- Гомологическая рекомбинация

Содержание
- 2. Ники (одноцепоченчные разрывы) в ДНК – часто встречающаяся штука, побочный продукт действия на ДНК множества разных
- 3. Некоторые бактериальные ферменты гомологической рекомбинации
- 4. Комплекс RecBCD RecBCD обладает многими активностями, среди которых хеликазная, экзо- и эндонуклеазная и АТФазная. Он связывается
- 5. Комплекс RecBCD При попадании RecBCD на хи-сайт его 3’-5’-экзо-нуклеазная активность подавляется, а 5’-3’-экзо-нуклеазная активность, напротив, стимулируется.
- 6. Хи-сайт – очень важная штука Если в ДНК случился дц разрыв в ходе репликации, в дело
- 7. RecF-рекомбинация Используется в случае оц разрывов ДНК, но редко – их проще сразу достроить Хеликаза RecQ
- 8. Модель работы белка RecA RecA, связанный с оц ДНК, в виде рибонуклеопротеинового комплекса внедряется в дуплекс
- 9. Структуры Холидея, образующиеся после спулинга Та самая трехцепочечная структура способна сформировать структуру Холидея. Она может либо
- 10. Ruv-белки формируют структуры Холидея Все начинается с той же структуры, с которой все начиналось на предыдущем
- 11. Ruv-белки разрешают структуры Холидея В результате в центре структуры Холидея создается достаточно пространства, чтобы RuvA мог
- 12. Интеграция линейных фрагментов в бактериальную хромосому (плазмиды, фаги…) В жизни любой бактерии наступает момент, когда в
- 13. Интеграция линейных фрагментов в бактериальную хромосому «Левая» часть синапса со временем сливается с вилкой репликации, и
- 14. Репарация ДНК Репарация есть процесс исправления повреждений в ДНК. ДНК подвержена воздействиям многих факторов, среди которых
- 15. Дезаминирование нуклеотидов Может происходить под действием дезаминирующих агентов (например, азотистой кислоты), что часто вызывает спаривание А-С.
- 16. Репарация дезаминированных нуклеотидов Специальная ДНК-гликозилаза вырезает неправильное основание, узнавая его. А потом приходит АР-эндонуклеаза и вносит
