- Классификация белков

Содержание
- 2. В любом живом организме содержатся тысячи белков, выполняющих разнообразные функции. Чтобы дать представление о многообразии белков,
- 4. ПЕПТИДЫ, природные или синтетические соединения, молекулы которых построены из остатков α-аминокислот, соединенных между собой пептидными (амидными)
- 5. Историческая справка Впервые пептиды были выделены из ферментативных гидролизатов белков. Термин "пептиды" предложен Эмиль Фишером. Первый
- 6. Историческая справка С применением N-карбобензоксиаминокислот были впервые синтезированы природные пептиды (глукатион, карнозин и др.). Важное достижение
- 7. Историческая справка Успехи синтетической химии пептидов были подготовлены достижениями в области разработки таких методов разделения, очистки
- 8. Белки либо протеины количественно доминируют над всеми иными макромолекулами живой клетки. Белки принимают участие во всех
- 9. Пептидные взаимосвязи возникают при взаимодействии α -аминогруппы одной аминокислоты с α -карбоксильной категорией иной аминокислоты: Пептидная
- 11. Функциональные качества белков ориентируются их конформацией, т.е. месторасположением полипептидной цепи в месте. Уникальность конформации для любого
- 12. По двум сторонам твердой пептидной взаимосвязи вполне вероятно вращение: y и j -углы, определяющие вращение что
- 13. Пептидная цепь покупает не свободную, а жестко явную конформацию, фиксируемую водородными взаимосвязями. Известны некоторое количество приемов
