- Молекулы адгезии

Содержание
- 2. В ходе развития иммунного ответа разные клетки взаимодействуют друг с другом. Известно как минимум 2 механизма
- 3. Молекулы иммунной системы: факторы межклеточного взаимодействия I. На поверхности клеток: (рецепторы-лиганды) а) Молекулы иммуноглобулинового суперсемейства: 1.
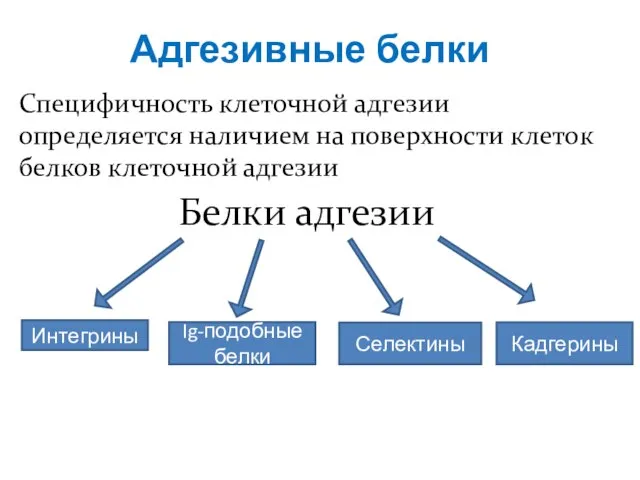
- 4. Адгезивные белки Специфичность клеточной адгезии определяется наличием на поверхности клеток белков клеточной адгезии Белки адгезии Интегрины
- 5. Клеточная адгезия – это соединение клеток, приводящее к формированию определённых правильных типов гистологических структур, специфичных для
- 6. Соединения клеток образуют пути сообщения, позволяя клеткам обмениваться сигналами, координирующими их поведение и регулирующими экспрессию генов.
- 7. Активным участником воспалительной реакции являются эндотелиальные клетки сосудов. Нарушение функции эндотелиальных клеток или их деструкция приводят
- 8. Адгезия клеток - процесс, когда мембранные молекулы одной клетки комплементарно связываются с мембранными молекулами другой клетки
- 9. Когда нормальные клетки прекращают делиться? Гены, включающие пролиферацию выключаются, если клетка получает сигнал, что все точки
- 10. Типы соединения молекул Адгезия может осуществляться на основе двух механизмов: а) гомофильного – молекулы адгезии одной
- 11. Клеточные контакты Клетка – клетка 1) Контакты простого типа: а) адгезионные б) интердигитация (пальцевые соединения) 2)
- 12. Архитектурные типы тканей Эпителиальные Много клеток – мало межклеточного вещества Межклеточные контакты Соединительные Много межклеточного вещества
- 13. Общая схема строения клеточных контактов Межклеточные контакты, а также контакты клетки с межклеточными контактами образуются по
- 14. Контакты простого типа. Адгезионные соединения Это простое сближение плазмолемм соседних клеток на расстояние 15-20 нм без
- 15. Интердигитация (пальцевидное соединение) (на рисунке) представляет собой контакт, при котором плазмолемма двух клеток, сопровождая друг друга,
- 16. Контакты простого типа Встречаются в эпителиальных тканях, здесь они образуют вокруг каждой клетки поясок (зона прилипания);
- 17. Контакты сцепляющего типа Десмосомы Полудесмосомы Поясок сцепления
- 18. Контакты сцепляющего типа Десмосома Десмосома представляет собой небольшое округлое образование, содержащее специфические внутри- и межклеточные элементы.
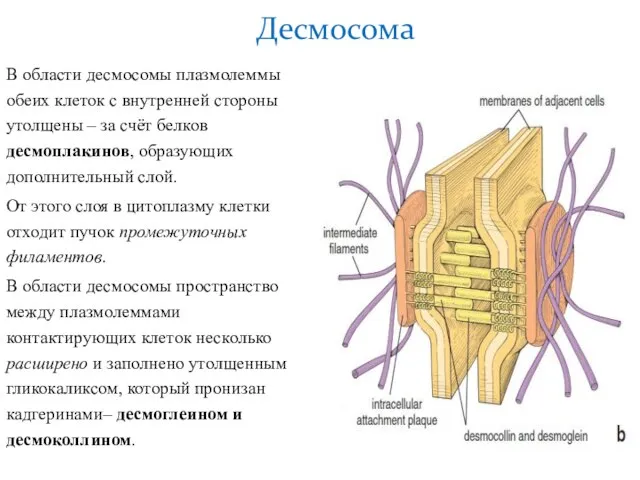
- 19. В области десмосомы плазмолеммы обеих клеток с внутренней стороны утолщены – за счёт белков десмоплакинов, образующих
- 20. Полудесмосома Полудесмосома обеспечивает контакт клеток с базальной мембраной. По структуре гемидесмосомы напоминают десмосомы и тоже содержат
- 21. Полудесмосома
- 22. Поясок сцепления Адгезивный поясок, (поясок сцепления, поясная десмосома) (zonula adherens), – парное образование в виде лент,
- 23. Белки поясков сцепления 1. Утолщение плазмолеммы со стороны цитоплазмы образовано винкулином; 2. Нити, отходящие в цитоплазму
- 24. Сравнительная таблица контактов сцепляющего типа
- 25. Контакты сцепляющего типа Десмосомы образуются между клетками тканей, подвергающихся механическим воздействиям (эпителиальные клетки, клетки сердечной мышцы);
- 26. Контакт запирающего типа Плотный контакт Плазмолеммы клеток прилегают друг к другу вплотную, сцепляясь с помощью специальных
- 27. Белки плотного контакта Основными белками плотных контактов являются клаудины и окклюдины. Через ряд специальных белков к
- 28. Контакты коммуникационного типа Щелевидные соединения (нексусы, электрические синапсы, эфапсы) Нексус имеет форму круга диаметром 0,5-0,3 мкм.
- 29. Таким образом, система клеточных контактов, механизмов клеточной адгезии и внеклеточного матрикса играет принципиальную роль во всех
- 30. Молекулы адгезии формируют несколько достаточно консервативных семейств. У млекопитающих известно 4 группы молекул адгезии : селектины
- 31. Характеристика факторов межклеточного взаимодействия иммунной системы Адгезины клеток иммунной системы (ICAM) CD – молекулы лейкоцитов и
- 32. Характеристика факторов межклеточного взаимодействия иммунной системы Селектины Небольшое семейство, представленное тремя молекулами, экспрессированными соответственно на: активированном
- 33. Селектины Селектины — трансмембранные белки на поверхности лимфоцитов, лейкоцитов и эндотелиоцитов. Общим для них является наличие
- 34. Селектины Р-селектин участвует в активации тромбоцитов и ранних этапах миграции лейкоцитов в очаг воспаления. Е-селектин —
- 35. Характеристика факторов межклеточного взаимодействия иммунной системы
- 36. Рецепторами L-селектинов служат вещества, называемые адрессинами — PNAd. Селектины и адрессины обеспечивают селективную адгезию клеток у
- 37. L-селектин формирует слабую связь с адрессинами; к тому же его молекула, как упомянуто выше, легко смывается
- 38. Характеристика факторов межклеточного взаимодействия иммунной системы Интегрины: Главные молекулы, опосредующие взаимодействие клеток с межклеточным веществом: они
- 39. Интегрины Интегрины — гетеродимерные белки, состоящие из крупной а-цепи и меньшей по размеру бета-цепи Интегрины —
- 40. В качестве рецепторов для β2-интегринов выступают мембранные молекулы, образующие группу ICAM содержащую 5 видов ICAM-1 (CD54),
- 41. Интегрины существуют в клетке в неактивной и активной формах (для некоторых интегринов описано промежуточное состояние). В
- 42. ИНТЕГРИНЫ Такой контакт не только обеспечивает надежную адгезию взаимодействующих клеток, но и служит источником вспомогательных сигналов,
- 43. АДГЕЗИЯ ЛЕЙКОЦИТОВ И ЭКСТРАВАЗАЦИЯ I этап: замедление движения лейкоцитов. Низкоаффинное взаимодействие адгезинов эндотелиальных клеток с лигандами
- 44. АДГЕЗИЯ ЛЕЙКОЦИТОВ И ЭКСТРАВАЗАЦИЯ
- 45. Эмиграция эффекторной Т-клетки в ткань при вирусной инфекции
- 46. Кадгерины Кадгерины проявляют свою адгезионную способность только в присутствии ионов Ca2+. По структуре классический кадгерин представляет
- 47. Таким образом, система клеточных контактов, механизмов клеточной адгезии и внеклеточного матрикса играет принципиальную роль во всех
- 49. Скачать презентацию

