- Нейрофизиология слуховой системы

Содержание
- 2. Мир, окружающий человека, полон звуков. Человек слышит не все звуки этого мира, а только те, которые
- 3. Среднее ухо отделено от наружного барабанной перепонкой, которая имеет вид конуса с вершиной, обращенной в барабанную
- 5. Строение среднего уха (схема). А —передача звуковых колебаний через среднее ухо к внутреннему уху; Б —улиточный
- 6. Внутреннее ухо включает два рецепторных аппарата: вестибулярный и слуховой. Опишем слуховой орган. Костный канал улитки разделен
- 8. Средняя лестница, заполненная эндолимфой, заряжена положительно (до +80 мВ) относительно двух других лестниц. Разность потенциала (эндокохлеарный
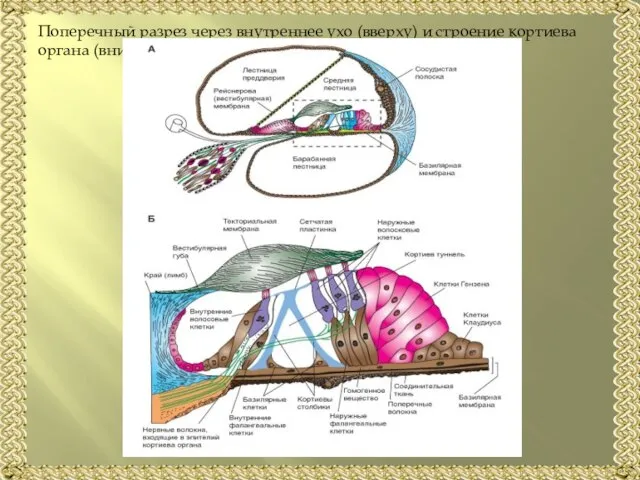
- 9. Поперечный разрез через внутреннее ухо (вверху) и строение кортиева органа (внизу):
- 10. Венгерский ученый Г. Бекеши (1951) предложил «теорию бегущей волны», позволяющую понять, как звуковая волна определенной частоты
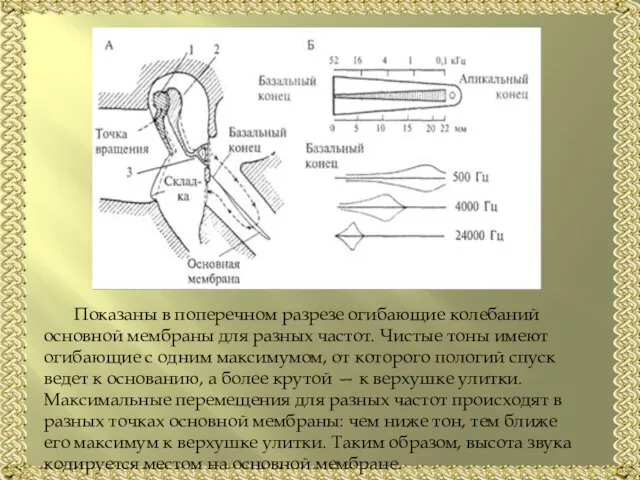
- 11. Показаны в поперечном разрезе огибающие колебаний основной мембраны для разных частот. Чистые тоны имеют огибающие с
- 12. Физиология путей и центров слуховой системы. Нейроны 1-го порядка (биполярные нейроны) находятся в спиральном ганглии, который
