- Нуклеиновые кислоты

Содержание
- 2. Нуклеиновые кислоты являются высокомолекулярными органическими соединениями. Существует два типа НК: рибонуклеиновые кислоты (РНК) и дезоксирибонуклеиновые кислоты
- 3. Следует иметь ввиду, что одни НК не могут быть достаточно стабильными хранителями генетической информации, и тем
- 4. Молекулы НК являются самыми большими биологическими полимерами. Так молекулярная масса РНК может достигать 10 – 14
- 5. Такая молекула крайне нестабильна и в отсутствии стабилизирующих её белков легко разламывается. При всех своих гигантских
- 6. При полном гидролизе НК в гидролизате могут быть обнаружены соединения трёх типов: фосфорная кислота; углевод (
- 8. На долю аденина (А), гуанина (Г), цитозина (Ц) и тимина (Т) приходится 99 -98% от общего
- 9. Минорные азотистые основания Минорные азотистые основания –это редко встречающиеся азотистые основания. Они составляют 1 -2% в
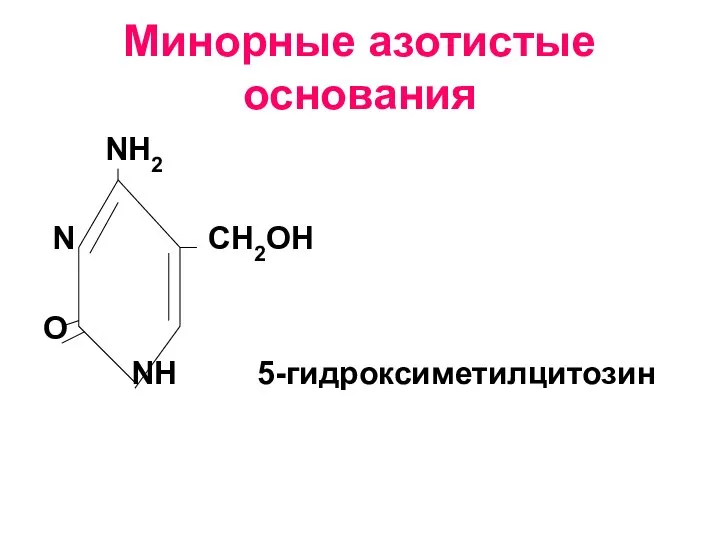
- 10. Минорные азотистые основания NH2 N СН2ОН O NH 5-гидроксиметилцитозин
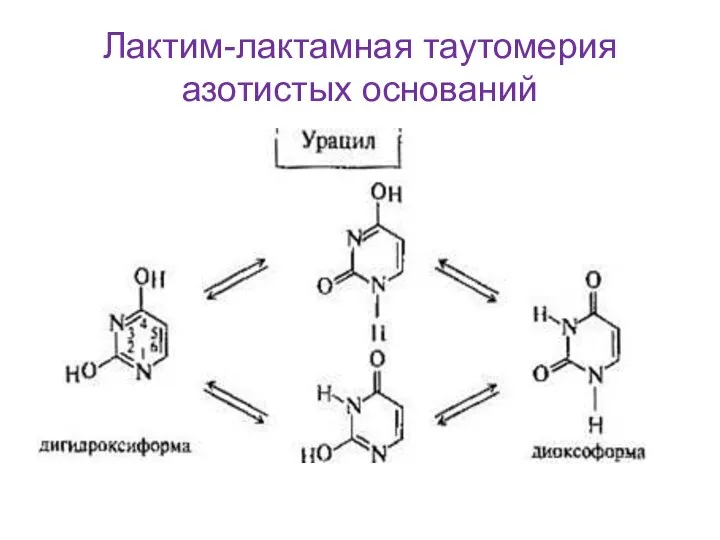
- 11. Азотистые основания пиримидинового или пуринового ряда, имеющие в своём составе кислород, могут существовать в двух формах:
- 12. Лактим-лактамная таутомерия азотистых оснований
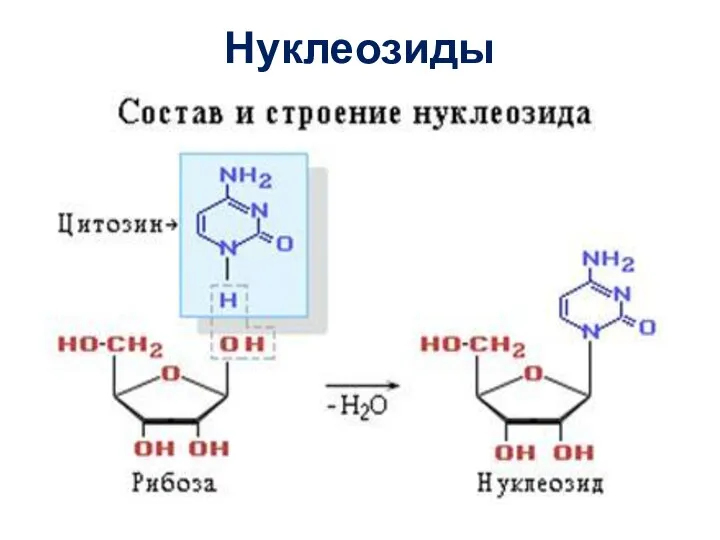
- 13. Нуклеозиды
- 14. Нуклеозиды
- 15. Нуклеотиды Если к остатку моносахарида в нуклеозиде присоединён остаток фосфорной кислоты, то такая структура получила название
- 16. Нуклеотиды
- 18. Нуклеотиды Мононуклеотиды в клетках могут: входить в состав НК; функционировать самостоятельно, выступая или в качестве коферментов,
- 19. Нуклеотиды Энергия временно аккумулируется в пирофосфатных связях молекулы АТФ.
- 20. Примером нуклеотида, выполняющего в клетках роль регулятора обменных процессов, является циклическая аденозинмонофосфорная кислота или цАМФ. Циклические
- 21. цАМФ
- 22. цАМФ В структуре цАМФ один остаток фосфорной кислоты соединён одновременно и с третьим(С3), и с пятым
- 23. НАД Н2/НАД+
- 24. Первичная структура НК В процессе образования полинуклеотида 2 мононуклеотида соединяются друг с другом за счёт взаимодействия
- 25. Первичная структура НК
- 26. Первичная структура НК После выделения молекулы воды 2 мононуклеотида оказываются соединёнными сложноэфирной связью между остатком фосфорной
- 27. Первичная структура НК Образовавшийся нуклеотид имеет на одном конце остаток фосфорной кислоты, а на другом -
- 28. Первичная структура НК Количество мономерных звеньев в полимерной цепи может достигать 108 -109 единиц. Мононуклеотиды связаны
- 29. Первичная структура НК Полинуклеотидная цепь имеет начало и окончание. Началом считается тот конец полинуклеотида, на котором
