- MOLEKUlyarnaya_biologia_LPZ_1

Содержание
- 2. Молекулярная биология – цель, задачи и связь с другими отраслями биологии, химии, физики. МБ - отрасль
- 3. Истоками молекулярной биологии являются: органическая химия, занимающаяся изучением химической структуры макромолекул; биохимия, целью которой является изучение
- 4. Исключит-ное внимание привлекают в наст. время 2 осн. класса макромолекул - белки и нуклеиновые кислоты: Белкам
- 5. 2.Доказательства генетической функции ДНК и РНК. Структура нуклеотидов. Принцип комплементарности. История изучения нуклеиновых кислот начинается с
- 6. 1. Английский. бактериолог Ф.Гриффит (1928) продемонстрировал следующий опыт: способность пневмококков к трансформации. Было выдвинуто предполож. о
- 7. Гриффит заключил, что живые микробы непатогенного штамма в присутствии клеток штамма патогенного приобретают наследственно закрепленные св-ва
- 9. О. Звери, К. Мак-Леод и М. Мак-Карти, в 1944 г. точно определили химич. природу «транс-формирующего агента»
- 10. 3. В 1950 г. американский биохимик Э.Чаргафф установил важнейшую закономерность химич. строения ДНК, согласно кот. сумма
- 11. В 1953 г. Дж. Уотсон и Ф. Крик на основании результатов рентгеноструктурного анализа и биохимич. данных
- 12. К настоящему времени : обнаружено, что ДНК мож. повреждаться и мож. восстанавливаться, молекулы ДНК мог. обменив.
- 13. Структура нуклеотидов. Нуклеиновые кислоты – это биологич. высокомолекуляр. полимерн. соединения, мономерами кот. явл. нуклеотиды. Нуклеотиды –
- 14. В нуклеиновых кислотах сахар представлен пентозой. В РНК пентоза является рибозой, а в ДНК – дезоксирибозой.
- 15. Изучая нуклеотидный состав ДНК различ. видов орг-мов, сделали след. выводы: 1. нуклеотидный состав ДНК разн. тканей
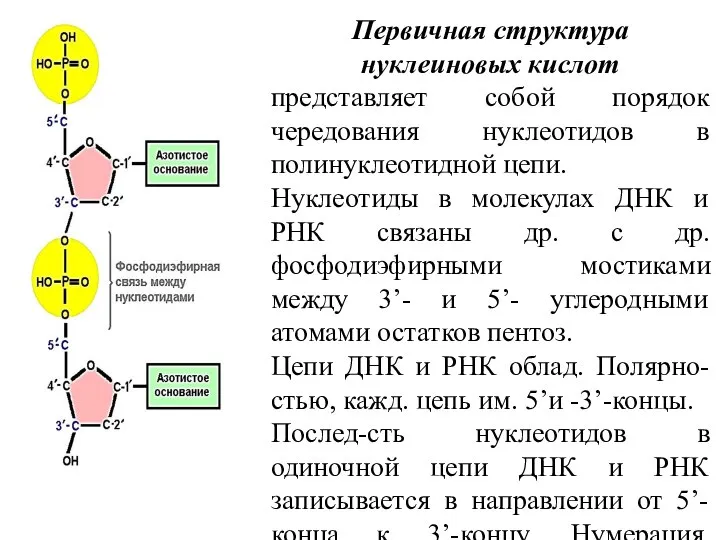
- 16. Первичная структура нуклеиновых кислот представляет собой порядок чередования нуклеотидов в полинуклеотидной цепи. Нуклеотиды в молекулах ДНК
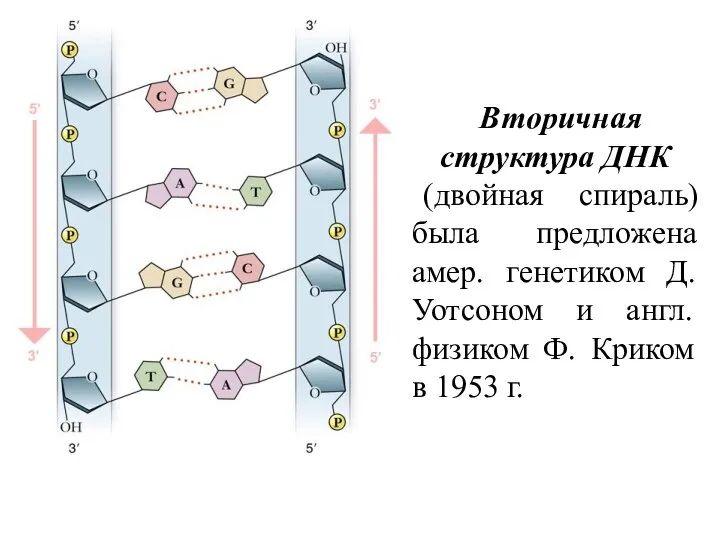
- 17. Вторичная структура ДНК (двойная спираль) была предложена амер. генетиком Д. Уотсоном и англ. физиком Ф. Криком
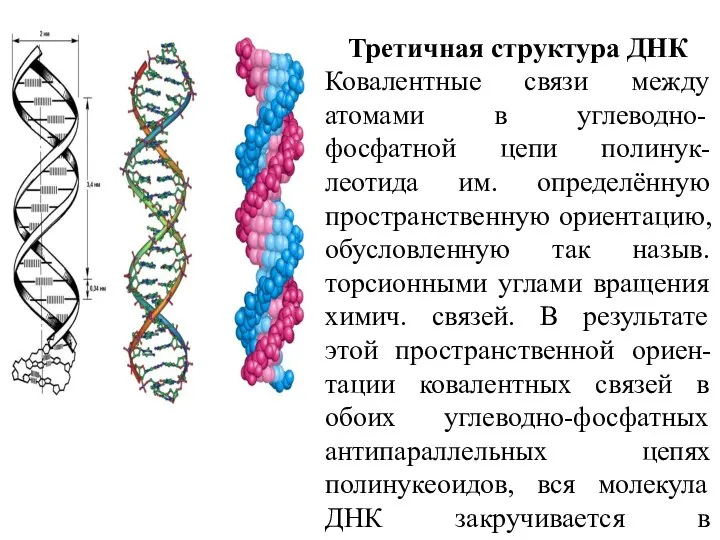
- 18. Третичная структура ДНК Ковалентные связи между атомами в углеводно-фосфатной цепи полинук-леотида им. определённую пространственную ориентацию, обусловленную
- 19. Формы ДНК А-форма ДНК: двухспиральная ДНК, содержащая прим. 11 остатков на 1 оборот спирали. В этой
- 21. Комплементарность – это пространственная взаимодополняемость молекул или их частей, приводящая к образованию водородных связей. Комплементарность каждой
- 22. Водородные связи, возникающие между пуринами и пиримидинами, удерживают комплементарные полинуклеотидные цепи в системе единой молекулы. Поскольку
- 23. Цепи ДНК (модель Уотсона и Крика) при закручивании в двойную спираль образ. большую и малую борозды,
- 24. 3. Коды ДНК. Гибкость двойной спирали. В 1954 г. амер. физик-теоретик Г.А. Гамов предпол., что кодирование
- 25. Генетич. код не содержит знаков препинания, и кодирующие триплеты следуют один за другим. Генетич. код явл.
- 26. Характеристика кода: 1. вырожденность (избыточность) - аминок-ты шифруются неск. триплетами. Вырожденность генетич. кода позволяет при мутационной
- 27. однонаправленность считывания (5′→3′), т.е. считывание информации с мРНК при синтезе белка происходит с её 5′-конца в
