- Мембранный транспорт

Содержание
- 2. отграничить живое от неживого организовать внутри клетки компартменты с различными свойствами контролировать проникновение в клетку и
- 3. Полупроницаемый барьер.
- 4. Формирование дефектных зон в липидном бислое лежит в основе неспецифической диффузии
- 5. Полупроницаемый барьер.
- 6. Содержание основных ионов (мМ) в клетках и внеклеточной жидкости различных животных, в сравнении с составом морской
- 8. Транспортные белки: помпы, каналы, транспортеры Первичный, активный Вторичный, пассивный
- 9. Мембранные помпы
- 10. Бактериородопсин Halobacterium halobium Энергия света
- 11. Glu204 Бактериородопсин
- 12. Родопсины. bacteriorhodopsin halorhodopsin sensory rhodopsin squid rhodopsin
- 13. Типы насосов (транспортных АТФаз) Семейство АТФаз Р-типа Семейство АТФаз F- и V-типа Семейство АТФаз АВС
- 14. Конформация О – связывающий центр доступен для АДФ и Фн Конформация L – связывание АДФ и
- 17. Конформация О – связывающий центр доступен для АДФ и Фн Конформация L – связывание АДФ и
- 18. Роль АТФаз V-типа в обеспечении функций различных клеток Передача сигнала, рецепторы
- 20. Р-АТФ-азы Β-аспартил-фосфат SERCA1
- 21. Модель активации мышечной Ca2+ ATPase, локализованной на мембране саркоплазматического ретикулума P-класс SERCA1
- 22. P-класс 12mM 145mM 4mM 139mM
- 23. Структура Na,K-АТФазы Нуклеотид-связывающий домен Активаторный домен 5 4 6 8 19 кДа фрагмент Фосфорилируемый домен 1
- 24. Модель рабоыты Na+/K+ ATPase на плазматической мембране.
- 25. Na,K –ATФаза ингибируется уабаином В 1785 году Визеринг начал использовать листья наперстянки для лечения сердечной недостаточности.
- 26. P4-ATPase, флиппаза
- 27. Distribution of PS throughout the cell.
- 28. ABC транспортеры (ATP-Binding “Cassette”)
- 29. ABC транспортеры (ATP-Binding “Cassette”)
- 30. Захват гистидина грамм-отрицательными бактериями permease
- 31. ABC транспортеры
- 33. Импорт
- 34. α-гемолизинE.coli протеазы E. chrysanthemi колицинVE.coli субтилин B.subtilis капсулярный полисахарид E.coli по M. Fathеt al., 1993 Оперон
- 36. ABC транспортеры эукариот
- 37. АВС-А1 и транспорт холестерола
- 38. Множественная лекарственная устойчивость
- 39. Множественная лекарственная устойчивость
- 42. MDR1 Pgp
- 43. MDR1
- 44. МНС I
- 45. Трансмембранный регулятор муковисцидоза CFTP-cystic fibrosis transmembrane regulator транспортер
- 46. Основные классы АТФаз в клетках и болезни, связанные с нарушением их функционирования
- 47. Белки-переносчики Движущими силами пассивного транспорта могут служить следующие градиенты: -концентрационный – для нейтральных молекул -электрохимический –
- 48. Мембранные переносчики major facilitator superfamily (MFS)
- 49. Концентрация S определяет скорость связывания и освобождения. Лимитирующая стадия – скорость конформационного изменения. +заряд 102-104 мол/сек
- 51. Унипорт GLUT1 эритроциты
- 52. Симпорт SGLT1 желудок
- 53. Симпорт GLUT2 Унипорт SGLT1
- 54. Симпорт
- 55. Примеры транспортеров симпортеров
- 56. Примеры транспортеров симпортеров
- 57. Антипорт
- 58. ATP/ADP Транслоказа Антипорт
- 59. Примеры транспортеров антипортеров
- 60. Na-Ca antiport Кардиомиоциты Антипорт
- 61. Анионный транспорт через мембрану эритроцита в системном и легочном капиллярах.
- 62. Закисление полости желудка париетальными клетками. P-class pump
- 63. Транспорт йонов и сахарозы в растительной вакуоли. Мембрана вакуоли содержит 2 типа протонных помп: V-class H+
- 64. Ингибиторы белков-переносчиков
- 65. Каналы
- 66. Функции йонных каналов: Регуляция водного обмена клетки: объём и тургор. 2. Регуляция pH: закисление и защелачивание.
- 67. По строению (родству их химического строения и происхождения образующих их белков). По строению (структуре) и по
- 68. Строение канала.
- 72. Схема строения ионного канала (на примере K+-канала Streptomyces lividans) Сформирован из 4 TVGYG-мотивов Открытие канала регулируется
- 74. Селективность Na 0,095нм К 0,13нм
- 75. Калиевый канал Селективность Натриевый канал
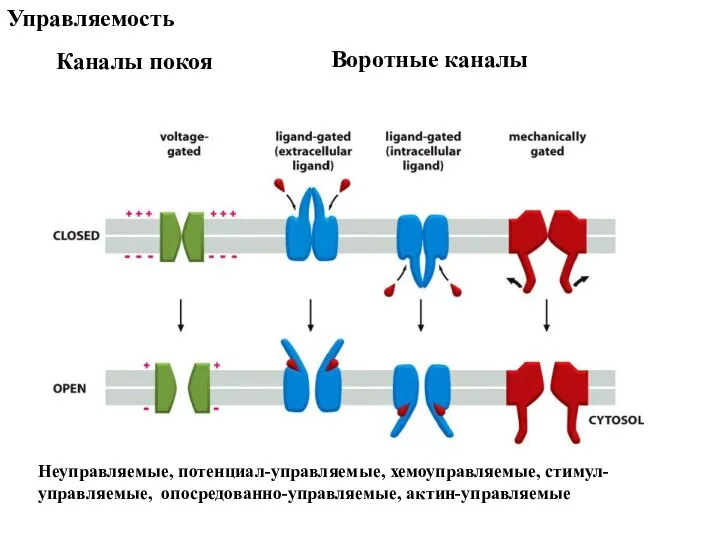
- 78. Каналы покоя Воротные каналы Управляемость Неуправляемые, потенциал-управляемые, хемоуправляемые, стимул-управляемые, опосредованно-управляемые, актин-управляемые
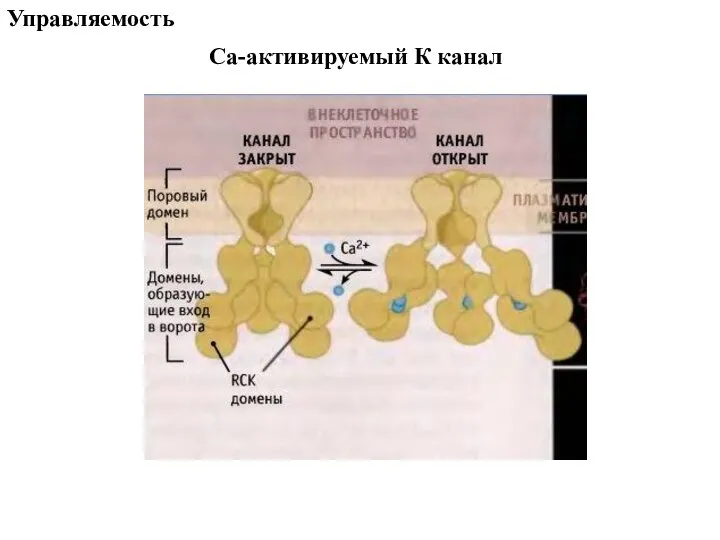
- 79. Са-активируемый К канал Управляемость
- 81. Гомоцистеин Глутамат N-метил-D-аспартат NMDA-рецептор – пример лиганд-управляемого ионного канала (Carlson, 1998) NR1 NR2
- 82. +2b субъединицы
- 83. Потенциал-управляемый К канал
- 84. 1. Открытое. Канал открыт и через него происходит перемещение ионов. 2. Закрытое. Канал закрыт и ионы
- 85. Активация Инактивация: Быстрая Медленная
- 86. Тетродотоксин: токсин рыбы фугу. Блокатор Na-каналов
- 87. Динофлагелляты: водоросли, ответственные за образование «красных приливов» Сакситоксин блокирует потенциал-зависимые Na-каналы
- 88. Тубокурарин (кураре) Тубокурарин – блокирует натриевый канал, чувствительный к ацетилхолину
- 89. Дендротоксин из яда черной мамбы Дендротоксин – блокатор потенциал-зависимых калиевых каналов
- 90. Ингибиторы каналов
- 91. Мутации в генах, кодирующих каналы, вызывают заболевания
- 92. Carbonyl cyanide-p-trifluoromethoxyphenylhydrazone (FCCP) Gramicidin Valinomycin Nigericin Ионофоры
- 93. Порины A trimeric OmpF channel in the lipid bilayer of the outer bacterial membrane is a
- 94. Порины
- 95. Рorin superfamily I The mitochondrial and plastid porin (MPP) Porin Superfamily II (MspA Superfamily) 1.B.24 -
- 96. Биогенез поринов.
- 97. N. meningitidis E. coli mitochondriа Биогенез поринов.
- 98. Порины в митохондриях voltage-dependent anion channel (VDAC)
- 99. Порины в митохондриях
- 100. Model of the mechanism of uniport transport by GLUT1, which is believed to shuttle between two
- 102. Скачать презентацию
Слайд 2отграничить живое от неживого
организовать внутри клетки компартменты с различными свойствами
контролировать
отграничить живое от неживого
организовать внутри клетки компартменты с различными свойствами
контролировать

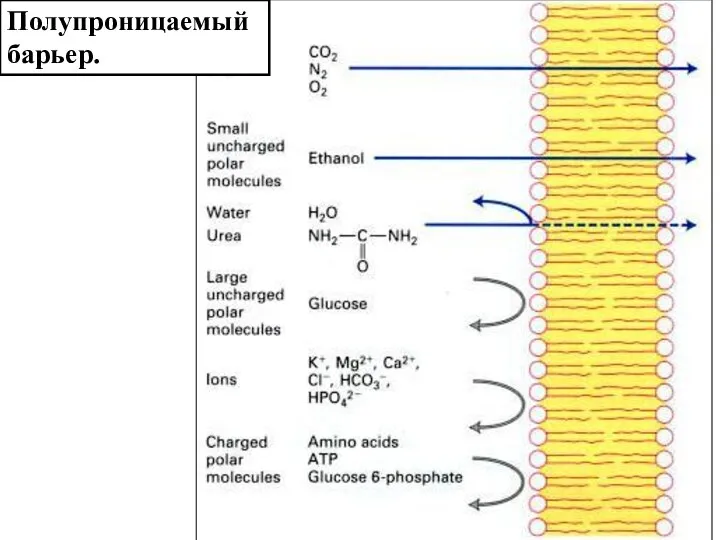
Слайд 3Полупроницаемый барьер.
Полупроницаемый барьер.
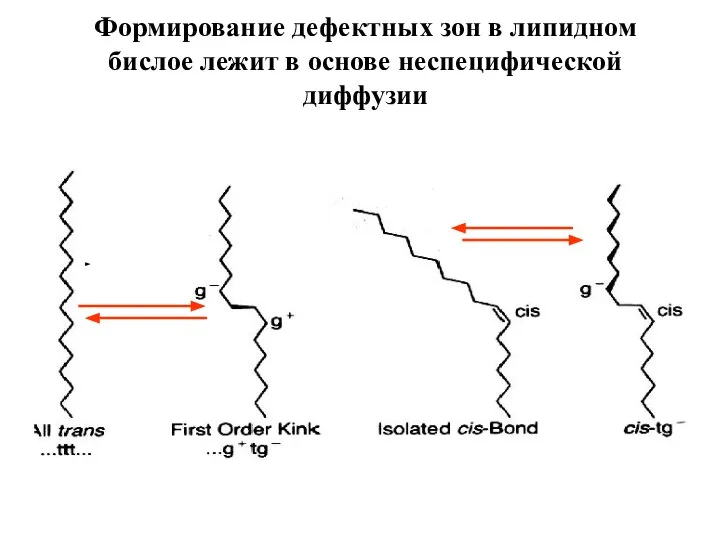
Слайд 4Формирование дефектных зон в липидном бислое лежит в основе неспецифической диффузии
Формирование дефектных зон в липидном бислое лежит в основе неспецифической диффузии
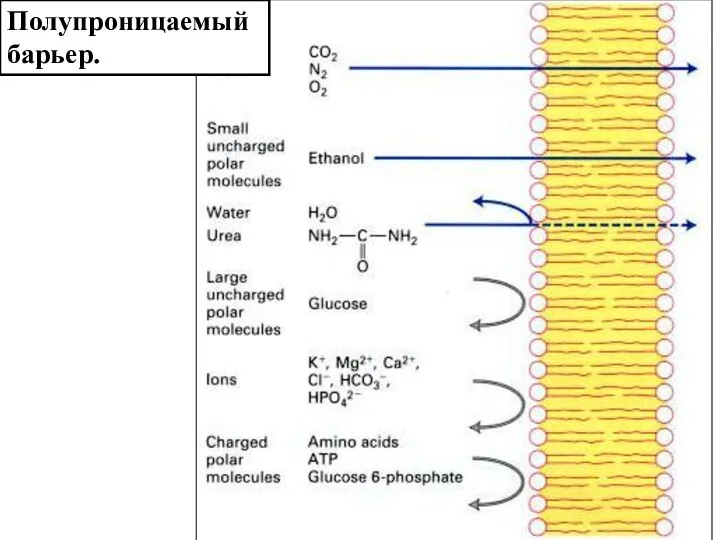
Слайд 5Полупроницаемый барьер.
Полупроницаемый барьер.