- 30. Первичная структура НК Окончанием полинуклеотида считается его конец, содержащий или свободный гидроксил у С3- пентозы или
- 31. Первичная структура НК Полинуклеотидную цепь принято изображать в виде последовательности сокращённых названий азотистых оснований, входящих в
- 32. Изучение нуклеотидного состава ДНК показало, что: - количество пиримидинов в молекуле эквивалентно количеству пуринов ( Т+Ц=А+Г
- 33. Тем не менее ДНК каждого отдельного вида животного, растения или микроорганизма обладает характерным для данного вида
- 34. Вторичная структура ДНК Согласно современным представлениям молекула ДНК представляет собой не одиночную, а двойную полинуклеотидную цепь,
- 35. Вторичная структура ДНК
- 36. Вторичная структура ДНК Спираль имеет правую закрутку и на каждый виток спирали приходится 10 пар нуклеотидов,
- 37. Вторичная структура ДНК Азотистые основания одной цепи взаимодействуют с азотистыми основаниями другой цепи за счёт образования
- 38. Вторичная структура ДНК Долгое время считалось, что стабильность двойной спирали ДНК обусловлена только водородными связями между
- 39. Стэкинг –взаимодействие - -суть его состоит в том, что плоскости азотистых оснований молекулы ДНК, перпендикулярные длинной
- 40. Стэкинг –взаимодействие В результата вся молекула оказывается «прошитой» по длине особым типом связей, которые получили название
- 41. Вторичная структура ДНК Стэкинг -взаимодействие
- 42. Вторичная структура ДНК Поскольку все азотистые основания одной полинуклеотидной цепи ДНК комплементарны азотистым основаниям её цепи,
- 43. Вторичная структура ДНК Регулярная пространственная структура, возникающая в результате взаимодействия двух антипараллельных комплементарных полинуклеотидных цепей и
- 44. РНК В отличие от ДНК молекулы РНК не имеют каких-либо выраженных закономерностей в отношении количественного состава
- 45. РНК В качестве элементов вторичной структуры РНК обычно рассматривают так называемые «шпильки», представляющие собой участки одной
- 46. РНК Полной комплементарности между восходящим и нисходящим участками в полинуклеотидной цепи в такой шпильке нет и
- 47. РНК В то время как состав и структура ДНК постоянны для клеток данного организма, молекулу РНК
- 48. РНК Рибосомальная РНК – входит в состав рибосом, клеточных органелл, на которых происходит сборка полипептидных цепей
- 50. Скачать презентацию
Слайд 2Нуклеиновые кислоты являются высокомолекулярными органическими соединениями. Существует два типа НК: рибонуклеиновые кислоты
Нуклеиновые кислоты являются высокомолекулярными органическими соединениями. Существует два типа НК: рибонуклеиновые кислоты