- 28. 4. Ассоциации ДНК с олигонуклетидами Олигонуклеотид - короткая однонитевая молекула ДНК и РНК (16-30 пар нуклеотидов).
- 29. Триплексы: специфичность образования Часто для прикладных задач необходимо, чтобы олигонуклеотид связался с ДНК в одном или
- 30. Выделяют 3 основных типа ошибок в узнавании олигонуклеотидом двухцепочечной ДНК.: неправильное спаривание в середине сайта, при
- 31. Существует неск. подходов к проблеме неспецифичности связывания олигонуклеотидов. скорость образования специфич. комплексов больше, чем неспецифич., а
- 32. специфичный комплекс еще достаточно стабилен, а неспецифич. уже нет. Напр. при температурах, близких к темпер. плавления,
- 33. Антисмысловой олигонуклеотид (antisense oligonucleotide) - олигонуклеотид, кот. комплементарен фрагменту мРНК, благодаря чему способен образовывать с ней
- 34. Хугстиновское спаривание оснований (Hoogsteen base pairing) (по им. К. Хугстина) - вариация спаривания нуклеотидов в нуклеиновых
- 35. В двуспиральных структурах таких пар не обнаружено, однако не исключается возможность образования тройных спиралей, в кот.
- 36. ДНК: H-форма H-форма ДНК – неканонич. структура в гомопурин- гомопиримидиновых участках ДНК. В нач. 80-х годов
- 37. Кроме того используют методы: Двумерный электрофорез H-формы ДНК H-форма ДНК модели с помощью зондирования различными химическими
- 38. 5. Ассоциация ДНК с белками Транскрипционные факторы: Принципы классификации Специфич-сть взаимодействия транскрип-ционных факторов с распознаваемыми ими
- 39. ДНК-связывающих доменов разделены на 4 суперкласса: Суперкласс 1. Факторы, ДНК-связывающий домен кот. обогащен положит-но заряженными аминок-тными
- 40. Комплекс лейциновой застёжки (показана синим цветом) с ДНК. Остатки лейцина, обеспечивающ. закреп-ление белков. спиралей обознач. красным
- 41. Цинковый палец (от. англ. цинк. палец) - тип белковой структуры, небольш. белковый мотив, или одним стабилизированный
- 42. Цинк. палец типа Cys 2 Его 2 включает альфа-спираль и антипараллель-ную бета-структуру. Ион цинка связан коор-динацион.
- 43. Факторы транскрипции с доменом "спираль-поворот-спираль» Тип пептидных доменов, специфически распознающих регуляторные послед-сти на ДНК, характерен для
- 44. Структуры полипептидных доменов типа (а) "спираль-поворот-спираль" (б) "лейциновая застежка" L - остатки Leu
- 45. Транскрипционные факторы: краткое определение Факторы транскрипции (transcription factors) - белки или белковые комплексы, непосредственно не участвующие
- 46. По функциональному признаку различают три класса транскрипционных факторов: а) основные, обеспечивающие нерегулируемый базальный уровень транскрипции и
- 47. Транскрипционный фактор – это белок, кот. после его перемещения в ядро клетки регулирует транскрипцию, специфически взаимодействуя
- 48. Антигены: силы взаимодействия с антителами (электростатическое притяжение) В основе взаимодействия антиген - антитело лежат те же
- 49. 3. Гидрофобные взаимодействия между неполярными гидрофобными группами, обеспечивающие по некоторым оценкам до 50% сродства между антителом
- 50. Организация хроматина эукариот. Вопрос о структурной организации хроматина в интерфазных ядрах далек от своего разрешения. Это