- 17. Репарация 8-оксигуанина (GO) Гуанин гораздо больше других нуклеотидов подвержен действию АФК. Активные формы кислорода могут приводить
- 18. Алкилирование гуанина Здесь представлен самый частый случай алкилирования, приводящий к образованию О6-метилгуанина. Он может спариваться с
- 19. Адаптивный ответ клетки на алкилирующие агенты Система бактериального адаптивного ответа состоит из четырех генов. В норме
- 20. Тиминовые димеры При действии УФ-света на клетку два соседних тимина в ДНК (а также некоторые другие
- 21. Репарация (фотореактивация) тиминовых димеров Процесс осуществляется специальными фотолиазами, которые активируются светом видимого диапазона. После такой активации
- 22. Mismatch-репарация Иногда бывает так, что в ДНК вместо нормальных нуклеотидов встраиваются химические соединения, очень на них
- 23. Mismatch-репарация Основной вопрос, встающий перед системой: в какой цепи ошибочный нуклеотид? Ответить на него помогает метилирование.
- 24. MutSLH-механизм mismatch-репарации Итак, имеется свежереплицированная ДНК (часто еще в процессе репликации) с мисмэтчем. Рядышком обязательно окажется
- 25. MutSLH-механизм mismatch-репарации Здесь в дело вступают экзонуклеазы, превращающие ник в «gap», и хеликаза UvrD, чувствующая, где
- 26. Nucleotide excision repair (эксцизионная репарация нуклеотидов) В отличие от рассмотренных ранее способов репарации повреждений, здесь репарации
- 27. Связь репарации и репликации Мы говорили о тех случаях, когда репарация случилась до репликации. Тогда все
- 28. Связь репарации и репликации Но ведь такая ситуация идеальна для гомологической рекомбинации! Тут же на сцене
- 29. SOS-репарация В жизни клетки бывает так, что повреждений в ДНК происходит сразу очень много. Тогда и
- 30. SOS-репарация При сильных повреждениях ДНК в клетке неизбежно появляются короткие оц фрагменты ДНК. У белка RecA
