстратами служат нуклеозидтрифосфаты, тимину ДНК
- 6. Оперон и транскриптон Единицей транскрипции у прокариот является оперон Единицей транскрипции у эукариот транскриптон
- 7. Структура оперона Промотор - место инициации транскрипции, к которому присоединяется фермент РНК-полимераза; в ДНК E. coli
- 8. Структура оперона
- 9. Структура транскриптона
- 10. Промотор прокариот У бактерий функцию промотора выполняют две последовательности нуклеотидов на 5′-конце молекулы. Одна из них
- 11. Промотор эукариот Эукариотические гены, кодирующие белки, имеют блок Хогнесса (ТАТААА) в положении -25, а также ЦААТ-блок
- 12. Промотор прокариот и эукариот
- 13. Терминация синтеза Терминацию синтеза РНК вызывают длинные блоки АТ-последовательностей нуклеотидов в ДНК - терминатор (стоп-сигнал); У
- 14. Факторы, необходимые для транскрипции Матрица, которой является неспаренная цепь ДНК. В отличие от репликации транскрипция происходит
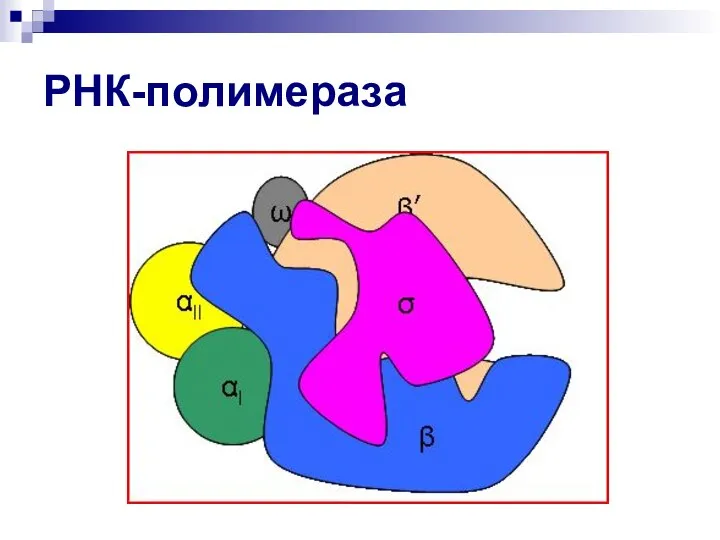
- 15. РНК-полимераза РНК-полимераза E.coli состоит из 5 субъединиц (α2ββ′ω, м.м. 390 000) и шестой σ-субъединицы. Фермент, состоящий
- 16. РНК-полимераза
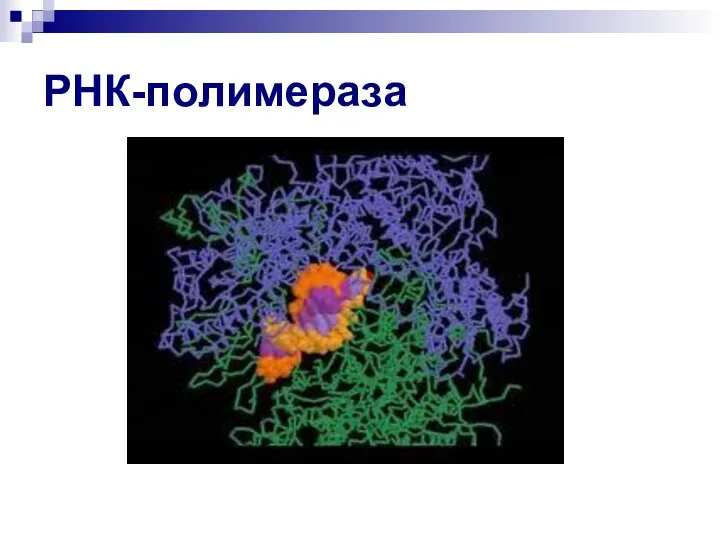
- 17. РНК-полимераза (продолжение) В эукариотических клетках присутствуют 3 ядерные РНК-полимеразы – I, II, III. РНК полимераза I
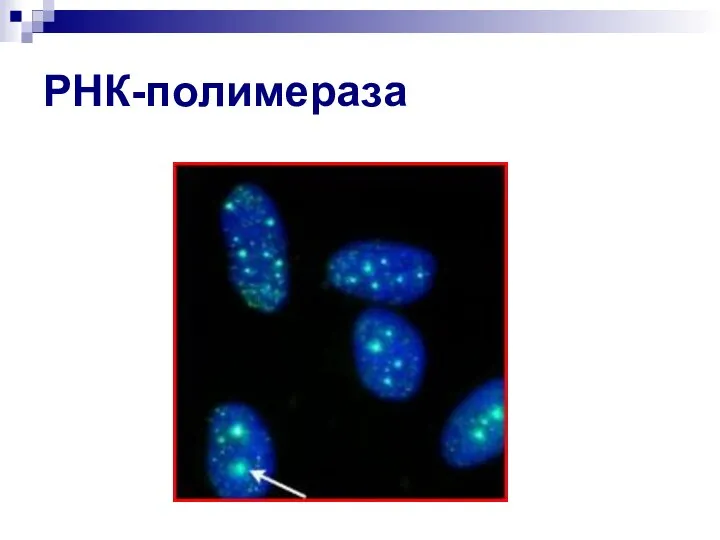
- 18. РНК-полимераза
- 19. РНК-полимераза
- 20. Стадии транскрипции Инициация Элонгация Терминация
- 21. Стадия инициации РНК-полимераза с помощью σ-субъединицы через серию случайных актов ассоциации-диссоциации находит промотор и происходит присоединение
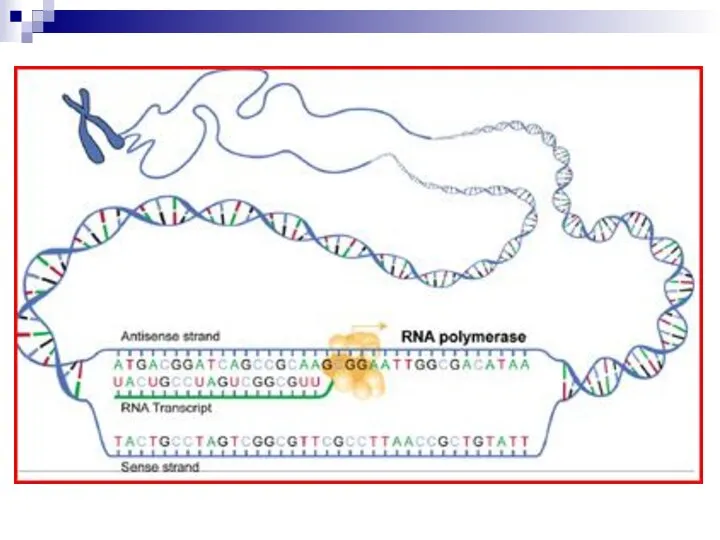
- 22. Стадия элонгации РНК полимераза синтезирует цепь РНК в направлении 5′→ 3′ антипараллельно матричной цепи ДНК (т.е.