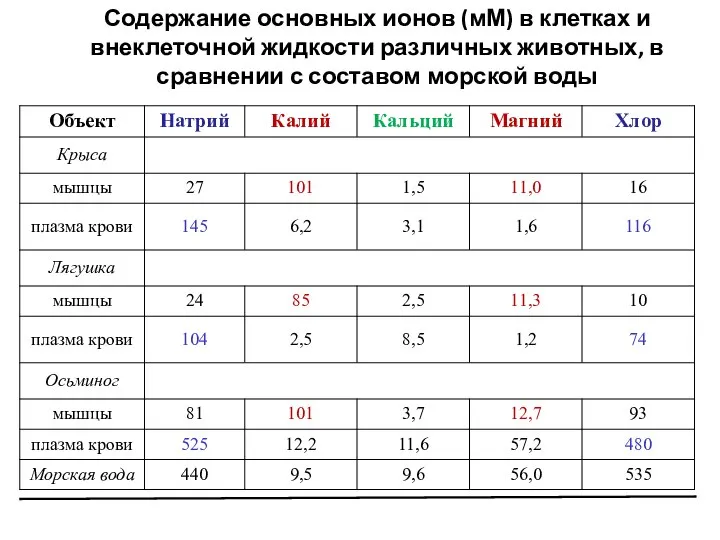
Слайд 6Содержание основных ионов (мМ) в клетках и внеклеточной жидкости различных животных, в
Содержание основных ионов (мМ) в клетках и внеклеточной жидкости различных животных, в
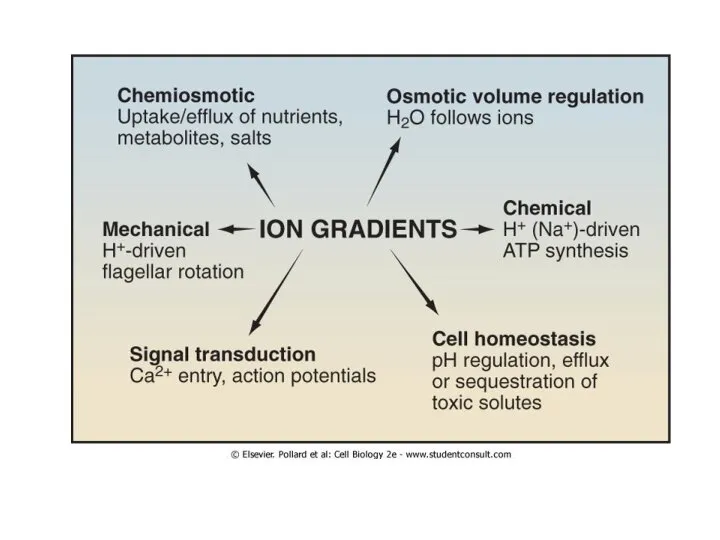
Слайд 8Транспортные белки: помпы, каналы, транспортеры
Первичный, активный
Вторичный, пассивный
Транспортные белки: помпы, каналы, транспортеры
Первичный, активный
Вторичный, пассивный
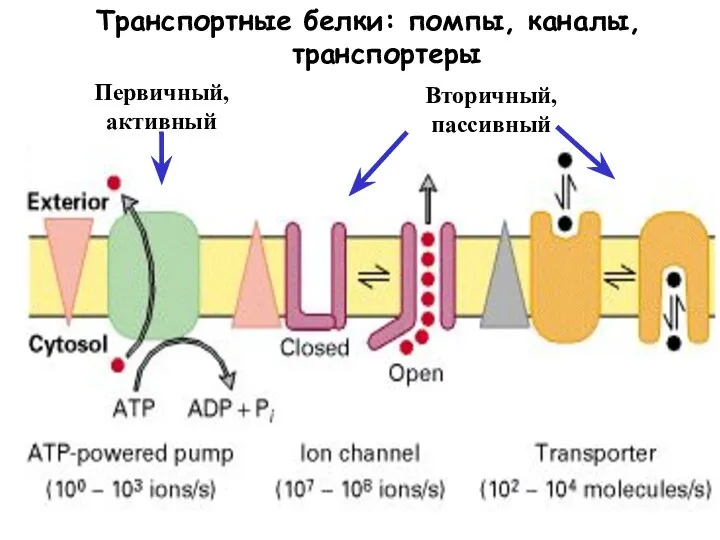
Слайд 9Мембранные помпы
Мембранные помпы
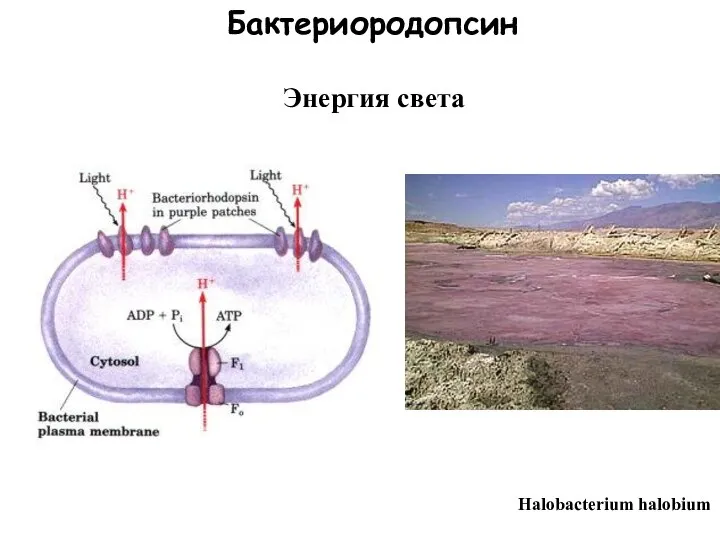
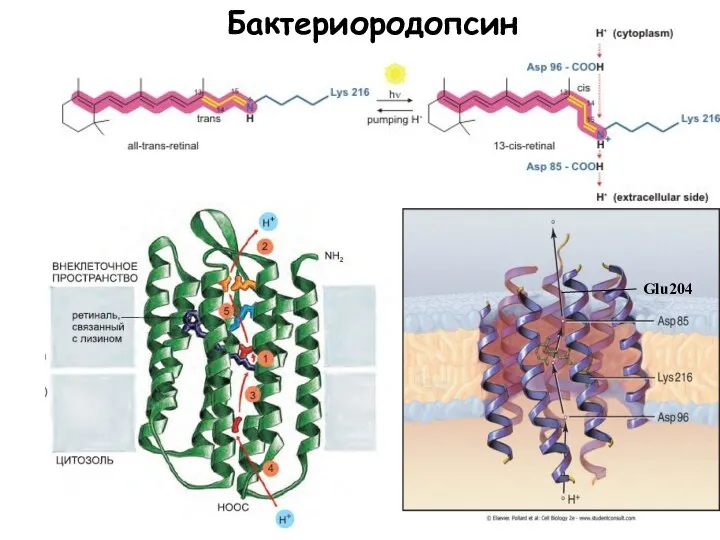
Слайд 10Бактериородопсин
Halobacterium halobium
Энергия света
Бактериородопсин
Halobacterium halobium
Энергия света
Слайд 11Glu204
Бактериородопсин
Glu204
Бактериородопсин
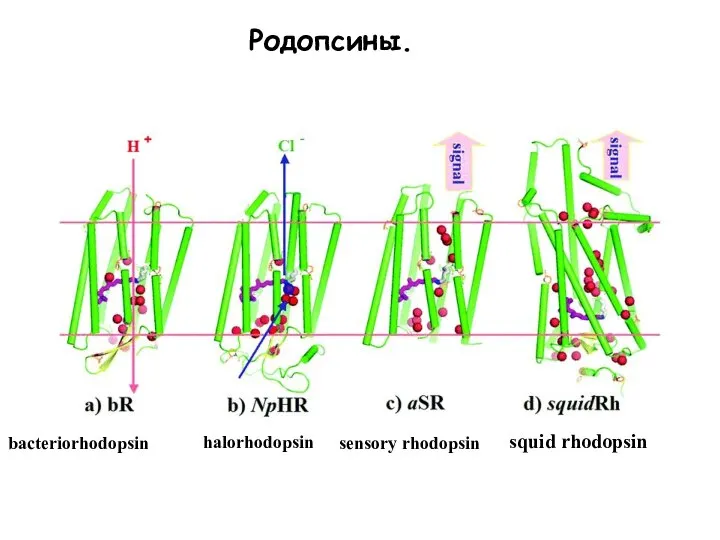
Слайд 12Родопсины.
bacteriorhodopsin
halorhodopsin
sensory rhodopsin
squid rhodopsin
Родопсины.
bacteriorhodopsin
halorhodopsin
sensory rhodopsin
squid rhodopsin
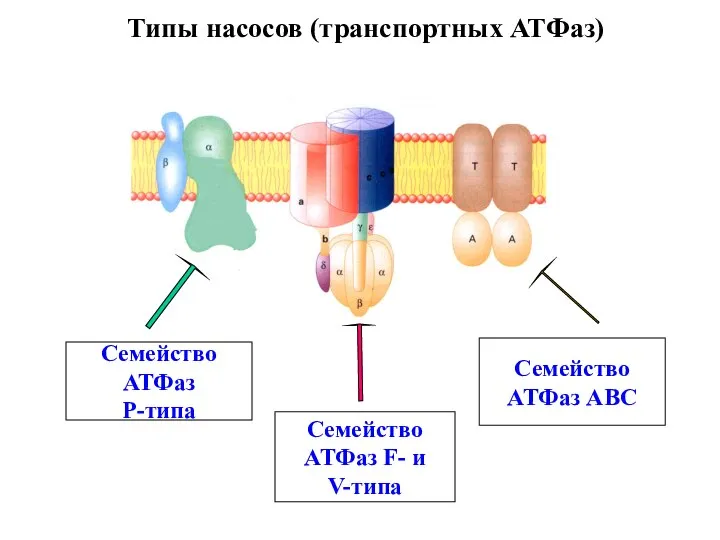
Слайд 13Типы насосов (транспортных АТФаз)
Семейство
АТФаз
Р-типа
Семейство
АТФаз F- и
V-типа
Семейство
АТФаз АВС
Типы насосов (транспортных АТФаз)
Семейство
АТФаз
Р-типа
Семейство
АТФаз F- и
V-типа
Семейство
АТФаз АВС
Слайд 14Конформация О – связывающий центр доступен для АДФ и Фн
Конформация L –
Конформация О – связывающий центр доступен для АДФ и Фн
Конформация L –
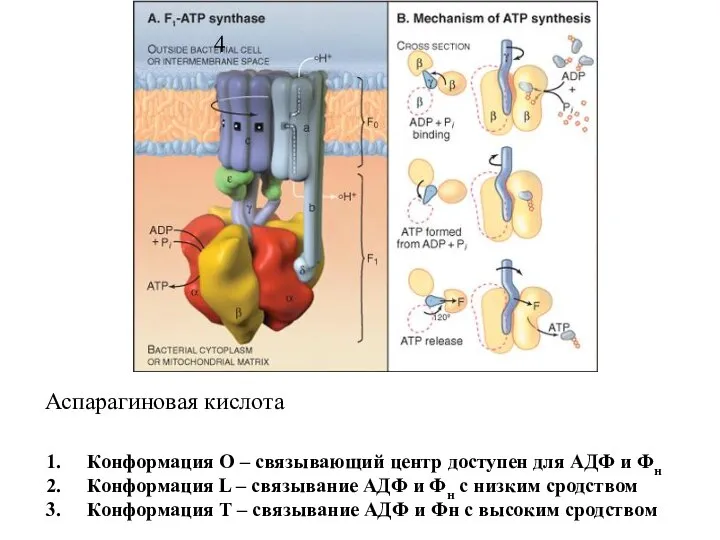
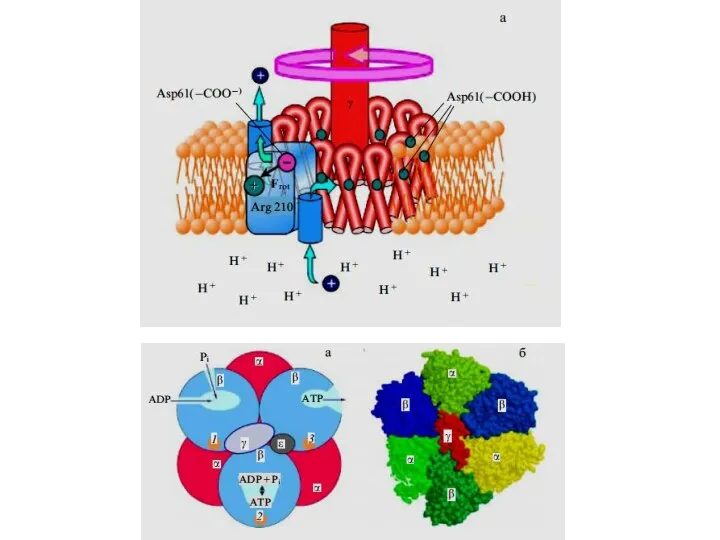
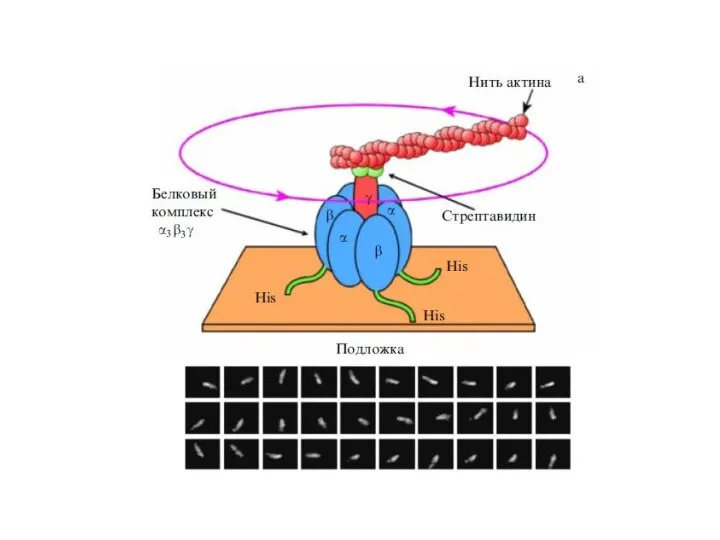
Слайд 17Конформация О – связывающий центр доступен для АДФ и Фн
Конформация L –
Конформация О – связывающий центр доступен для АДФ и Фн
Конформация L –
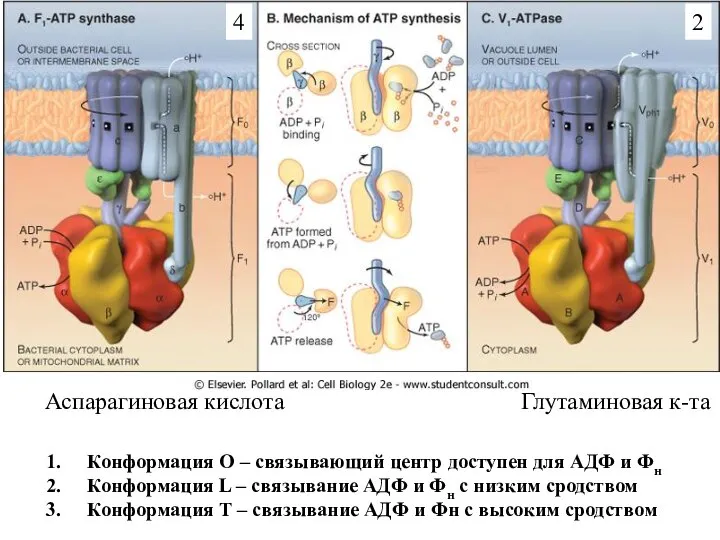
Слайд 18Роль АТФаз V-типа в обеспечении функций различных клеток