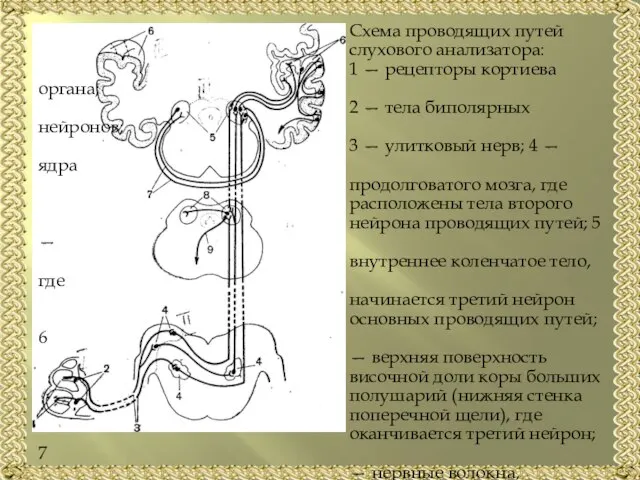
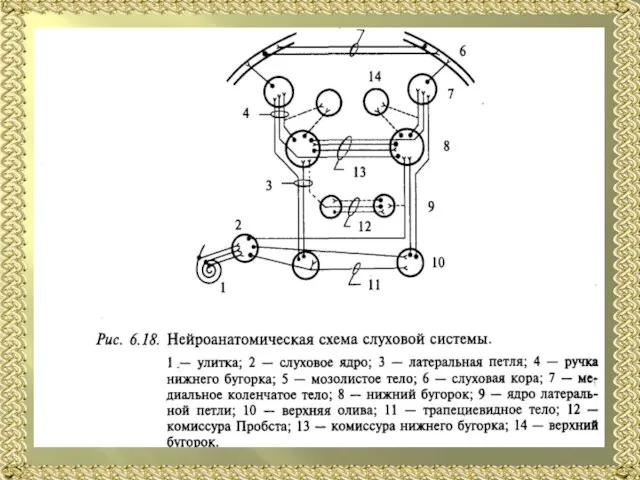
- 13. Схема проводящих путей слухового анализатора: 1 — рецепторы кортиева органа; 2 — тела биполярных нейронов; 3
- 14. Нейроны 2-го порядка образуют в продолговатом мозге комплекс слуховых ядер. Этот комплекс делят на дорсальный и
- 16. Следующий уровень слуховой системы находится на уровне моста и представлен ядрами верхней оливы (медиальным и латеральным)
- 17. Нижнее двухолмие, расположенное на дорсальной поверхности среднего мозга, является важнейшим центром анализа звуковых сигналов. На этом
- 19. Медиальное коленчатое тело является таламическим центром слуховой системы. В нем различают крупноклеточную и мелкоклеточную (основную) части.
- 20. Слуховая кора представляет высший центр слуховой системы и располагается в височной доле. У человека в ее
- 21. Частотно-пороговые характеристики нейронов слуховой системы. Частотную избирательность нейронов слуховой системы описывают частотно-пороговой кривой (ЧПК), отражающей зависимость
- 24. Скачать презентацию
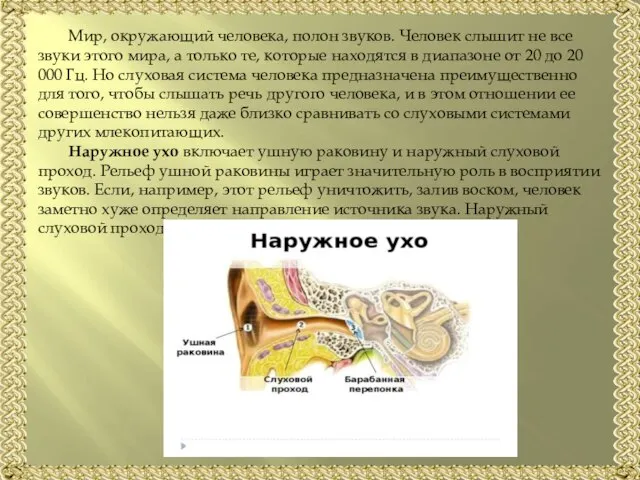
Слайд 3Среднее ухо отделено от наружного барабанной перепонкой, которая имеет вид конуса с
Среднее ухо отделено от наружного барабанной перепонкой, которая имеет вид конуса с