- 51. ХРОМАТИН, нуклеопротеид клеточ. ядра, составл. основу хромосом. В состав X. входят: ДНК (30-40% по массе), гистоны
- 52. Согласно пространств. модели А. Клуга кор-частица выглядит как плоский диск диам. 11 нм, толщ. 5,7 нм,
- 53. Гистоны составляют больш. осн. белков хроматина и находятся примерно в том же кол-ве, что и ДНК.
- 54. Гистоны первых четырех классов им. значит-ное кол-во как кислых, так и основных аминок-т. Поэтому эти белки
- 55. Пятый класс представлен гистонами, оч. богатыми лизином; он сост. из неск. достаточно близко-родственных белков с перекрывающимися
- 56. негистоны - это все др. белки хроматина. Предполагается поэтому, что они облад. большими видовыми и тканевыми
- 57. Упаковка генетич. материала достигается путем спирализации (конденсации) Первый уровень упаковки ДНК - нуклеосомный. Нуклеосома предст. собой
- 58. Третий уровень упаковки - хроматидный (петлевой). Супернуклеосомная нить спирализуется с образованием петель и изгибов. Она составляет
- 59. Схема строения нуклеосомы: 1 – нуклеосомная частица; 2 – октамер, охватывающий четыре пары гистонов; 3 –
- 60. Схема начальных уровней компактизации хроматина: 1 – нуклеосомный; 2 – нуклеомерный; 3 – хромомерный (петлевой домен);
- 62. Сборка нуклеосом 1 способ. обусловлен способностью тетрамера Н32Н42 организовывать ДНК в частицы, которые несколько напоминают минимальную
- 63. Антитела, полученные против этого белка, реагируют с белками нуклеоплазмы многих эукариот. След-но, можно предположить, что этот
- 64. Фейзинг. Терм. "фазирование" обознач. неслучайное располож. нуклеосом относит-но конкретной послед-сти нуклеотидов ДНК в определенных участках генома.
- 65. Во время транскрипции часть нуклеосомных белков остается связанной с ДНК. Нуклеосомы как частицы видны на хроматиновых
- 66. Предлагаются 2 варианта изменения структуры нуклеосом при синтезе РНК: нуклеосома «расщепляется» на 2 полунуклеосомы, а ДНК
- 67. НЕКОДИРУЮЩИЕ РЕГУЛЯТОРНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ Геном животных и человека - сложнейшая система с многоуровневой системой регуляции. Важную роль
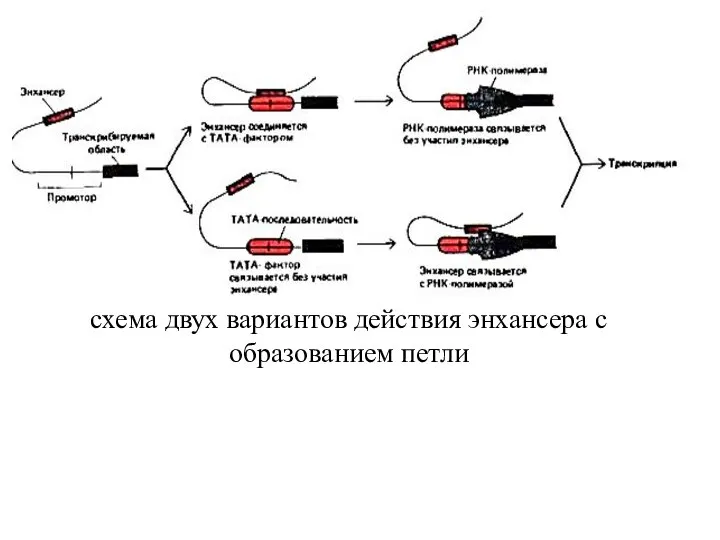
- 68. Энхансеры высших эукариот высших эукариот способны активировать гены на больших расст. (до десятков тыс. пар нуклеотидов).
- 69. схема двух вариантов действия энхансера с образованием петли
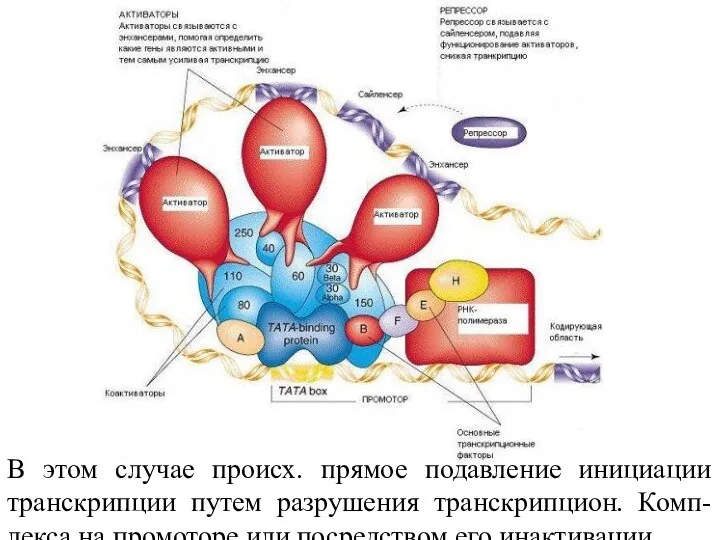
- 70. Активаторы (Activators) - белки, кот. связываются с энхансерами, кот. помогают РНК-полимеразе правильно начать транскрипцию. Репрессоры (Repressor)
- 71. Транскрипцию нужно не только активировать, но и подавлять. Для этого существуют сайленсеры. Сайленсеры репрессируют активность генов,
- 72. В этом случае происх. прямое подавление инициации транскрипции путем разрушения транскрипцион. Комп-лекса на промоторе или посредством
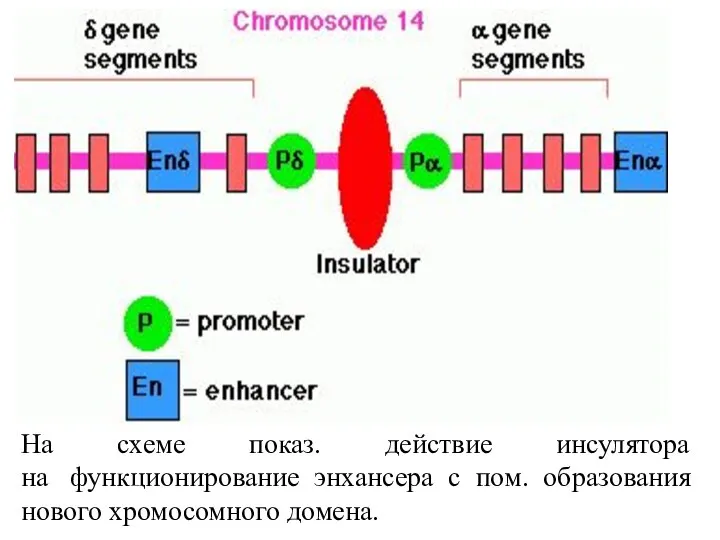
- 73. Инсуляторы (англ. insulate - изолировать) - послед-сти ДНК, особ. регуляторные элементы, кот. облад. способ-стью блокир. сигналы,
- 74. На схеме показ. действие инсулятора на функционирование энхансера с пом. образования нового хромосомного домена.
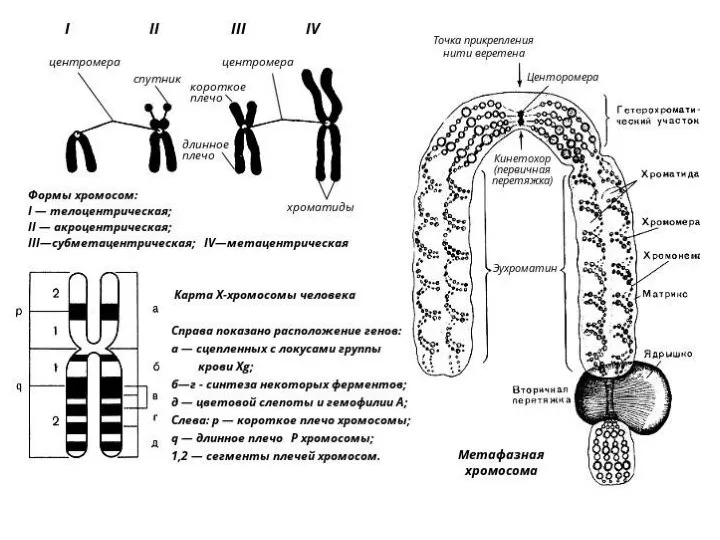
- 75. Внутриядерная архитектура хромосом. Явление трансвекции Клетки тела многоклеточ. эукариот им. Диплоид. (двойной) набор хромосом, складывающийся из
- 78. Скачать презентацию