- 24. Стадия элонгации (продолжение) Максимальная скорость элонгации составляет примерно 50 нуклеотидов в секунду. В отличие от ДНК-полимеразы
- 25. Стадия терминации Сигнал терминации синтеза молекулы РНК представляет собой определенную последовательность нуклеотидов, расположенную в рамках кодирующей
- 27. Посттрансляционная модификация РНК В результате транскрипции образуются три типа предшественников РНК (первичные транскрипты): предшественник мРНК, или
- 28. Посттрансляционная модификация РНК Вырезание неинформативных участков (интронов) из пре-мРНК. Сращивание информативных участков (экзонов) – сплайсинг. Модификация
- 29. Посттрансляционная модификация мРНК Вырезание неинформативных участков пре-мРНК происходит с помощью рибонуклеаз и/или рибозимов В ядре происходит
- 30. Посттрансляционная модификация тРНК Расщепление большого предшественника на молекулы тРНК. Присоединение характерного ЦЦА-триплета к 3′-концу молекулы (акцепторный
- 32. Посттрансляционная модификация рРНК Метилирование на участках, формирующих в дальнейшем зрелые молекулы рРНК. Расщепление общего первичного транскрипта
- 33. Обратная транскрипция
- 34. ОБРАТНАЯ ТРАНСКРИПЦИЯ Обратная транскриптаза содержится в РНК-ретровирусах, обладает тремя видами активности: 1) РНК-зависимой ДНК-полимеразной; 2) рибонуклеазной;
- 35. Стадии обратной транскрипции На матрице РНК по принципу комплементарности синтезируется ДНК - комплементарная ДНК (кДНК). Происходит
- 37. Ингибиторы обратной транскриптазы По принципу действия ингибиторы обратной транскриптазы делятся на нуклеозидные (НИОТ), нуклеотидные (НтИОТ) и
- 39. Скачать презентацию
Слайд 2Транскрипция - биосинтез РНК на матрице ДНК
Последовательность рибонуклеотидов в молекуле РНК комплементарна
Транскрипция - биосинтез РНК на матрице ДНК
Последовательность рибонуклеотидов в молекуле РНК комплементарна