Слайд 3Молекулы иммунной системы:
факторы межклеточного взаимодействия
I. На поверхности клеток: (рецепторы-лиганды)
а) Молекулы иммуноглобулинового
Молекулы иммунной системы:
факторы межклеточного взаимодействия
I. На поверхности клеток: (рецепторы-лиганды)
а) Молекулы иммуноглобулинового

Слайд 4Адгезивные белки
Специфичность клеточной адгезии определяется наличием на поверхности клеток белков клеточной адгезии
Белки
Адгезивные белки
Специфичность клеточной адгезии определяется наличием на поверхности клеток белков клеточной адгезии
Белки
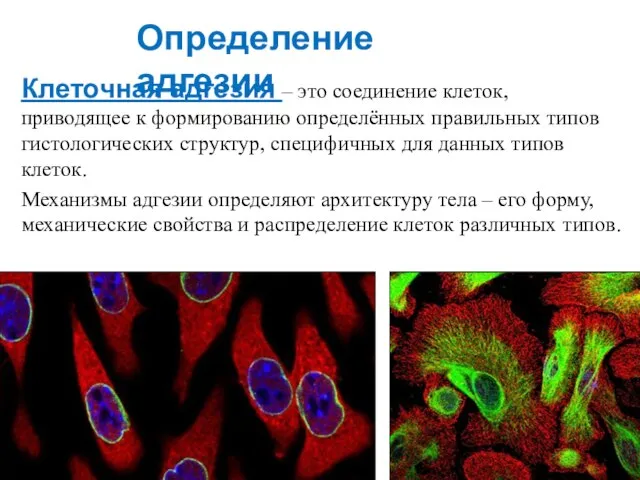
Слайд 5Клеточная адгезия – это соединение клеток, приводящее к формированию определённых правильных типов
Клеточная адгезия – это соединение клеток, приводящее к формированию определённых правильных типов
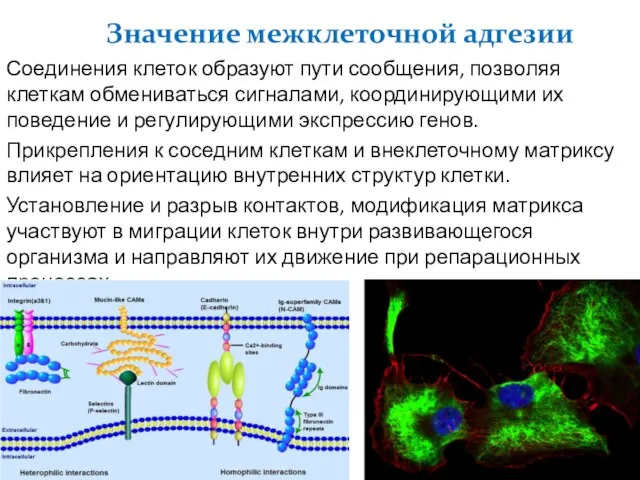
Слайд 6Соединения клеток образуют пути сообщения, позволяя клеткам обмениваться сигналами, координирующими их поведение
Соединения клеток образуют пути сообщения, позволяя клеткам обмениваться сигналами, координирующими их поведение
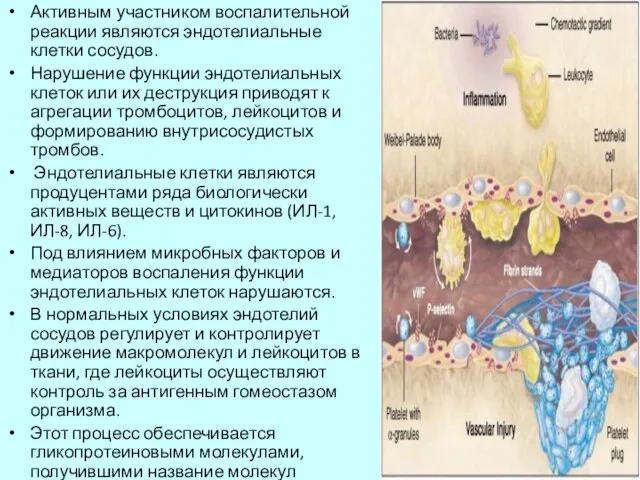
Слайд 7Активным участником воспалительной реакции являются эндотелиальные клетки сосудов.
Нарушение функции эндотелиальных клеток
Активным участником воспалительной реакции являются эндотелиальные клетки сосудов.
Нарушение функции эндотелиальных клеток
Слайд 8Адгезия клеток - процесс, когда мембранные молекулы одной клетки комплементарно связываются с
Адгезия клеток - процесс, когда мембранные молекулы одной клетки комплементарно связываются с