Передача сигнала, рецепторы
Роль АТФаз V-типа в обеспечении функций различных клеток
Передача сигнала, рецепторы
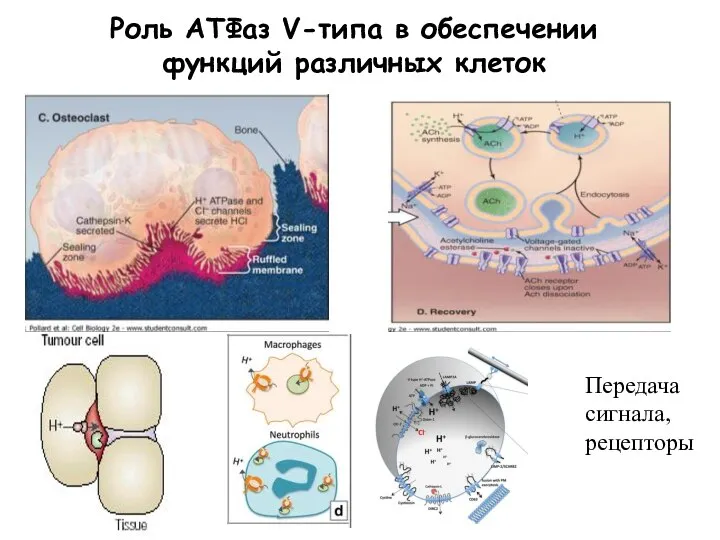
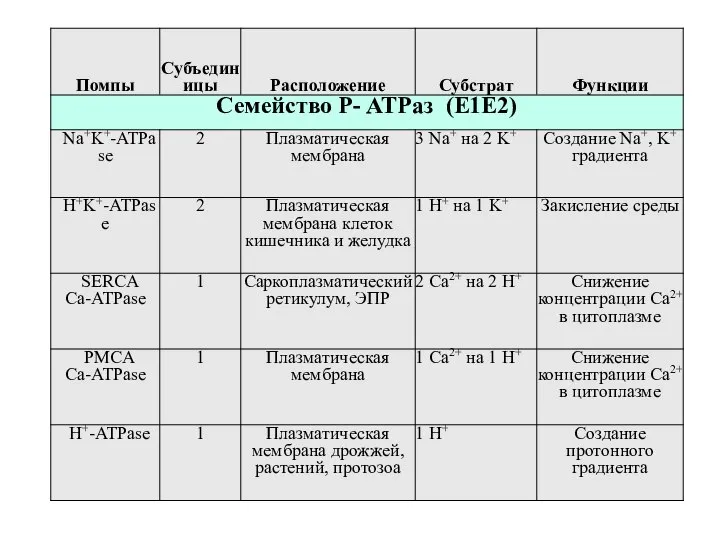
Слайд 20Р-АТФ-азы
Β-аспартил-фосфат
SERCA1
Р-АТФ-азы
Β-аспартил-фосфат
SERCA1
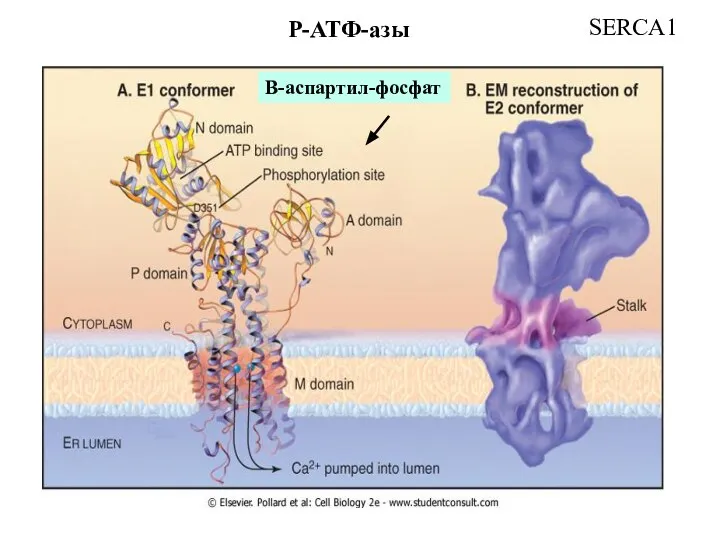
Слайд 21Модель активации мышечной Ca2+ ATPase, локализованной на мембране саркоплазматического ретикулума
P-класс
SERCA1
Модель активации мышечной Ca2+ ATPase, локализованной на мембране саркоплазматического ретикулума
P-класс
SERCA1
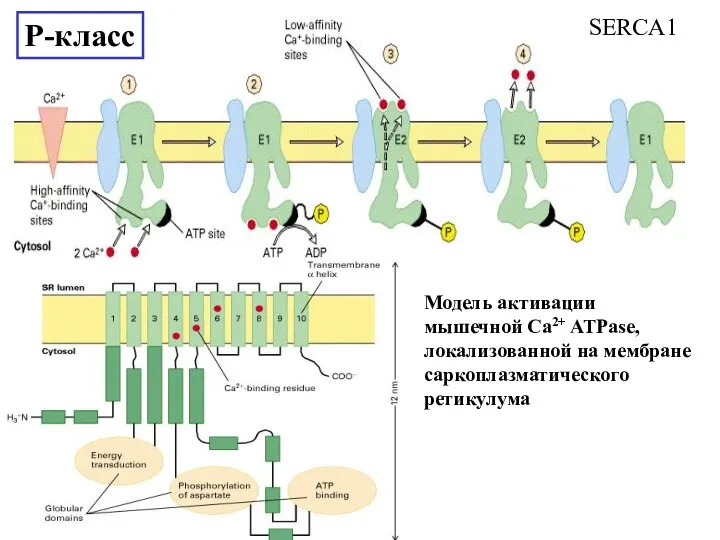
Слайд 22P-класс
12mM
145mM
4mM
139mM
P-класс
12mM
145mM
4mM
139mM
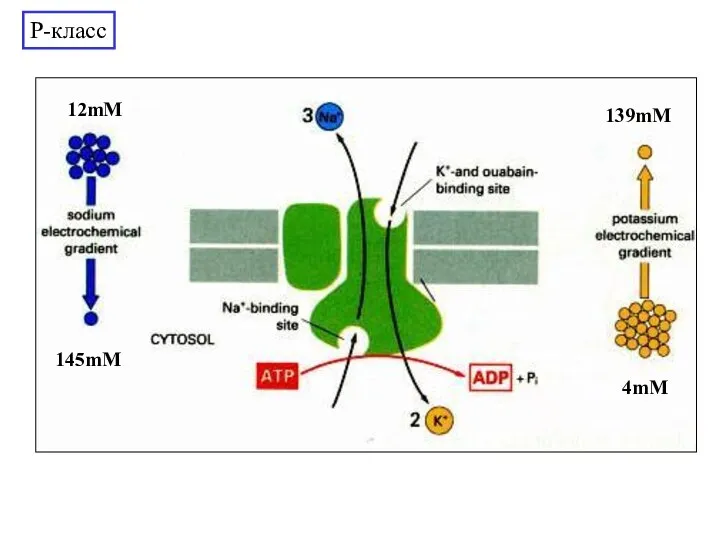
Слайд 23Структура Na,K-АТФазы
Нуклеотид-связывающий
домен
Активаторный домен
5
4
6
8
19 кДа фрагмент
Фосфорилируемый
домен
1
2
3
7
9
10
Участки гликозилирования
P-класс
Структура Na,K-АТФазы
Нуклеотид-связывающий
домен
Активаторный домен
5
4
6
8
19 кДа фрагмент
Фосфорилируемый
домен
1
2
3
7
9
10
Участки гликозилирования
P-класс
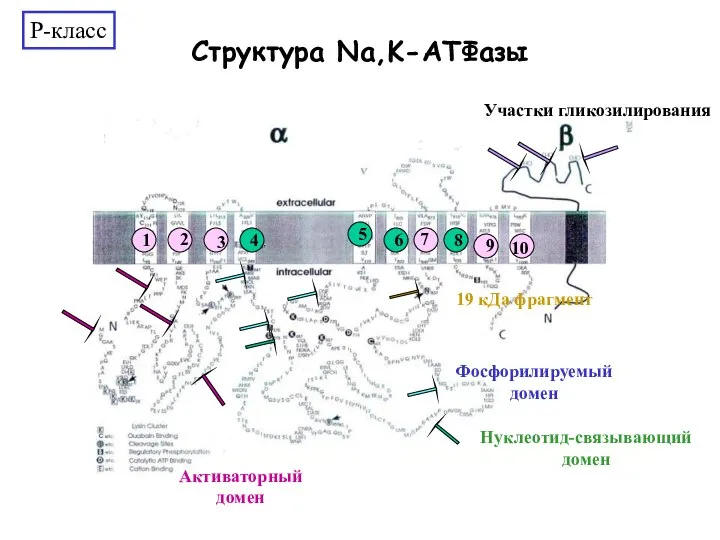
Слайд 24Модель рабоыты Na+/K+ ATPase на плазматической мембране.
Модель рабоыты Na+/K+ ATPase на плазматической мембране.
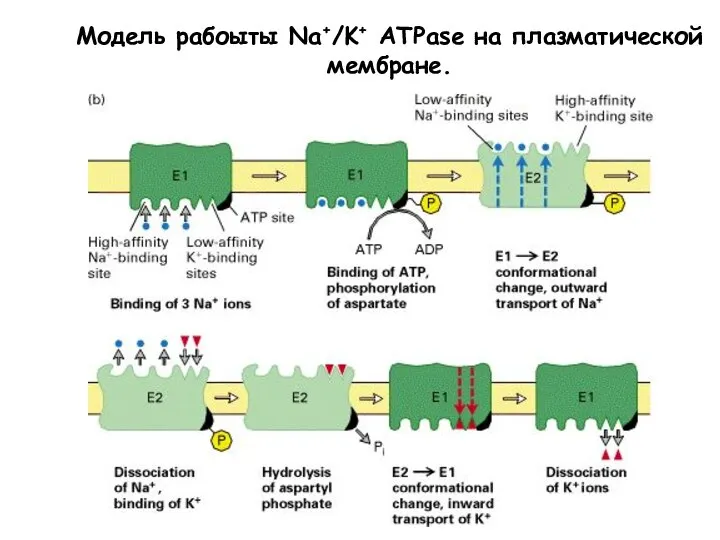
Слайд 25Na,K –ATФаза ингибируется уабаином
В 1785 году Визеринг начал использовать листья наперстянки для
Na,K –ATФаза ингибируется уабаином
В 1785 году Визеринг начал использовать листья наперстянки для
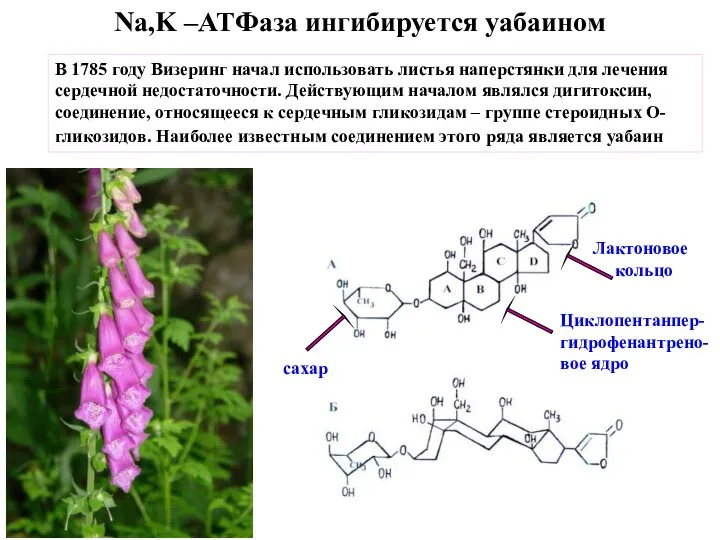
Слайд 26P4-ATPase, флиппаза
P4-ATPase, флиппаза
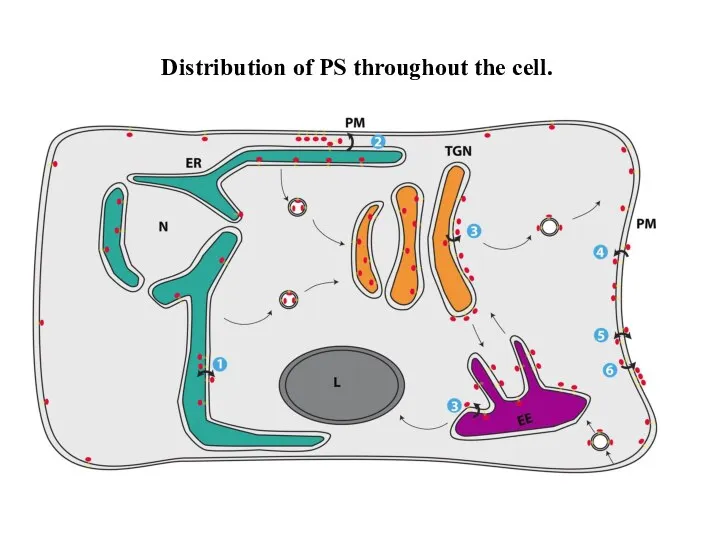
Слайд 27Distribution of PS throughout the cell.
Distribution of PS throughout the cell.
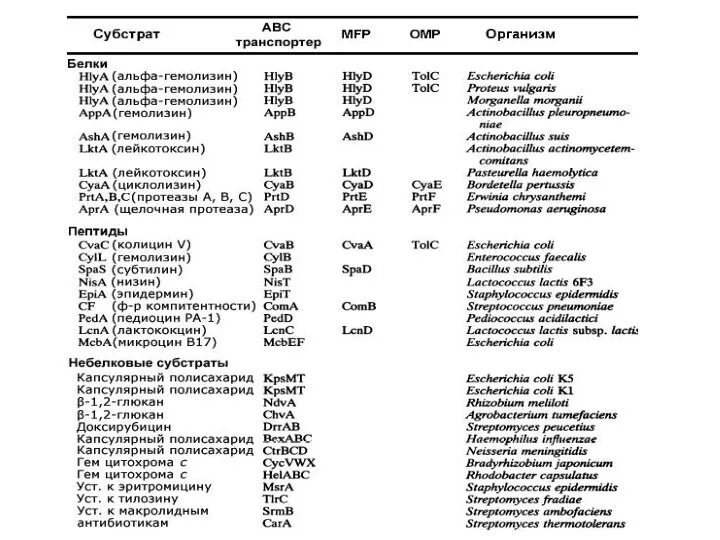
Слайд 28ABC транспортеры
(ATP-Binding “Cassette”)