Слайд 3Считывание информации с ДНК-матрицы на РНК, синтез идет с помощью РНК-полимераз.
У
Считывание информации с ДНК-матрицы на РНК, синтез идет с помощью РНК-полимераз. У

Слайд 4Механизм РНК-полимеразной реакции тот же, что и ДНК-полимеразной, направление синтеза 5’?3’, (субстратами
Механизм РНК-полимеразной реакции тот же, что и ДНК-полимеразной, направление синтеза 5’?3’, (субстратами

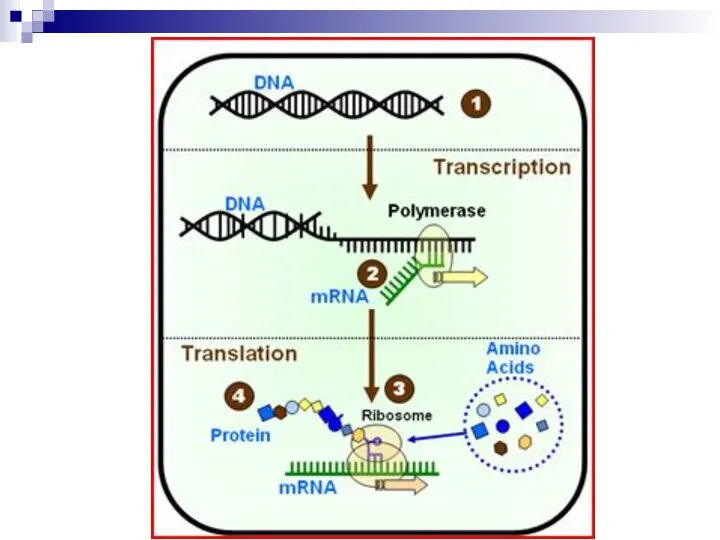
Слайд 6Оперон и транскриптон
Единицей транскрипции у прокариот является оперон
Единицей транскрипции у эукариот
Оперон и транскриптон
Единицей транскрипции у прокариот является оперон
Единицей транскрипции у эукариот

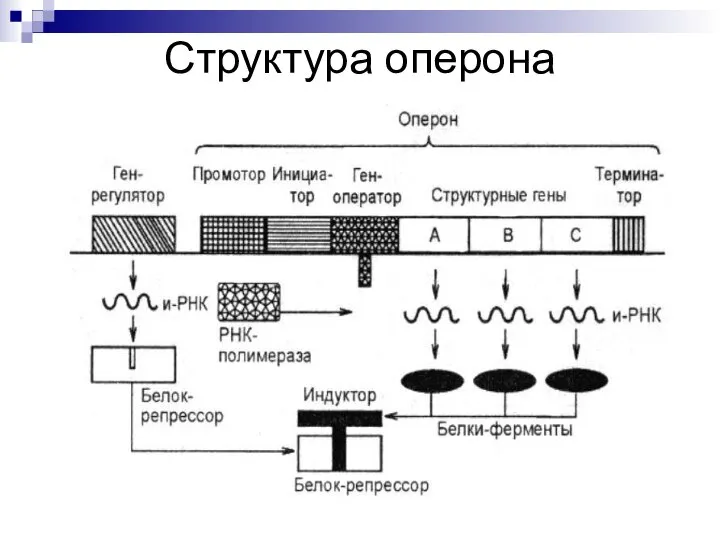
Слайд 7Структура оперона
Промотор - место инициации транскрипции, к которому присоединяется фермент РНК-полимераза; в
Структура оперона
Промотор - место инициации транскрипции, к которому присоединяется фермент РНК-полимераза; в