Слайд 3Истоками молекулярной биологии являются:
органическая химия, занимающаяся изучением химической структуры макромолекул;
биохимия, целью
Истоками молекулярной биологии являются:
органическая химия, занимающаяся изучением химической структуры макромолекул;
биохимия, целью

Слайд 4Исключит-ное внимание привлекают в наст. время 2 осн. класса макромолекул - белки
Исключит-ное внимание привлекают в наст. время 2 осн. класса макромолекул - белки

Слайд 52.Доказательства генетической функции ДНК и РНК.
Структура нуклеотидов.
Принцип комплементарности.
История изучения нуклеиновых кислот
2.Доказательства генетической функции ДНК и РНК.
Структура нуклеотидов.
Принцип комплементарности.
История изучения нуклеиновых кислот

Слайд 61. Английский. бактериолог Ф.Гриффит (1928) продемонстрировал следующий опыт:
способность пневмококков к трансформации.
1. Английский. бактериолог Ф.Гриффит (1928) продемонстрировал следующий опыт:
способность пневмококков к трансформации.

Слайд 7Гриффит заключил, что живые микробы непатогенного штамма в присутствии клеток штамма патогенного
Гриффит заключил, что живые микробы непатогенного штамма в присутствии клеток штамма патогенного

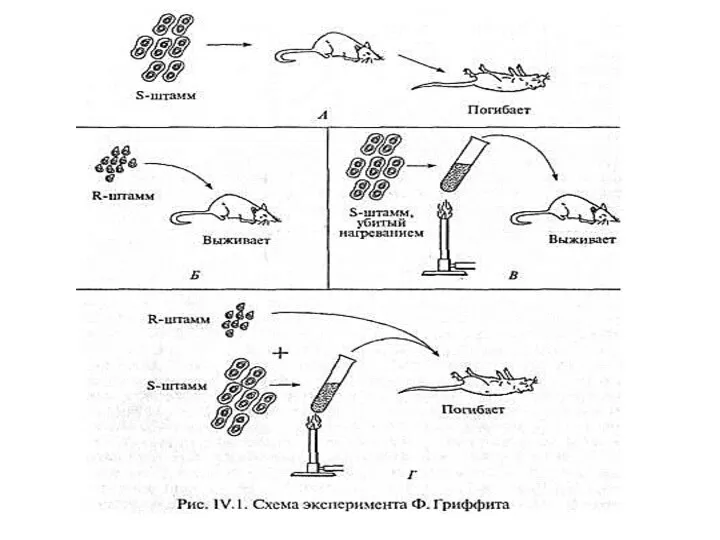
Слайд 9О. Звери, К. Мак-Леод и М. Мак-Карти, в 1944 г. точно определили
О. Звери, К. Мак-Леод и М. Мак-Карти, в 1944 г. точно определили

Слайд 103. В 1950 г. американский биохимик Э.Чаргафф установил важнейшую закономерность химич. строения
3. В 1950 г. американский биохимик Э.Чаргафф установил важнейшую закономерность химич. строения

Слайд 11В 1953 г. Дж. Уотсон и Ф. Крик на основании результатов рентгеноструктурного
В 1953 г. Дж. Уотсон и Ф. Крик на основании результатов рентгеноструктурного

Слайд 12К настоящему времени :
обнаружено, что ДНК мож. повреждаться и мож. восстанавливаться,
молекулы
К настоящему времени :
обнаружено, что ДНК мож. повреждаться и мож. восстанавливаться,
молекулы

Слайд 13Структура нуклеотидов.
Нуклеиновые кислоты – это биологич. высокомолекуляр. полимерн. соединения, мономерами кот. явл. нуклеотиды.
Нуклеотиды –
Структура нуклеотидов.
Нуклеиновые кислоты – это биологич. высокомолекуляр. полимерн. соединения, мономерами кот. явл. нуклеотиды.
Нуклеотиды –

Слайд 14В нуклеиновых кислотах сахар представлен пентозой.
В РНК пентоза является рибозой, а в
В нуклеиновых кислотах сахар представлен пентозой.
В РНК пентоза является рибозой, а в

Слайд 15Изучая нуклеотидный состав ДНК различ. видов орг-мов, сделали след. выводы:
1. нуклеотидный состав ДНК
Изучая нуклеотидный состав ДНК различ. видов орг-мов, сделали след. выводы:
1. нуклеотидный состав ДНК

Слайд 16Первичная структура нуклеиновых кислот
представляет собой порядок чередования нуклеотидов в полинуклеотидной цепи.
Нуклеотиды
Первичная структура нуклеиновых кислот
представляет собой порядок чередования нуклеотидов в полинуклеотидной цепи.
Нуклеотиды
Слайд 17Вторичная структура ДНК
(двойная спираль) была предложена амер. генетиком Д. Уотсоном и англ.
Вторичная структура ДНК
(двойная спираль) была предложена амер. генетиком Д. Уотсоном и англ.
Слайд 18Третичная структура ДНК
Ковалентные связи между атомами в углеводно-фосфатной цепи полинук-леотида им. определённую
Третичная структура ДНК
Ковалентные связи между атомами в углеводно-фосфатной цепи полинук-леотида им. определённую
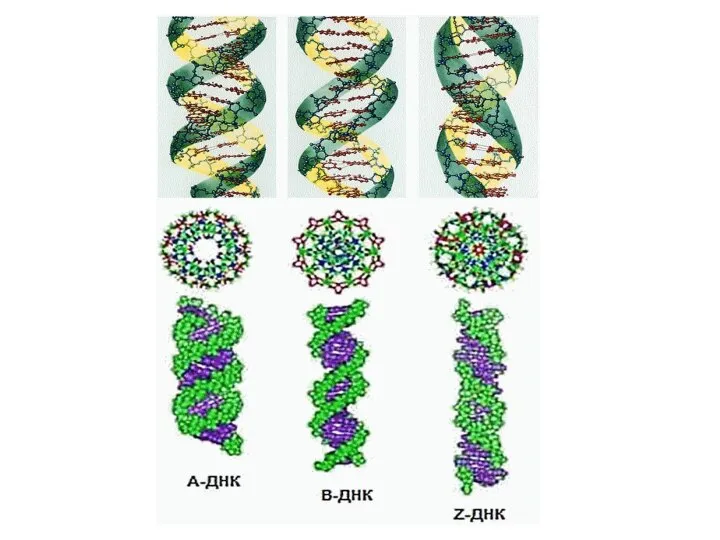
Слайд 19Формы ДНК
А-форма ДНК: двухспиральная ДНК, содержащая прим. 11 остатков на 1 оборот спирали.
Формы ДНК
А-форма ДНК: двухспиральная ДНК, содержащая прим. 11 остатков на 1 оборот спирали.