- 31. SOS-репарация: мутасома pol V Итак, в норме экспрессия генов umuC и umuD подавлена белком LexA. При
- 32. SOS-репарация: мутасома pol V Отдельные мономеры белка RecA способны покинуть филаменты, образованные им на оцДНК, и
- 34. Скачать презентацию
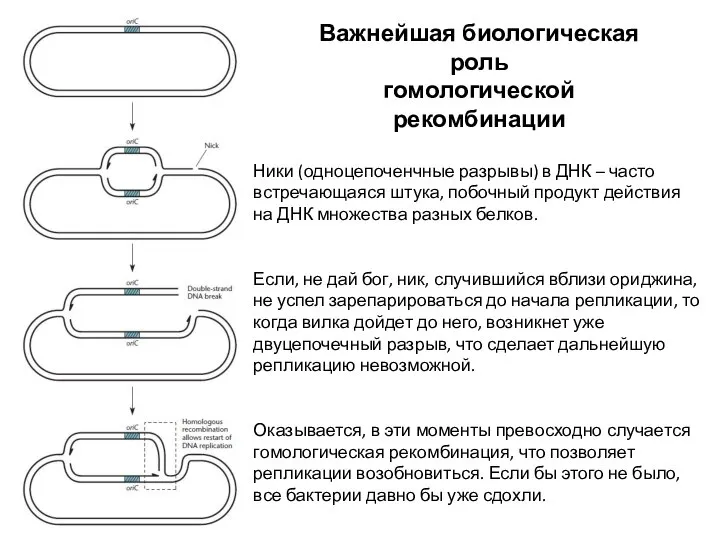
Слайд 3Некоторые бактериальные ферменты
гомологической рекомбинации
Некоторые бактериальные ферменты
гомологической рекомбинации
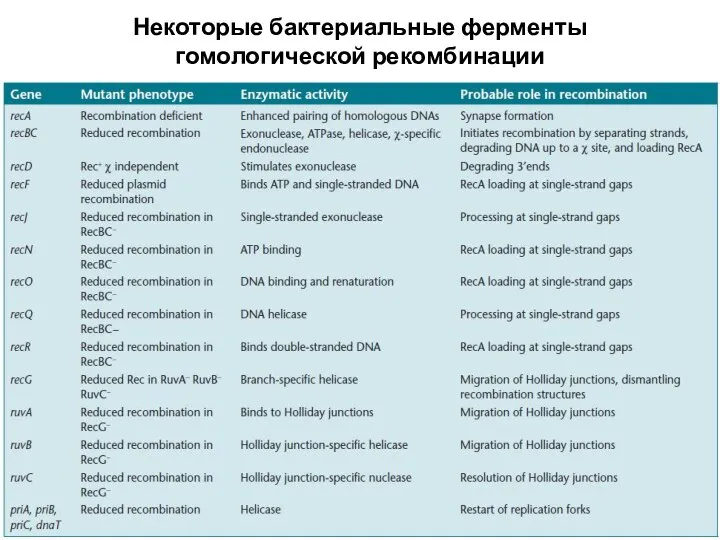
Слайд 4Комплекс RecBCD
RecBCD обладает многими активностями, среди которых хеликазная, экзо- и эндонуклеазная и
Комплекс RecBCD
RecBCD обладает многими активностями, среди которых хеликазная, экзо- и эндонуклеазная и
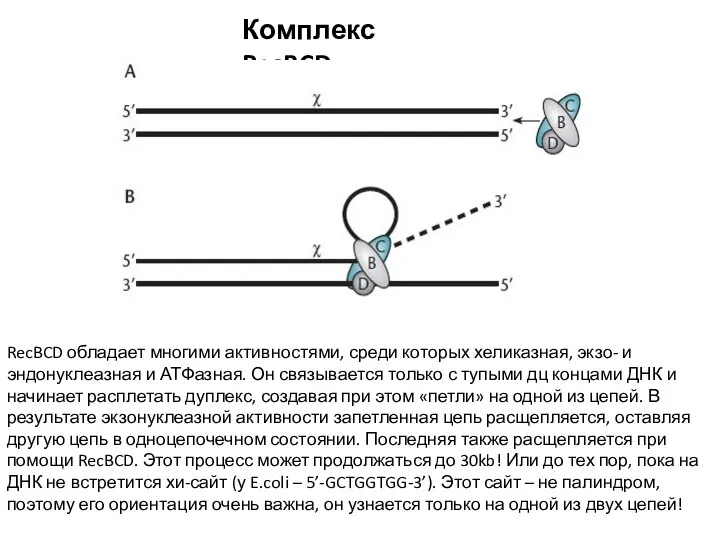
Слайд 5Комплекс RecBCD
При попадании RecBCD на хи-сайт его 3’-5’-экзо-нуклеазная активность подавляется, а 5’-3’-экзо-нуклеазная
Комплекс RecBCD
При попадании RecBCD на хи-сайт его 3’-5’-экзо-нуклеазная активность подавляется, а 5’-3’-экзо-нуклеазная
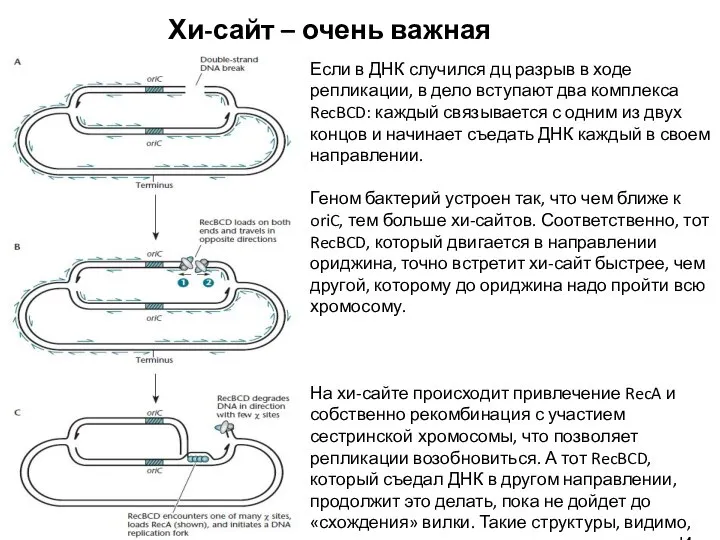
Слайд 6Хи-сайт – очень важная штука
Если в ДНК случился дц разрыв в ходе
Хи-сайт – очень важная штука
Если в ДНК случился дц разрыв в ходе
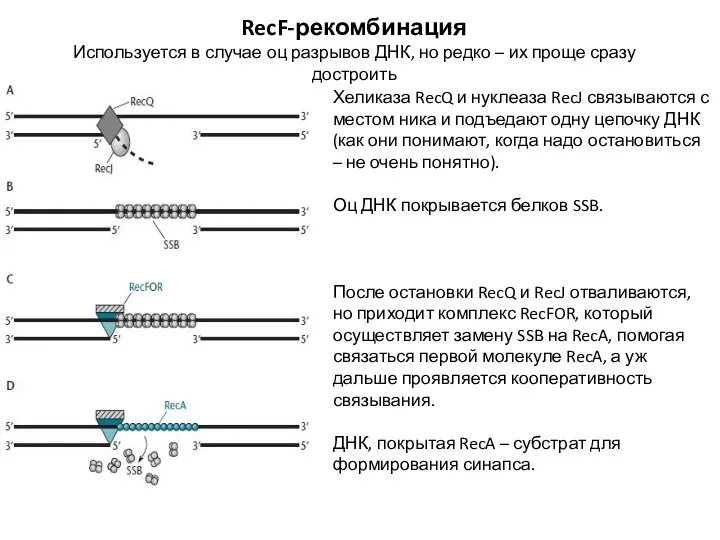
Слайд 7RecF-рекомбинация
Используется в случае оц разрывов ДНК, но редко – их проще сразу
RecF-рекомбинация
Используется в случае оц разрывов ДНК, но редко – их проще сразу
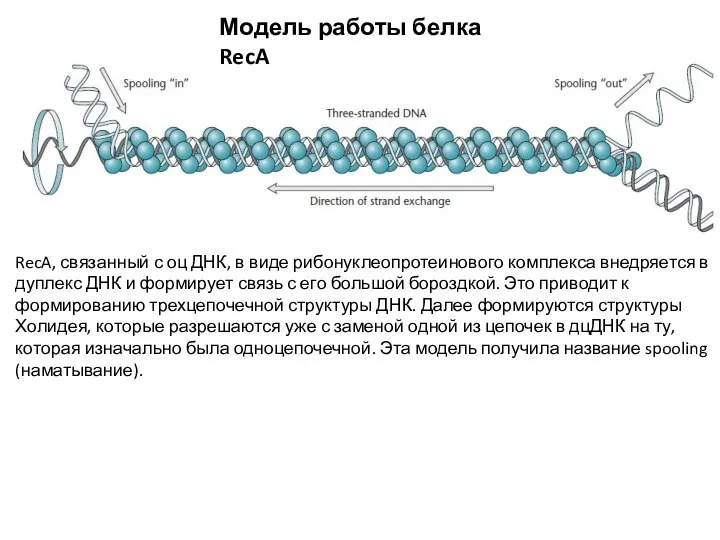
Слайд 8Модель работы белка RecA
RecA, связанный с оц ДНК, в виде рибонуклеопротеинового комплекса
Модель работы белка RecA
RecA, связанный с оц ДНК, в виде рибонуклеопротеинового комплекса
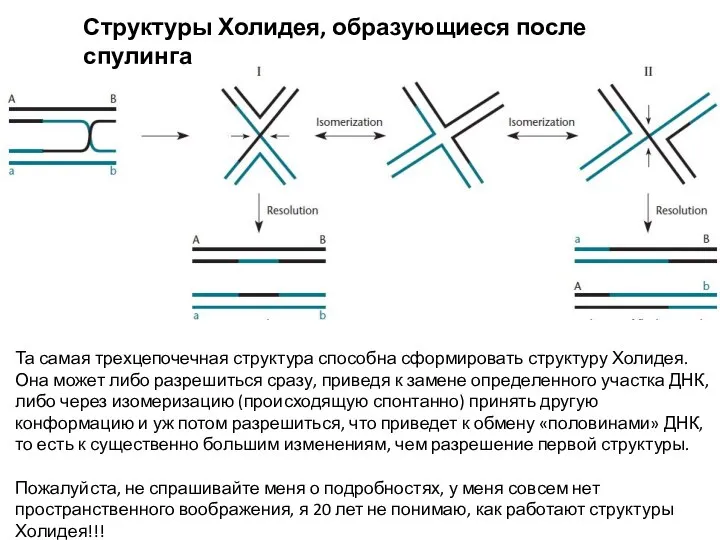
Слайд 9Структуры Холидея, образующиеся после спулинга
Та самая трехцепочечная структура способна сформировать структуру Холидея.
Структуры Холидея, образующиеся после спулинга
Та самая трехцепочечная структура способна сформировать структуру Холидея.
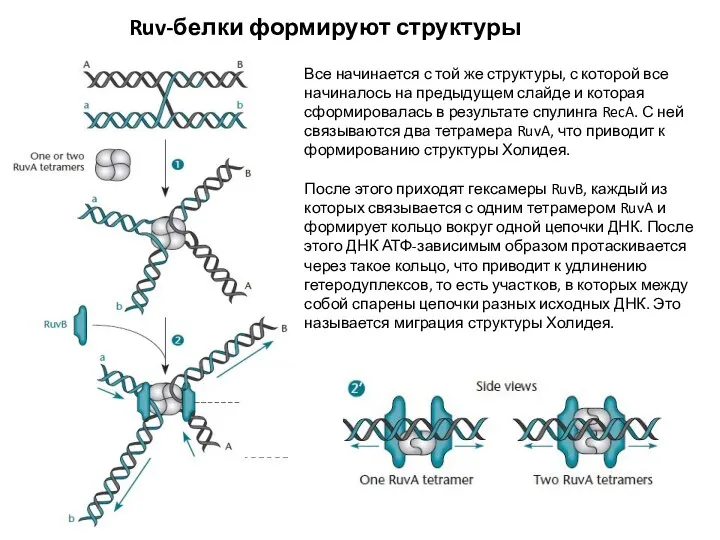
Слайд 10Ruv-белки формируют структуры Холидея