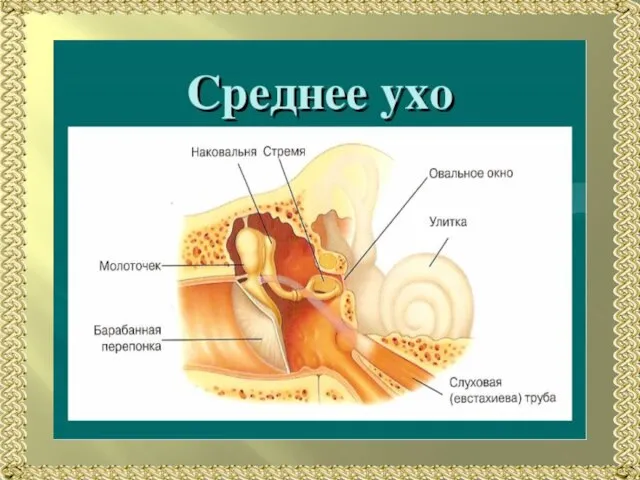
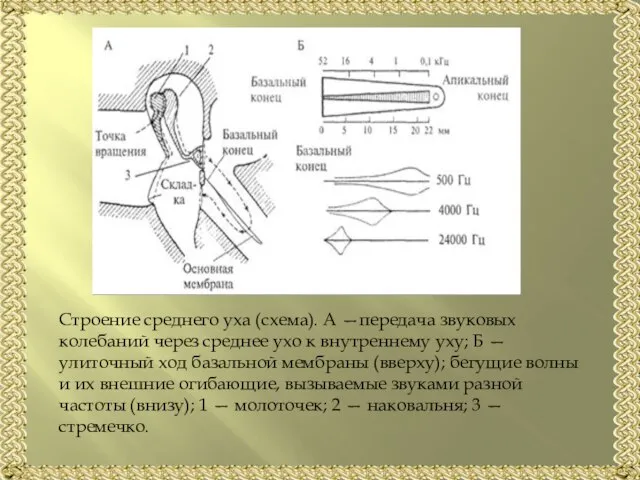
Слайд 5Строение среднего уха (схема). А —передача звуковых колебаний через среднее ухо к
Строение среднего уха (схема). А —передача звуковых колебаний через среднее ухо к
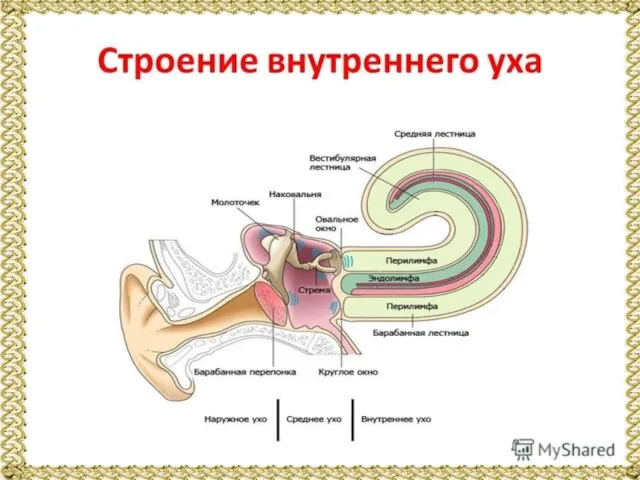
Слайд 6Внутреннее ухо включает два рецепторных аппарата: вестибулярный и слуховой. Опишем слуховой орган.
Внутреннее ухо включает два рецепторных аппарата: вестибулярный и слуховой. Опишем слуховой орган.