Слайд 3Следует иметь ввиду, что одни НК не могут быть достаточно стабильными хранителями
Следует иметь ввиду, что одни НК не могут быть достаточно стабильными хранителями

Слайд 4Молекулы НК являются самыми большими биологическими полимерами. Так молекулярная масса РНК может
Молекулы НК являются самыми большими биологическими полимерами. Так молекулярная масса РНК может

Слайд 5Такая молекула крайне нестабильна и в отсутствии стабилизирующих её белков легко разламывается.
При
Такая молекула крайне нестабильна и в отсутствии стабилизирующих её белков легко разламывается.
При

Слайд 6При полном гидролизе НК в гидролизате могут быть обнаружены соединения трёх типов:
При полном гидролизе НК в гидролизате могут быть обнаружены соединения трёх типов:

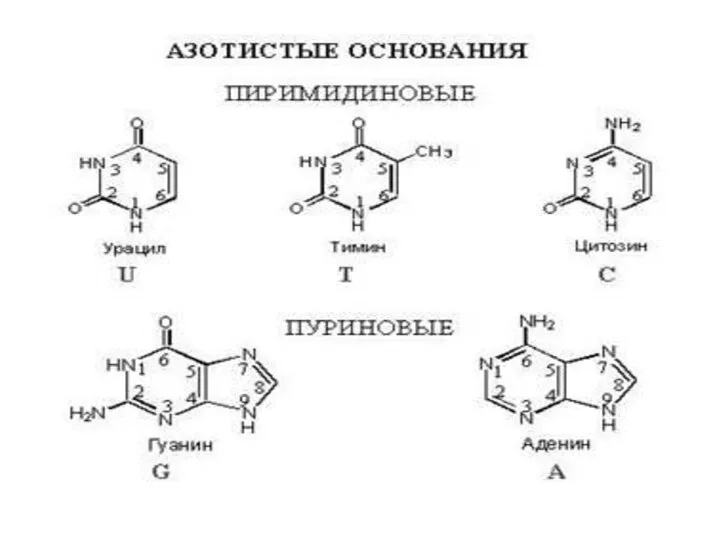
Слайд 8На долю аденина (А), гуанина (Г), цитозина (Ц) и тимина (Т)
приходится
На долю аденина (А), гуанина (Г), цитозина (Ц) и тимина (Т)
приходится

Слайд 9Минорные азотистые основания
Минорные азотистые основания –это редко встречающиеся азотистые основания. Они составляют
Минорные азотистые основания
Минорные азотистые основания –это редко встречающиеся азотистые основания. Они составляют

Слайд 10Минорные азотистые основания
NH2
N СН2ОН
O
NH 5-гидроксиметилцитозин
Минорные азотистые основания
NH2
N СН2ОН
O
NH 5-гидроксиметилцитозин
Слайд 11Азотистые основания пиримидинового или пуринового ряда, имеющие в своём составе кислород, могут
Азотистые основания пиримидинового или пуринового ряда, имеющие в своём составе кислород, могут