Слайд 9Когда нормальные клетки прекращают делиться?
Гены, включающие пролиферацию выключаются, если клетка получает сигнал,
Когда нормальные клетки прекращают делиться?
Гены, включающие пролиферацию выключаются, если клетка получает сигнал,

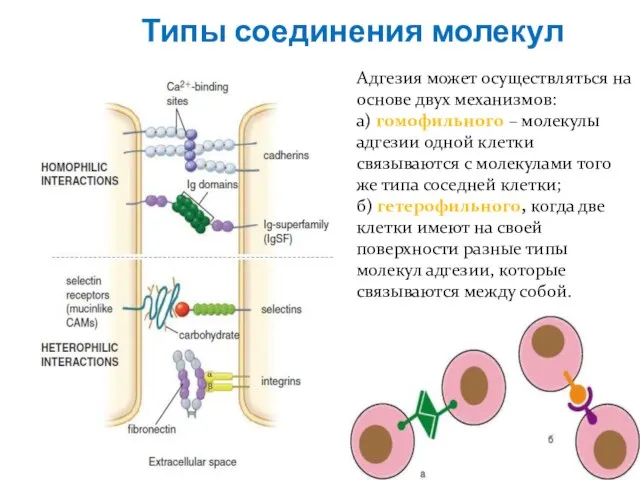
Слайд 10Типы соединения молекул
Адгезия может осуществляться на основе двух механизмов:
а) гомофильного – молекулы
Типы соединения молекул
Адгезия может осуществляться на основе двух механизмов:
а) гомофильного – молекулы
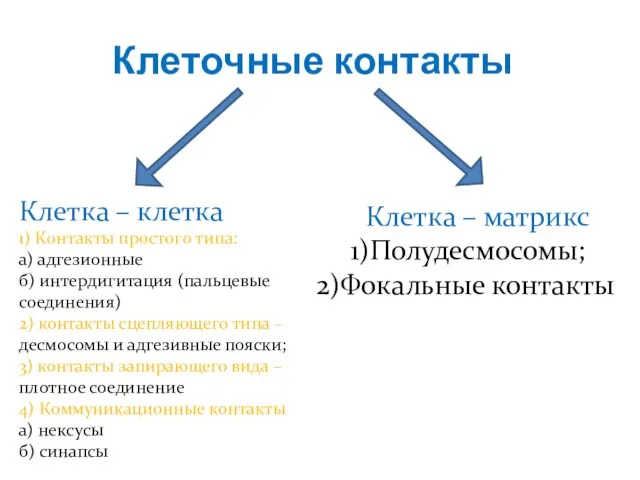
Слайд 11Клеточные контакты
Клетка – клетка
1) Контакты простого типа:
а) адгезионные
б) интердигитация (пальцевые соединения)
2) контакты
Клеточные контакты
Клетка – клетка
1) Контакты простого типа:
а) адгезионные
б) интердигитация (пальцевые соединения)
2) контакты
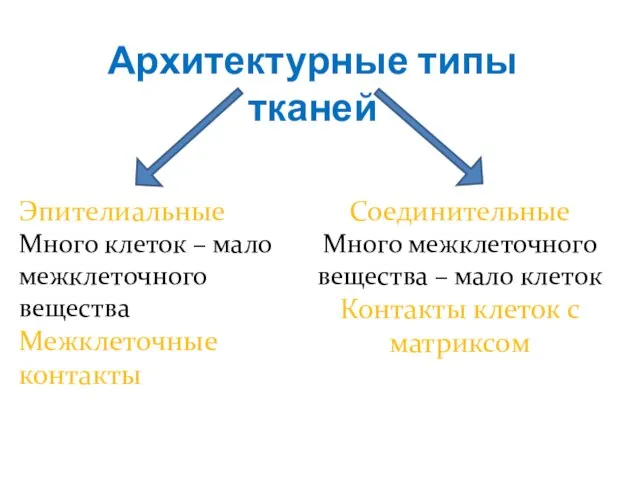
Слайд 12Архитектурные типы тканей
Эпителиальные
Много клеток – мало межклеточного вещества
Межклеточные контакты
Соединительные
Много межклеточного вещества –
Архитектурные типы тканей
Эпителиальные
Много клеток – мало межклеточного вещества
Межклеточные контакты
Соединительные
Много межклеточного вещества –
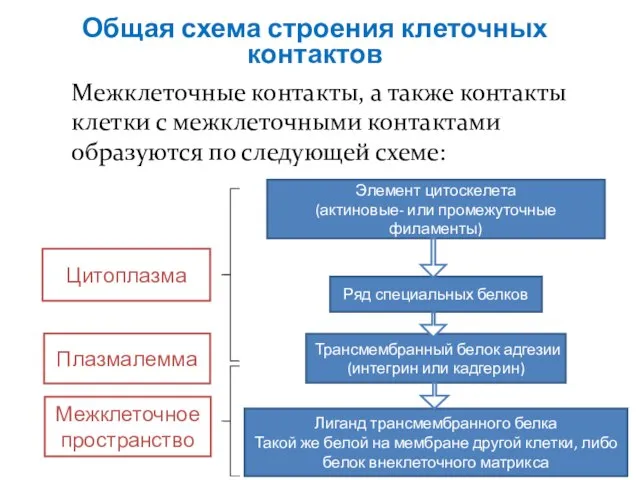
Слайд 13Общая схема строения клеточных контактов
Межклеточные контакты, а также контакты клетки с межклеточными
Общая схема строения клеточных контактов
Межклеточные контакты, а также контакты клетки с межклеточными
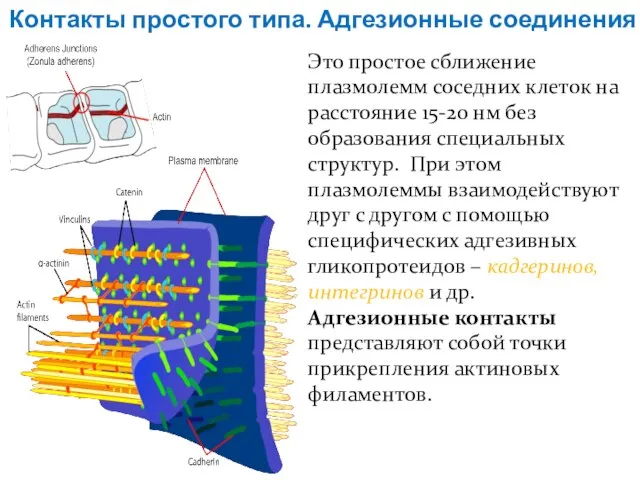
Слайд 14Контакты простого типа. Адгезионные соединения
Это простое сближение плазмолемм соседних клеток на расстояние
Контакты простого типа. Адгезионные соединения
Это простое сближение плазмолемм соседних клеток на расстояние
Слайд 15Интердигитация (пальцевидное соединение) (на рисунке) представляет собой контакт, при котором плазмолемма двух
Интердигитация (пальцевидное соединение) (на рисунке) представляет собой контакт, при котором плазмолемма двух

