- Гормоны периферических желез внутренней секреции

Содержание
- 2. Структура молекулы инсулина: Молекула инсулина построена из двух пептидных цепей: цепь А содержит 21 аминокислотный остаток,
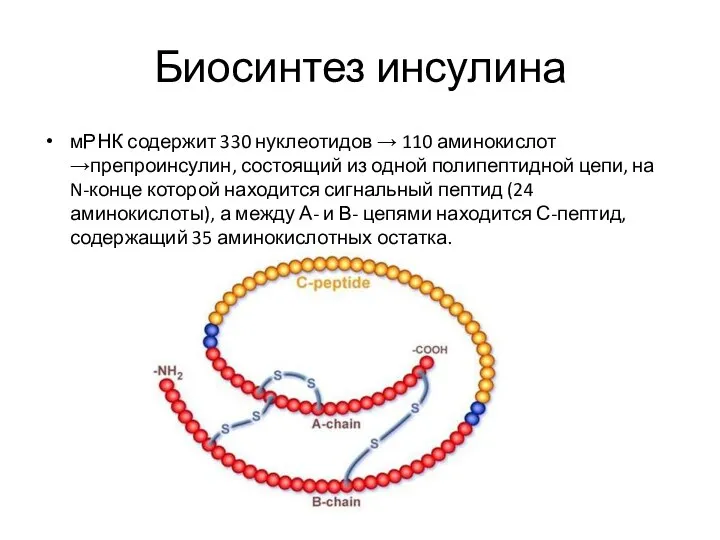
- 3. Биосинтез инсулина мРНК содержит 330 нуклеотидов → 110 аминокислот →препроинсулин, состоящий из одной полипептидной цепи, на
- 4. Синтез препроинсулина происходит на полирибосомах, связанных с эндоплазматическим ретикулумом. Препроинсулин проникает в люмен ретикулума, где от
- 5. В аппарате Гольджи и секреторных гранулах происходит превращение проинсулина в инсулин. В этом участвуют две эндопептидазы:
- 7. При стимуляции глюкозой инсулин быстро освобождается из секреторных гранул, а количество инсулиновой мРНК в клетке возрастает
- 8. Глюкоза стимулирует синтез инсулиновой мРНК при продолжительной инкубации (2 - 72 часа). При инкубации в течение
- 9. Секреция инсулина и С-пептида происходит путем экзоцитоза. Инсулин в растворе легко образует олигомерные агрегаты, преимущественно димеры
- 10. Глюкоза, аминокислоты (особенно аргинин и лизин), кетоновые тела и жирные кислоты в физиологических концентрациях стимулируют секрецию
- 12. Распад инсулина Время полураспада инсулина в крови - 3-10 мин, С-пептида - около 30 мин. Кровь
- 14. Скачать презентацию
Слайд 2Структура молекулы инсулина:
Молекула инсулина построена из двух пептидных цепей: цепь А содержит
Структура молекулы инсулина:
Молекула инсулина построена из двух пептидных цепей: цепь А содержит

Слайд 3Биосинтез инсулина
мРНК содержит 330 нуклеотидов → 110 аминокислот →препроинсулин, состоящий из одной
Биосинтез инсулина
мРНК содержит 330 нуклеотидов → 110 аминокислот →препроинсулин, состоящий из одной
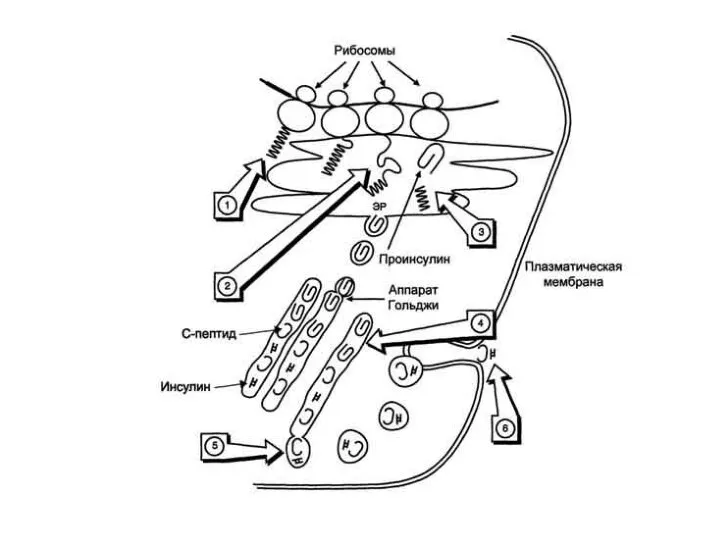
Слайд 4Синтез препроинсулина происходит на полирибосомах, связанных с эндоплазматическим ретикулумом.
Препроинсулин проникает в
Синтез препроинсулина происходит на полирибосомах, связанных с эндоплазматическим ретикулумом.
Препроинсулин проникает в