Все начинается с той же структуры, с которой все
Ruv-белки формируют структуры Холидея
Все начинается с той же структуры, с которой все
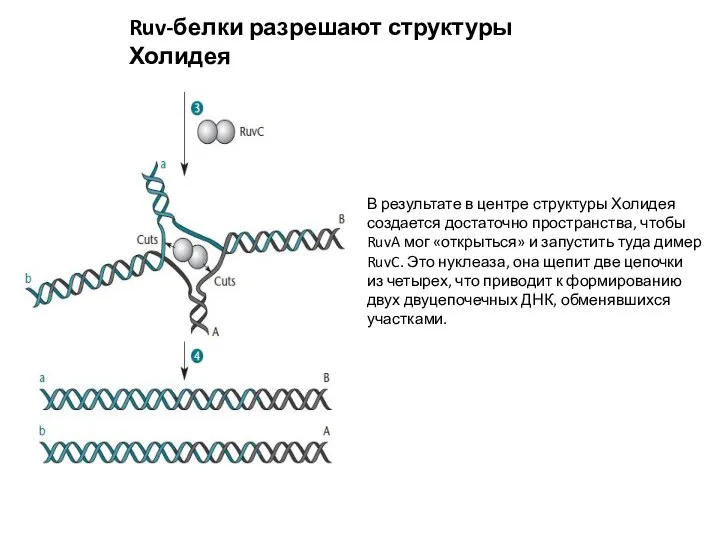
Слайд 11Ruv-белки разрешают структуры Холидея
В результате в центре структуры Холидея создается достаточно пространства,
Ruv-белки разрешают структуры Холидея
В результате в центре структуры Холидея создается достаточно пространства,
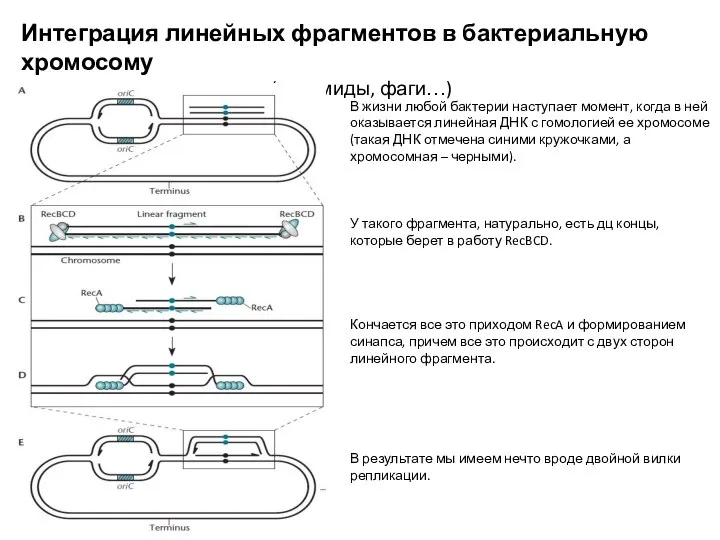
Слайд 12Интеграция линейных фрагментов в бактериальную хромосому
(плазмиды, фаги…)
В жизни любой бактерии наступает момент,
Интеграция линейных фрагментов в бактериальную хромосому
(плазмиды, фаги…)
В жизни любой бактерии наступает момент,
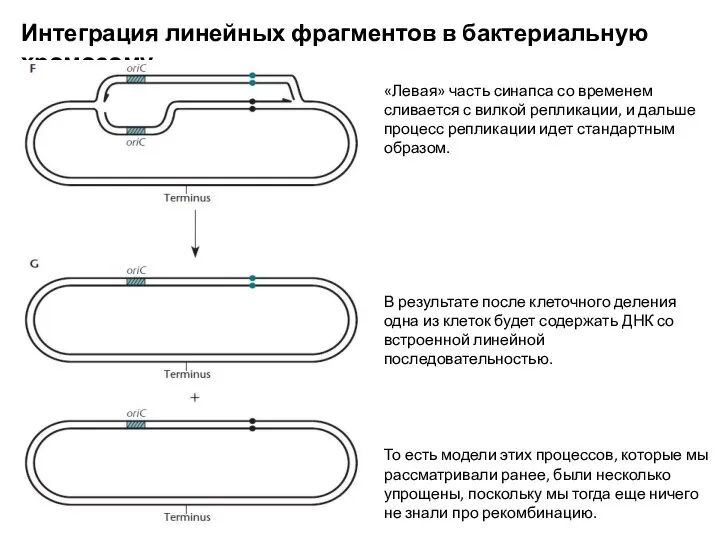
Слайд 13Интеграция линейных фрагментов в бактериальную хромосому
«Левая» часть синапса со временем сливается с
Интеграция линейных фрагментов в бактериальную хромосому
«Левая» часть синапса со временем сливается с
Слайд 14Репарация ДНК
Репарация есть процесс исправления повреждений в ДНК.
ДНК подвержена воздействиям многих факторов,
Репарация ДНК
Репарация есть процесс исправления повреждений в ДНК.
ДНК подвержена воздействиям многих факторов,

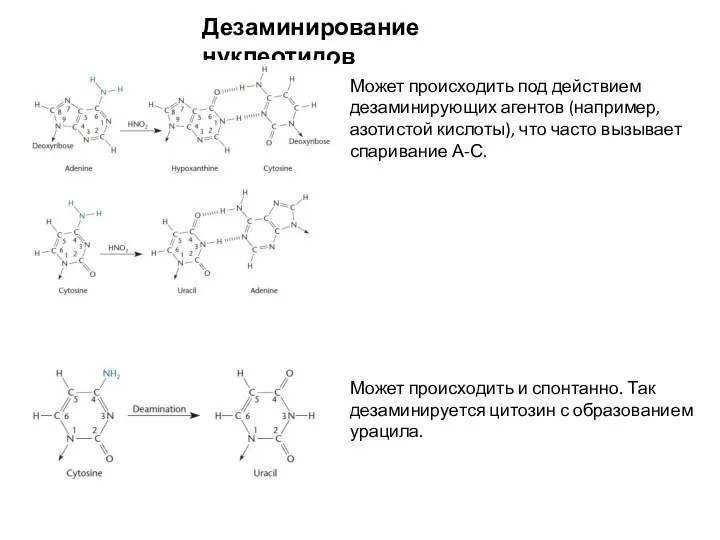
Слайд 15Дезаминирование нуклеотидов
Может происходить под действием дезаминирующих агентов (например, азотистой кислоты), что часто
Дезаминирование нуклеотидов
Может происходить под действием дезаминирующих агентов (например, азотистой кислоты), что часто
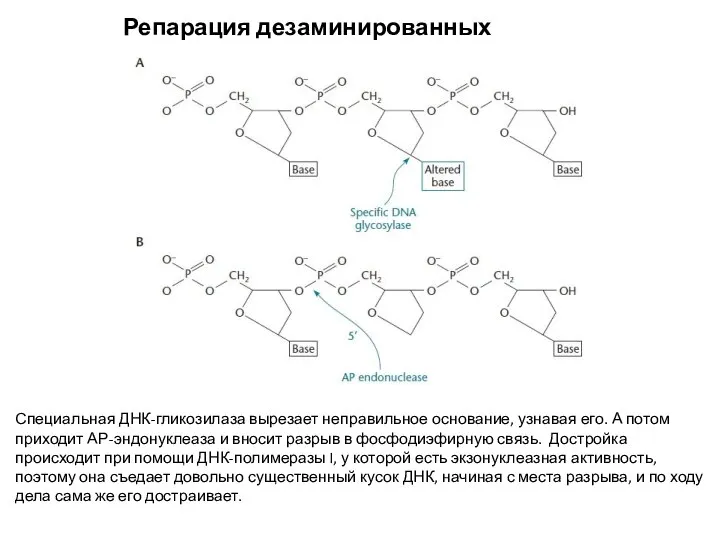
Слайд 16Репарация дезаминированных нуклеотидов
Специальная ДНК-гликозилаза вырезает неправильное основание, узнавая его. А потом приходит
Репарация дезаминированных нуклеотидов
Специальная ДНК-гликозилаза вырезает неправильное основание, узнавая его. А потом приходит
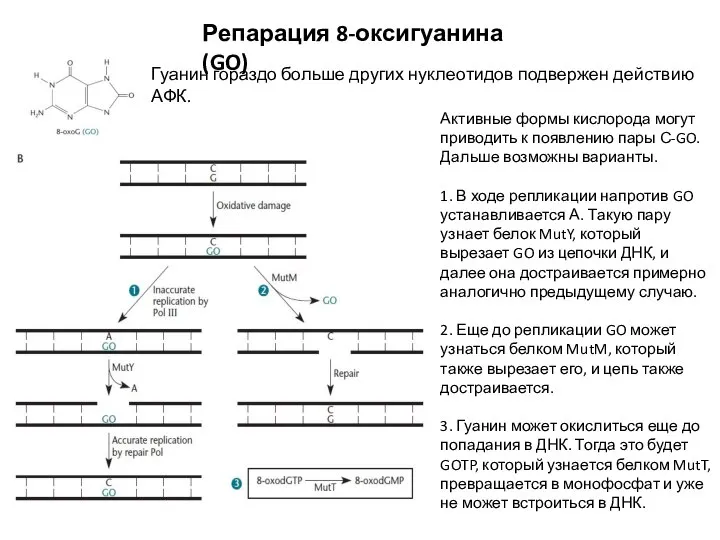
Слайд 17Репарация 8-оксигуанина (GO)
Гуанин гораздо больше других нуклеотидов подвержен действию АФК.