Слайд 8Структура оперона
Структура оперона
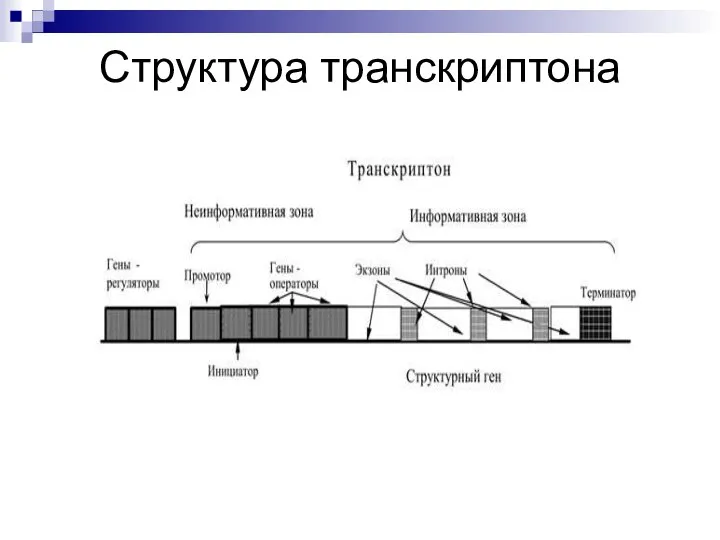
Слайд 9Структура транскриптона
Структура транскриптона
Слайд 10Промотор прокариот
У бактерий функцию промотора выполняют две последовательности нуклеотидов на 5′-конце молекулы.
Промотор прокариот
У бактерий функцию промотора выполняют две последовательности нуклеотидов на 5′-конце молекулы.

Слайд 11Промотор эукариот
Эукариотические гены, кодирующие белки, имеют блок Хогнесса (ТАТААА) в положении -25,
Промотор эукариот
Эукариотические гены, кодирующие белки, имеют блок Хогнесса (ТАТААА) в положении -25,

Слайд 12Промотор прокариот и эукариот
Промотор прокариот и эукариот
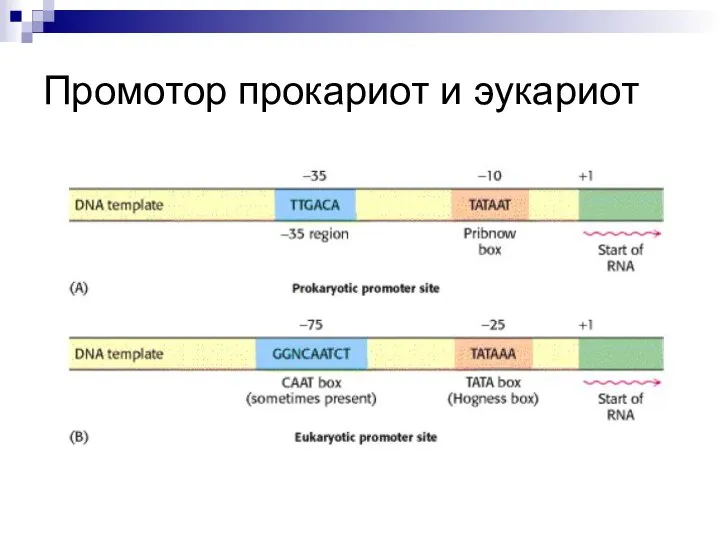
Слайд 13Терминация синтеза
Терминацию синтеза РНК вызывают длинные блоки АТ-последовательностей нуклеотидов в ДНК -
Терминация синтеза
Терминацию синтеза РНК вызывают длинные блоки АТ-последовательностей нуклеотидов в ДНК -

Слайд 14Факторы, необходимые для транскрипции
Матрица, которой является неспаренная цепь ДНК. В отличие от
Факторы, необходимые для транскрипции
Матрица, которой является неспаренная цепь ДНК. В отличие от

Слайд 15РНК-полимераза
РНК-полимераза E.coli состоит из 5 субъединиц (α2ββ′ω, м.м. 390 000) и шестой σ-субъединицы.
РНК-полимераза
РНК-полимераза E.coli состоит из 5 субъединиц (α2ββ′ω, м.м. 390 000) и шестой σ-субъединицы.