Слайд 21Комплементарность – это пространственная взаимодополняемость молекул или их частей, приводящая к образованию водородных
Комплементарность – это пространственная взаимодополняемость молекул или их частей, приводящая к образованию водородных

Слайд 22Водородные связи, возникающие между пуринами и пиримидинами, удерживают комплементарные полинуклеотидные цепи в
Водородные связи, возникающие между пуринами и пиримидинами, удерживают комплементарные полинуклеотидные цепи в

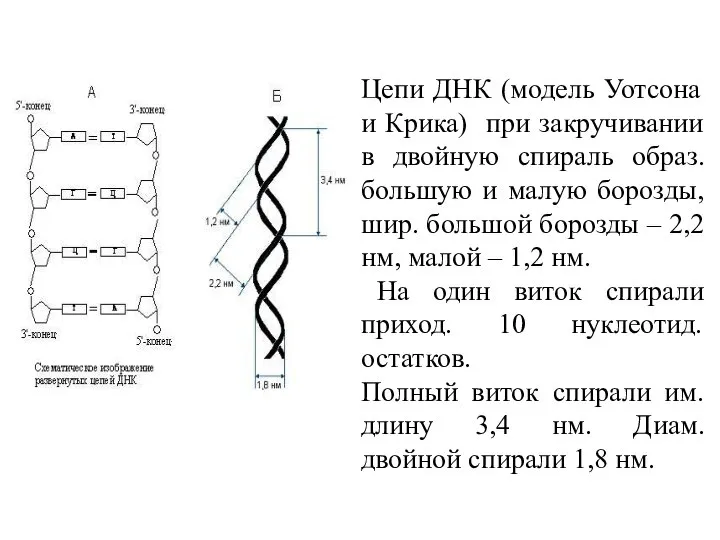
Слайд 23Цепи ДНК (модель Уотсона и Крика) при закручивании в двойную спираль образ.
Цепи ДНК (модель Уотсона и Крика) при закручивании в двойную спираль образ.
Слайд 243. Коды ДНК. Гибкость двойной спирали.
В 1954 г. амер. физик-теоретик Г.А. Гамов
3. Коды ДНК. Гибкость двойной спирали.
В 1954 г. амер. физик-теоретик Г.А. Гамов

Слайд 25Генетич. код не содержит знаков препинания, и кодирующие триплеты следуют один за
Генетич. код не содержит знаков препинания, и кодирующие триплеты следуют один за

Слайд 26Характеристика кода:
1. вырожденность (избыточность) - аминок-ты шифруются неск. триплетами. Вырожденность генетич. кода позволяет при
Характеристика кода:
1. вырожденность (избыточность) - аминок-ты шифруются неск. триплетами. Вырожденность генетич. кода позволяет при

Слайд 27однонаправленность считывания (5′→3′), т.е. считывание информации с мРНК при синтезе белка происходит
однонаправленность считывания (5′→3′), т.е. считывание информации с мРНК при синтезе белка происходит

Слайд 284. Ассоциации ДНК с олигонуклетидами
Олигонуклеотид - короткая однонитевая молекула ДНК и
4. Ассоциации ДНК с олигонуклетидами
Олигонуклеотид - короткая однонитевая молекула ДНК и

Слайд 29Триплексы: специфичность образования
Часто для прикладных задач необходимо, чтобы олигонуклеотид связался с ДНК
Триплексы: специфичность образования
Часто для прикладных задач необходимо, чтобы олигонуклеотид связался с ДНК

Слайд 30Выделяют 3 основных типа ошибок в узнавании олигонуклеотидом двухцепочечной ДНК.:
неправильное спаривание в
Выделяют 3 основных типа ошибок в узнавании олигонуклеотидом двухцепочечной ДНК.:
неправильное спаривание в

Слайд 31Существует неск. подходов к проблеме неспецифичности связывания олигонуклеотидов.
скорость образования специфич.
Существует неск. подходов к проблеме неспецифичности связывания олигонуклеотидов.
скорость образования специфич.

Слайд 32 специфичный комплекс еще достаточно стабилен, а неспецифич. уже нет. Напр. при
специфичный комплекс еще достаточно стабилен, а неспецифич. уже нет. Напр. при

Слайд 33Антисмысловой олигонуклеотид (antisense oligonucleotide) - олигонуклеотид, кот. комплементарен фрагменту мРНК, благодаря чему способен
Антисмысловой олигонуклеотид (antisense oligonucleotide) - олигонуклеотид, кот. комплементарен фрагменту мРНК, благодаря чему способен

Слайд 34Хугстиновское спаривание оснований (Hoogsteen base pairing) (по им. К. Хугстина) - вариация спаривания
Хугстиновское спаривание оснований (Hoogsteen base pairing) (по им. К. Хугстина) - вариация спаривания

Слайд 35В двуспиральных структурах таких пар не обнаружено, однако не исключается возможность образования
В двуспиральных структурах таких пар не обнаружено, однако не исключается возможность образования

Слайд 36ДНК: H-форма
H-форма ДНК – неканонич. структура в гомопурин- гомопиримидиновых участках ДНК.
В
ДНК: H-форма
H-форма ДНК – неканонич. структура в гомопурин- гомопиримидиновых участках ДНК.
В

Слайд 37Кроме того используют методы:
Двумерный электрофорез H-формы ДНК
H-форма ДНК модели с помощью зондирования
Кроме того используют методы:
Двумерный электрофорез H-формы ДНК
H-форма ДНК модели с помощью зондирования

Слайд 38
5. Ассоциация ДНК с белками
Транскрипционные факторы: Принципы классификации
Специфич-сть взаимодействия транскрип-ционных факторов с
5. Ассоциация ДНК с белками
Транскрипционные факторы: Принципы классификации
Специфич-сть взаимодействия транскрип-ционных факторов с

Слайд 39ДНК-связывающих доменов разделены на 4 суперкласса:
Суперкласс 1. Факторы, ДНК-связывающий домен кот. обогащен
ДНК-связывающих доменов разделены на 4 суперкласса:
Суперкласс 1. Факторы, ДНК-связывающий домен кот. обогащен