Активные формы кислорода
Репарация 8-оксигуанина (GO)
Гуанин гораздо больше других нуклеотидов подвержен действию АФК.
Активные формы кислорода
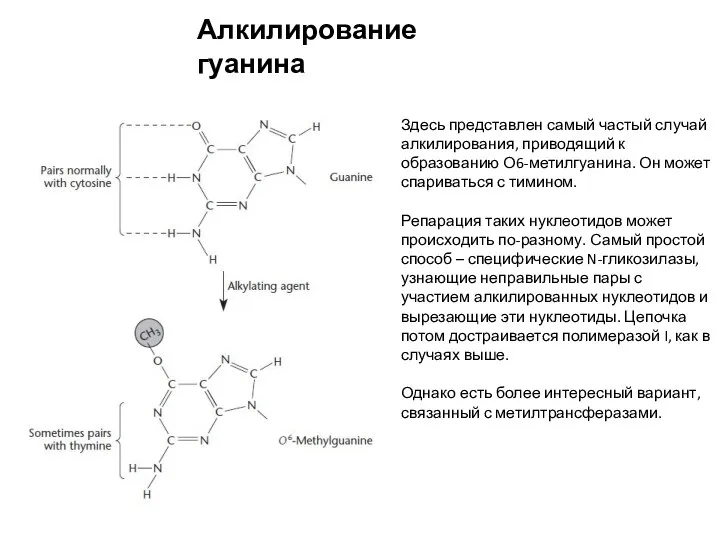
Слайд 18Алкилирование гуанина
Здесь представлен самый частый случай алкилирования, приводящий к образованию О6-метилгуанина. Он
Алкилирование гуанина
Здесь представлен самый частый случай алкилирования, приводящий к образованию О6-метилгуанина. Он
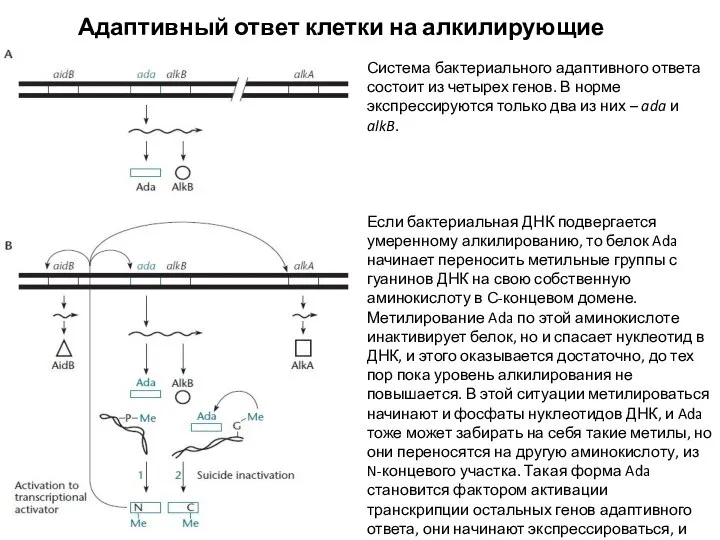
Слайд 19Адаптивный ответ клетки на алкилирующие агенты
Система бактериального адаптивного ответа состоит из четырех
Адаптивный ответ клетки на алкилирующие агенты
Система бактериального адаптивного ответа состоит из четырех
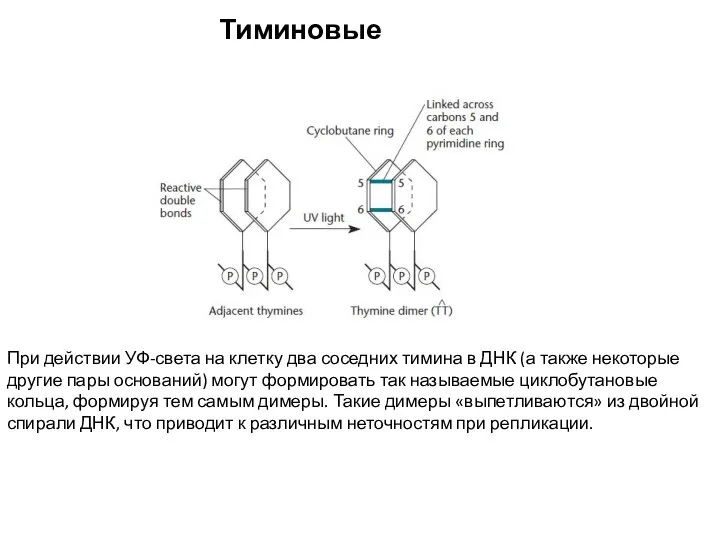
Слайд 20Тиминовые димеры
При действии УФ-света на клетку два соседних тимина в ДНК (а
Тиминовые димеры
При действии УФ-света на клетку два соседних тимина в ДНК (а
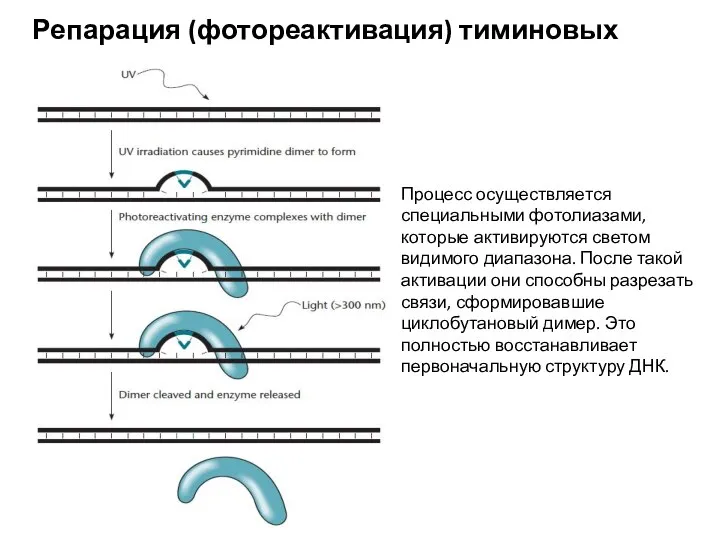
Слайд 21Репарация (фотореактивация) тиминовых димеров
Процесс осуществляется специальными фотолиазами, которые активируются светом видимого диапазона.