Слайд 12Лактим-лактамная таутомерия азотистых оснований
Лактим-лактамная таутомерия азотистых оснований
Слайд 13Нуклеозиды
Нуклеозиды
Слайд 14Нуклеозиды
Нуклеозиды
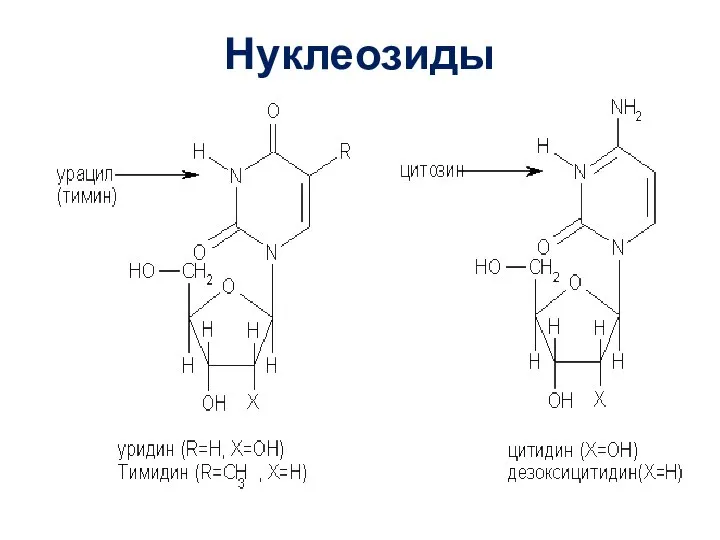
Слайд 15Нуклеотиды
Если к остатку моносахарида в нуклеозиде присоединён остаток фосфорной кислоты, то такая
Нуклеотиды
Если к остатку моносахарида в нуклеозиде присоединён остаток фосфорной кислоты, то такая

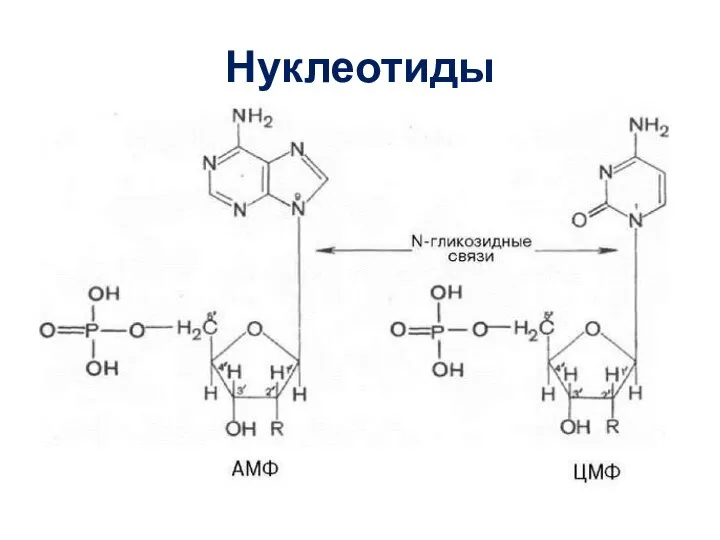
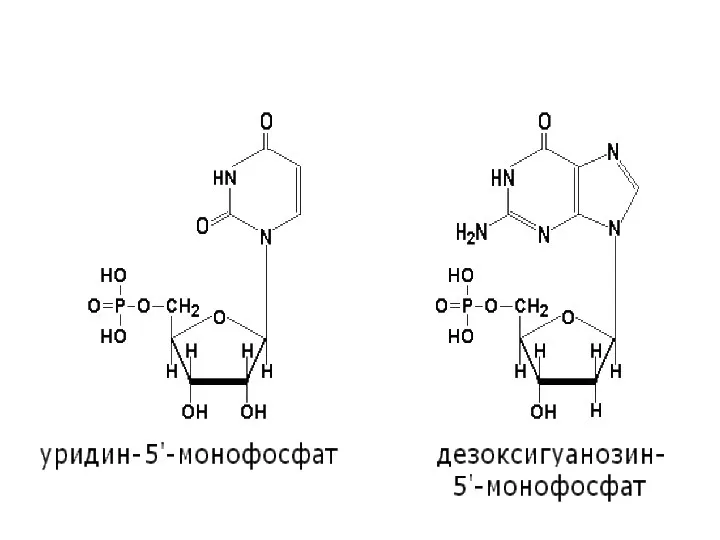
Слайд 16Нуклеотиды
Нуклеотиды
Слайд 18Нуклеотиды
Мононуклеотиды в клетках могут:
входить в состав НК;
функционировать самостоятельно, выступая или в
Нуклеотиды
Мононуклеотиды в клетках могут:
входить в состав НК;
функционировать самостоятельно, выступая или в

