- Линейная хромосома

Содержание
- 2. Хромосомы – нуклеопротеиновые тела, в которых хранится, передается потомству и реализуется наследственная информация (Жимулев) . Первые
- 3. Прокариоты (археи и бактерии, в том числе митохондрии и пластиды, постоянно обитающие в клетках большинства эукариот)
- 4. Особенности репликации линейных геномов Кольцевые замкнутые геномы характерны для многих бактерий, их плазмид и некоторых вирусов.
- 5. Рис. 1. Проблема репликации отстающей цепи ДНК линейных хромосом. 1 – отстающая цепь реплицирующейся ДНК синтезируется
- 6. Линейные хромосомы бактерий. Афоризм Жака Моно (французский биохимик и микробиолог): "То, что верно для E. coli,
- 7. Если у вирусов синтез ДНК инициируется на конце хромосомы с использованием в качестве затравки TP (концевые
- 8. Рис. 2. Модель достройки теломерных участков хромосом и плазмид Streptomyces а – структура теломеры после репликации:
- 9. В соответствии с первой моделью одноцепочечный участок теломеры, содержащий TIR-последовательность, образует концевую шпильку путем комплементарных взаимодействий
- 10. Неизвестно, как много форм линейных бактериальных хромосом существует в природе. Не изучены и таксономические проблемы, связанные
- 11. Список литературы Филипченко Ю. А. Генетика. Жимулев И.Ф. Общая и молекулярная генетика Коряков Д. Е., Жимулев
- 13. Скачать презентацию
Слайд 2Хромосомы – нуклеопротеиновые тела, в которых хранится, передается потомству и реализуется наследственная
Хромосомы – нуклеопротеиновые тела, в которых хранится, передается потомству и реализуется наследственная

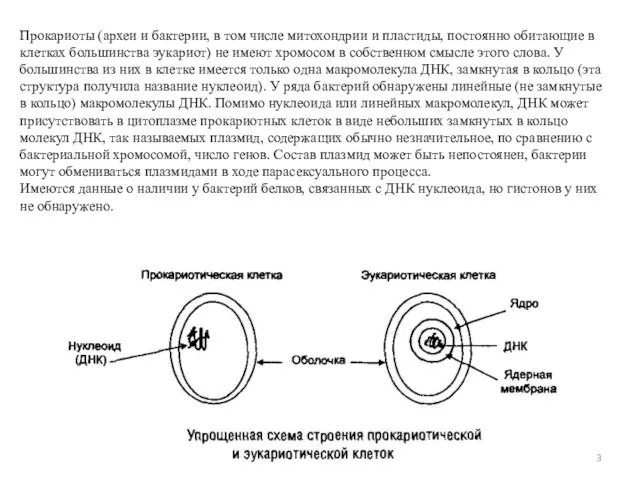
Слайд 3Прокариоты (археи и бактерии, в том числе митохондрии и пластиды, постоянно обитающие в клетках большинства эукариот) не имеют хромосом
Прокариоты (археи и бактерии, в том числе митохондрии и пластиды, постоянно обитающие в клетках большинства эукариот) не имеют хромосом
Слайд 4Особенности репликации линейных геномов
Кольцевые замкнутые геномы характерны для многих бактерий, их плазмид
Особенности репликации линейных геномов
Кольцевые замкнутые геномы характерны для многих бактерий, их плазмид

Слайд 5Рис. 1. Проблема репликации отстающей цепи ДНК линейных хромосом.
1 – отстающая цепь реплицирующейся ДНК
Рис. 1. Проблема репликации отстающей цепи ДНК линейных хромосом.
1 – отстающая цепь реплицирующейся ДНК

Слайд 6Линейные хромосомы бактерий.
Афоризм Жака Моно (французский биохимик и микробиолог): "То, что верно
Линейные хромосомы бактерий.
Афоризм Жака Моно (французский биохимик и микробиолог): "То, что верно

Слайд 7Если у вирусов синтез ДНК инициируется на конце хромосомы с использованием в качестве
Если у вирусов синтез ДНК инициируется на конце хромосомы с использованием в качестве