Слайд 5В аппарате Гольджи и секреторных гранулах происходит превращение проинсулина в инсулин.
В
В аппарате Гольджи и секреторных гранулах происходит превращение проинсулина в инсулин.
В

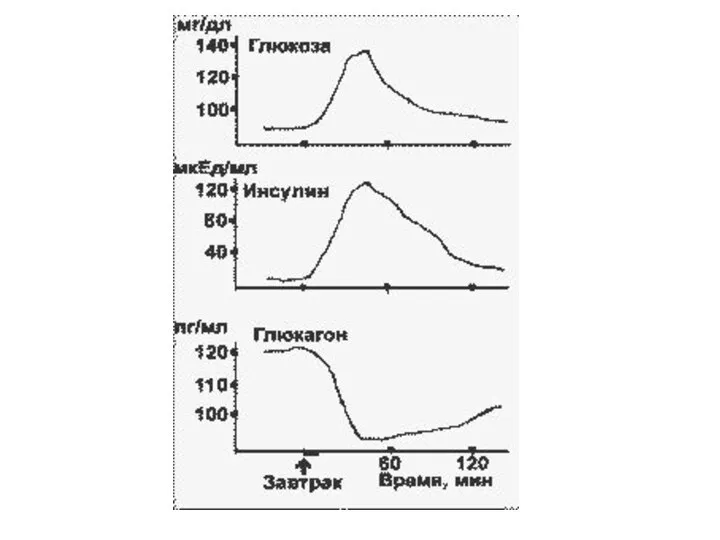
Слайд 7При стимуляции глюкозой инсулин быстро освобождается из секреторных гранул, а количество инсулиновой
При стимуляции глюкозой инсулин быстро освобождается из секреторных гранул, а количество инсулиновой

Слайд 8Глюкоза стимулирует синтез инсулиновой мРНК при продолжительной инкубации (2 - 72 часа).
Глюкоза стимулирует синтез инсулиновой мРНК при продолжительной инкубации (2 - 72 часа).

Слайд 9Секреция инсулина и С-пептида происходит путем экзоцитоза. Инсулин в растворе легко образует
Секреция инсулина и С-пептида происходит путем экзоцитоза. Инсулин в растворе легко образует

Слайд 10Глюкоза, аминокислоты (особенно аргинин и лизин), кетоновые тела и жирные кислоты в
Глюкоза, аминокислоты (особенно аргинин и лизин), кетоновые тела и жирные кислоты в

Слайд 12Распад инсулина
Время полураспада инсулина в крови - 3-10 мин, С-пептида - около
Распад инсулина
Время полураспада инсулина в крови - 3-10 мин, С-пептида - около