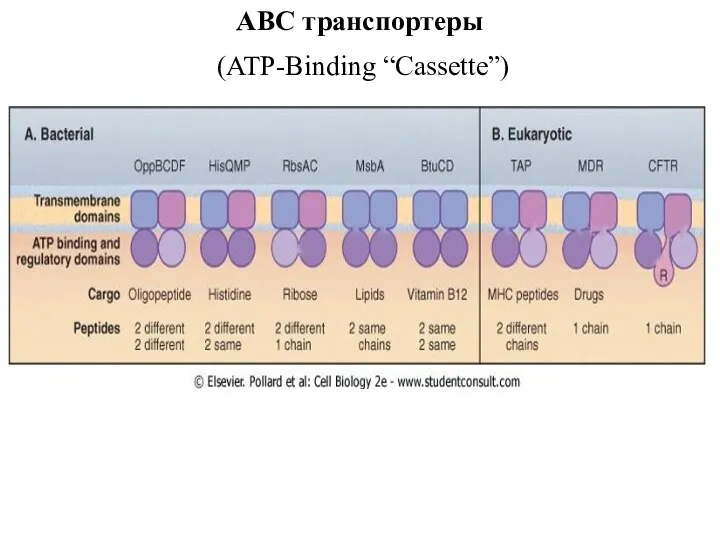
ABC транспортеры
(ATP-Binding “Cassette”)
Слайд 29ABC транспортеры
(ATP-Binding “Cassette”)
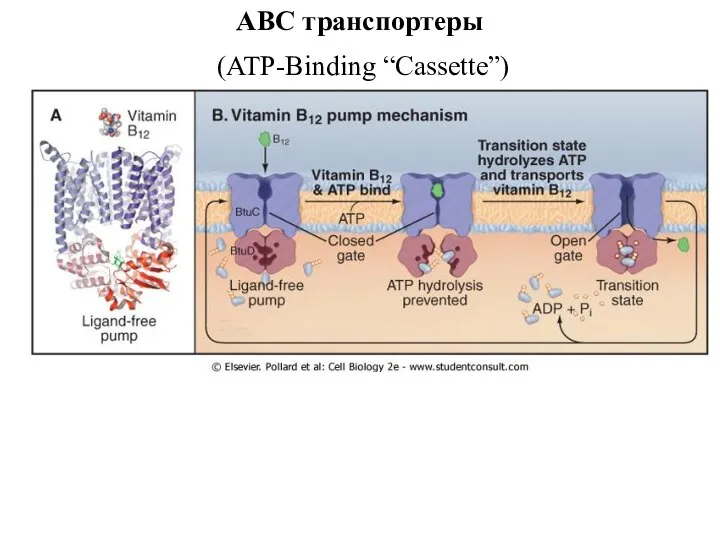
ABC транспортеры
(ATP-Binding “Cassette”)
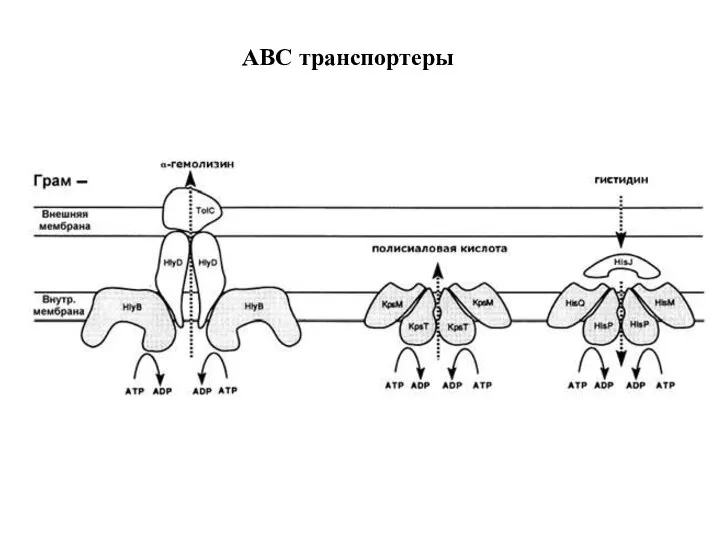
Слайд 30Захват гистидина грамм-отрицательными бактериями
permease
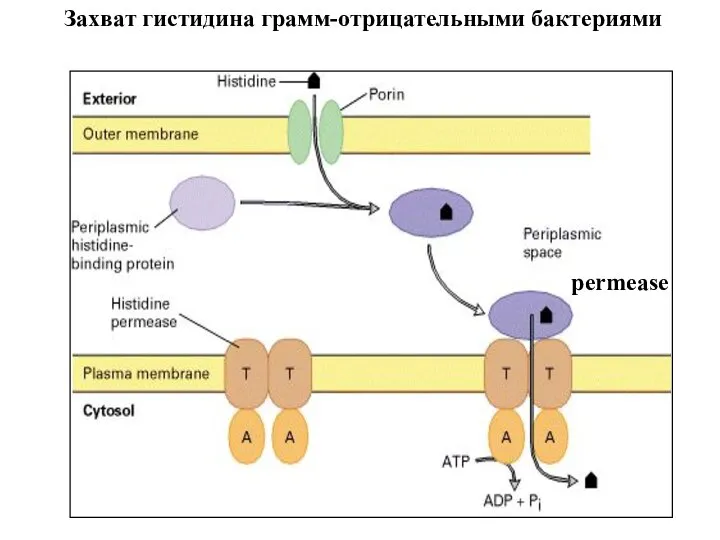
Захват гистидина грамм-отрицательными бактериями
permease
Слайд 31ABC транспортеры
ABC транспортеры
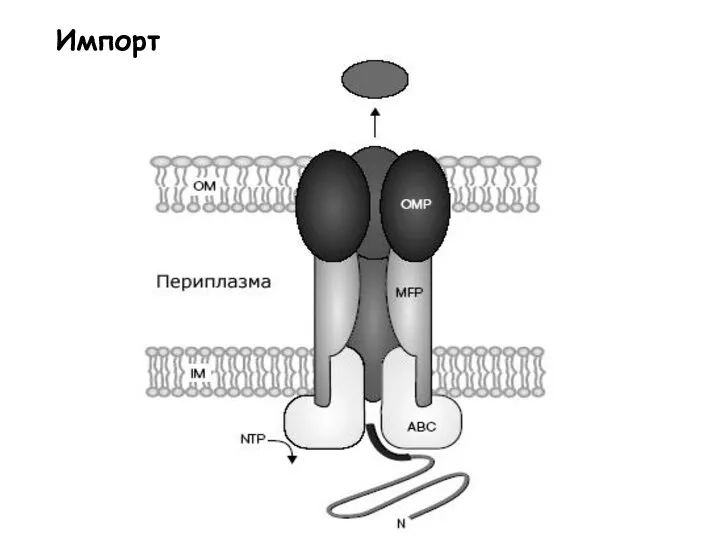
Слайд 33Импорт
Импорт
Слайд 34α-гемолизинE.coli
протеазы E. chrysanthemi
колицинVE.coli
субтилин B.subtilis
капсулярный полисахарид E.coli
по M. Fathеt al., 1993
Оперон
α-гемолизинE.coli
протеазы E. chrysanthemi
колицинVE.coli
субтилин B.subtilis
капсулярный полисахарид E.coli
по M. Fathеt al., 1993
Оперон
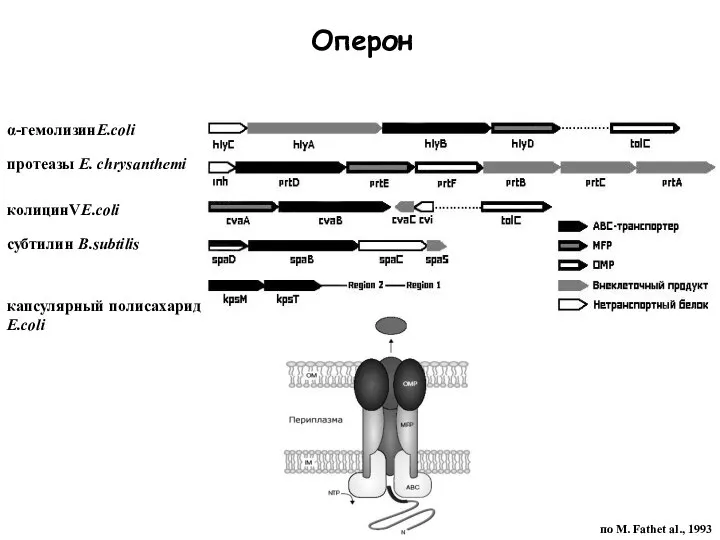
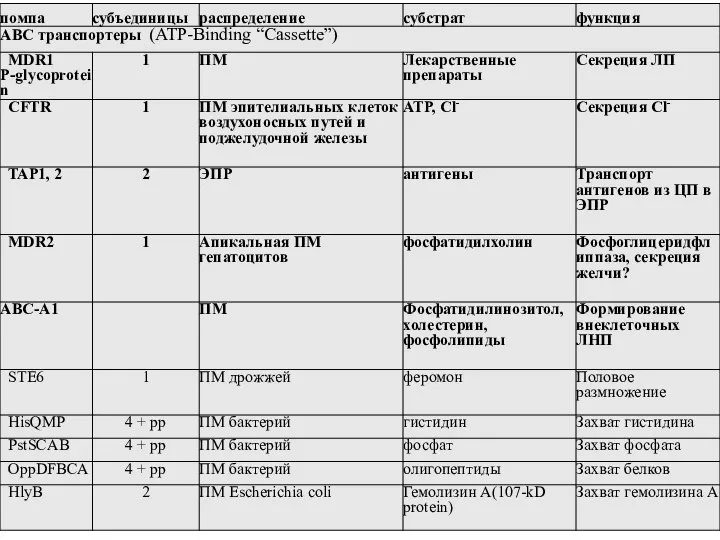
Слайд 36ABC транспортеры эукариот
ABC транспортеры эукариот
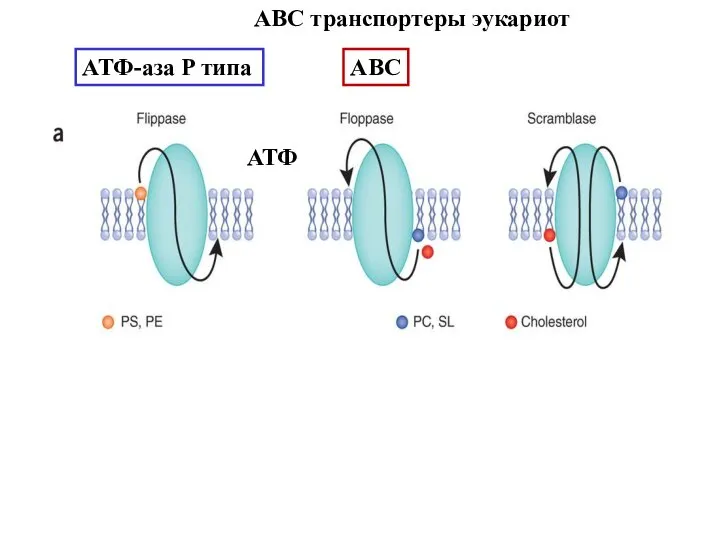
Слайд 37АВС-А1
и транспорт холестерола
АВС-А1
и транспорт холестерола
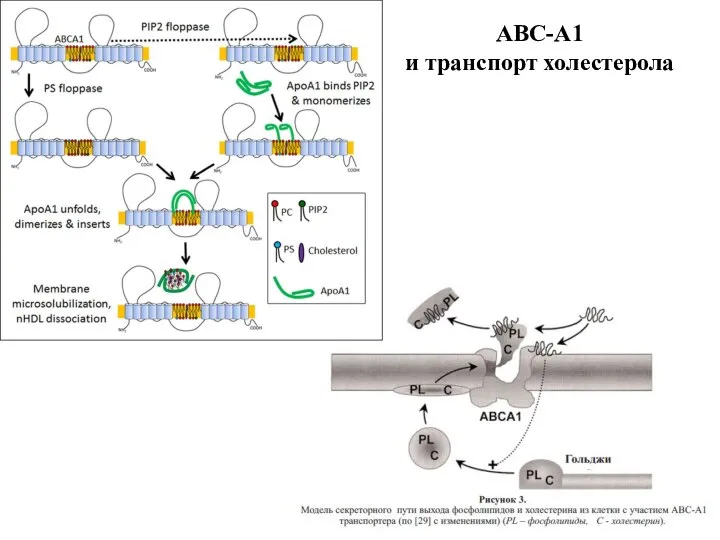
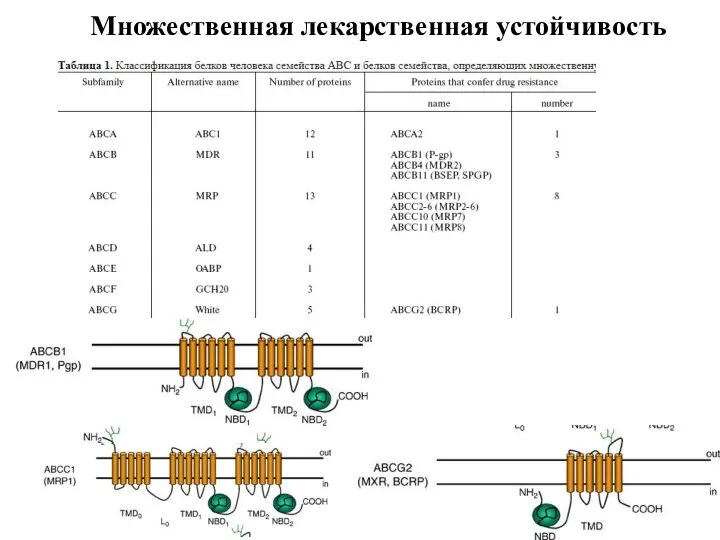
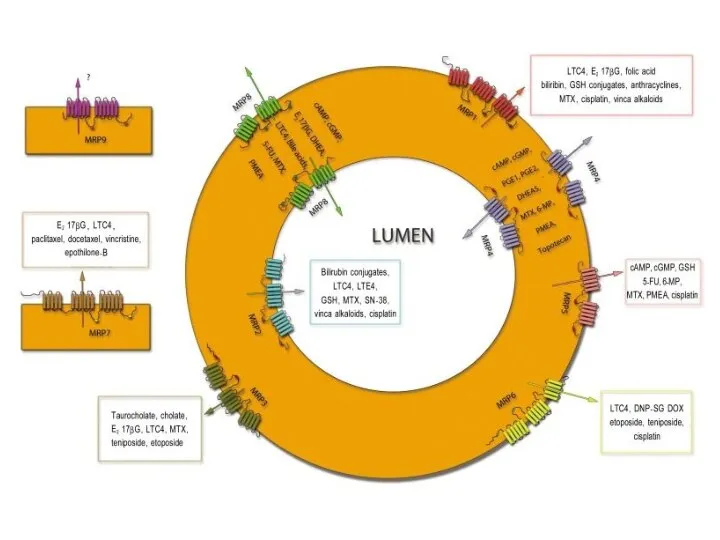
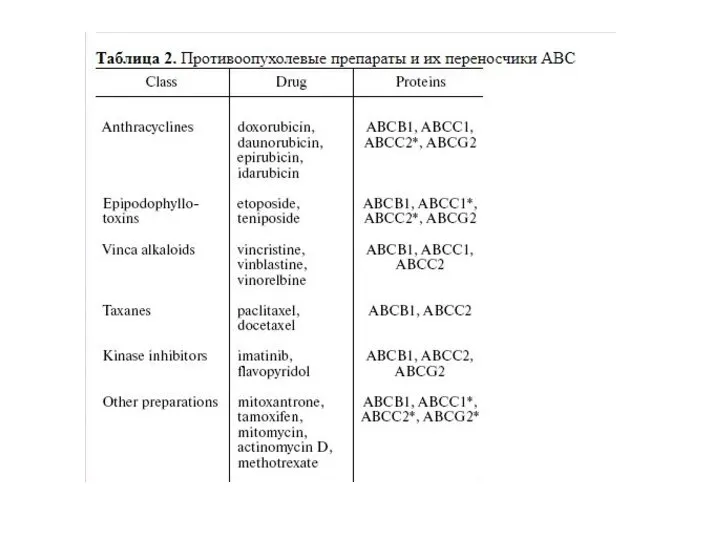
Слайд 38Множественная лекарственная устойчивость
Множественная лекарственная устойчивость
Слайд 39Множественная лекарственная устойчивость
Множественная лекарственная устойчивость
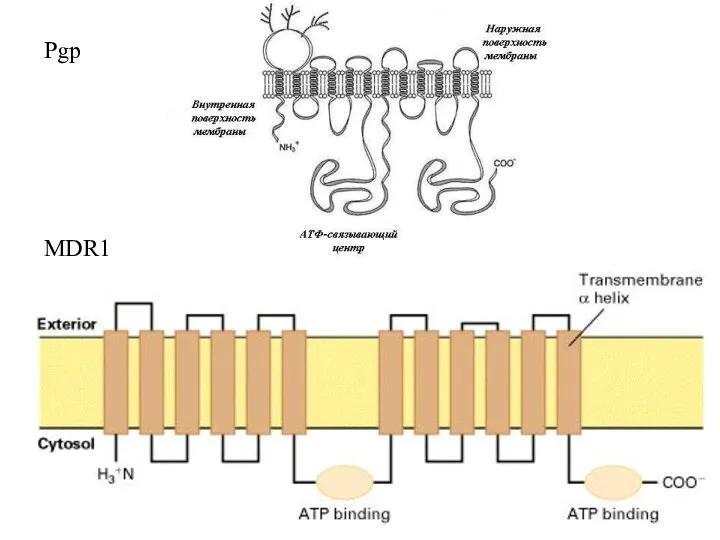
Слайд 42MDR1
Pgp
MDR1
Pgp
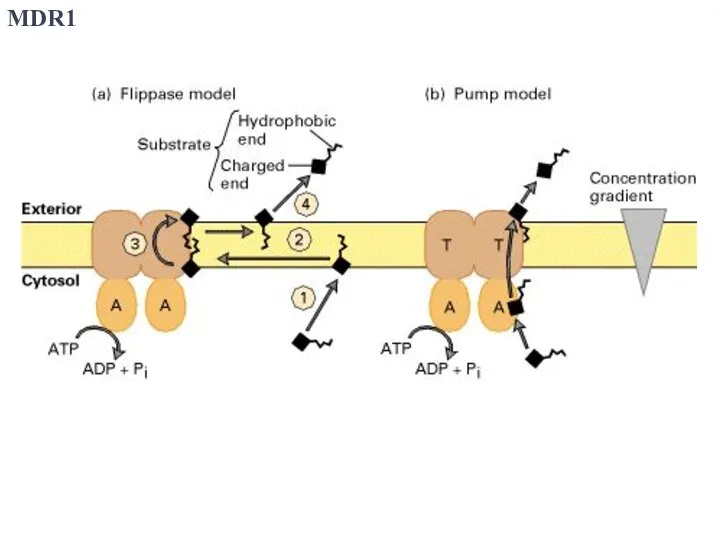
Слайд 43MDR1
MDR1
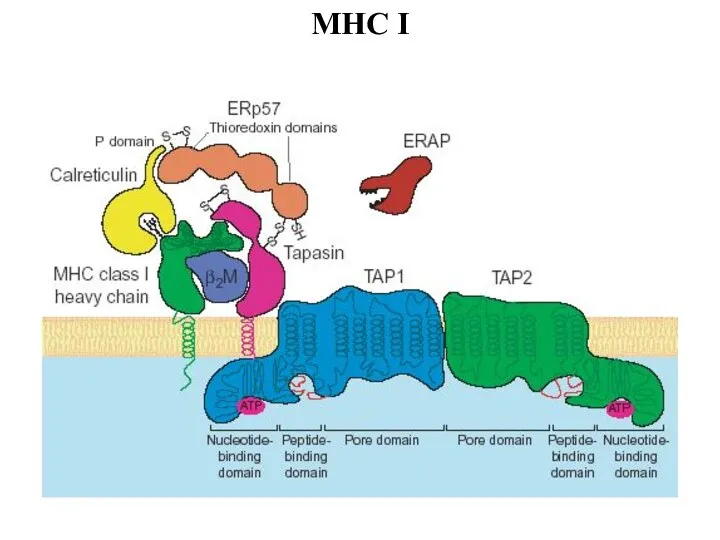
Слайд 44МНС I
МНС I
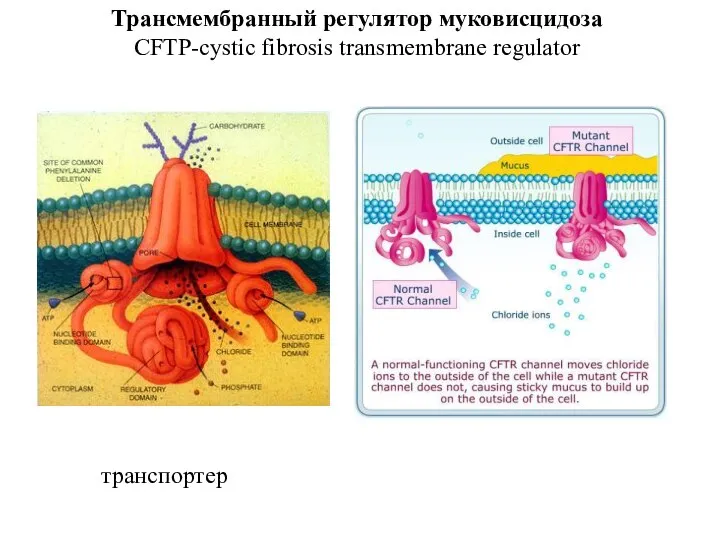
Слайд 45Трансмембранный регулятор муковисцидоза CFTP-cystic fibrosis transmembrane regulator
транспортер