Слайд 8Средняя лестница, заполненная эндолимфой, заряжена положительно (до +80 мВ) относительно двух других
Средняя лестница, заполненная эндолимфой, заряжена положительно (до +80 мВ) относительно двух других

Слайд 9Поперечный разрез через внутреннее ухо (вверху) и строение кортиева органа (внизу):
Поперечный разрез через внутреннее ухо (вверху) и строение кортиева органа (внизу):
Слайд 10Венгерский ученый Г. Бекеши (1951) предложил «теорию бегущей волны», позволяющую понять, как
Венгерский ученый Г. Бекеши (1951) предложил «теорию бегущей волны», позволяющую понять, как

Слайд 11Показаны в поперечном разрезе огибающие колебаний основной мембраны для разных частот. Чистые
Показаны в поперечном разрезе огибающие колебаний основной мембраны для разных частот. Чистые
Слайд 12Физиология путей и центров слуховой системы. Нейроны 1-го порядка (биполярные нейроны) находятся
Физиология путей и центров слуховой системы. Нейроны 1-го порядка (биполярные нейроны) находятся

Слайд 13 Схема проводящих путей
слухового анализатора:
1 — рецепторы кортиева органа;
Схема проводящих путей
слухового анализатора:
1 — рецепторы кортиева органа;
Слайд 14Нейроны 2-го порядка образуют в продолговатом мозге комплекс слуховых ядер. Этот комплекс
Нейроны 2-го порядка образуют в продолговатом мозге комплекс слуховых ядер. Этот комплекс

Слайд 16Следующий уровень слуховой системы находится на уровне моста и представлен ядрами верхней
Следующий уровень слуховой системы находится на уровне моста и представлен ядрами верхней

Слайд 17Нижнее двухолмие, расположенное на дорсальной поверхности среднего мозга, является важнейшим центром анализа
Нижнее двухолмие, расположенное на дорсальной поверхности среднего мозга, является важнейшим центром анализа

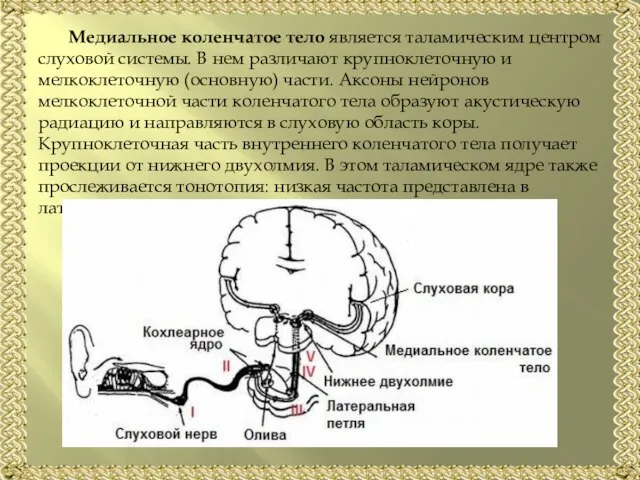
Слайд 19Медиальное коленчатое тело является таламическим центром слуховой системы. В нем различают крупноклеточную
Медиальное коленчатое тело является таламическим центром слуховой системы. В нем различают крупноклеточную
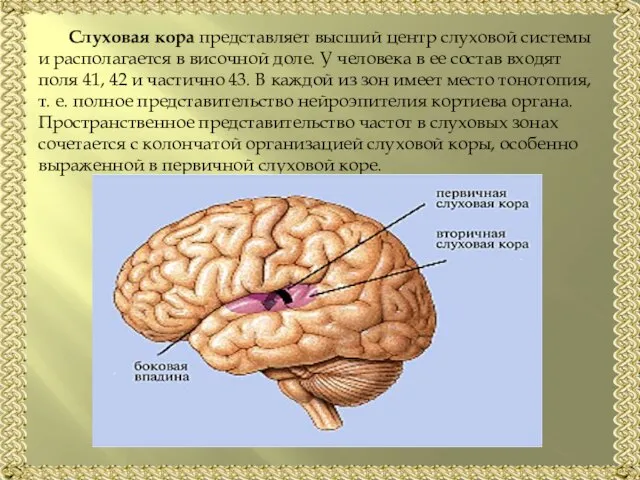
Слайд 20Слуховая кора представляет высший центр слуховой системы и располагается в височной доле.
Слуховая кора представляет высший центр слуховой системы и располагается в височной доле.
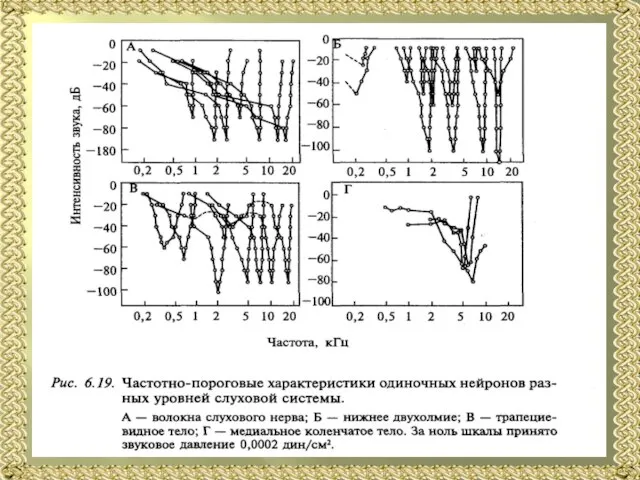
Слайд 21Частотно-пороговые характеристики нейронов слуховой системы. Частотную избирательность нейронов слуховой системы описывают частотно-пороговой
Частотно-пороговые характеристики нейронов слуховой системы. Частотную избирательность нейронов слуховой системы описывают частотно-пороговой