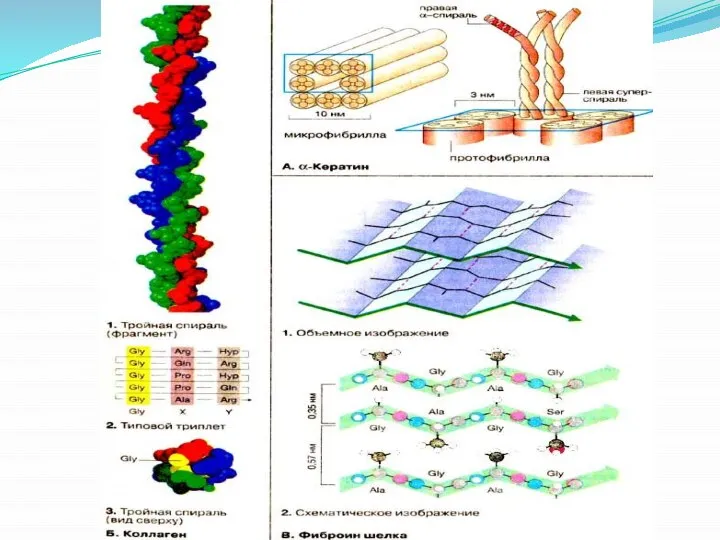
- 14. Содержание α -спиралей и β -структур в различных белках различно: у фибриллярных белков - лишь α
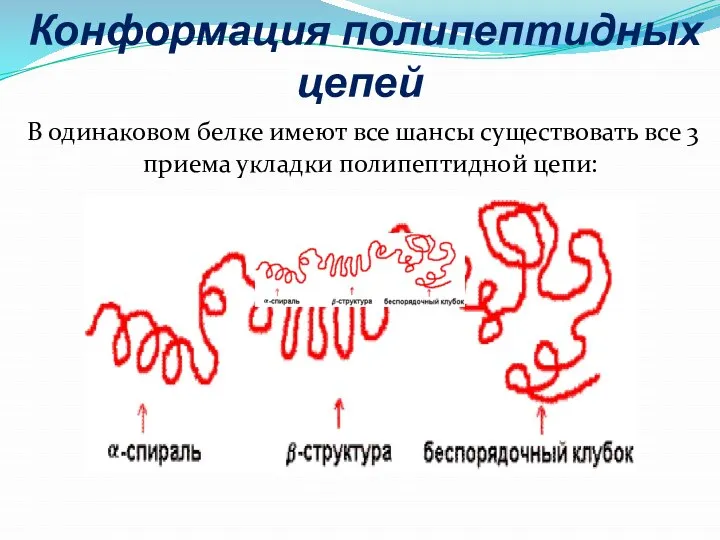
- 15. Конформация полипептидных цепей В одинаковом белке имеют все шансы существовать все 3 приема укладки полипептидной цепи:
- 16. Третичная текстура глобулярных белков предполагает ориентацию в месте полипептидной цепи, содержащей α -спирали, β-структуры и участки
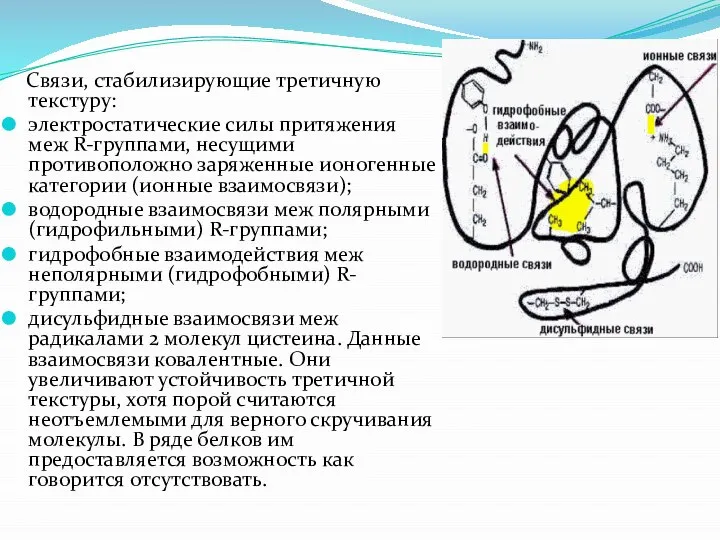
- 17. Связи, стабилизирующие третичную текстуру: электростатические силы притяжения меж R-группами, несущими противоположно заряженные ионогенные категории (ионные взаимосвязи);
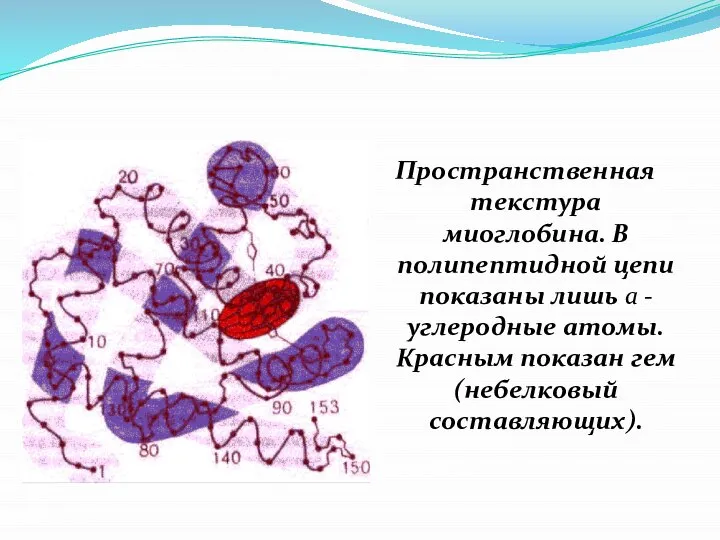
- 18. Пространственная текстура миоглобина. В полипептидной цепи показаны лишь a -углеродные атомы. Красным показан гем (небелковый составляющих).
- 19. Денатурация. Перемена температуры, ионной силы, рН, и еще обработка органическими либо какими-либо дестабилизирующими агентами имеет возможность
- 20. Ренативация При явных условиях денатурированный белок быть может ренативирован. Данное случается при удалении денатурирующего либо дестабилизирующего
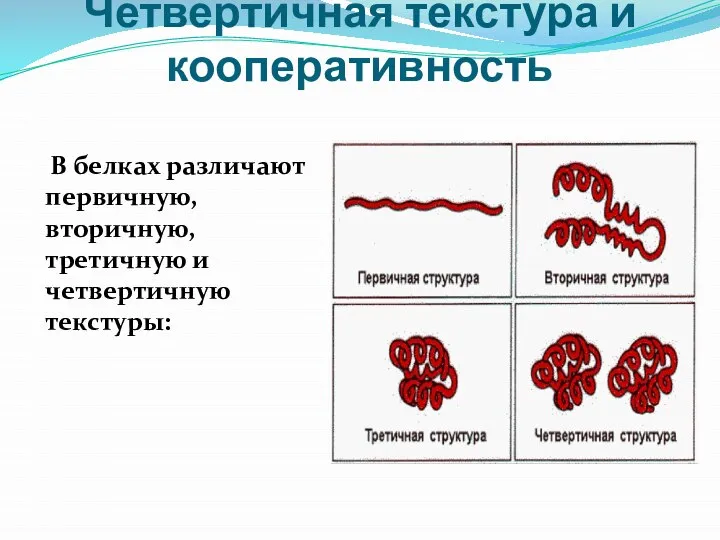
- 21. Четвертичная текстура и кооперативность В белках различают первичную, вторичную, третичную и четвертичную текстуры:
- 23. Скачать презентацию