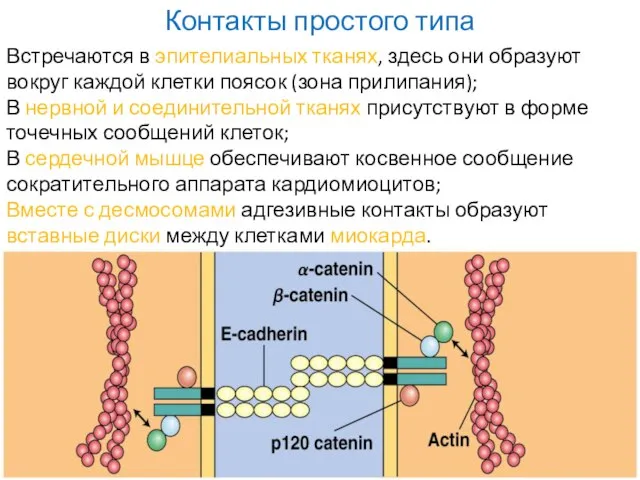
Слайд 16Контакты простого типа
Встречаются в эпителиальных тканях, здесь они образуют вокруг каждой клетки
Контакты простого типа
Встречаются в эпителиальных тканях, здесь они образуют вокруг каждой клетки
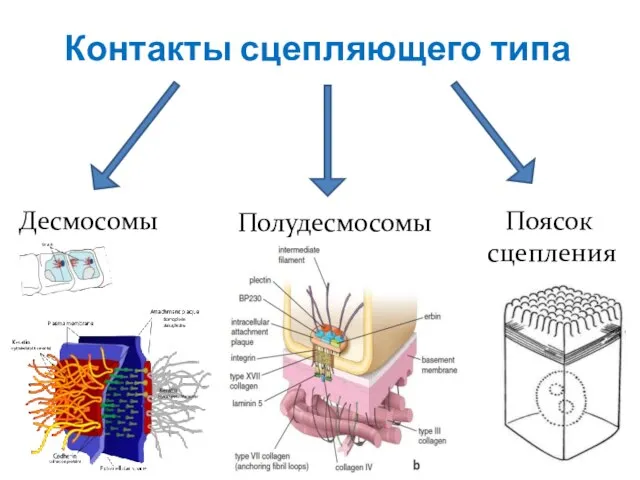
Слайд 17Контакты сцепляющего типа
Десмосомы
Полудесмосомы
Поясок
сцепления
Контакты сцепляющего типа
Десмосомы
Полудесмосомы
Поясок
сцепления
Слайд 18Контакты сцепляющего типа
Десмосома
Десмосома представляет собой небольшое округлое образование, содержащее специфические внутри- и
Контакты сцепляющего типа
Десмосома
Десмосома представляет собой небольшое округлое образование, содержащее специфические внутри- и
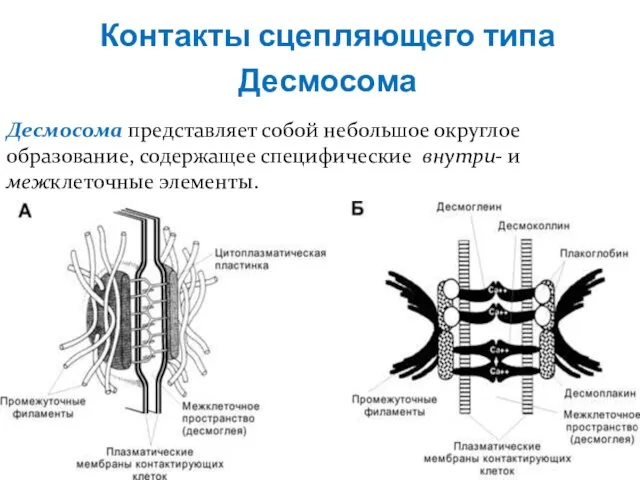
Слайд 19В области десмосомы плазмолеммы обеих клеток с внутренней стороны утолщены – за
В области десмосомы плазмолеммы обеих клеток с внутренней стороны утолщены – за
Слайд 20Полудесмосома
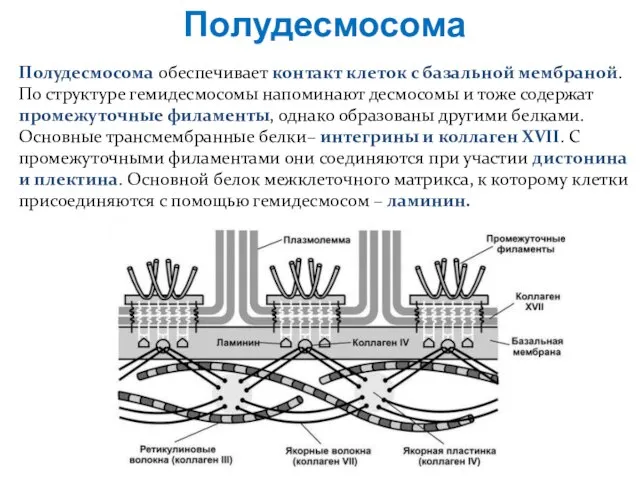
Полудесмосома обеспечивает контакт клеток с базальной мембраной.
По структуре гемидесмосомы напоминают десмосомы и
Полудесмосома