Репарация (фотореактивация) тиминовых димеров
Процесс осуществляется специальными фотолиазами, которые активируются светом видимого диапазона.
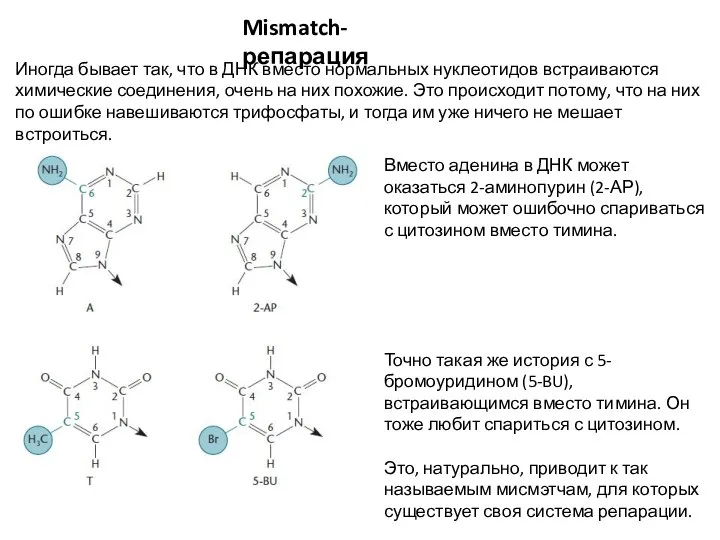
Слайд 22Mismatch-репарация
Иногда бывает так, что в ДНК вместо нормальных нуклеотидов встраиваются химические соединения,
Mismatch-репарация
Иногда бывает так, что в ДНК вместо нормальных нуклеотидов встраиваются химические соединения,
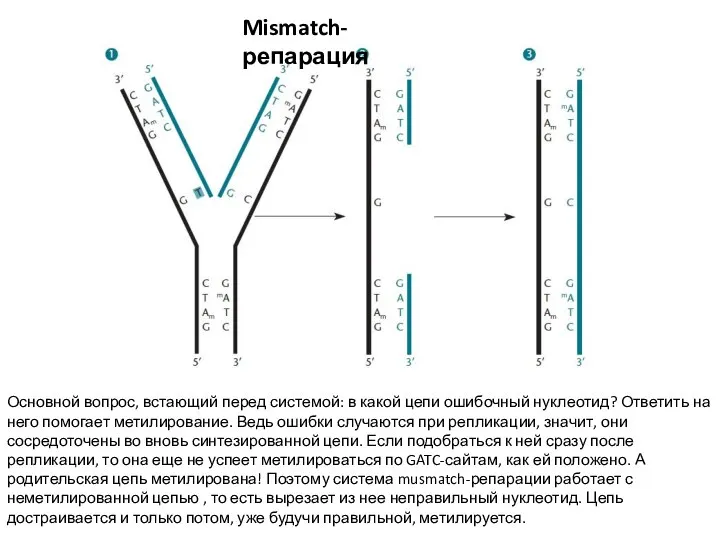
Слайд 23Mismatch-репарация
Основной вопрос, встающий перед системой: в какой цепи ошибочный нуклеотид? Ответить на
Mismatch-репарация
Основной вопрос, встающий перед системой: в какой цепи ошибочный нуклеотид? Ответить на
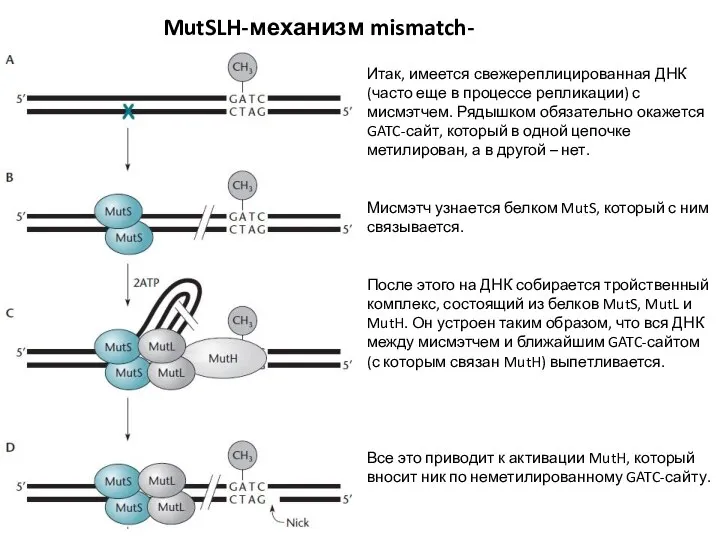
Слайд 24MutSLH-механизм mismatch-репарации
Итак, имеется свежереплицированная ДНК (часто еще в процессе репликации) с мисмэтчем.
MutSLH-механизм mismatch-репарации
Итак, имеется свежереплицированная ДНК (часто еще в процессе репликации) с мисмэтчем.