Слайд 4 ПЕПТИДЫ, природные или синтетические соединения, молекулы которых построены из остатков α-аминокислот,
ПЕПТИДЫ, природные или синтетические соединения, молекулы которых построены из остатков α-аминокислот,

Слайд 5Историческая справка
Впервые пептиды были выделены из ферментативных гидролизатов белков. Термин "пептиды"
Историческая справка
Впервые пептиды были выделены из ферментативных гидролизатов белков. Термин "пептиды"

Слайд 6Историческая справка
С применением N-карбобензоксиаминокислот были впервые синтезированы природные пептиды (глукатион, карнозин
Историческая справка
С применением N-карбобензоксиаминокислот были впервые синтезированы природные пептиды (глукатион, карнозин

Слайд 7Историческая справка
Успехи синтетической химии пептидов были подготовлены достижениями в области разработки таких
Историческая справка
Успехи синтетической химии пептидов были подготовлены достижениями в области разработки таких

Слайд 8 Белки либо протеины количественно доминируют над всеми иными макромолекулами живой клетки.
Белки либо протеины количественно доминируют над всеми иными макромолекулами живой клетки.

Слайд 9 Пептидные взаимосвязи возникают при взаимодействии α -аминогруппы одной аминокислоты с α
Пептидные взаимосвязи возникают при взаимодействии α -аминогруппы одной аминокислоты с α

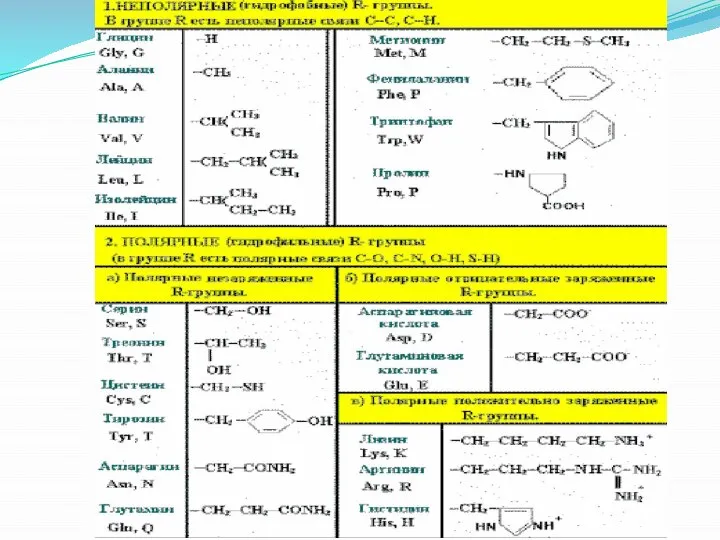
Слайд 11 Функциональные качества белков ориентируются их конформацией, т.е. месторасположением полипептидной цепи в
Функциональные качества белков ориентируются их конформацией, т.е. месторасположением полипептидной цепи в

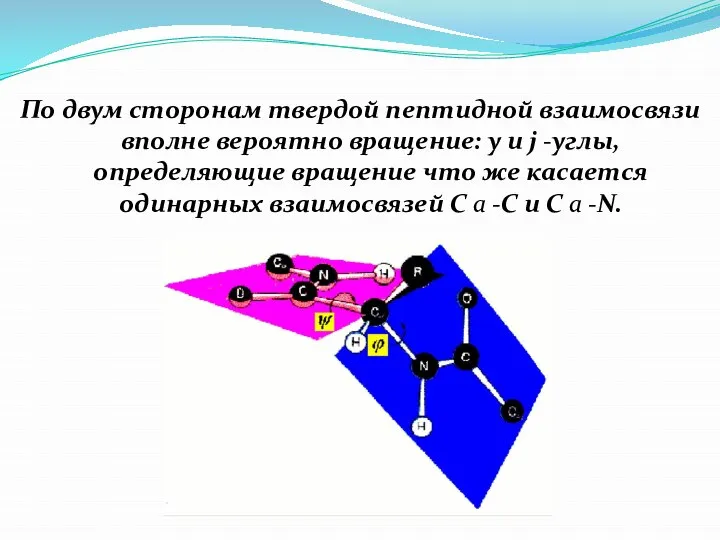
Слайд 12По двум сторонам твердой пептидной взаимосвязи вполне вероятно вращение: y и j
По двум сторонам твердой пептидной взаимосвязи вполне вероятно вращение: y и j
Слайд 13 Пептидная цепь покупает не свободную, а жестко явную конформацию, фиксируемую водородными
Пептидная цепь покупает не свободную, а жестко явную конформацию, фиксируемую водородными