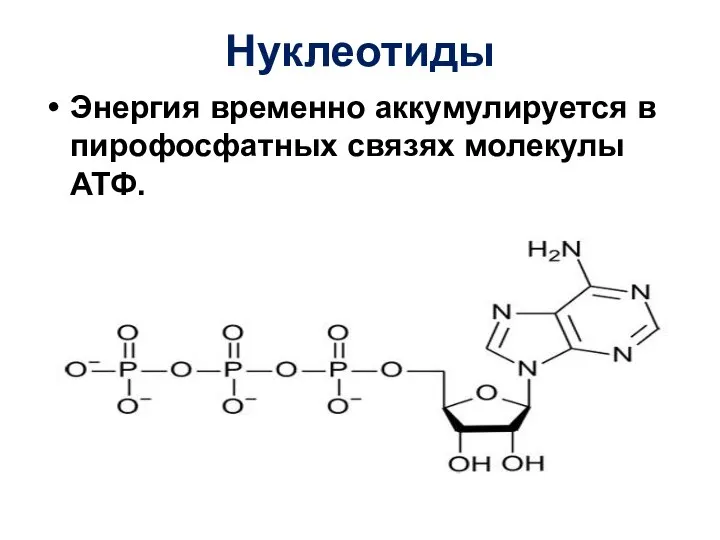
Слайд 19Нуклеотиды
Энергия временно аккумулируется в пирофосфатных связях молекулы АТФ.
Нуклеотиды
Энергия временно аккумулируется в пирофосфатных связях молекулы АТФ.
Слайд 20Примером нуклеотида, выполняющего в клетках роль регулятора обменных процессов, является циклическая аденозинмонофосфорная
Примером нуклеотида, выполняющего в клетках роль регулятора обменных процессов, является циклическая аденозинмонофосфорная

Слайд 21цАМФ
цАМФ
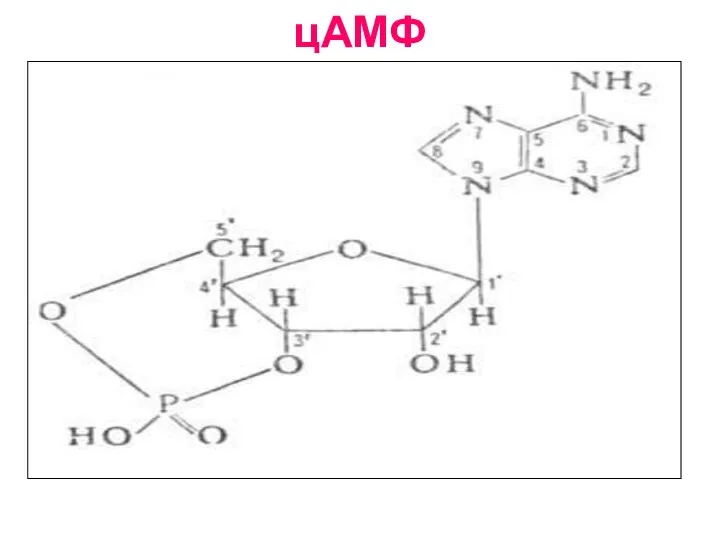
Слайд 22цАМФ
В структуре цАМФ один остаток фосфорной кислоты соединён одновременно и с третьим(С3),
цАМФ
В структуре цАМФ один остаток фосфорной кислоты соединён одновременно и с третьим(С3),

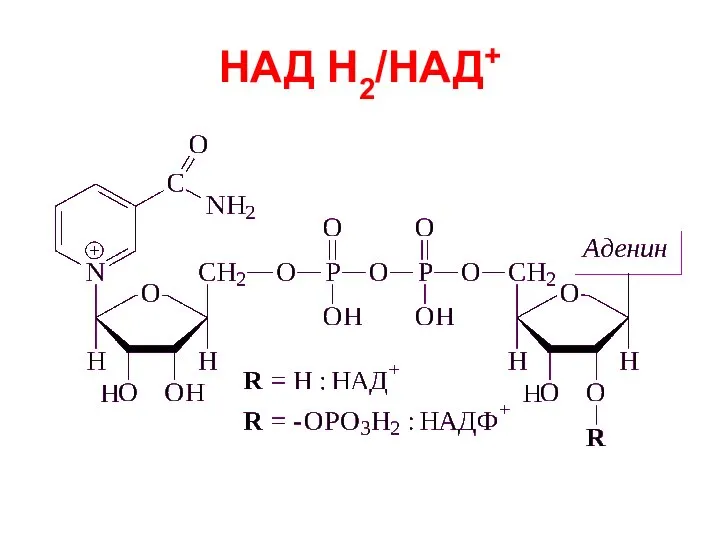
Слайд 23НАД Н2/НАД+
НАД Н2/НАД+
Слайд 24Первичная структура НК
В процессе образования полинуклеотида 2 мононуклеотида соединяются друг с другом
Первичная структура НК
В процессе образования полинуклеотида 2 мононуклеотида соединяются друг с другом