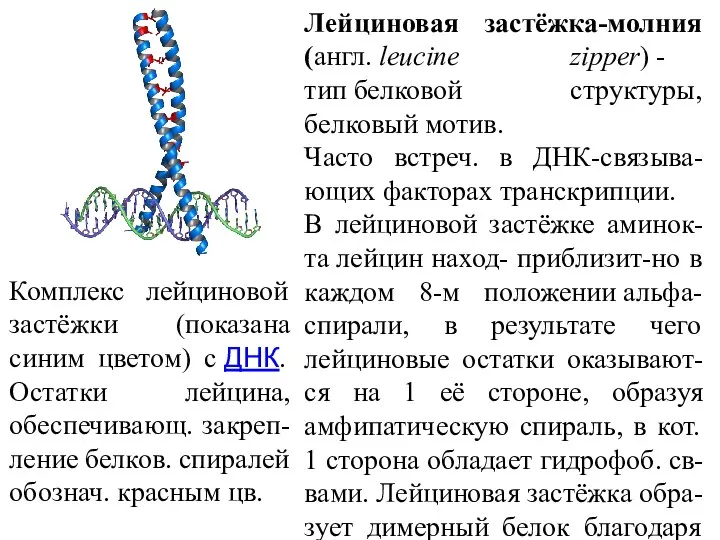
Слайд 40Комплекс лейциновой застёжки (показана синим цветом) с ДНК. Остатки лейцина, обеспечивающ. закреп-ление белков.
Комплекс лейциновой застёжки (показана синим цветом) с ДНК. Остатки лейцина, обеспечивающ. закреп-ление белков.
Слайд 41Цинковый палец (от. англ. цинк. палец) - тип белковой структуры, небольш. белковый мотив, или одним
Цинковый палец (от. англ. цинк. палец) - тип белковой структуры, небольш. белковый мотив, или одним

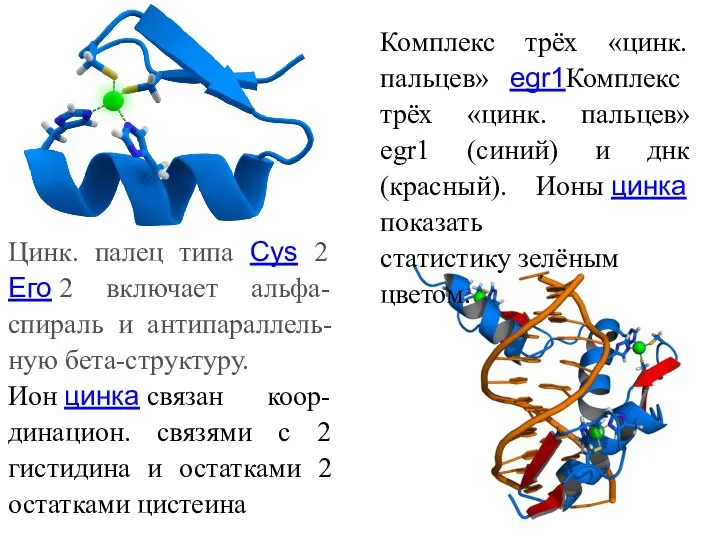
Слайд 42Цинк. палец типа Cys 2 Его 2 включает альфа-спираль и антипараллель-ную бета-структуру.
Ион цинка связан коор-динацион.
Цинк. палец типа Cys 2 Его 2 включает альфа-спираль и антипараллель-ную бета-структуру.
Ион цинка связан коор-динацион.
Слайд 43Факторы транскрипции с доменом "спираль-поворот-спираль»
Тип пептидных доменов, специфически распознающих регуляторные послед-сти на
Факторы транскрипции с доменом "спираль-поворот-спираль»
Тип пептидных доменов, специфически распознающих регуляторные послед-сти на

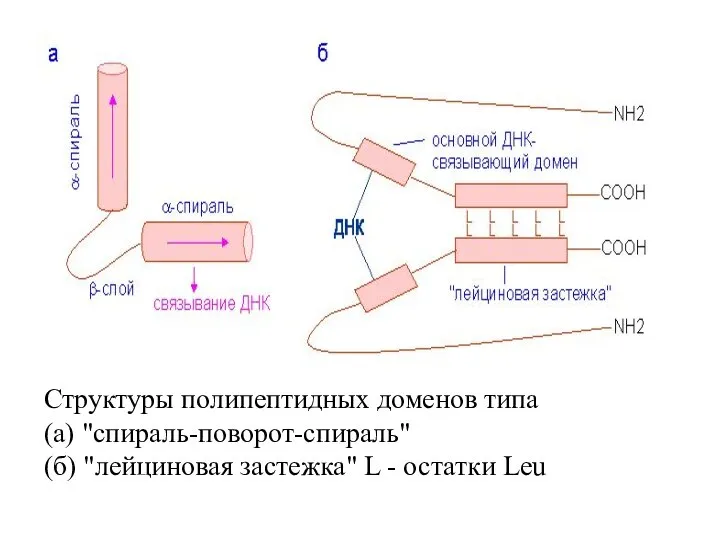
Слайд 44Структуры полипептидных доменов типа
(а) "спираль-поворот-спираль"
(б) "лейциновая застежка" L - остатки Leu
Структуры полипептидных доменов типа
(а) "спираль-поворот-спираль"
(б) "лейциновая застежка" L - остатки Leu
Слайд 45Транскрипционные факторы: краткое определение
Факторы транскрипции (transcription factors) - белки или белковые комплексы,
Транскрипционные факторы: краткое определение Факторы транскрипции (transcription factors) - белки или белковые комплексы,

Слайд 46По функциональному признаку различают
три класса транскрипционных факторов:
а) основные, обеспечивающие нерегулируемый
По функциональному признаку различают
три класса транскрипционных факторов:
а) основные, обеспечивающие нерегулируемый

Слайд 47Транскрипционный фактор – это белок, кот. после его перемещения в ядро клетки
Транскрипционный фактор – это белок, кот. после его перемещения в ядро клетки

Слайд 48Антигены: силы взаимодействия с антителами (электростатическое притяжение)
В основе взаимодействия антиген - антитело лежат те же законы
Антигены: силы взаимодействия с антителами (электростатическое притяжение)
В основе взаимодействия антиген - антитело лежат те же законы

Слайд 493. Гидрофобные взаимодействия между неполярными гидрофобными группами, обеспечивающие по некоторым оценкам до
3. Гидрофобные взаимодействия между неполярными гидрофобными группами, обеспечивающие по некоторым оценкам до