Трансмембранный регулятор муковисцидоза CFTP-cystic fibrosis transmembrane regulator
транспортер
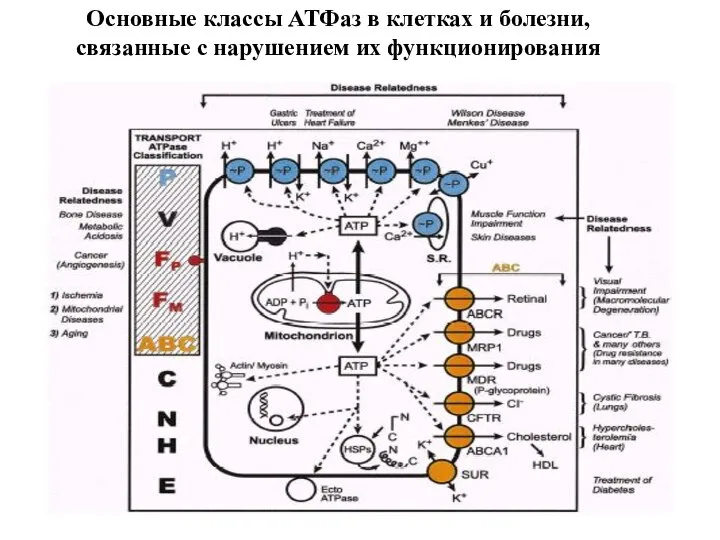
Слайд 46Основные классы АТФаз в клетках и болезни, связанные с нарушением их функционирования
Основные классы АТФаз в клетках и болезни, связанные с нарушением их функционирования
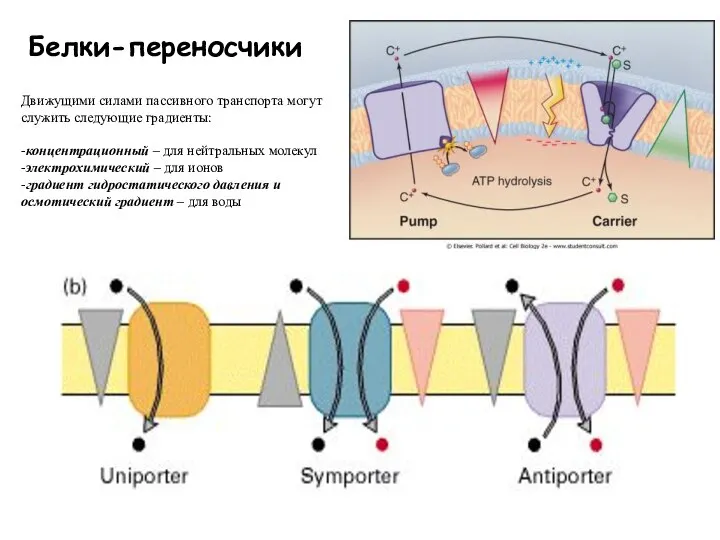
Слайд 47Белки-переносчики
Движущими силами пассивного транспорта могут служить следующие градиенты:
-концентрационный – для нейтральных молекул
-электрохимический
Белки-переносчики
Движущими силами пассивного транспорта могут служить следующие градиенты:
-концентрационный – для нейтральных молекул
-электрохимический
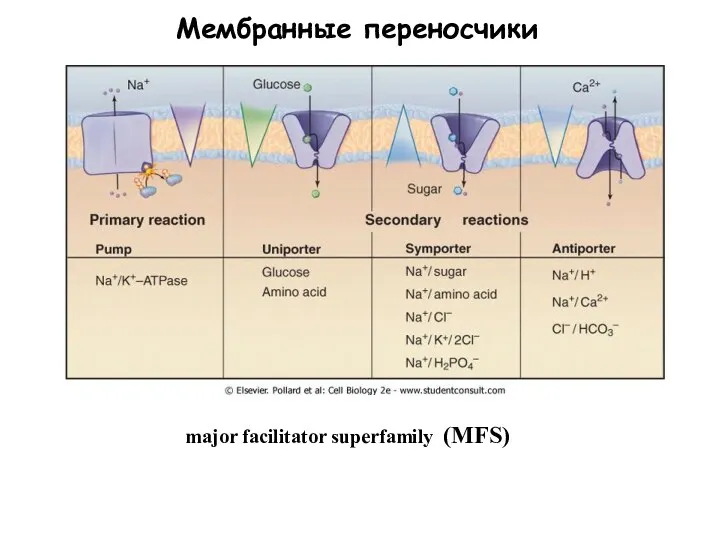
Слайд 48Мембранные переносчики
major facilitator superfamily (MFS)
Мембранные переносчики
major facilitator superfamily (MFS)
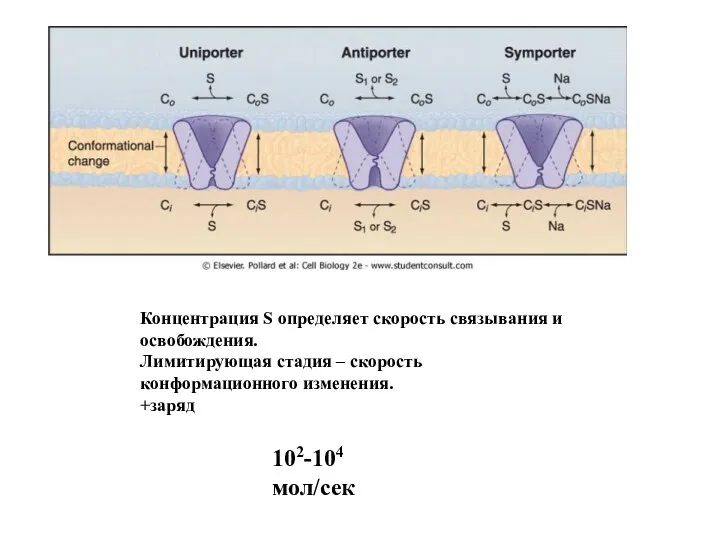
Слайд 49Концентрация S определяет скорость связывания и освобождения.
Лимитирующая стадия – скорость конформационного изменения.
+заряд
102-104
Концентрация S определяет скорость связывания и освобождения.
Лимитирующая стадия – скорость конформационного изменения.
+заряд
102-104
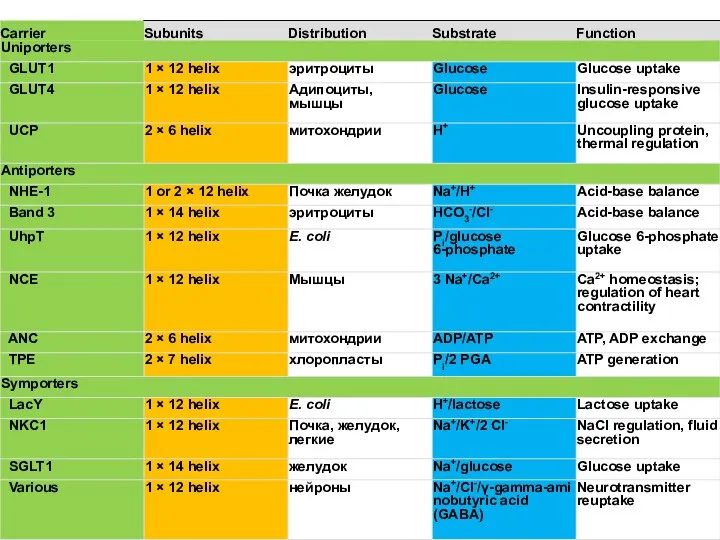
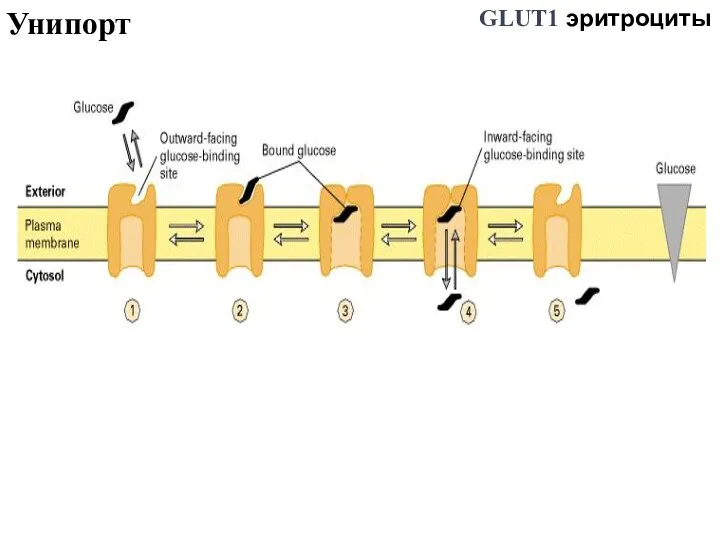
Слайд 51Унипорт
GLUT1 эритроциты
Унипорт
GLUT1 эритроциты
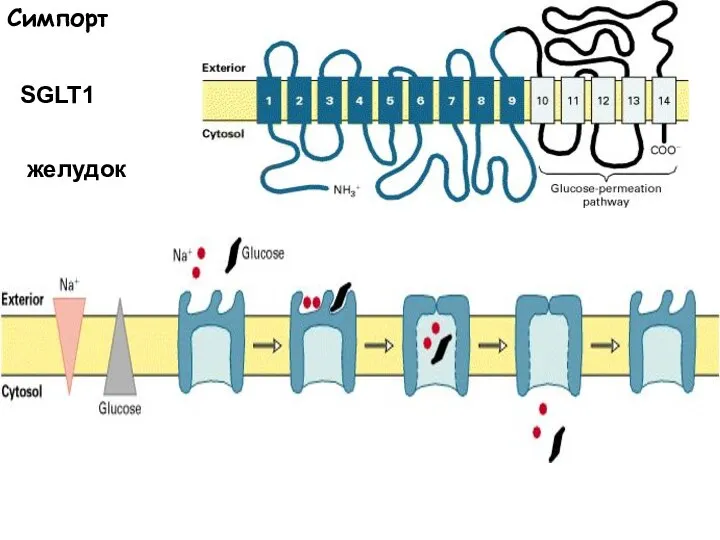
Слайд 52Симпорт
SGLT1
желудок
Симпорт
SGLT1
желудок
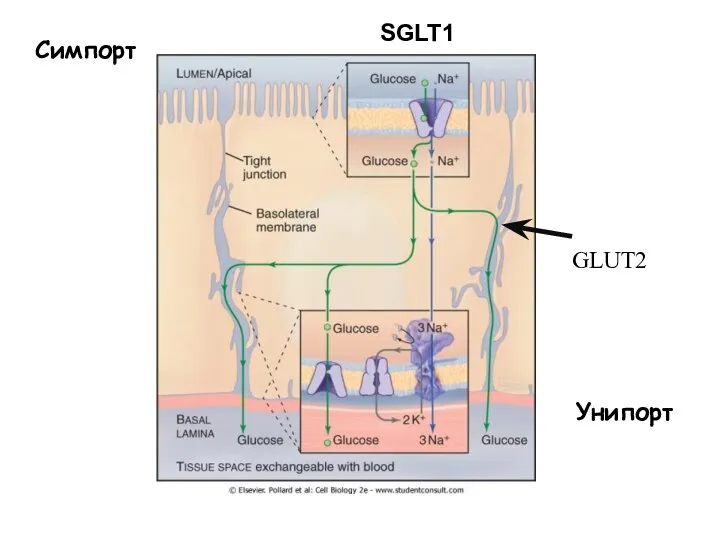
Слайд 53Симпорт
GLUT2
Унипорт
SGLT1
Симпорт
GLUT2
Унипорт
SGLT1
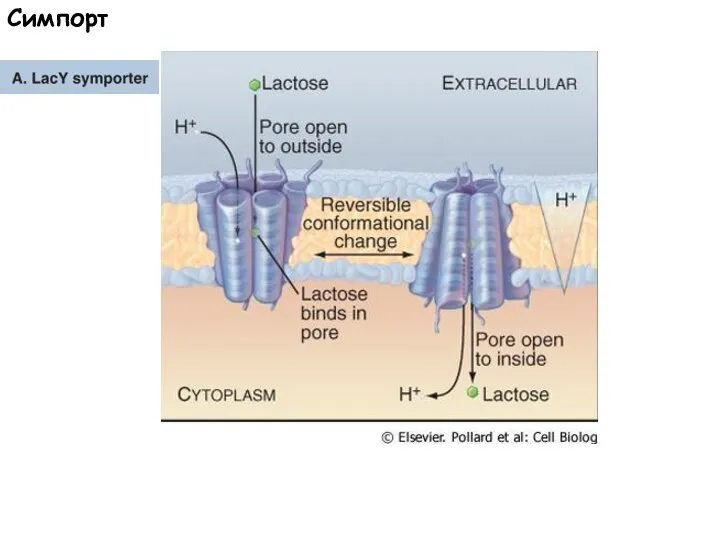
Слайд 54Симпорт
Симпорт
Слайд 55Примеры транспортеров симпортеров
Примеры транспортеров симпортеров
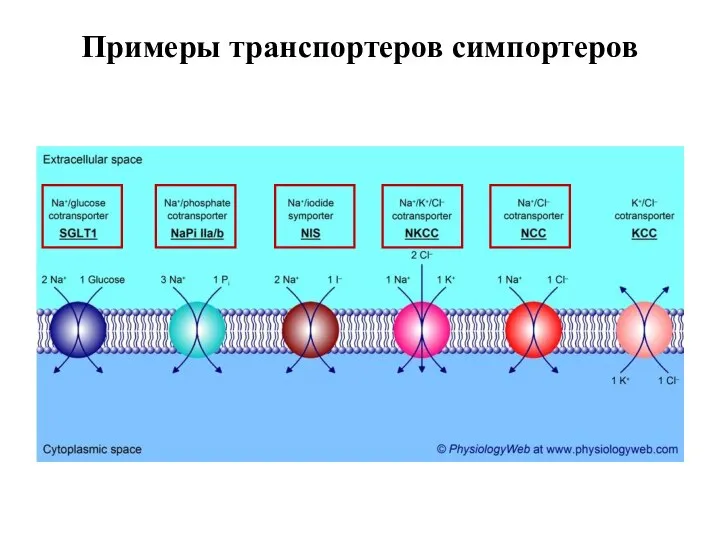
Слайд 56Примеры транспортеров симпортеров
Примеры транспортеров симпортеров
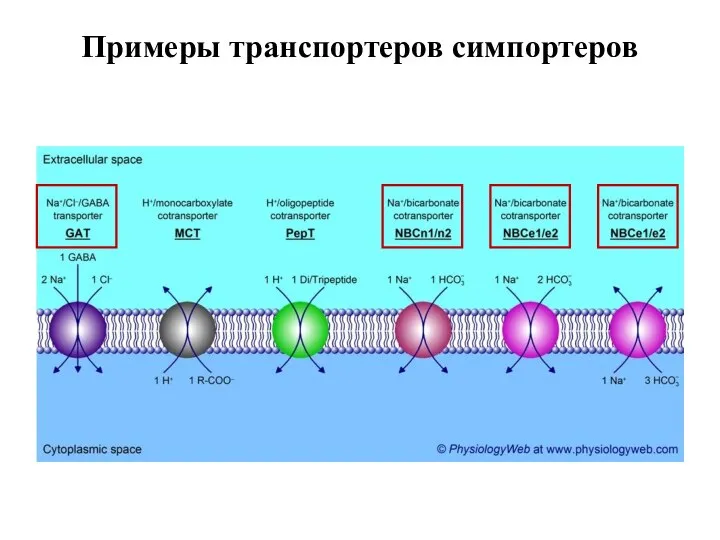