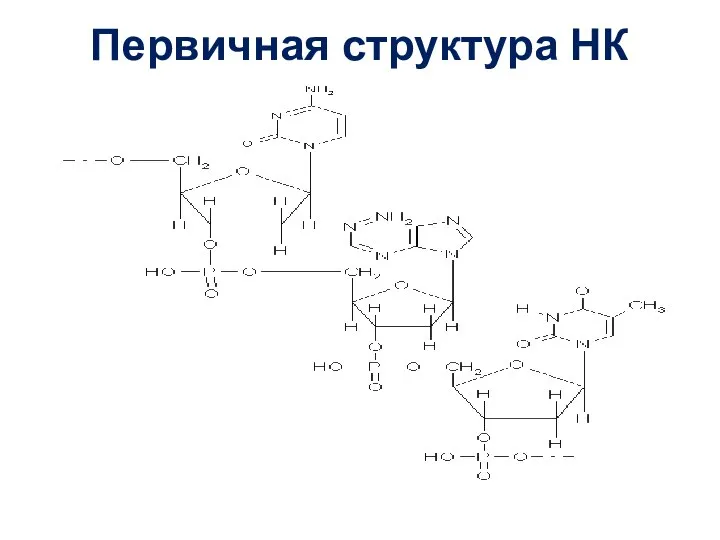
Слайд 25Первичная структура НК
Первичная структура НК
Слайд 26Первичная структура НК
После выделения молекулы воды 2 мононуклеотида оказываются соединёнными сложноэфирной связью
Первичная структура НК
После выделения молекулы воды 2 мононуклеотида оказываются соединёнными сложноэфирной связью

Слайд 27Первичная структура НК
Образовавшийся нуклеотид имеет на одном конце остаток фосфорной кислоты, а
Первичная структура НК
Образовавшийся нуклеотид имеет на одном конце остаток фосфорной кислоты, а

Слайд 28Первичная структура НК
Количество мономерных звеньев в полимерной цепи может достигать 108 -109
Первичная структура НК
Количество мономерных звеньев в полимерной цепи может достигать 108 -109

Слайд 29Первичная структура НК
Полинуклеотидная цепь имеет начало и окончание.
Началом считается тот конец
Первичная структура НК
Полинуклеотидная цепь имеет начало и окончание.
Началом считается тот конец

Слайд 30Первичная структура НК
Окончанием полинуклеотида считается его конец, содержащий или свободный гидроксил у
Первичная структура НК
Окончанием полинуклеотида считается его конец, содержащий или свободный гидроксил у

Слайд 31Первичная структура НК
Полинуклеотидную цепь принято изображать в виде последовательности сокращённых названий азотистых
Первичная структура НК
Полинуклеотидную цепь принято изображать в виде последовательности сокращённых названий азотистых

Слайд 32Изучение нуклеотидного состава ДНК показало, что:
- количество пиримидинов в молекуле эквивалентно количеству
Изучение нуклеотидного состава ДНК показало, что:
- количество пиримидинов в молекуле эквивалентно количеству

Слайд 33Тем не менее ДНК каждого отдельного вида животного, растения или микроорганизма обладает
Тем не менее ДНК каждого отдельного вида животного, растения или микроорганизма обладает

Слайд 34Вторичная структура ДНК
Согласно современным представлениям молекула ДНК представляет собой не одиночную, а
Вторичная структура ДНК
Согласно современным представлениям молекула ДНК представляет собой не одиночную, а