 3Kletochnaya_teoria
3Kletochnaya_teoria ЦНС - центральная нервная система
ЦНС - центральная нервная система Строение и функции цитоплазматической мембраны
Строение и функции цитоплазматической мембраны Малообъемная гидрооника (Сириус)
Малообъемная гидрооника (Сириус) Животные Саратовкой области (Красная книга)
Животные Саратовкой области (Красная книга) Загадочная плесень
Загадочная плесень Адаптация к охлаждающему воздействию
Адаптация к охлаждающему воздействию Сибирский багульник
Сибирский багульник Секреторная мембранная система. Аппарат Гольджи. Лекция 9
Секреторная мембранная система. Аппарат Гольджи. Лекция 9 Физиология растений
Физиология растений Методы изучения наследственности человека
Методы изучения наследственности человека Бактериофаги и бактериофагия
Бактериофаги и бактериофагия Тип Членистоногие. Класс Насекомые (Insécta). 7 класс
Тип Членистоногие. Класс Насекомые (Insécta). 7 класс Кожа. Структура кожи
Кожа. Структура кожи Які зміни відбуваються в неживій природі
Які зміни відбуваються в неживій природі prezentatsiya-k-uroku-virusnye-zabolevaniya
prezentatsiya-k-uroku-virusnye-zabolevaniya Развитие животного мира на Земле
Развитие животного мира на Земле Каталитикалық РНК
Каталитикалық РНК Анатомия, физиология, патология органов речи, слуха и зрения
Анатомия, физиология, патология органов речи, слуха и зрения Отдел Zygomycota. Класс Zygomycetes
Отдел Zygomycota. Класс Zygomycetes Класс млекопитающие. Отряд мозоленогие, отряд парнокопытные
Класс млекопитающие. Отряд мозоленогие, отряд парнокопытные Методическое пособие к уроку: Разнообразие биологических систем и процессов
Методическое пособие к уроку: Разнообразие биологических систем и процессов Структурная организация клеток прокариот и эукариот. Лекция 1
Структурная организация клеток прокариот и эукариот. Лекция 1 Класс Ракообразные
Класс Ракообразные Leishmania
Leishmania Отряд простейших. Тип саркомостигофоры
Отряд простейших. Тип саркомостигофоры Семейство Пасленовые. Класс Двудольные
Семейство Пасленовые. Класс Двудольные Презентация на тему Скелет
Презентация на тему Скелет