Слайд 57Антипорт
Антипорт
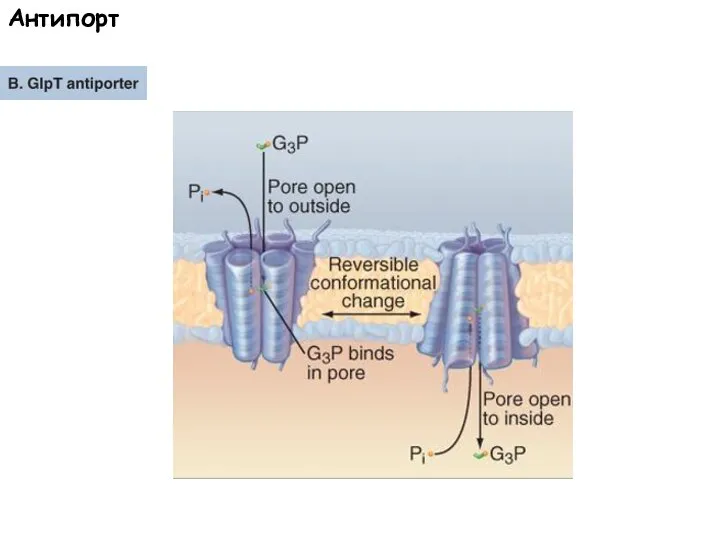
Слайд 58ATP/ADP Транслоказа
Антипорт
ATP/ADP Транслоказа
Антипорт
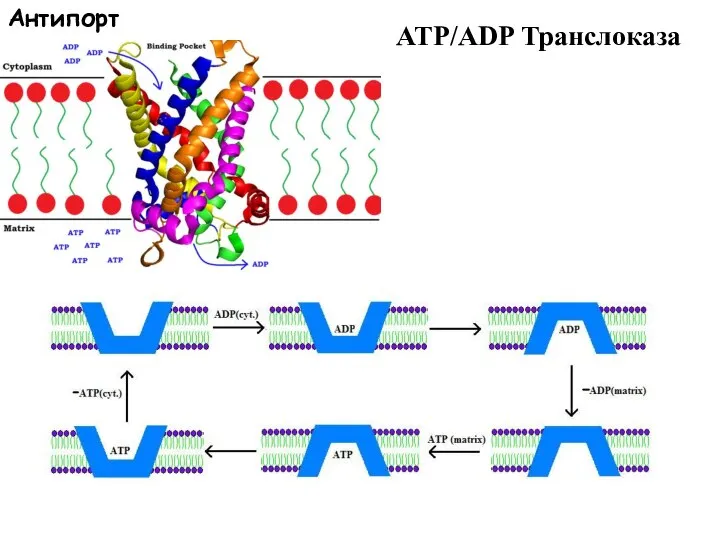
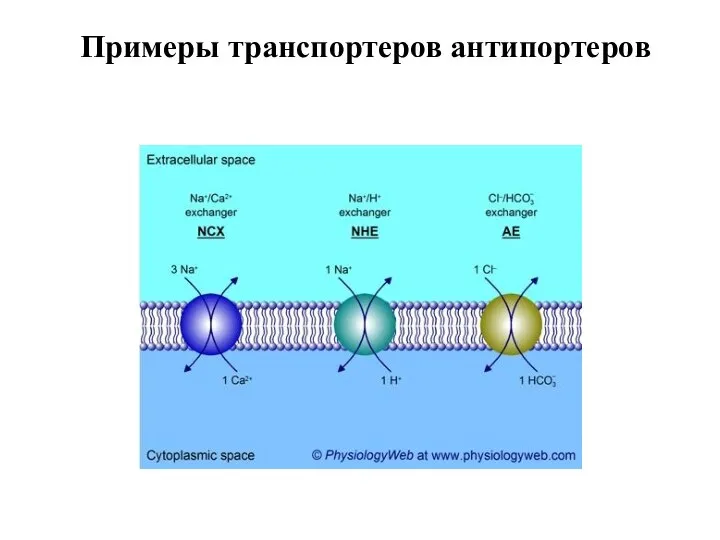
Слайд 59Примеры транспортеров антипортеров
Примеры транспортеров антипортеров
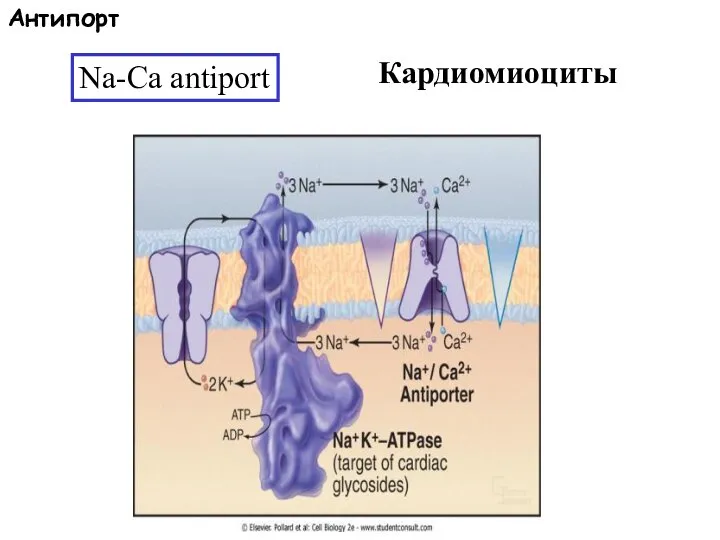
Слайд 60Na-Ca antiport
Кардиомиоциты
Антипорт
Na-Ca antiport
Кардиомиоциты
Антипорт
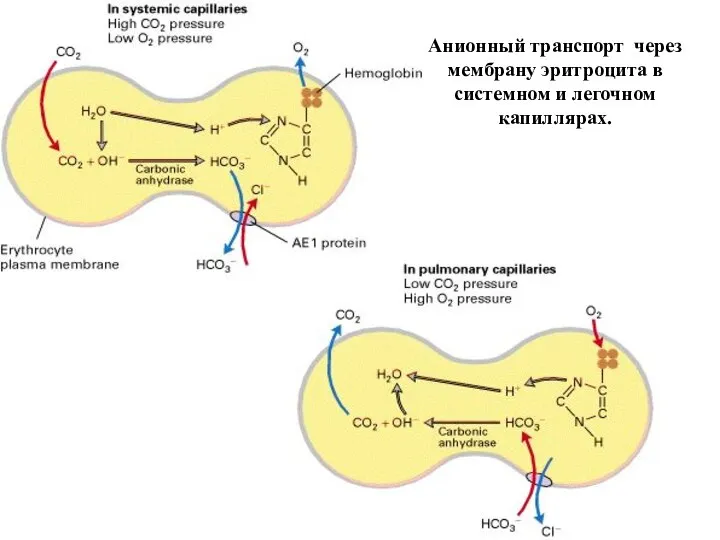
Слайд 61Анионный транспорт через мембрану эритроцита в системном и легочном капиллярах.
Анионный транспорт через мембрану эритроцита в системном и легочном капиллярах.
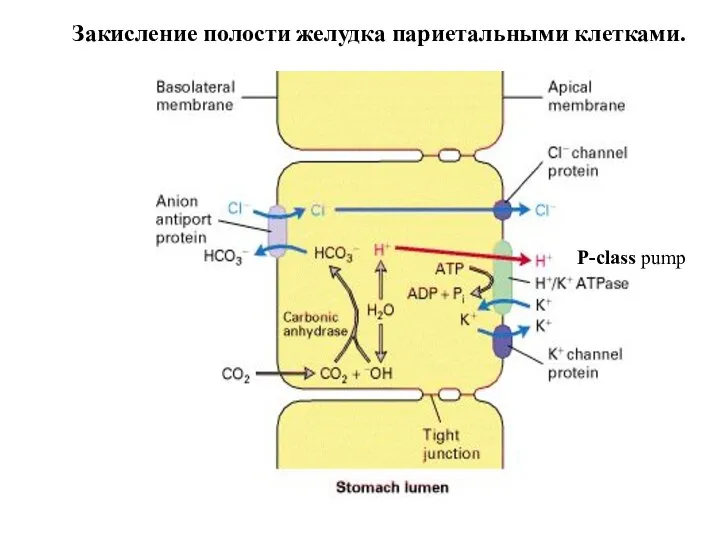
Слайд 62
Закисление полости желудка париетальными клетками.
P-class pump
Закисление полости желудка париетальными клетками.
P-class pump
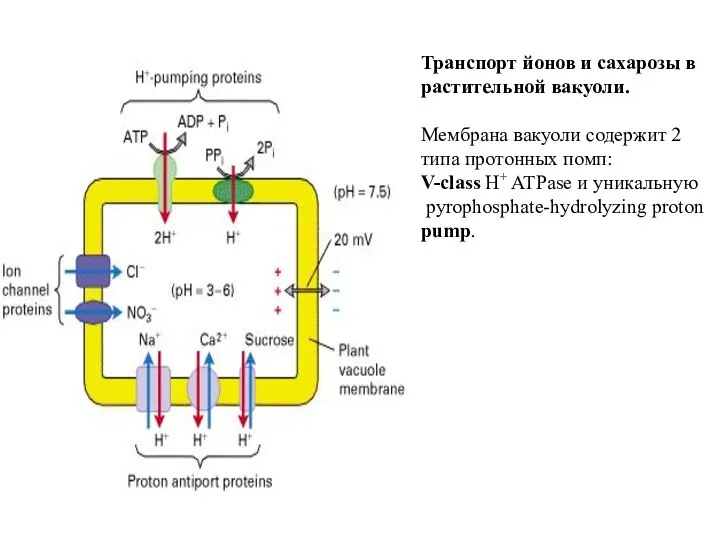
Слайд 63Транспорт йонов и сахарозы в растительной вакуоли.
Мембрана вакуоли содержит 2 типа
Транспорт йонов и сахарозы в растительной вакуоли.
Мембрана вакуоли содержит 2 типа
Слайд 64Ингибиторы белков-переносчиков
Ингибиторы белков-переносчиков

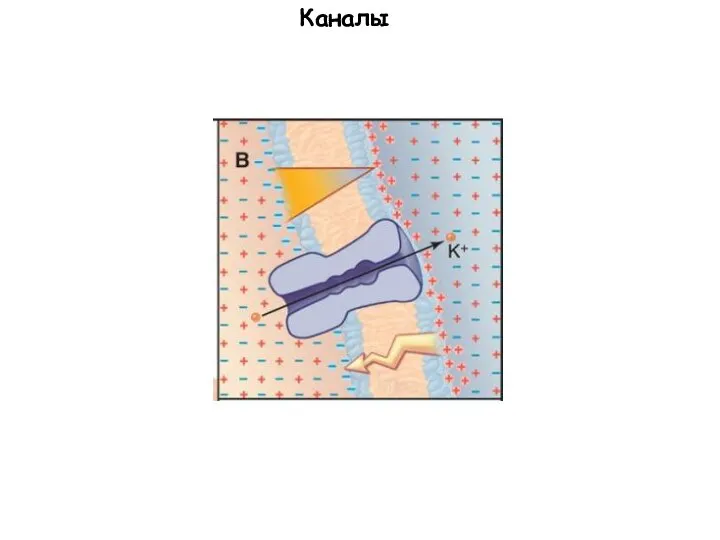
Слайд 65Каналы
Каналы
Слайд 66Функции йонных каналов:
Регуляция водного обмена клетки: объём и тургор.
2. Регуляция pH: закисление
Функции йонных каналов:
Регуляция водного обмена клетки: объём и тургор.
2. Регуляция pH: закисление

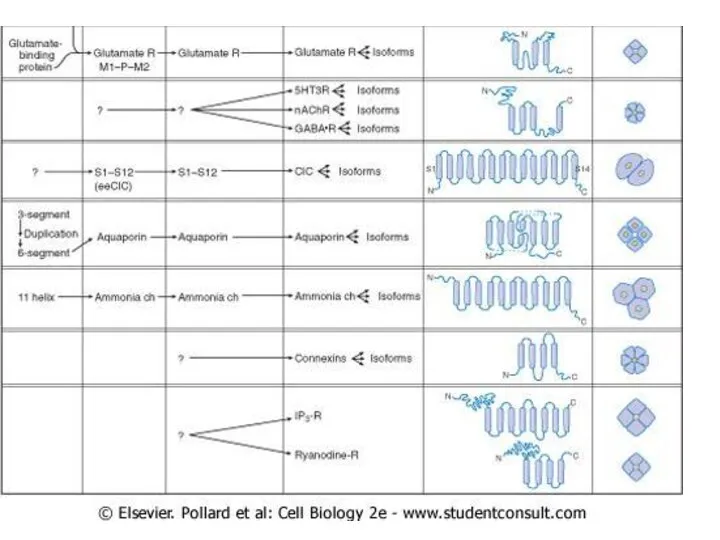
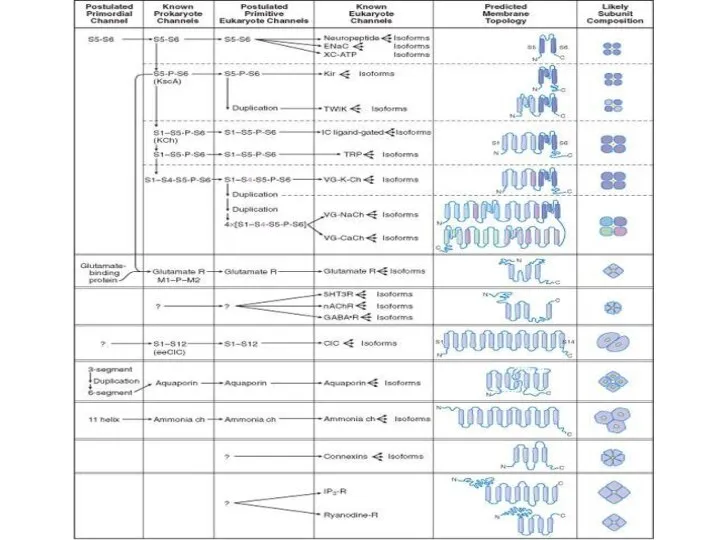
Слайд 67По строению (родству их химического строения и происхождения образующих их белков). По
По строению (родству их химического строения и происхождения образующих их белков). По

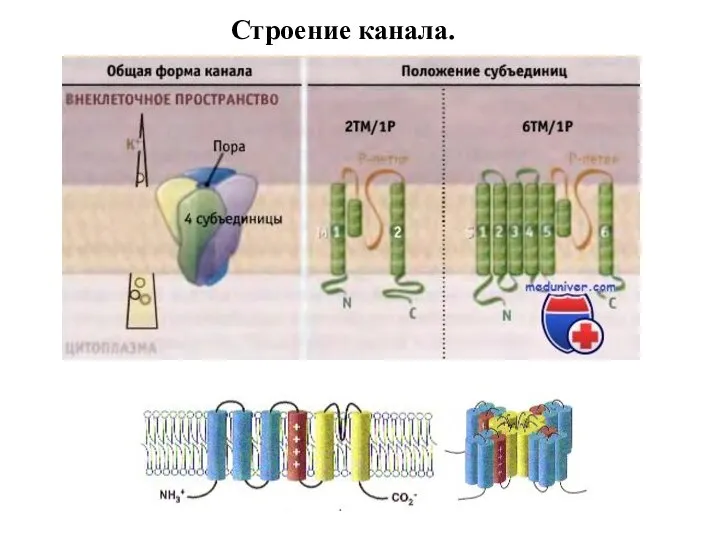
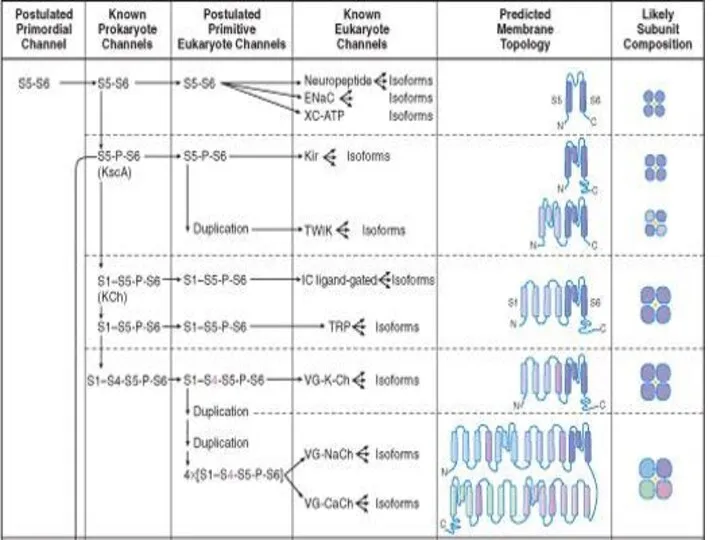
Слайд 68Строение канала.
Строение канала.