Полудесмосома обеспечивает контакт клеток с базальной мембраной.
По структуре гемидесмосомы напоминают десмосомы и
Слайд 21Полудесмосома
Полудесмосома
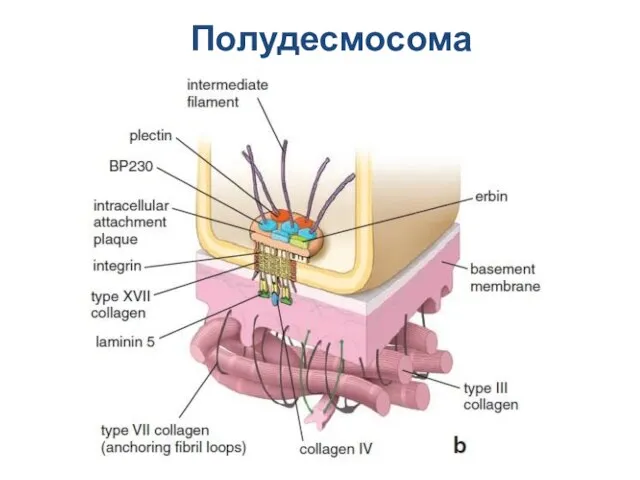
Слайд 22Поясок сцепления
Адгезивный поясок, (поясок сцепления, поясная десмосома) (zonula adherens), – парное образование
Поясок сцепления
Адгезивный поясок, (поясок сцепления, поясная десмосома) (zonula adherens), – парное образование
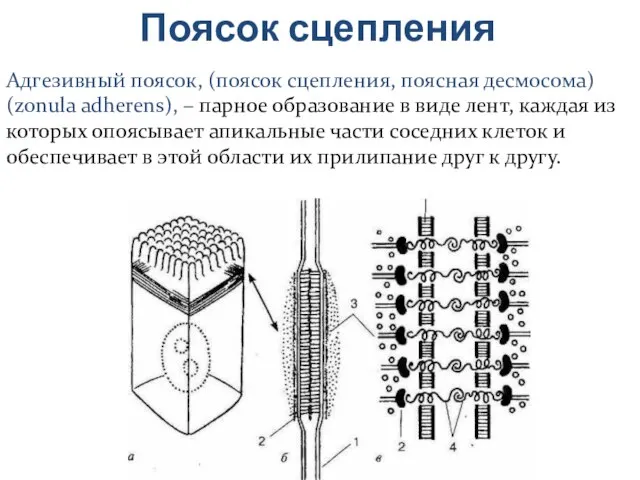
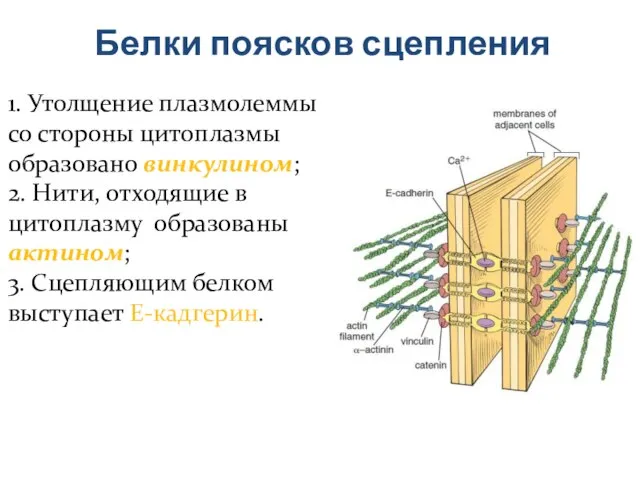
Слайд 23Белки поясков сцепления
1. Утолщение плазмолеммы со стороны цитоплазмы образовано винкулином;
2. Нити, отходящие
Белки поясков сцепления
1. Утолщение плазмолеммы со стороны цитоплазмы образовано винкулином;
2. Нити, отходящие
Слайд 24Сравнительная таблица контактов сцепляющего типа
Сравнительная таблица контактов сцепляющего типа
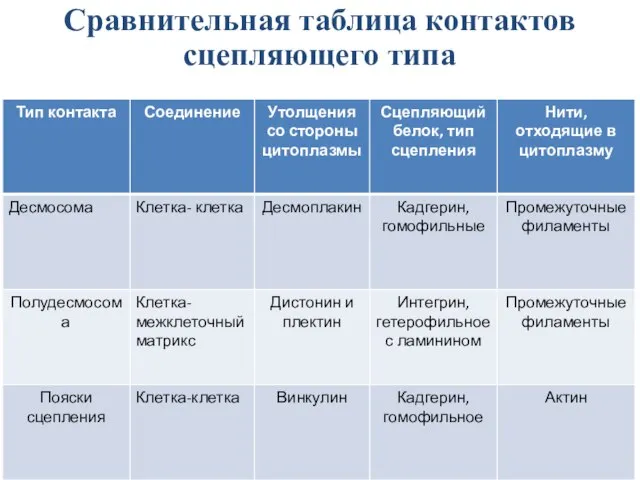
Слайд 25Контакты сцепляющего типа
Десмосомы образуются между клетками тканей, подвергающихся механическим воздействиям (эпителиальные клетки,
Контакты сцепляющего типа
Десмосомы образуются между клетками тканей, подвергающихся механическим воздействиям (эпителиальные клетки,