Слайд 16РНК-полимераза
РНК-полимераза
Слайд 17РНК-полимераза (продолжение)
В эукариотических клетках присутствуют 3 ядерные РНК-полимеразы – I, II, III.
РНК-полимераза (продолжение)
В эукариотических клетках присутствуют 3 ядерные РНК-полимеразы – I, II, III.

Слайд 18РНК-полимераза
РНК-полимераза
Слайд 19РНК-полимераза
РНК-полимераза
Слайд 20Стадии транскрипции
Инициация
Элонгация
Терминация
Стадии транскрипции
Инициация
Элонгация
Терминация

Слайд 21Стадия инициации
РНК-полимераза с помощью σ-субъединицы через серию случайных актов ассоциации-диссоциации находит промотор
Стадия инициации
РНК-полимераза с помощью σ-субъединицы через серию случайных актов ассоциации-диссоциации находит промотор

Слайд 22Стадия элонгации
РНК полимераза синтезирует цепь РНК в направлении 5′→ 3′ антипараллельно матричной
Стадия элонгации
РНК полимераза синтезирует цепь РНК в направлении 5′→ 3′ антипараллельно матричной

Слайд 24Стадия элонгации (продолжение)
Максимальная скорость элонгации составляет примерно 50 нуклеотидов в секунду.
Стадия элонгации (продолжение)
Максимальная скорость элонгации составляет примерно 50 нуклеотидов в секунду.

Слайд 25Стадия терминации
Сигнал терминации синтеза молекулы РНК представляет собой определенную последовательность нуклеотидов, расположенную
Стадия терминации
Сигнал терминации синтеза молекулы РНК представляет собой определенную последовательность нуклеотидов, расположенную

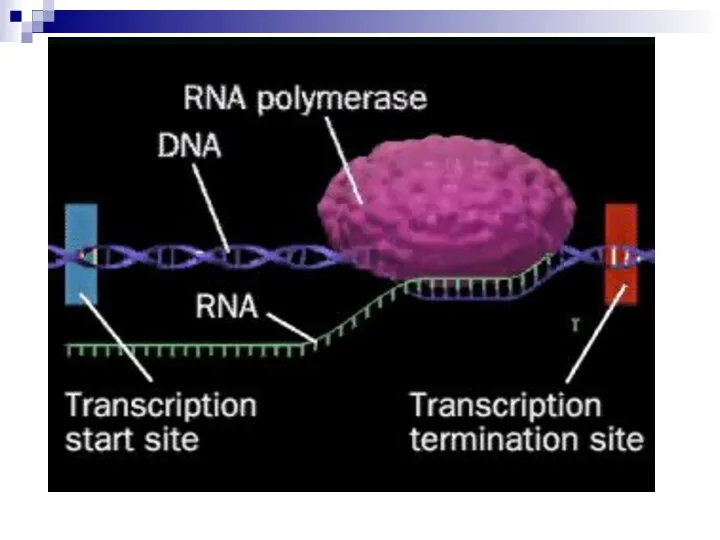
Слайд 27Посттрансляционная модификация РНК
В результате транскрипции образуются три типа предшественников РНК (первичные транскрипты):
Посттрансляционная модификация РНК
В результате транскрипции образуются три типа предшественников РНК (первичные транскрипты):

Слайд 28Посттрансляционная модификация РНК
Вырезание неинформативных участков (интронов) из пре-мРНК.
Сращивание информативных участков (экзонов) –
Посттрансляционная модификация РНК
Вырезание неинформативных участков (интронов) из пре-мРНК.
Сращивание информативных участков (экзонов) –

Слайд 29Посттрансляционная модификация мРНК
Вырезание неинформативных участков пре-мРНК происходит с помощью рибонуклеаз и/или рибозимов
Посттрансляционная модификация мРНК
Вырезание неинформативных участков пре-мРНК происходит с помощью рибонуклеаз и/или рибозимов

Слайд 30Посттрансляционная модификация тРНК
Расщепление большого предшественника на молекулы тРНК.
Присоединение характерного ЦЦА-триплета к
Посттрансляционная модификация тРНК
Расщепление большого предшественника на молекулы тРНК.
Присоединение характерного ЦЦА-триплета к