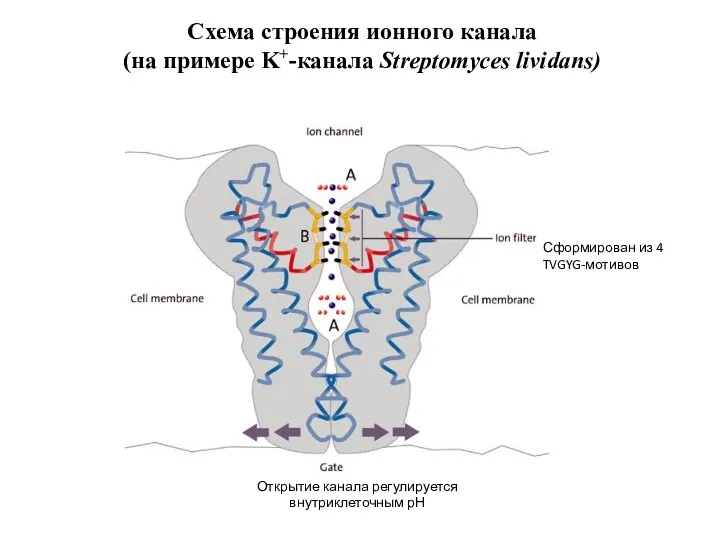
Слайд 72Схема строения ионного канала
(на примере K+-канала Streptomyces lividans)
Сформирован из 4 TVGYG-мотивов
Открытие канала
Схема строения ионного канала
(на примере K+-канала Streptomyces lividans)
Сформирован из 4 TVGYG-мотивов
Открытие канала
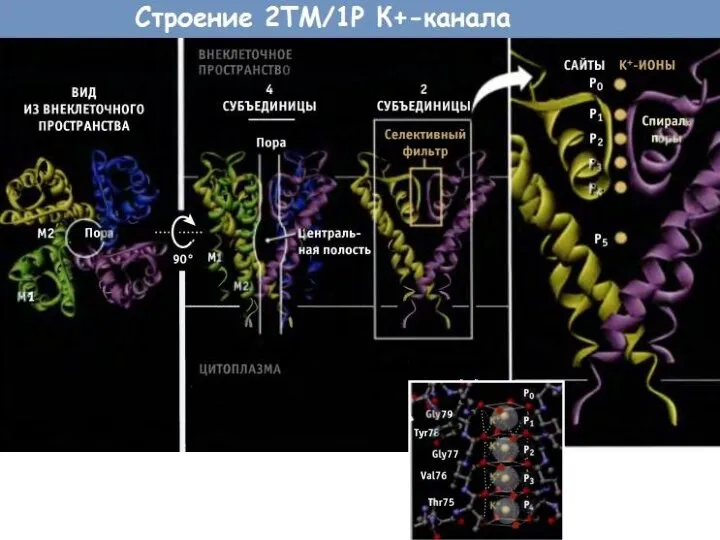
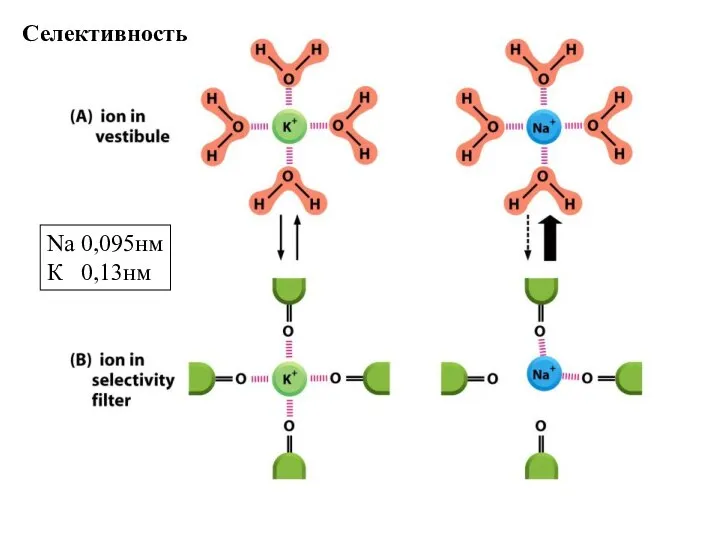
Слайд 74Селективность
Na 0,095нм
К 0,13нм
Селективность
Na 0,095нм
К 0,13нм
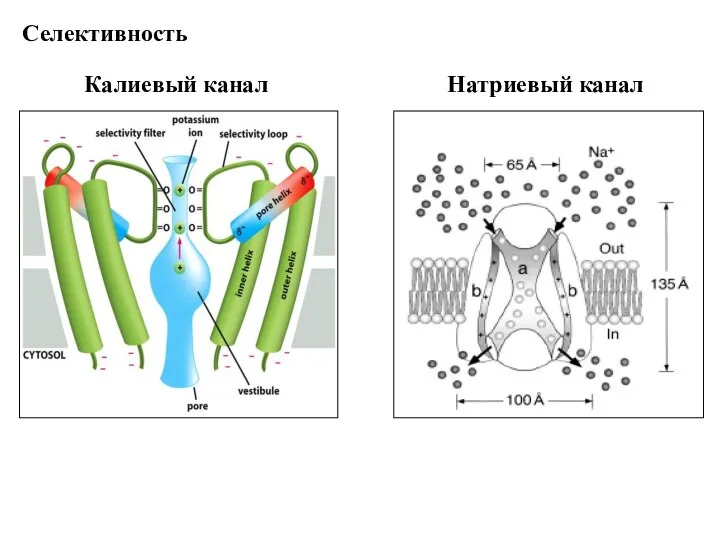
Слайд 75Калиевый канал
Селективность
Натриевый канал
Калиевый канал
Селективность
Натриевый канал
Слайд 78Каналы покоя
Воротные каналы
Управляемость
Неуправляемые, потенциал-управляемые, хемоуправляемые, стимул-управляемые, опосредованно-управляемые, актин-управляемые
Каналы покоя
Воротные каналы
Управляемость
Неуправляемые, потенциал-управляемые, хемоуправляемые, стимул-управляемые, опосредованно-управляемые, актин-управляемые
Слайд 79Са-активируемый К канал
Управляемость
Са-активируемый К канал
Управляемость
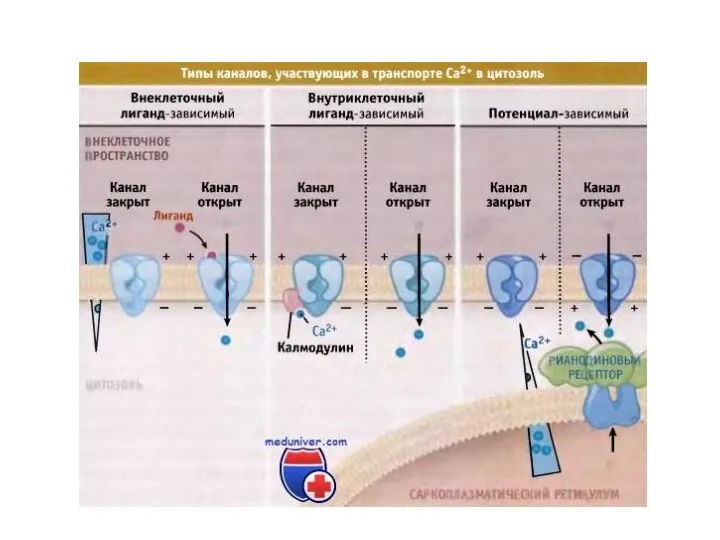
Слайд 81Гомоцистеин
Глутамат
N-метил-D-аспартат
NMDA-рецептор – пример лиганд-управляемого ионного канала
(Carlson, 1998)
NR1
NR2
Гомоцистеин
Глутамат
N-метил-D-аспартат
NMDA-рецептор – пример лиганд-управляемого ионного канала
(Carlson, 1998)
NR1
NR2
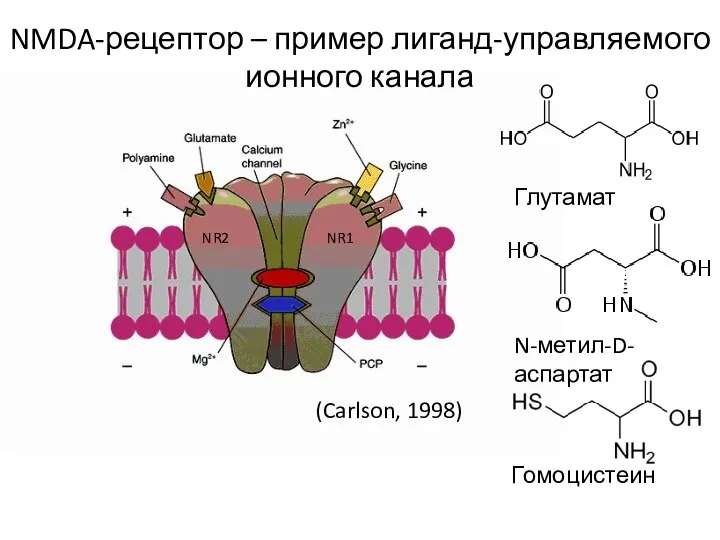
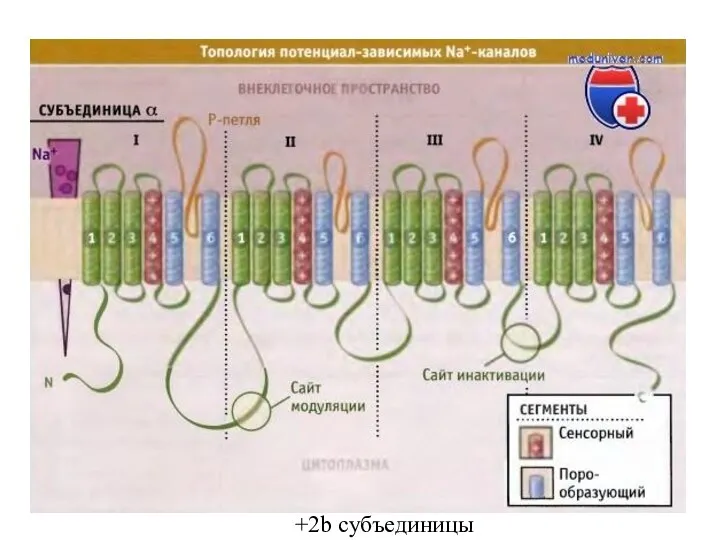
Слайд 82+2b субъединицы
+2b субъединицы
Слайд 83Потенциал-управляемый К канал
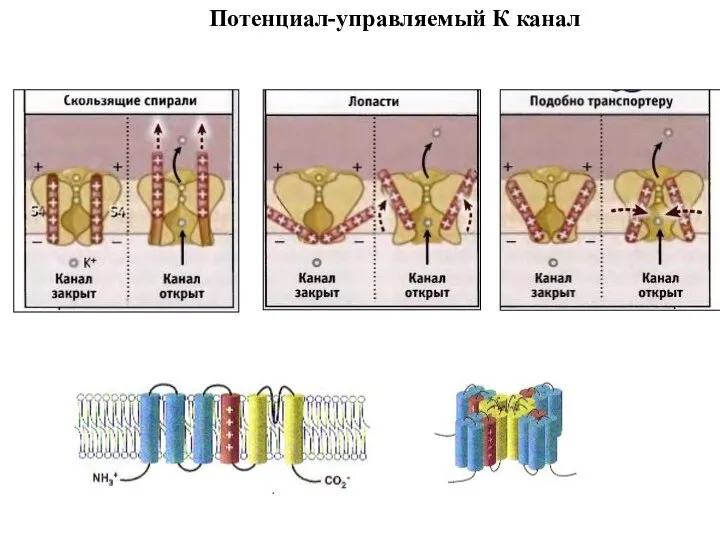
Потенциал-управляемый К канал
Слайд 841. Открытое. Канал открыт и через него происходит перемещение ионов.
2. Закрытое. Канал
1. Открытое. Канал открыт и через него происходит перемещение ионов.
2. Закрытое. Канал

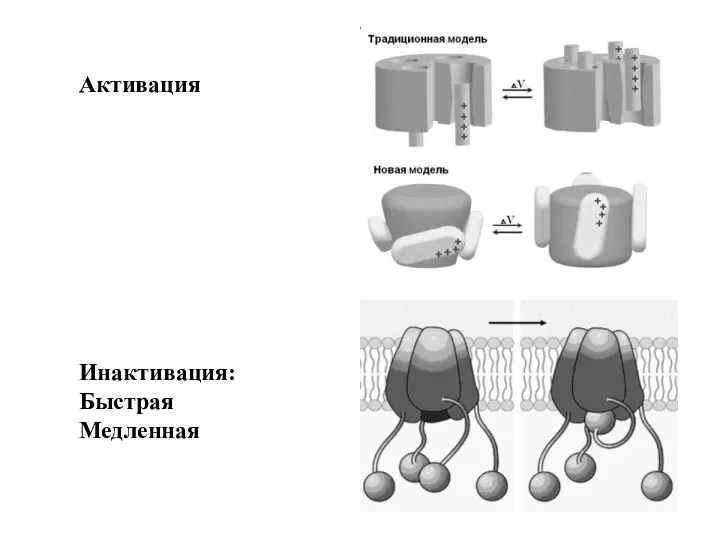
Слайд 85Активация
Инактивация:
Быстрая
Медленная
Активация
Инактивация:
Быстрая
Медленная
Слайд 86Тетродотоксин: токсин рыбы фугу.
Блокатор Na-каналов
Тетродотоксин: токсин рыбы фугу.