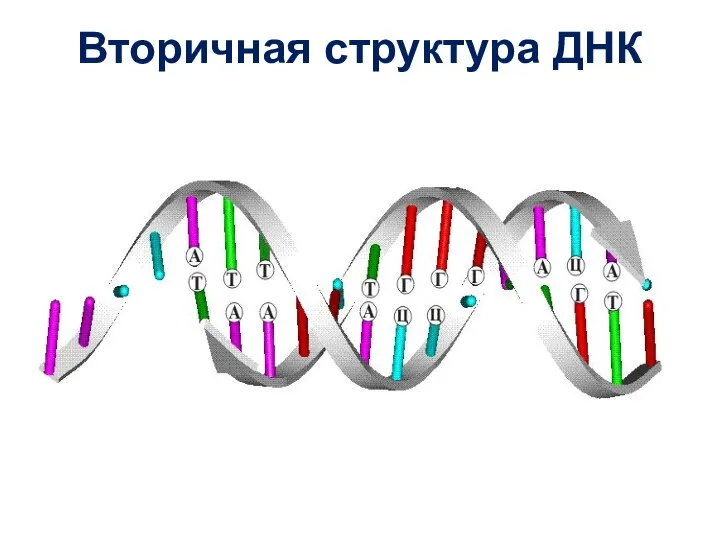
Слайд 35Вторичная структура ДНК
Вторичная структура ДНК
Слайд 36Вторичная структура ДНК
Спираль имеет правую закрутку и на каждый виток спирали приходится
Вторичная структура ДНК
Спираль имеет правую закрутку и на каждый виток спирали приходится

Слайд 37Вторичная структура ДНК
Азотистые основания одной цепи взаимодействуют с азотистыми основаниями другой цепи
Вторичная структура ДНК
Азотистые основания одной цепи взаимодействуют с азотистыми основаниями другой цепи

Слайд 38Вторичная структура ДНК
Долгое время считалось, что стабильность двойной спирали ДНК обусловлена только
Вторичная структура ДНК
Долгое время считалось, что стабильность двойной спирали ДНК обусловлена только

Слайд 39Стэкинг –взаимодействие -
-суть его состоит в том, что плоскости азотистых оснований молекулы
Стэкинг –взаимодействие -
-суть его состоит в том, что плоскости азотистых оснований молекулы

Слайд 40Стэкинг –взаимодействие
В результата вся молекула оказывается «прошитой» по длине особым типом связей,
Стэкинг –взаимодействие
В результата вся молекула оказывается «прошитой» по длине особым типом связей,

Слайд 41Вторичная структура ДНК
Стэкинг -взаимодействие
Вторичная структура ДНК
Стэкинг -взаимодействие
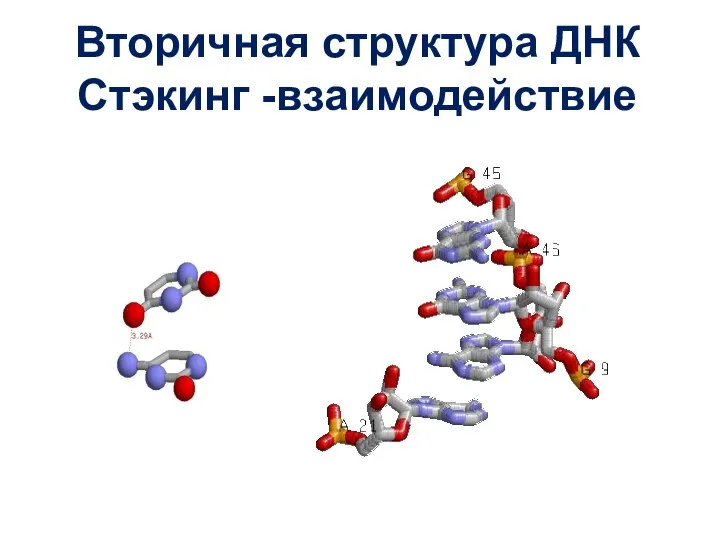
Слайд 42Вторичная структура ДНК
Поскольку все азотистые основания одной полинуклеотидной цепи ДНК комплементарны азотистым
Вторичная структура ДНК
Поскольку все азотистые основания одной полинуклеотидной цепи ДНК комплементарны азотистым

Слайд 43Вторичная структура ДНК
Регулярная пространственная структура, возникающая в результате взаимодействия двух антипараллельных комплементарных
Вторичная структура ДНК
Регулярная пространственная структура, возникающая в результате взаимодействия двух антипараллельных комплементарных

Слайд 44РНК
В отличие от ДНК молекулы РНК не имеют каких-либо выраженных закономерностей в
РНК
В отличие от ДНК молекулы РНК не имеют каких-либо выраженных закономерностей в