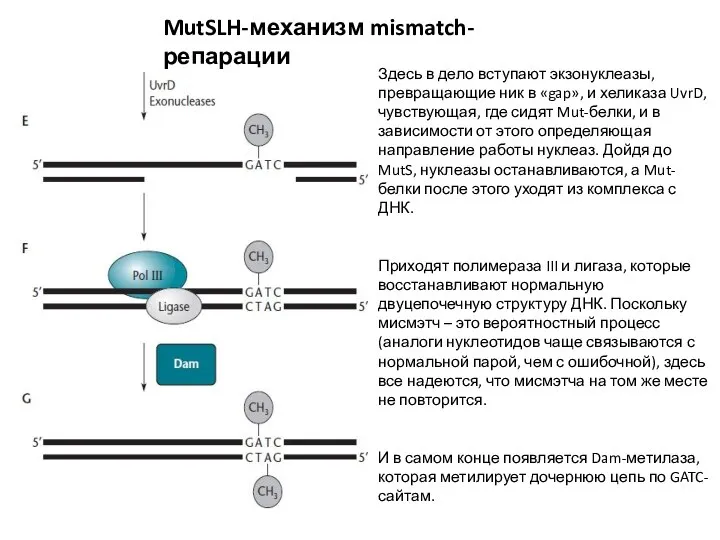
Слайд 25MutSLH-механизм mismatch-репарации
Здесь в дело вступают экзонуклеазы, превращающие ник в «gap», и хеликаза
MutSLH-механизм mismatch-репарации
Здесь в дело вступают экзонуклеазы, превращающие ник в «gap», и хеликаза
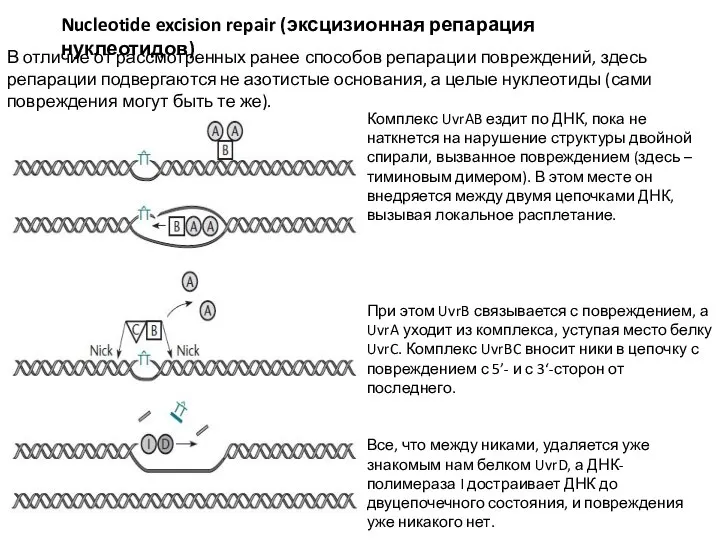
Слайд 26Nucleotide excision repair (эксцизионная репарация нуклеотидов)
В отличие от рассмотренных ранее способов репарации
Nucleotide excision repair (эксцизионная репарация нуклеотидов)
В отличие от рассмотренных ранее способов репарации
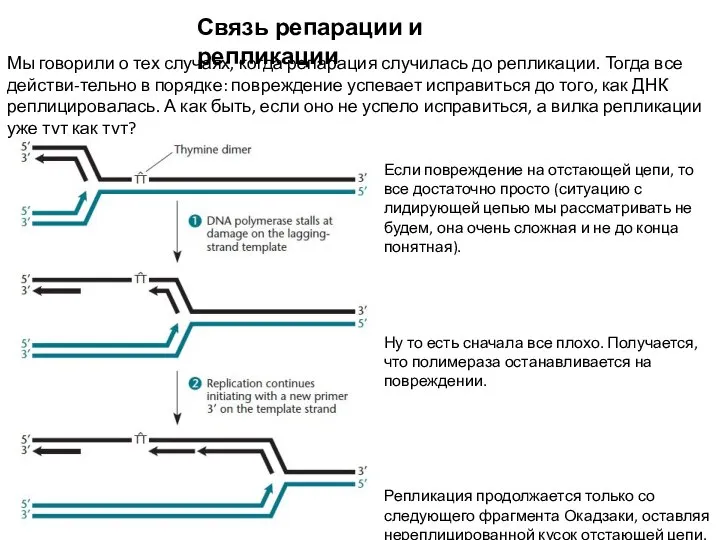
Слайд 27Связь репарации и репликации
Мы говорили о тех случаях, когда репарация случилась до
Связь репарации и репликации
Мы говорили о тех случаях, когда репарация случилась до
Слайд 28Связь репарации и репликации
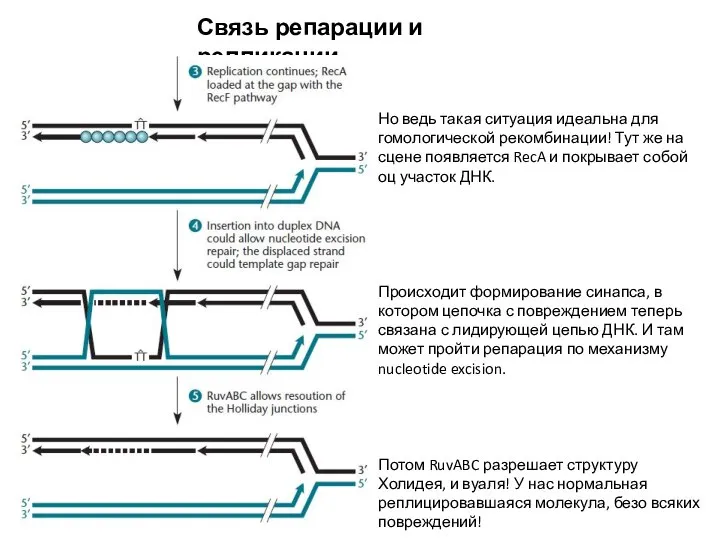
Но ведь такая ситуация идеальна для гомологической рекомбинации! Тут
Связь репарации и репликации
Но ведь такая ситуация идеальна для гомологической рекомбинации! Тут
Слайд 29SOS-репарация
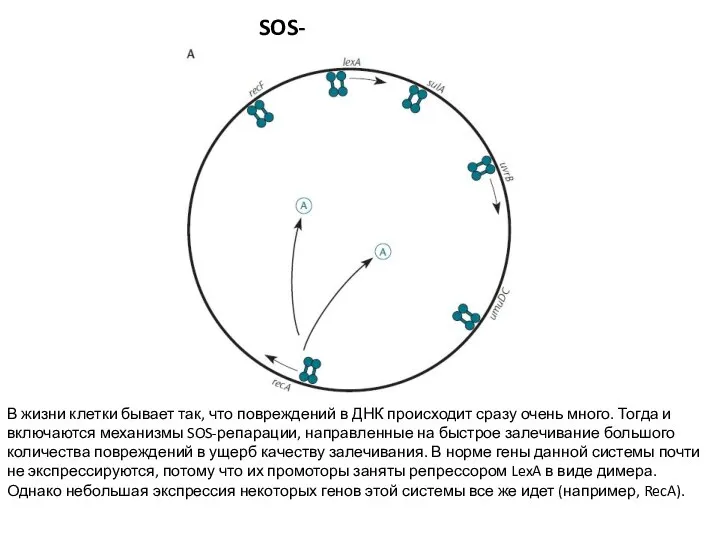
В жизни клетки бывает так, что повреждений в ДНК происходит сразу очень
SOS-репарация
В жизни клетки бывает так, что повреждений в ДНК происходит сразу очень
Слайд 30SOS-репарация
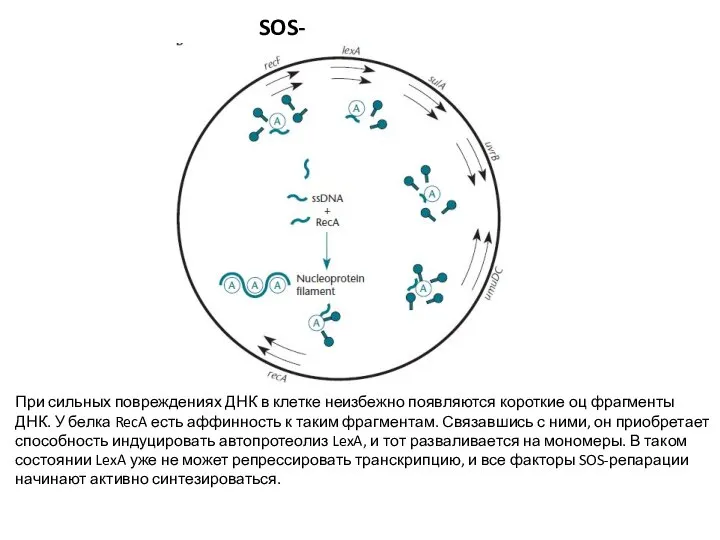
При сильных повреждениях ДНК в клетке неизбежно появляются короткие оц фрагменты ДНК.