Блокатор Na-каналов

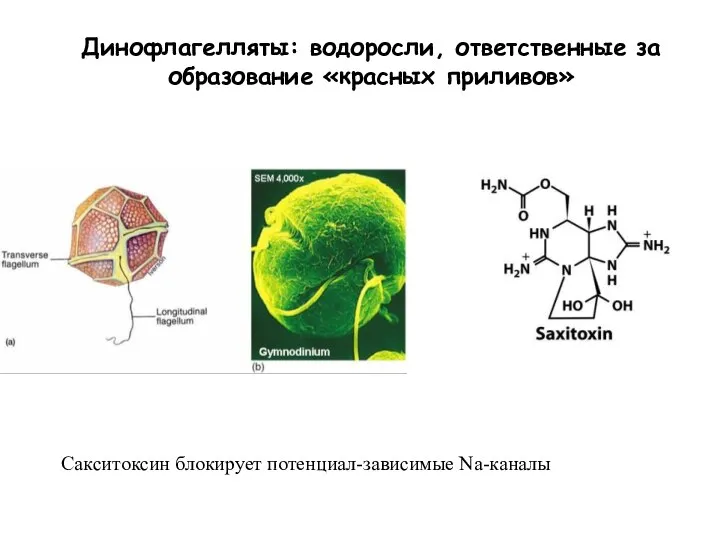
Слайд 87Динофлагелляты: водоросли, ответственные за образование «красных приливов»
Сакситоксин блокирует потенциал-зависимые Na-каналы
Динофлагелляты: водоросли, ответственные за образование «красных приливов»
Сакситоксин блокирует потенциал-зависимые Na-каналы
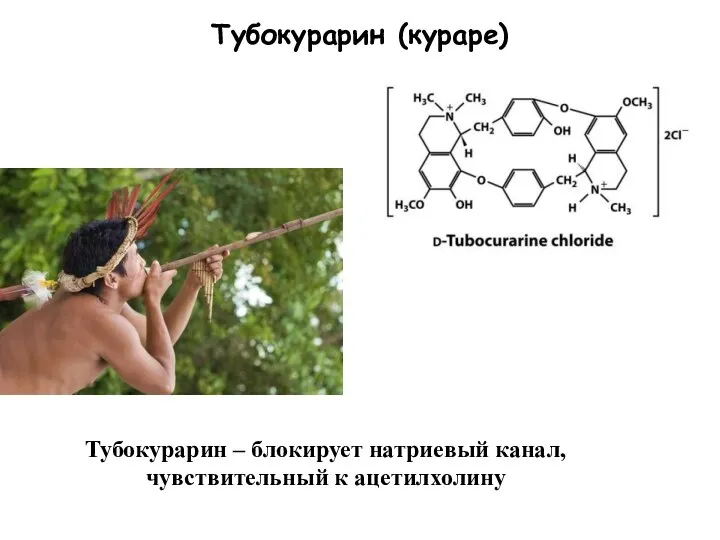
Слайд 88Тубокурарин (кураре)
Тубокурарин – блокирует натриевый канал, чувствительный к ацетилхолину
Тубокурарин (кураре)
Тубокурарин – блокирует натриевый канал, чувствительный к ацетилхолину
Слайд 89Дендротоксин из яда черной мамбы
Дендротоксин – блокатор потенциал-зависимых калиевых каналов
Дендротоксин из яда черной мамбы
Дендротоксин – блокатор потенциал-зависимых калиевых каналов

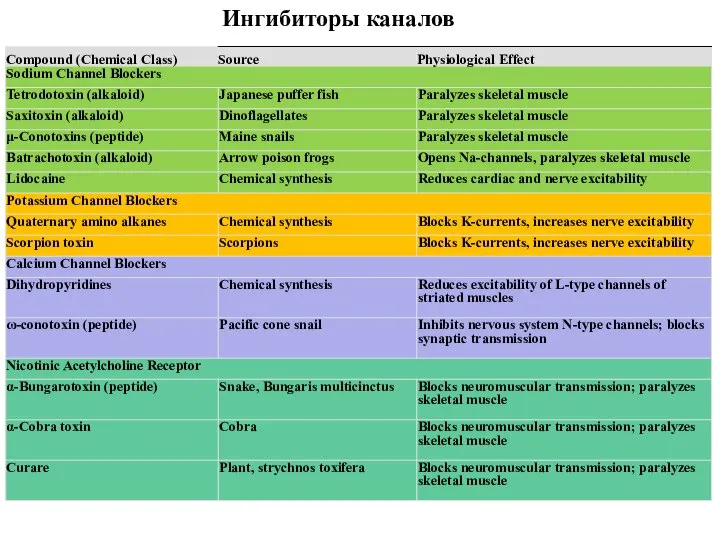
Слайд 90Ингибиторы каналов
Ингибиторы каналов
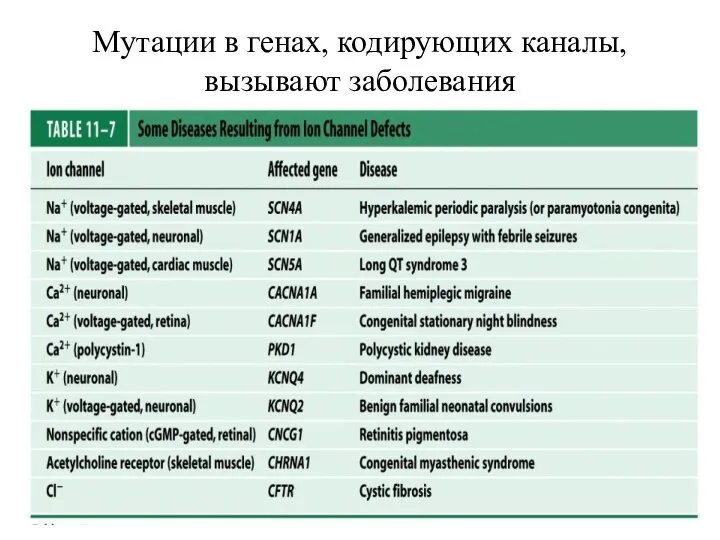
Слайд 91Мутации в генах, кодирующих каналы, вызывают заболевания
Мутации в генах, кодирующих каналы, вызывают заболевания
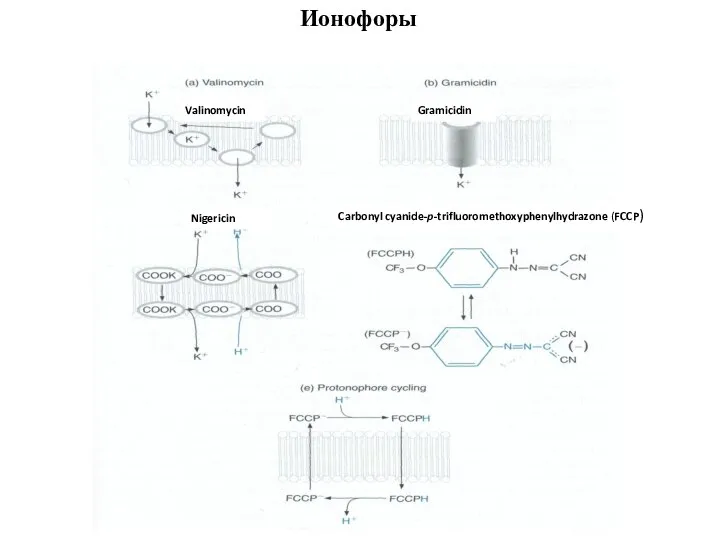
Слайд 92Carbonyl cyanide-p-trifluoromethoxyphenylhydrazone (FCCP)
Gramicidin
Valinomycin
Nigericin
Ионофоры
Carbonyl cyanide-p-trifluoromethoxyphenylhydrazone (FCCP)
Gramicidin
Valinomycin
Nigericin
Ионофоры
Слайд 93Порины
A trimeric OmpF channel in the lipid bilayer of the outer bacterial
Порины
A trimeric OmpF channel in the lipid bilayer of the outer bacterial
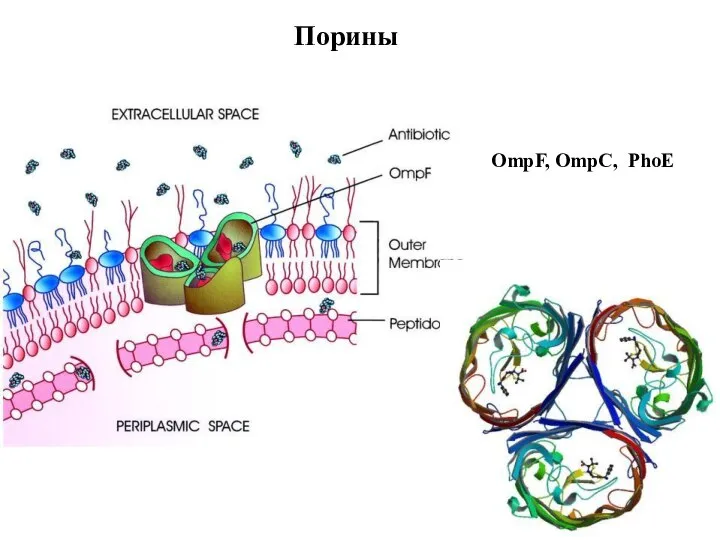
Слайд 94Порины
Порины
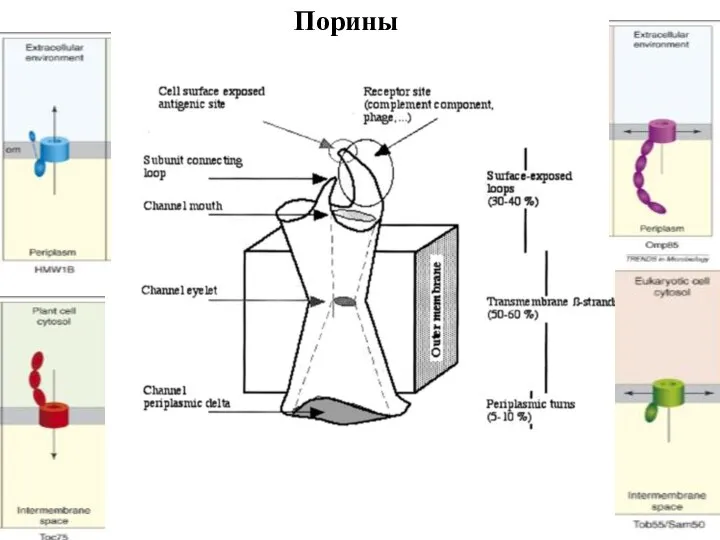
Слайд 95Рorin superfamily I
The mitochondrial and plastid porin (MPP)
Porin Superfamily II
Рorin superfamily I
The mitochondrial and plastid porin (MPP)
Porin Superfamily II

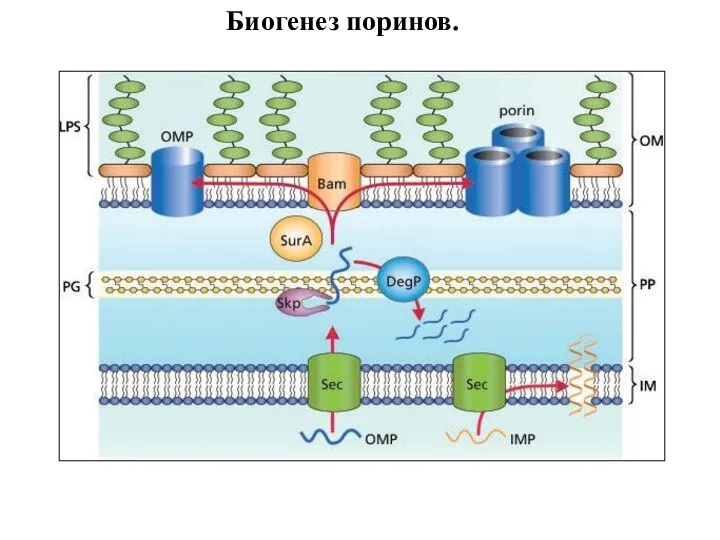
Слайд 96Биогенез поринов.
Биогенез поринов.
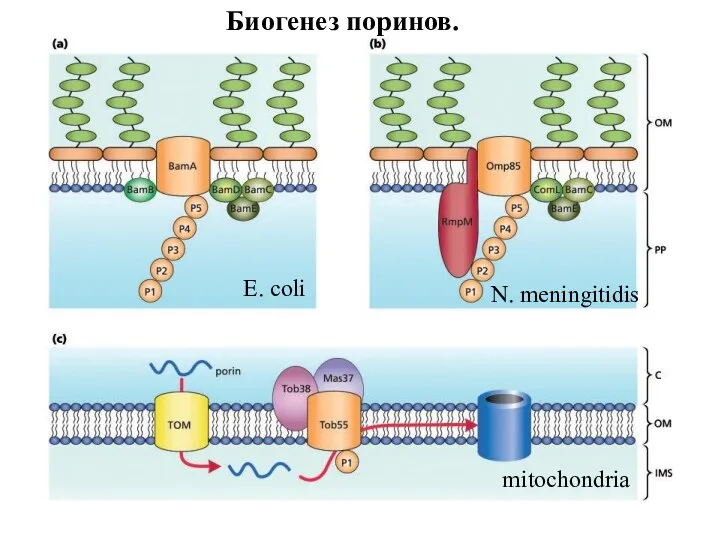
Слайд 97N. meningitidis
E. coli
mitochondriа
Биогенез поринов.
N. meningitidis
E. coli
mitochondriа
Биогенез поринов.
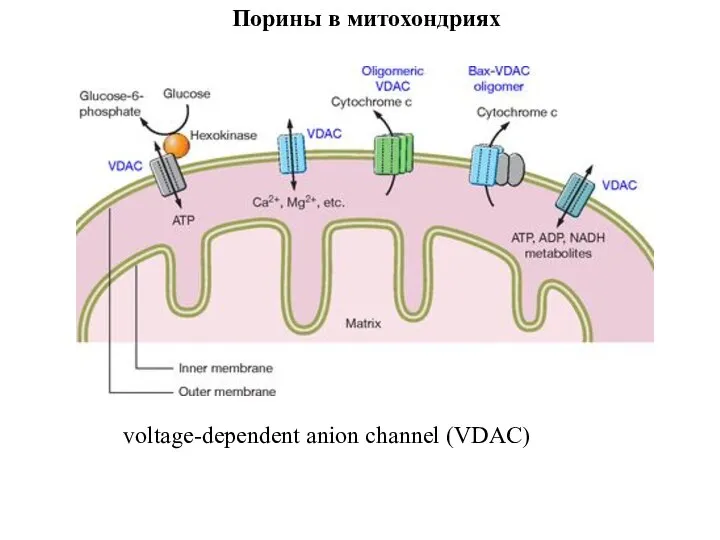
Слайд 98Порины в митохондриях
voltage-dependent anion channel (VDAC)
Порины в митохондриях
voltage-dependent anion channel (VDAC)
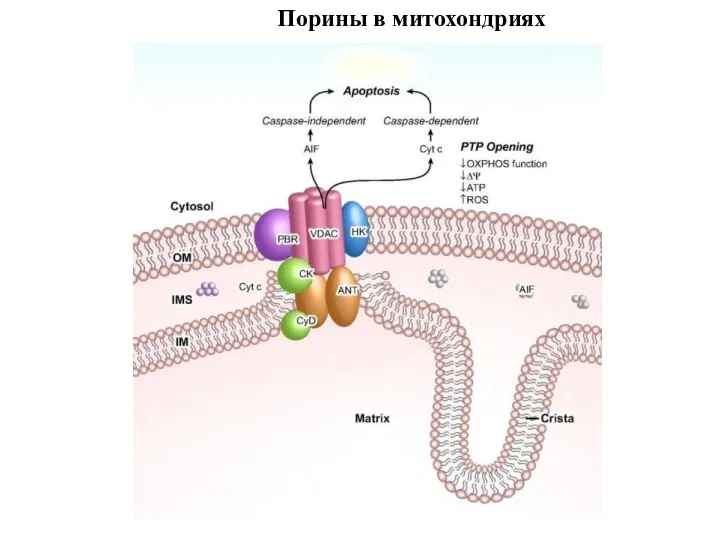
Слайд 99Порины в митохондриях
Порины в митохондриях
Слайд 100Model of the mechanism of uniport transport by GLUT1, which is believed
Model of the mechanism of uniport transport by GLUT1, which is believed
 Эндокринная система
Эндокринная система Белки
Белки Виртуальное путешествие в микромир
Виртуальное путешествие в микромир Могут ли растения производить потомство без помощи семян?
Могут ли растения производить потомство без помощи семян? Чудеса родного края
Чудеса родного края Этапы пищеварения
Этапы пищеварения Презентация на тему "Класс Двустворчатые моллюски" - презентации по Биологии
Презентация на тему "Класс Двустворчатые моллюски" - презентации по Биологии Анатомия и физиология печени и поджелудочной железы
Анатомия и физиология печени и поджелудочной железы Строение тела человека как единая система
Строение тела человека как единая система Гигиена питания
Гигиена питания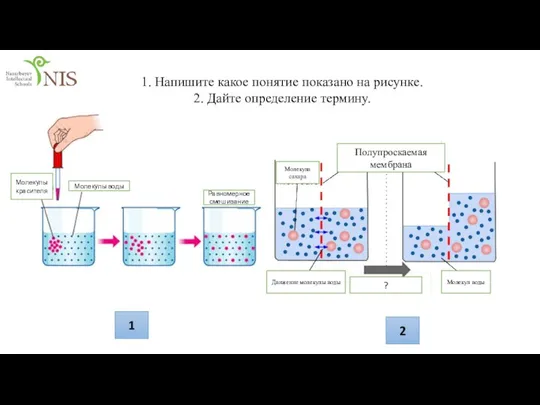 презен транспорт рус (Мырзаханулы Асан)
презен транспорт рус (Мырзаханулы Асан) Синичкин день
Синичкин день Анализ лесовосстановительных процессов в дубравах Южного Урала с применением молекулярно-биологических методов исследований
Анализ лесовосстановительных процессов в дубравах Южного Урала с применением молекулярно-биологических методов исследований Экоцарство - природное государство. Экологическая мозаика
Экоцарство - природное государство. Экологическая мозаика УМК И. Н. Пономаревой 7 класс концентрический курс
УМК И. Н. Пономаревой 7 класс концентрический курс Молекулярная биология
Молекулярная биология Половая система чекловека. Тест
Половая система чекловека. Тест Протисты, эукариотические организмы, не входящие в состав животных, грибов и растений
Протисты, эукариотические организмы, не входящие в состав животных, грибов и растений В лесу родилась елочка, или мир хвойных деревьев
В лесу родилась елочка, или мир хвойных деревьев Химия ферментов
Химия ферментов Мышечный аппарат глаза
Мышечный аппарат глаза Зимующие птицы
Зимующие птицы Размножение и развитие растений
Размножение и развитие растений Африканский страус
Африканский страус Биологический механизм запахов
Биологический механизм запахов Координация и регуляция. Структура и функции палочек и колбочек, волосковых клеток
Координация и регуляция. Структура и функции палочек и колбочек, волосковых клеток Презентация на тему Национальный парк «Беловежская пуща»
Презентация на тему Национальный парк «Беловежская пуща»  Менералды заттек (сілтілі)
Менералды заттек (сілтілі)