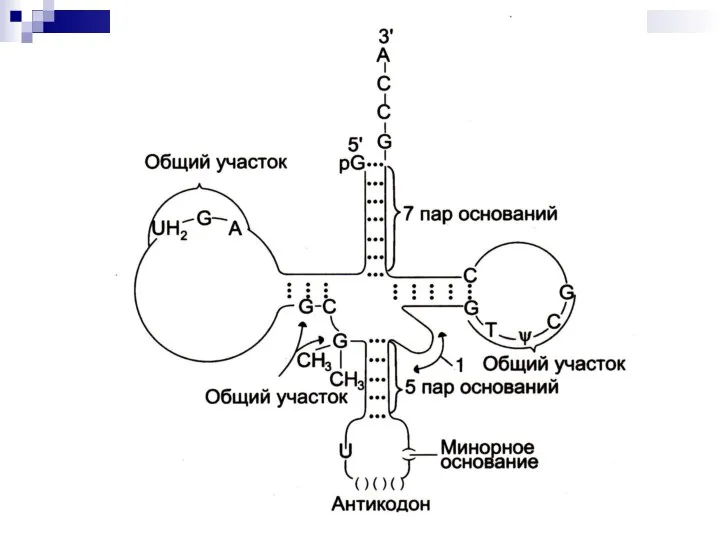
Слайд 32Посттрансляционная модификация рРНК
Метилирование на участках, формирующих в дальнейшем зрелые молекулы рРНК.
Расщепление
Посттрансляционная модификация рРНК
Метилирование на участках, формирующих в дальнейшем зрелые молекулы рРНК.
Расщепление

Слайд 33Обратная транскрипция
Обратная транскрипция

Слайд 34ОБРАТНАЯ ТРАНСКРИПЦИЯ
Обратная транскриптаза содержится в РНК-ретровирусах, обладает тремя видами активности:
1) РНК-зависимой ДНК-полимеразной;
2)
ОБРАТНАЯ ТРАНСКРИПЦИЯ
Обратная транскриптаза содержится в РНК-ретровирусах, обладает тремя видами активности:
1) РНК-зависимой ДНК-полимеразной;
2)

Слайд 35Стадии обратной транскрипции
На матрице РНК по принципу комплементарности синтезируется ДНК - комплементарная
Стадии обратной транскрипции
На матрице РНК по принципу комплементарности синтезируется ДНК - комплементарная

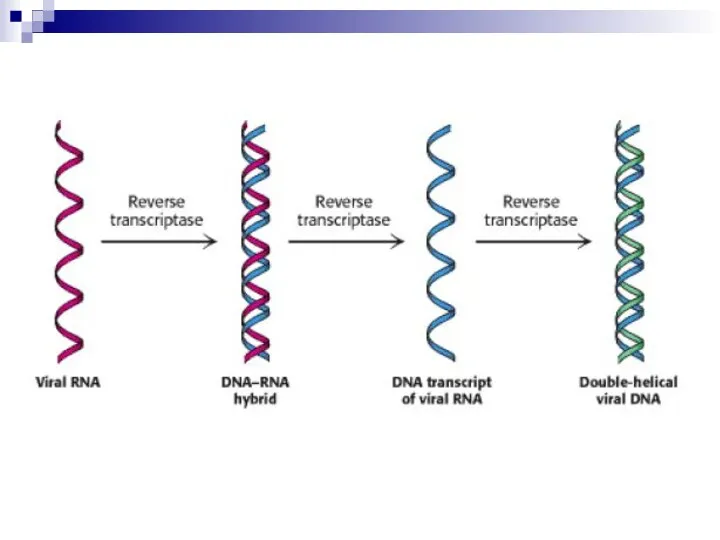
Слайд 37Ингибиторы обратной транскриптазы
По принципу действия ингибиторы обратной транскриптазы делятся на нуклеозидные (НИОТ),
Ингибиторы обратной транскриптазы
По принципу действия ингибиторы обратной транскриптазы делятся на нуклеозидные (НИОТ),