Слайд 50Организация хроматина эукариот.
Вопрос о структурной организации хроматина в интерфазных ядрах далек
Организация хроматина эукариот.
Вопрос о структурной организации хроматина в интерфазных ядрах далек

Слайд 51ХРОМАТИН, нуклеопротеид клеточ. ядра, составл. основу хромосом.
В состав X. входят: ДНК
ХРОМАТИН, нуклеопротеид клеточ. ядра, составл. основу хромосом.
В состав X. входят: ДНК

Слайд 52Согласно пространств. модели А. Клуга кор-частица выглядит как плоский диск диам. 11
Согласно пространств. модели А. Клуга кор-частица выглядит как плоский диск диам. 11

Слайд 53Гистоны составляют больш. осн. белков хроматина и находятся примерно в том же
Гистоны составляют больш. осн. белков хроматина и находятся примерно в том же

Слайд 54Гистоны первых четырех классов им. значит-ное кол-во как кислых, так и основных
Гистоны первых четырех классов им. значит-ное кол-во как кислых, так и основных

Слайд 55Пятый класс представлен гистонами, оч. богатыми лизином; он сост. из неск. достаточно
Пятый класс представлен гистонами, оч. богатыми лизином; он сост. из неск. достаточно

Слайд 56негистоны - это все др. белки хроматина.
Предполагается поэтому, что они облад.
негистоны - это все др. белки хроматина.
Предполагается поэтому, что они облад.

Слайд 57Упаковка генетич. материала достигается путем спирализации (конденсации)
Первый уровень упаковки ДНК - нуклеосомный.
Упаковка генетич. материала достигается путем спирализации (конденсации)
Первый уровень упаковки ДНК - нуклеосомный.

Слайд 58Третий уровень упаковки - хроматидный (петлевой). Супернуклеосомная нить спирализуется с образованием петель
Третий уровень упаковки - хроматидный (петлевой). Супернуклеосомная нить спирализуется с образованием петель

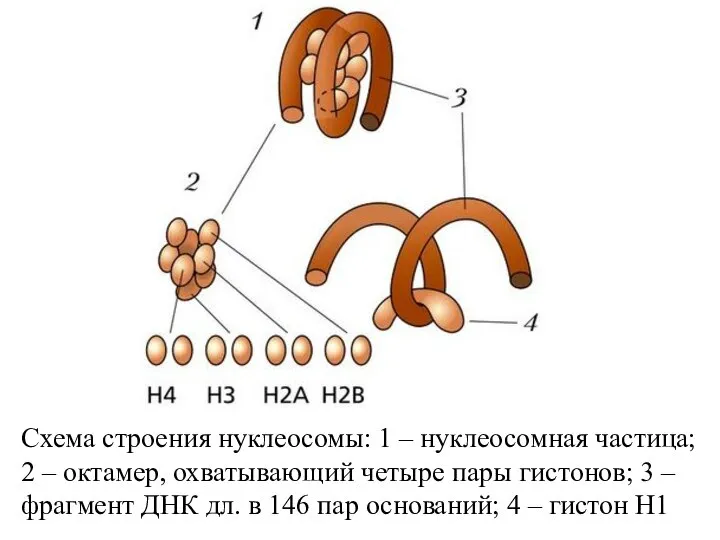
Слайд 59Схема строения нуклеосомы: 1 – нуклеосомная частица; 2 – октамер, охватывающий четыре
Схема строения нуклеосомы: 1 – нуклеосомная частица; 2 – октамер, охватывающий четыре
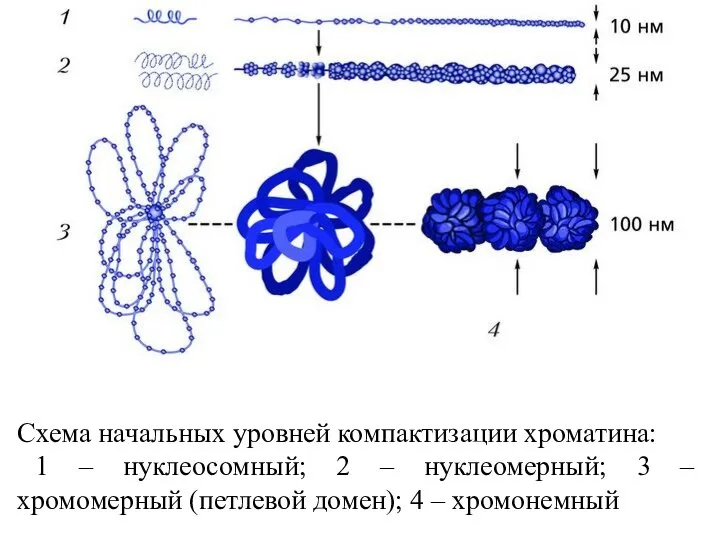
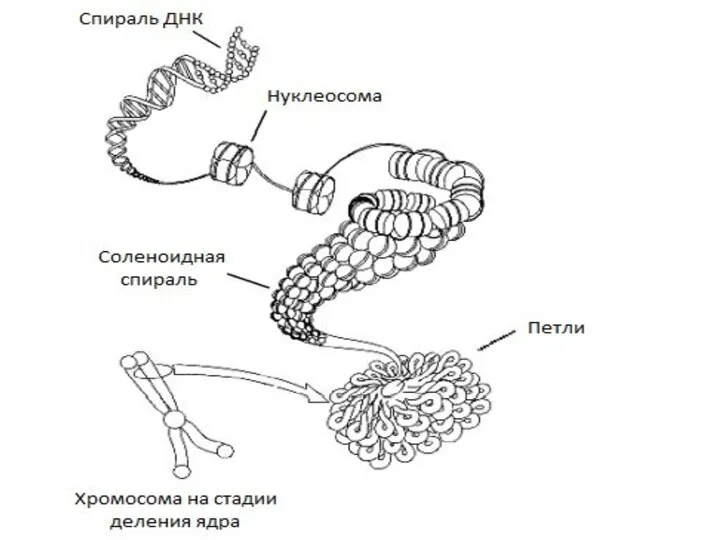
Слайд 60Схема начальных уровней компактизации хроматина:
1 – нуклеосомный; 2 – нуклеомерный; 3
Схема начальных уровней компактизации хроматина:
1 – нуклеосомный; 2 – нуклеомерный; 3
Слайд 62Сборка нуклеосом
1 способ. обусловлен способностью тетрамера Н32Н42 организовывать ДНК в частицы, которые
Сборка нуклеосом
1 способ. обусловлен способностью тетрамера Н32Н42 организовывать ДНК в частицы, которые