 Сурок Байбак
Сурок Байбак День наблюдения птиц
День наблюдения птиц Розробка та аналіз біогазового комплексу, що використовує біомасу рослин
Розробка та аналіз біогазового комплексу, що використовує біомасу рослин Весна. Перелетные птицы
Весна. Перелетные птицы Законы Менделя
Законы Менделя Презентация по биологии на тему_ _Жизненный цикл мхов на примере кукушкина льна и сфаг
Презентация по биологии на тему_ _Жизненный цикл мхов на примере кукушкина льна и сфаг Жизнь в морях и океанах
Жизнь в морях и океанах Органы дыхания и газообмен. 7 класс
Органы дыхания и газообмен. 7 класс Плауны, хвощи, папоротники
Плауны, хвощи, папоротники Комунікація тварин
Комунікація тварин prezentatsia_Obschaya_TsNS_2018
prezentatsia_Obschaya_TsNS_2018 Фотосинтез как ассимиляционный процесс. Значение и механизм фотосинтеза
Фотосинтез как ассимиляционный процесс. Значение и механизм фотосинтеза Грибы
Грибы Вегетативные ядра в ЦНС. Высшие вегетативные центры
Вегетативные ядра в ЦНС. Высшие вегетативные центры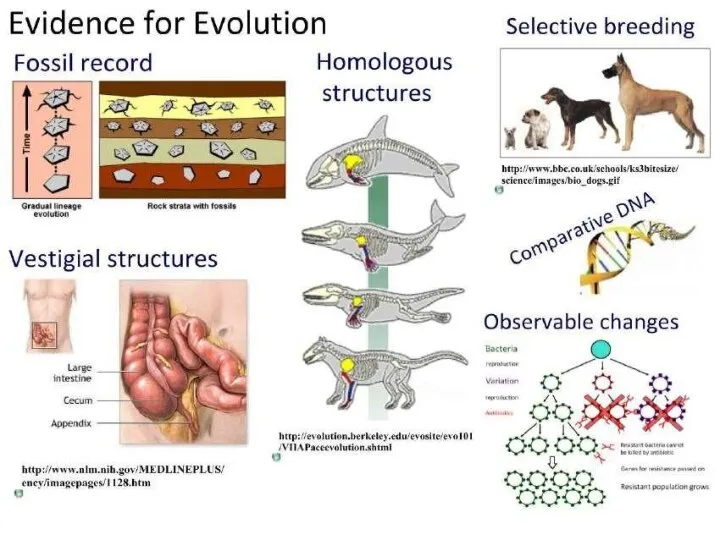 Evidence for Evolution
Evidence for Evolution Биологичекая защита. Энтомофаги вредителей плодово-ягодных культур
Биологичекая защита. Энтомофаги вредителей плодово-ягодных культур Бурый медведь
Бурый медведь 11.7 Хромомная теория. Генетика пола. Наследование, сцепленное с полом
11.7 Хромомная теория. Генетика пола. Наследование, сцепленное с полом Пока дышу – живу. Модедь Дондерса
Пока дышу – живу. Модедь Дондерса Промысловые рыбы. Их использование и охрана
Промысловые рыбы. Их использование и охрана Селекция в животноводстве
Селекция в животноводстве Координация и регуляция
Координация и регуляция Класс ленточные черви
Класс ленточные черви Движение крови в организме. Органы кровообращения
Движение крови в организме. Органы кровообращения Выявление доминирующего полушария у учеников старшей школы гимназии №2
Выявление доминирующего полушария у учеников старшей школы гимназии №2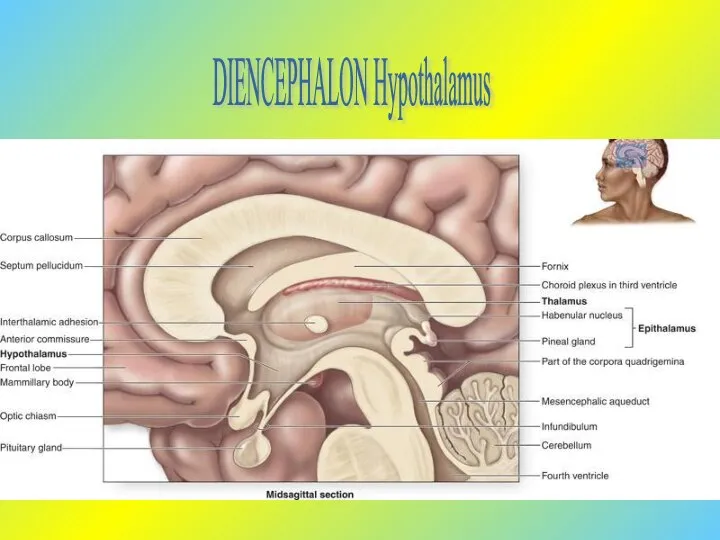 Гипоталамус
Гипоталамус Сравнение свойств организма человека и животных
Сравнение свойств организма человека и животных Устойчивость экосистем
Устойчивость экосистем