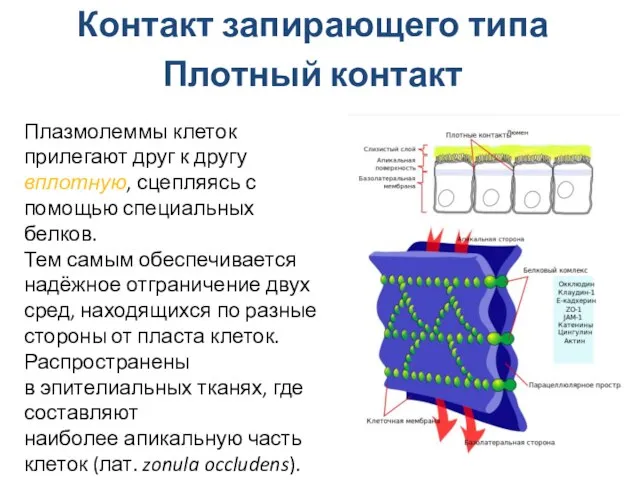
Слайд 26Контакт запирающего типа
Плотный контакт
Плазмолеммы клеток прилегают друг к другу вплотную, сцепляясь с
Контакт запирающего типа
Плотный контакт
Плазмолеммы клеток прилегают друг к другу вплотную, сцепляясь с
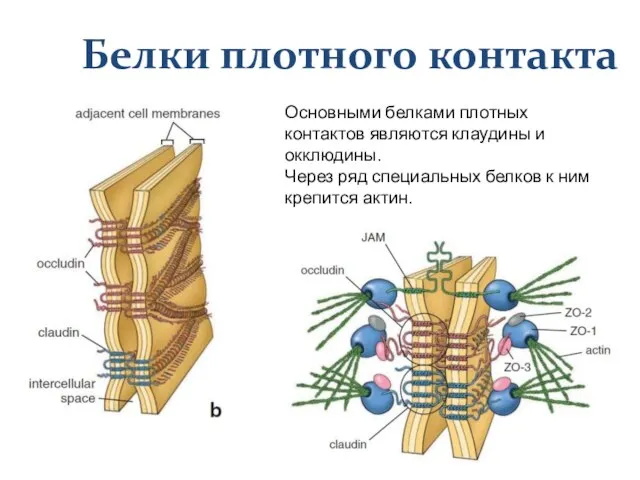
Слайд 27Белки плотного контакта
Основными белками плотных контактов являются клаудины и окклюдины.
Через ряд специальных
Белки плотного контакта
Основными белками плотных контактов являются клаудины и окклюдины.
Через ряд специальных
Слайд 28Контакты коммуникационного типа
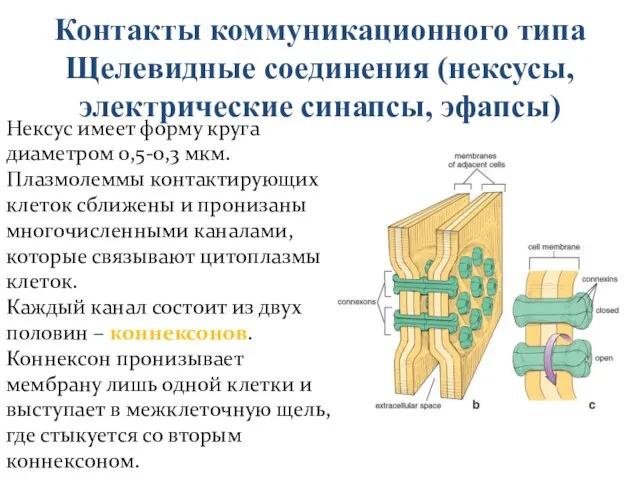
Щелевидные соединения (нексусы, электрические синапсы, эфапсы)
Нексус имеет форму круга диаметром
Контакты коммуникационного типа
Щелевидные соединения (нексусы, электрические синапсы, эфапсы)
Нексус имеет форму круга диаметром
Слайд 29Таким образом, система клеточных контактов, механизмов клеточной адгезии и внеклеточного матрикса играет
Таким образом, система клеточных контактов, механизмов клеточной адгезии и внеклеточного матрикса играет
Слайд 30Молекулы адгезии формируют несколько достаточно консервативных семейств.
У млекопитающих известно 4 группы
Молекулы адгезии формируют несколько достаточно консервативных семейств.
У млекопитающих известно 4 группы