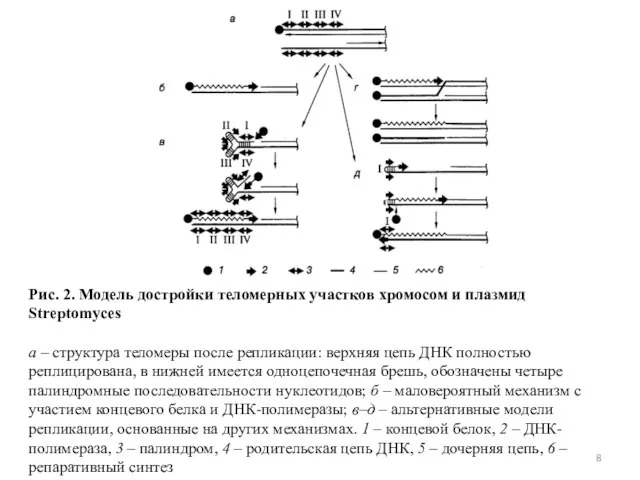
Слайд 8Рис. 2. Модель достройки теломерных участков хромосом и плазмид Streptomyces
а – структура теломеры после репликации:
Рис. 2. Модель достройки теломерных участков хромосом и плазмид Streptomyces а – структура теломеры после репликации:
Слайд 9В соответствии с первой моделью одноцепочечный участок теломеры, содержащий TIR-последовательность, образует концевую
В соответствии с первой моделью одноцепочечный участок теломеры, содержащий TIR-последовательность, образует концевую

Слайд 10Неизвестно, как много форм линейных бактериальных хромосом существует в природе. Не изучены
Неизвестно, как много форм линейных бактериальных хромосом существует в природе. Не изучены

Слайд 11Список литературы
Филипченко Ю. А. Генетика. Жимулев И.Ф. Общая и молекулярная генетика
Коряков Д. Е., Жимулев И. Ф. Хромосомы.
Список литературы
Филипченко Ю. А. Генетика. Жимулев И.Ф. Общая и молекулярная генетика
Коряков Д. Е., Жимулев И. Ф. Хромосомы.

 Формы естественного отбора
Формы естественного отбора Органы чувств
Органы чувств Биотехнология - использование живых систем, клеток, организмов для практических нужд человека
Биотехнология - использование живых систем, клеток, организмов для практических нужд человека Понятия проницаемости и селективности. Типы мембранного транспорта
Понятия проницаемости и селективности. Типы мембранного транспорта Сознание и мышление человека. Речь
Сознание и мышление человека. Речь Рептилии Саратовской области
Рептилии Саратовской области Общая характеристика класса Млекопитающие
Общая характеристика класса Млекопитающие Подтип Трахейные. Класс Насекомые
Подтип Трахейные. Класс Насекомые Грибы рода Aspergillus
Грибы рода Aspergillus Осенние листья в дендрарии Санкт-Петербургского ботанического сада
Осенние листья в дендрарии Санкт-Петербургского ботанического сада Сердечно- сосудистые заболевания. К\в Человек и его здоровье
Сердечно- сосудистые заболевания. К\в Человек и его здоровье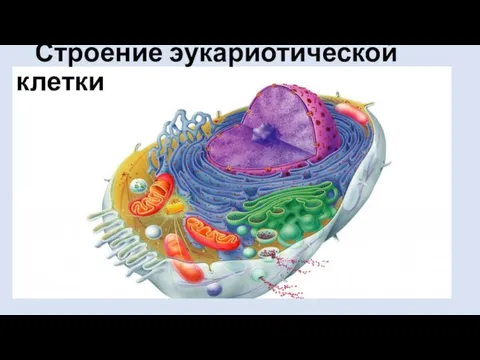 Строение эукариотической клетки
Строение эукариотической клетки Презентация на тему Экология и здоровье человека
Презентация на тему Экология и здоровье человека  Практическая работа
Практическая работа Вегетативные органы растений. Ярмарка знаний
Вегетативные органы растений. Ярмарка знаний Белка
Белка Мир птиц
Мир птиц Семейства цветковых растений
Семейства цветковых растений Биология – наука о жизни
Биология – наука о жизни Жизненные формы растений
Жизненные формы растений Строение и функции лимфатической системы, движение лимфы в организме
Строение и функции лимфатической системы, движение лимфы в организме Влияние домашнего содержания на развитие амфибии - амбистомы
Влияние домашнего содержания на развитие амфибии - амбистомы Зелёная морская черепах
Зелёная морская черепах Занимательная экология
Занимательная экология Биомолекулы: ДНК
Биомолекулы: ДНК Презентация на тему Витамины 10 класс
Презентация на тему Витамины 10 класс  Тела, вещества, частицы. Урок окружающего мира, 3 класс
Тела, вещества, частицы. Урок окружающего мира, 3 класс Строение цветка
Строение цветка