 Бабочка семейства Павлиноглазки Samia Cynthia, или Айлантовый шелкопряд. Опыт разведения в неволе
Бабочка семейства Павлиноглазки Samia Cynthia, или Айлантовый шелкопряд. Опыт разведения в неволе Презентация на тему Методы селекции растений
Презентация на тему Методы селекции растений  Ткани животных
Ткани животных Луга нашей Родины
Луга нашей Родины bf9c797d6665ca36da02d2ec9ab78d9a
bf9c797d6665ca36da02d2ec9ab78d9a Питание растений и грибов
Питание растений и грибов Типы взаимодействия между организмами
Типы взаимодействия между организмами Структура и строение кости
Структура и строение кости Изучение мутагенного действия нового вещества in vitro. План
Изучение мутагенного действия нового вещества in vitro. План 2_Selektsia_zhivotnykh
2_Selektsia_zhivotnykh Мышцы кисти
Мышцы кисти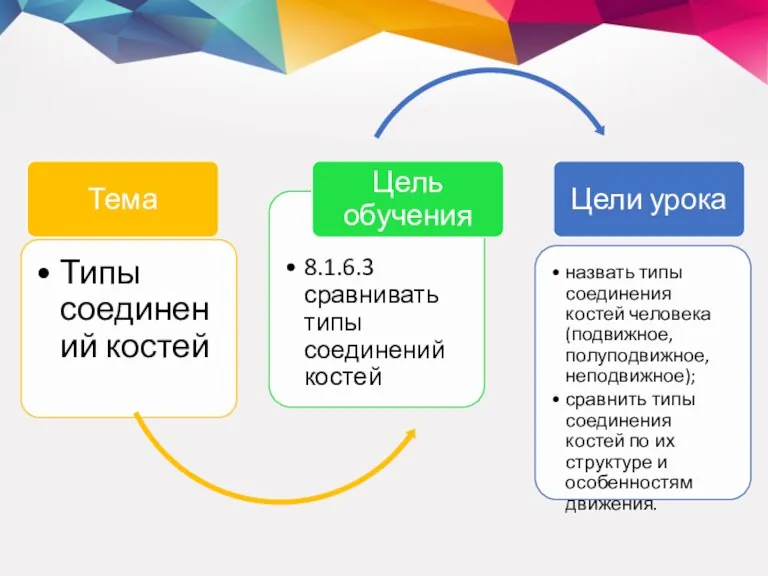 Типы соединений костей
Типы соединений костей Школьная научно-практическая работа Рыбы нашего района. 7 класс
Школьная научно-практическая работа Рыбы нашего района. 7 класс Презентация на тему Сосальщики и ленточные черви
Презентация на тему Сосальщики и ленточные черви  Спланхнология. Внутренние органы человека
Спланхнология. Внутренние органы человека Автор: Филатова Анжелика, ученица 6 класса Руководитель: Филатова С.Н., учитель биологии МБОУ СОШ №6 город Ливны 2012 год
Автор: Филатова Анжелика, ученица 6 класса Руководитель: Филатова С.Н., учитель биологии МБОУ СОШ №6 город Ливны 2012 год Органы пищеварения
Органы пищеварения Законы наследственности
Законы наследственности Proekt
Proekt Проект Planet Micro
Проект Planet Micro Развитие жизни на Земле
Развитие жизни на Земле Покровные ткани растений
Покровные ткани растений Паразитизм как экологическое явление.Задачи медицинской паразитологии
Паразитизм как экологическое явление.Задачи медицинской паразитологии Мышцы плечевого пояса и свободной верхней конечности
Мышцы плечевого пояса и свободной верхней конечности Домики, которые построила природа
Домики, которые построила природа Виявлення наявності маркерів вірусного гепатиту В у плазмі крові донорів
Виявлення наявності маркерів вірусного гепатиту В у плазмі крові донорів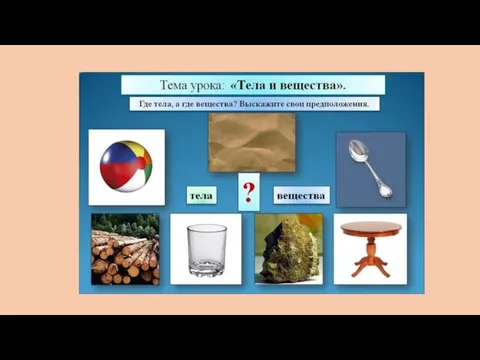 Тела и вещества
Тела и вещества Жизнедеятельность клетки
Жизнедеятельность клетки