SOS-репарация
При сильных повреждениях ДНК в клетке неизбежно появляются короткие оц фрагменты ДНК.
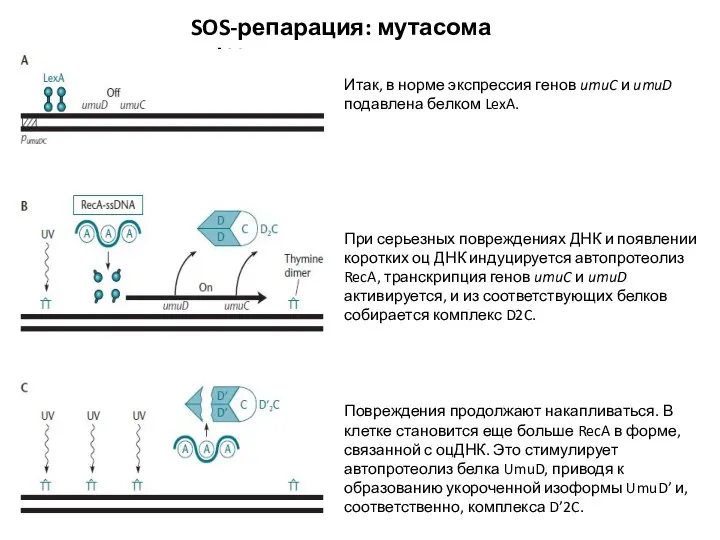
Слайд 31SOS-репарация: мутасома pol V
Итак, в норме экспрессия генов umuC и umuD подавлена
SOS-репарация: мутасома pol V
Итак, в норме экспрессия генов umuC и umuD подавлена
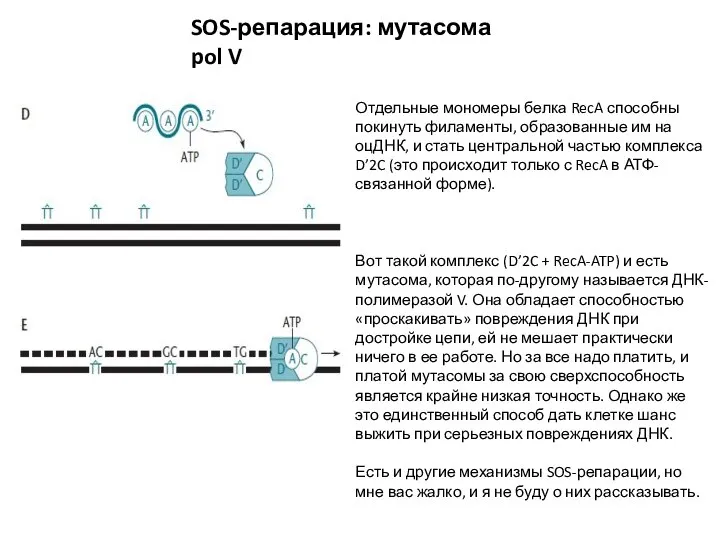
Слайд 32SOS-репарация: мутасома pol V
Отдельные мономеры белка RecA способны покинуть филаменты, образованные им
SOS-репарация: мутасома pol V
Отдельные мономеры белка RecA способны покинуть филаменты, образованные им
 Шырайгүл, петуния
Шырайгүл, петуния Bai giang ky thuat y sinh
Bai giang ky thuat y sinh Опасные растения и насекомые Крыма
Опасные растения и насекомые Крыма Презентация на тему Ель, сосна – хвойные растения
Презентация на тему Ель, сосна – хвойные растения  Многообразие губок
Многообразие губок Рыба пиранья
Рыба пиранья Сосуды основания сердца
Сосуды основания сердца Презентация на тему Строение клетки (10 класс)
Презентация на тему Строение клетки (10 класс)  Головной мозг. Отделы головного мозга
Головной мозг. Отделы головного мозга Бактерии – разрушители органических веществ
Бактерии – разрушители органических веществ Презентация на тему Влияние солнечной активности на социальные процессы
Презентация на тему Влияние солнечной активности на социальные процессы  Физика и кошка
Физика и кошка Методы определения концентрации спермиев
Методы определения концентрации спермиев Физиология возбудимых тканей
Физиология возбудимых тканей Генетическое оружие нейтрофила
Генетическое оружие нейтрофила Низшие растения. Водоросли
Низшие растения. Водоросли Селекция (часть 2)
Селекция (часть 2) Криптонит для растений
Криптонит для растений Україна від А до Я. Природа (слайди)
Україна від А до Я. Природа (слайди) Посмотри и улыбнись. Животные
Посмотри и улыбнись. Животные Обмен веществ и энергии в клетках
Обмен веществ и энергии в клетках Презентация на тему ОТРЯД НАСЕКОМОЯДНЫЕ
Презентация на тему ОТРЯД НАСЕКОМОЯДНЫЕ  Появление Человека Разумного. Урок № 13
Появление Человека Разумного. Урок № 13 Эндокринная система человека
Эндокринная система человека Деревья Калифорнии
Деревья Калифорнии Понятие морфогенов и градиентов их концентраций
Понятие морфогенов и градиентов их концентраций Дезоксирибонуклеиновая кислота ДНК
Дезоксирибонуклеиновая кислота ДНК Мікробіологічні препарати у виноградарстві
Мікробіологічні препарати у виноградарстві