Слайд 14 Содержание α -спиралей и β -структур в различных белках различно: у
Содержание α -спиралей и β -структур в различных белках различно: у
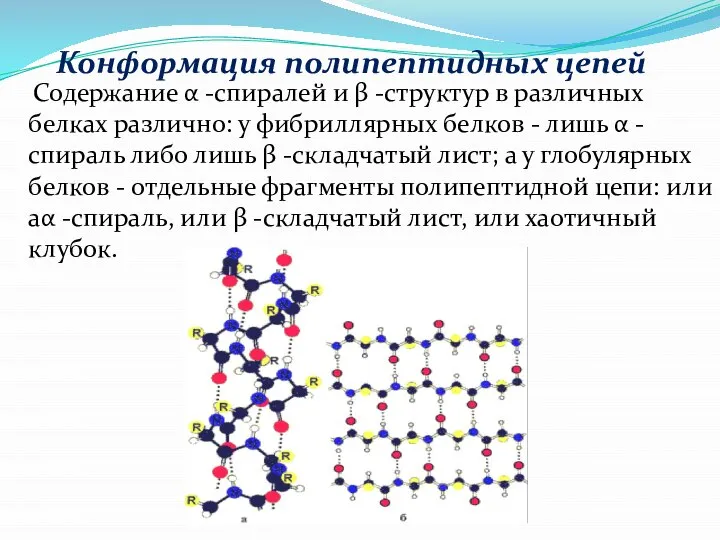
Слайд 15 Конформация полипептидных цепей
В одинаковом белке имеют все шансы существовать все 3
Конформация полипептидных цепей
В одинаковом белке имеют все шансы существовать все 3
Слайд 16 Третичная текстура глобулярных белков предполагает ориентацию в месте полипептидной цепи, содержащей
Третичная текстура глобулярных белков предполагает ориентацию в месте полипептидной цепи, содержащей

Слайд 17 Связи, стабилизирующие третичную текстуру:
электростатические силы притяжения меж R-группами, несущими противоположно
Связи, стабилизирующие третичную текстуру:
электростатические силы притяжения меж R-группами, несущими противоположно
Слайд 18Пространственная текстура миоглобина. В полипептидной цепи показаны лишь a -углеродные атомы. Красным
Пространственная текстура миоглобина. В полипептидной цепи показаны лишь a -углеродные атомы. Красным
Слайд 19Денатурация.
Перемена температуры, ионной силы, рН, и еще обработка органическими либо какими-либо
Денатурация.
Перемена температуры, ионной силы, рН, и еще обработка органическими либо какими-либо

Слайд 20 Ренативация
При явных условиях денатурированный белок быть может ренативирован. Данное случается
Ренативация
При явных условиях денатурированный белок быть может ренативирован. Данное случается

Слайд 21Четвертичная текстура и кооперативность
В белках различают первичную, вторичную, третичную и четвертичную
Четвертичная текстура и кооперативность
В белках различают первичную, вторичную, третичную и четвертичную
 Неполное доминирование. Дигибридное скрещивание
Неполное доминирование. Дигибридное скрещивание Пролапс слезной железы третьего века
Пролапс слезной железы третьего века Строение дыхательной системы
Строение дыхательной системы Павлины
Павлины Шаблон теста по биологии
Шаблон теста по биологии Понятие о ДНК как хранителе и носителе генетического материала. 7 класс
Понятие о ДНК как хранителе и носителе генетического материала. 7 класс Ферменты. История изучения, функции Ферменты
Ферменты. История изучения, функции Ферменты Физиология питания
Физиология питания Змеи Удмуртской республики
Змеи Удмуртской республики Амфібії. Ознаки пристосування земноводних до життя на суходолі
Амфібії. Ознаки пристосування земноводних до життя на суходолі Красная Книга Тывы. Класс Птицы. 5
Красная Книга Тывы. Класс Птицы. 5 Рабочая тетрадь. Биология 7 класс
Рабочая тетрадь. Биология 7 класс Экологические системы
Экологические системы Иммуноферментный анализ (ИФА)
Иммуноферментный анализ (ИФА) Растения степи. Ребусы
Растения степи. Ребусы Значение выделения в жизни организмов. Продукты выделения
Значение выделения в жизни организмов. Продукты выделения Размножение растений
Размножение растений Гаметогенез. Сперматогенез. Оогенез
Гаметогенез. Сперматогенез. Оогенез Тест Общие признаки птиц
Тест Общие признаки птиц Азотистый обмен. Общие пути
Азотистый обмен. Общие пути Регуляция дыхания
Регуляция дыхания Мидии и багрянки
Мидии и багрянки Протисты. Общая характеристика
Протисты. Общая характеристика Строение клетки
Строение клетки Класс Млекопитающие
Класс Млекопитающие Обмен веществ
Обмен веществ Полёты птиц
Полёты птиц Рептилии. 7 класс
Рептилии. 7 класс