Слайд 31Характеристика факторов межклеточного взаимодействия иммунной системы
Адгезины клеток иммунной системы (ICAM)
CD –
Характеристика факторов межклеточного взаимодействия иммунной системы
Адгезины клеток иммунной системы (ICAM)
CD –

Слайд 32Характеристика факторов межклеточного взаимодействия иммунной системы
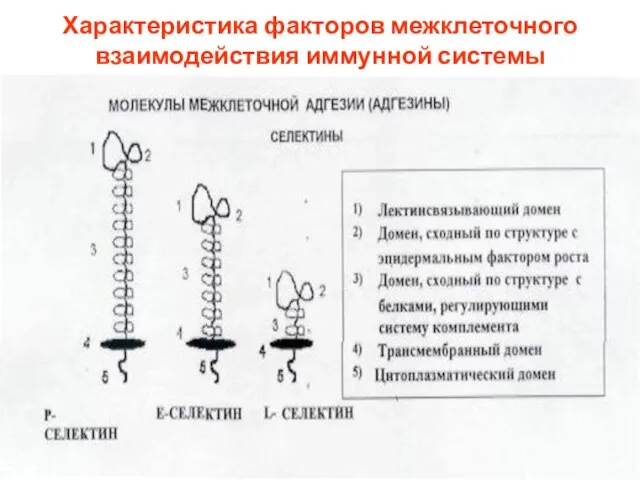
Селектины
Небольшое семейство, представленное тремя молекулами, экспрессированными соответственно
Характеристика факторов межклеточного взаимодействия иммунной системы
Селектины
Небольшое семейство, представленное тремя молекулами, экспрессированными соответственно

Слайд 33Селектины
Селектины — трансмембранные белки на поверхности лимфоцитов, лейкоцитов и эндотелиоцитов.
Общим для
Селектины
Селектины — трансмембранные белки на поверхности лимфоцитов, лейкоцитов и эндотелиоцитов.
Общим для

Слайд 34Селектины
Р-селектин участвует в активации тромбоцитов и ранних этапах миграции лейкоцитов в очаг
Селектины
Р-селектин участвует в активации тромбоцитов и ранних этапах миграции лейкоцитов в очаг

Слайд 35Характеристика факторов межклеточного взаимодействия иммунной системы
Характеристика факторов межклеточного взаимодействия иммунной системы
Слайд 36Рецепторами L-селектинов служат вещества, называемые адрессинами — PNAd.
Селектины и адрессины обеспечивают
Рецепторами L-селектинов служат вещества, называемые адрессинами — PNAd.
Селектины и адрессины обеспечивают

Слайд 37L-селектин формирует слабую связь с адрессинами; к тому же его молекула, как
L-селектин формирует слабую связь с адрессинами; к тому же его молекула, как

Слайд 38Характеристика факторов межклеточного взаимодействия иммунной системы
Интегрины:
Главные молекулы, опосредующие взаимодействие клеток с межклеточным
Характеристика факторов межклеточного взаимодействия иммунной системы
Интегрины:
Главные молекулы, опосредующие взаимодействие клеток с межклеточным

Слайд 39Интегрины
Интегрины — гетеродимерные белки, состоящие из крупной а-цепи и меньшей по размеру
Интегрины
Интегрины — гетеродимерные белки, состоящие из крупной а-цепи и меньшей по размеру

Слайд 40В качестве рецепторов для β2-интегринов выступают мембранные молекулы, образующие группу ICAM содержащую
В качестве рецепторов для β2-интегринов выступают мембранные молекулы, образующие группу ICAM содержащую

Слайд 41Интегрины существуют в клетке в неактивной и активной формах (для некоторых интегринов
Интегрины существуют в клетке в неактивной и активной формах (для некоторых интегринов

Слайд 42ИНТЕГРИНЫ
Такой контакт не только обеспечивает надежную адгезию взаимодействующих клеток, но и служит
ИНТЕГРИНЫ
Такой контакт не только обеспечивает надежную адгезию взаимодействующих клеток, но и служит

