- Механизмы проведения возбуждения

Содержание
- 2. Законы проведения возбуждения по нервам. 1. Закон физиологической непрерывности. Перерезка, перевязка, а также любое другое воздействие,
- 3. Классификация нервных волокон. Как только в какой-либо точке нервного или мышечного волокна возникает ПД и этот
- 4. Скорость проведения возбуждения в нервных волокнах разного диаметра
- 5. Мембрана в области перехвата специализирована для генерации возбуждения: количество Na+-каналов на единицу площади здесь примерно в
- 6. Классификация нервных волокон. Электрический ответ целого нерва является алгебраической суммой ПД отдельных его нервных волокон. Поэтому,
- 7. 3.2. НЕРВНО-МЫШЕЧНЫЙ СИНАПС: СТРОЕНИЕ, МЕХАНИЗМ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ, ОСОБЕННОСТИ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ В СИНАПСЕ ПО СРАВНЕНИЮ С НЕРВНЫМ
- 8. Роль медиатора заключается в повышении проницаемости постсинаптической мембраны для ионов Na+. Возникновение потока ионов Na+ из
- 9. Эта проницаемость возможна благодаря тому, что в результате деполяризации пресинаптической мембраны открываются ее кальциевые каналы. Ион
- 10. Синаптическая передача возбуждения по сравнению с распространением потенциала действия имеет рад свойств: 1) наличие медиатора в
- 12. Скачать презентацию
Слайд 2Законы проведения возбуждения по нервам.
1. Закон физиологической непрерывности. Перерезка, перевязка,
Законы проведения возбуждения по нервам.
1. Закон физиологической непрерывности. Перерезка, перевязка,

Слайд 3 Классификация нервных волокон. Как только в какой-либо точке нервного или мышечного
Классификация нервных волокон. Как только в какой-либо точке нервного или мышечного

Слайд 4
Скорость проведения возбуждения в нервных волокнах разного диаметра
Скорость проведения возбуждения в нервных волокнах разного диаметра

Слайд 5Мембрана в области перехвата специализирована для генерации возбуждения: количество Na+-каналов на единицу
Мембрана в области перехвата специализирована для генерации возбуждения: количество Na+-каналов на единицу

Слайд 6Классификация нервных волокон. Электрический ответ целого нерва является алгебраической суммой ПД отдельных
Классификация нервных волокон. Электрический ответ целого нерва является алгебраической суммой ПД отдельных

Слайд 73.2. НЕРВНО-МЫШЕЧНЫЙ СИНАПС: СТРОЕНИЕ, МЕХАНИЗМ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ, ОСОБЕННОСТИ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ В СИНАПСЕ
3.2. НЕРВНО-МЫШЕЧНЫЙ СИНАПС: СТРОЕНИЕ, МЕХАНИЗМ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ, ОСОБЕННОСТИ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ В СИНАПСЕ
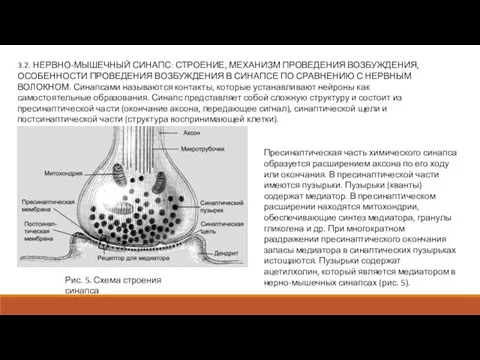
Слайд 8Роль медиатора заключается в повышении проницаемости постсинаптической мембраны для ионов Na+. Возникновение
Роль медиатора заключается в повышении проницаемости постсинаптической мембраны для ионов Na+. Возникновение

Слайд 9Эта проницаемость возможна благодаря тому, что в результате деполяризации пресинаптической мембраны открываются
Эта проницаемость возможна благодаря тому, что в результате деполяризации пресинаптической мембраны открываются

Слайд 10Синаптическая передача возбуждения по сравнению с распространением потенциала действия имеет рад свойств:
1)
Синаптическая передача возбуждения по сравнению с распространением потенциала действия имеет рад свойств:
1)

 Корма для лошадей
Корма для лошадей Интерактивный кроссворд по теме: Типы соцветий
Интерактивный кроссворд по теме: Типы соцветий Садовые ягоды
Садовые ягоды Голоса животных
Голоса животных Жизненные формы растений
Жизненные формы растений Современные представления о сенсорных системах
Современные представления о сенсорных системах Презентация на тему Деревья, которые живут в воде
Презентация на тему Деревья, которые живут в воде  Витамины. Виды витаминов
Витамины. Виды витаминов водоросли
водоросли Такие разные стебли
Такие разные стебли Щупальцевые, или Тентакуляты
Щупальцевые, или Тентакуляты Клетка. Строение
Клетка. Строение Игра Проверь себя. Знатоки насекомых
Игра Проверь себя. Знатоки насекомых Кишечнополостные
Кишечнополостные Витамин А Ретинол
Витамин А Ретинол Вегетативные органы растений. Побег
Вегетативные органы растений. Побег Тест по теме «Цветок» 6 класс
Тест по теме «Цветок» 6 класс Редкие и исчезающие виды животных, занесенные в Красную книгу Омской области
Редкие и исчезающие виды животных, занесенные в Красную книгу Омской области Хищничество и паразитизм
Хищничество и паразитизм Подцарство низшие растения. Лишайники
Подцарство низшие растения. Лишайники Сердечно-сосудистая система
Сердечно-сосудистая система Презентация на тему Строение и деятельность внутренних органов земноводных
Презентация на тему Строение и деятельность внутренних органов земноводных  Строение клетки. Ткани
Строение клетки. Ткани 5. Ядро, Организация генетического материала эукариот (3)
5. Ядро, Организация генетического материала эукариот (3) Prezentatsia_10_klass_Fotosintez
Prezentatsia_10_klass_Fotosintez Биология в естествознании
Биология в естествознании Дикие животные: леопард
Дикие животные: леопард изолируйся
изолируйся