Слайд 63Антитела, полученные против этого белка, реагируют с белками нуклеоплазмы многих эукариот. След-но,
Антитела, полученные против этого белка, реагируют с белками нуклеоплазмы многих эукариот. След-но,

Слайд 64Фейзинг.
Терм. "фазирование" обознач. неслучайное располож. нуклеосом относит-но конкретной послед-сти нуклеотидов ДНК в
Фейзинг.
Терм. "фазирование" обознач. неслучайное располож. нуклеосом относит-но конкретной послед-сти нуклеотидов ДНК в

Слайд 65Во время транскрипции часть нуклеосомных белков остается связанной с ДНК.
Нуклеосомы как
Во время транскрипции часть нуклеосомных белков остается связанной с ДНК.
Нуклеосомы как

Слайд 66Предлагаются 2 варианта изменения структуры нуклеосом при синтезе РНК:
нуклеосома «расщепляется» на 2
Предлагаются 2 варианта изменения структуры нуклеосом при синтезе РНК:
нуклеосома «расщепляется» на 2

Слайд 67НЕКОДИРУЮЩИЕ РЕГУЛЯТОРНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ
Геном животных и человека - сложнейшая система с многоуровневой системой
НЕКОДИРУЮЩИЕ РЕГУЛЯТОРНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ
Геном животных и человека - сложнейшая система с многоуровневой системой

Слайд 68Энхансеры высших эукариот высших эукариот способны активировать гены на больших расст. (до десятков тыс. пар нуклеотидов).
Энхансеры высших эукариот высших эукариот способны активировать гены на больших расст. (до десятков тыс. пар нуклеотидов).

Слайд 69схема двух вариантов действия энхансера с образованием петли
схема двух вариантов действия энхансера с образованием петли
Слайд 70Активаторы (Activators) - белки, кот. связываются с энхансерами, кот. помогают РНК-полимеразе правильно
Активаторы (Activators) - белки, кот. связываются с энхансерами, кот. помогают РНК-полимеразе правильно

Слайд 71Транскрипцию нужно не только активировать, но и подавлять.
Для этого существуют сайленсеры.
Транскрипцию нужно не только активировать, но и подавлять.
Для этого существуют сайленсеры.

Слайд 72В этом случае происх. прямое подавление инициации транскрипции путем разрушения транскрипцион. Комп-лекса
В этом случае происх. прямое подавление инициации транскрипции путем разрушения транскрипцион. Комп-лекса
Слайд 73Инсуляторы (англ. insulate - изолировать) - послед-сти ДНК, особ. регуляторные элементы, кот. облад.
Инсуляторы (англ. insulate - изолировать) - послед-сти ДНК, особ. регуляторные элементы, кот. облад.

Слайд 74На схеме показ. действие инсулятора на функционирование энхансера с пом. образования нового
На схеме показ. действие инсулятора на функционирование энхансера с пом. образования нового
Слайд 75Внутриядерная архитектура хромосом. Явление трансвекции
Клетки тела многоклеточ. эукариот им. Диплоид. (двойной) набор
Внутриядерная архитектура хромосом. Явление трансвекции
Клетки тела многоклеточ. эукариот им. Диплоид. (двойной) набор

 Плесневые грибы и дрожжи
Плесневые грибы и дрожжи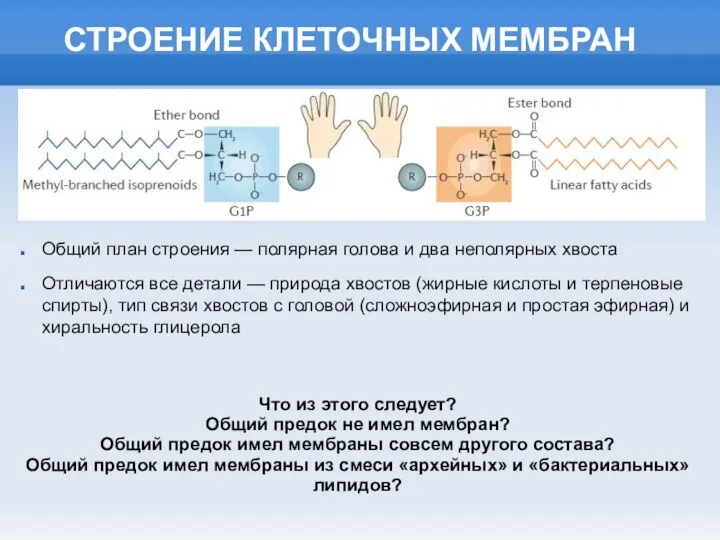 Строение клеточных мембран
Строение клеточных мембран Популяция – единица вида и эволюции
Популяция – единица вида и эволюции Биоэлемент фосфор для животных
Биоэлемент фосфор для животных Селекционеры в годы Великой Отечественной войны
Селекционеры в годы Великой Отечественной войны Способы передвижения животных. Полости тела
Способы передвижения животных. Полости тела Координация и регуляция
Координация и регуляция Птицы
Птицы Дисахариды. Физические свойства
Дисахариды. Физические свойства Промежуточный мозг (diencephalon)
Промежуточный мозг (diencephalon) Рокарий
Рокарий Животные Нижегородской области
Животные Нижегородской области Свиристель в Анжерке. Исследовательская работа
Свиристель в Анжерке. Исследовательская работа Взаимодействие генов
Взаимодействие генов Как размножаются растения
Как размножаются растения Мышцы кисти
Мышцы кисти Обмен углеводов. Гликолиз. Пентозофосфатный путь
Обмен углеводов. Гликолиз. Пентозофосфатный путь Понятие о дисциплине Дендрология. Жизненные формы. Жизненный цикл древесных растений
Понятие о дисциплине Дендрология. Жизненные формы. Жизненный цикл древесных растений Класс Птицы
Класс Птицы Органические вещества клетки. Биологическая роль веществ в клетке (на примере углеводов)
Органические вещества клетки. Биологическая роль веществ в клетке (на примере углеводов) Картоптың қара күйесі. Angiosorus solani. T
Картоптың қара күйесі. Angiosorus solani. T лекция 2 Физиология
лекция 2 Физиология Лес – наше богатство. Занятие №1
Лес – наше богатство. Занятие №1 Функциональная анатомия органов дыхания
Функциональная анатомия органов дыхания Голосовой аппарат человека. Занятие 6
Голосовой аппарат человека. Занятие 6 חיות בעולם
חיות בעולם «ПЕРЕЛЁТНЫЕ ПТИЦЫ»
«ПЕРЕЛЁТНЫЕ ПТИЦЫ» Терморегуляция организма. Закаливание
Терморегуляция организма. Закаливание