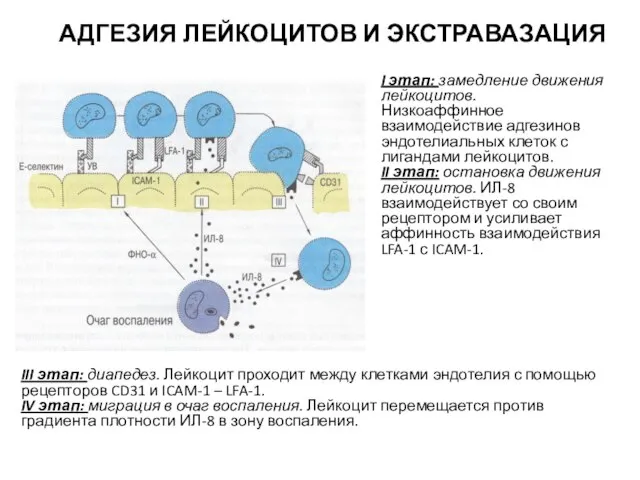
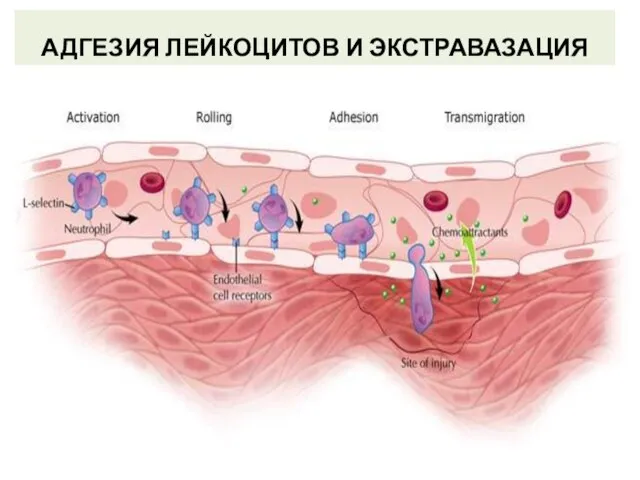
Слайд 43АДГЕЗИЯ ЛЕЙКОЦИТОВ И ЭКСТРАВАЗАЦИЯ
I этап: замедление движения лейкоцитов. Низкоаффинное взаимодействие адгезинов эндотелиальных
АДГЕЗИЯ ЛЕЙКОЦИТОВ И ЭКСТРАВАЗАЦИЯ
I этап: замедление движения лейкоцитов. Низкоаффинное взаимодействие адгезинов эндотелиальных
Слайд 44АДГЕЗИЯ ЛЕЙКОЦИТОВ И ЭКСТРАВАЗАЦИЯ
АДГЕЗИЯ ЛЕЙКОЦИТОВ И ЭКСТРАВАЗАЦИЯ
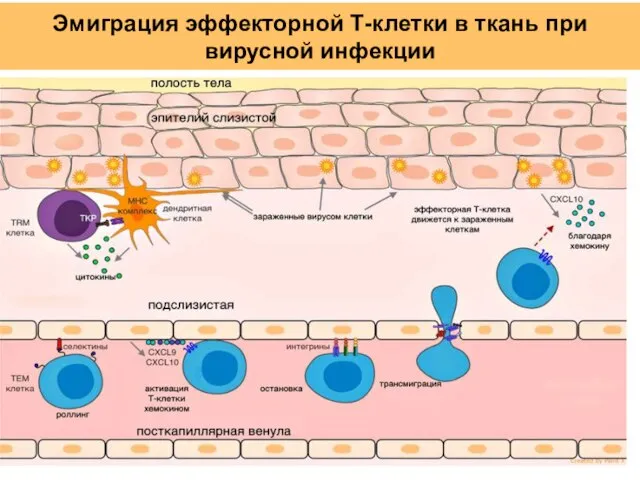
Слайд 45Эмиграция эффекторной Т-клетки в ткань при вирусной инфекции
Эмиграция эффекторной Т-клетки в ткань при вирусной инфекции
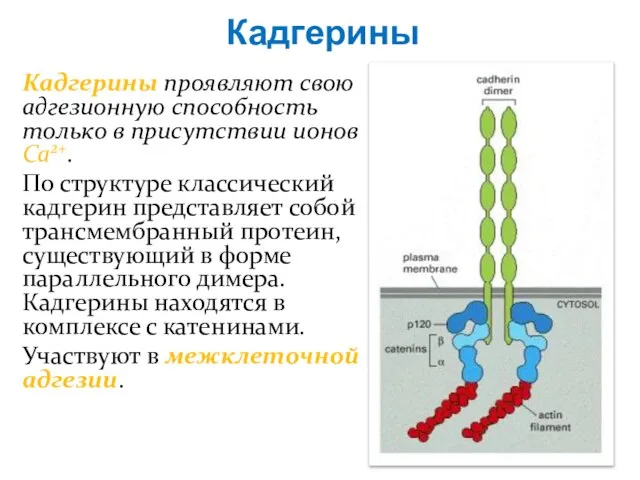
Слайд 46Кадгерины
Кадгерины проявляют свою адгезионную способность только в присутствии ионов Ca2+.
По структуре
Кадгерины
Кадгерины проявляют свою адгезионную способность только в присутствии ионов Ca2+.
По структуре
Слайд 47Таким образом, система клеточных контактов, механизмов клеточной адгезии и внеклеточного матрикса играет
Таким образом, система клеточных контактов, механизмов клеточной адгезии и внеклеточного матрикса играет
 Позахромосомна спадковість у людини
Позахромосомна спадковість у людини Домашние птицы
Домашние птицы Мезозойская эра
Мезозойская эра Питание и пищеварение
Питание и пищеварение Парящие над облаками
Парящие над облаками Насекомые
Насекомые Вплив тривалості та якості сну на розумову працездатність учнів середнього шкільного віку
Вплив тривалості та якості сну на розумову працездатність учнів середнього шкільного віку Индивидуальная работа. Генетические коллекции гороха
Индивидуальная работа. Генетические коллекции гороха Фрукты
Фрукты Чудесные цветники весной
Чудесные цветники весной Генетика
Генетика Что общего у разных растений?
Что общего у разных растений? волки
волки Презентация на тему Знатоки природы
Презентация на тему Знатоки природы  Комбинативная изменчивость
Комбинативная изменчивость Основы генетики
Основы генетики Лист и цветок. Оплодотворение
Лист и цветок. Оплодотворение Иммунитет
Иммунитет Воздействие биологии на формирование новых норм, установок и ориентаций культуры
Воздействие биологии на формирование новых норм, установок и ориентаций культуры Клеточный цикл. Митоз. Наблюдение митоза в клетках кончика корешка лука на готовых микропрепаратах
Клеточный цикл. Митоз. Наблюдение митоза в клетках кончика корешка лука на готовых микропрепаратах Презентация на тему Сообщество или биоценоз
Презентация на тему Сообщество или биоценоз  Пропорции фигуры человека
Пропорции фигуры человека Обмен веществ и энергии в организме
Обмен веществ и энергии в организме Закономерности наследования признаков. Решение задач
Закономерности наследования признаков. Решение задач Решение задач по почвоведению
Решение задач по почвоведению Развитие животных
Развитие животных Влияние излучения от Wi-Fi роутера на рост и развитие живых организмов
Влияние излучения от Wi-Fi роутера на рост и развитие живых организмов Свойства живых организмов
Свойства живых организмов