Слайд 45РНК
В качестве элементов вторичной структуры РНК обычно рассматривают так называемые «шпильки», представляющие
РНК
В качестве элементов вторичной структуры РНК обычно рассматривают так называемые «шпильки», представляющие

Слайд 46РНК
Полной комплементарности между восходящим и нисходящим участками в полинуклеотидной цепи в такой
РНК
Полной комплементарности между восходящим и нисходящим участками в полинуклеотидной цепи в такой

Слайд 47РНК
В то время как состав и структура ДНК постоянны для клеток данного
РНК
В то время как состав и структура ДНК постоянны для клеток данного

Слайд 48РНК
Рибосомальная РНК – входит в состав рибосом, клеточных органелл, на которых происходит
РНК
Рибосомальная РНК – входит в состав рибосом, клеточных органелл, на которых происходит

 Белки. Функции белков
Белки. Функции белков Белки и ферменты
Белки и ферменты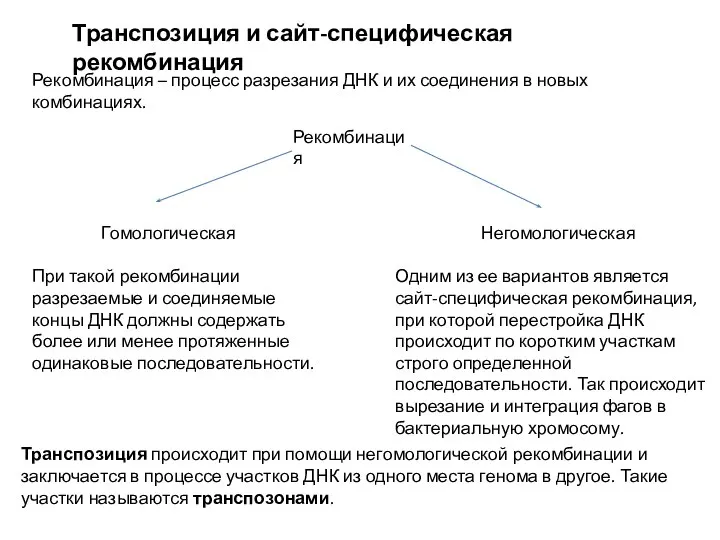 Транспозиция и сайт - специфическая рекомбинация
Транспозиция и сайт - специфическая рекомбинация Адаптация живых организмов к экстремальным значениям солености
Адаптация живых организмов к экстремальным значениям солености Презентация на тему Пищевые связи
Презентация на тему Пищевые связи  Корень: функции, морфология, анатомия, видоизменения
Корень: функции, морфология, анатомия, видоизменения Popular Dietary Practices
Popular Dietary Practices Растительная клетка (2)
Растительная клетка (2) Растения степей
Растения степей Презентация на тему Увеличительные приборы и приготовление микропрепарата
Презентация на тему Увеличительные приборы и приготовление микропрепарата  Голосеменные. Гинкго (Ginkgo biloba
Голосеменные. Гинкго (Ginkgo biloba Бактериялар фиологиясы
Бактериялар фиологиясы Закономерности наследования признаков
Закономерности наследования признаков Презентация на тему РАННИЕ ПОЛОВЫЕ СВЯЗИ И ИХ ПОСЛЕДСТВИЯ
Презентация на тему РАННИЕ ПОЛОВЫЕ СВЯЗИ И ИХ ПОСЛЕДСТВИЯ  Общая характеристика птиц
Общая характеристика птиц Строение корня и побега растений
Строение корня и побега растений Знакомые незнакомцы. Путешествие с домашними животными
Знакомые незнакомцы. Путешествие с домашними животными анализаторы
анализаторы Липиды. Классификация липидов
Липиды. Классификация липидов Особенности внутреннего строения птиц в связи с приспособленностью к полету
Особенности внутреннего строения птиц в связи с приспособленностью к полету Гипотеза панспермии
Гипотеза панспермии Породи собак
Породи собак Дыхание и кровообращение (3 класс)
Дыхание и кровообращение (3 класс) Насекомые. Бабочки и жуки
Насекомые. Бабочки и жуки День птиц
День птиц Волшебный витамин или химические элементы в живой клетке
Волшебный витамин или химические элементы в живой клетке Что растёт на клумбе
Что растёт на клумбе Аңқұстардың адам тіршілігіне әсері
Аңқұстардың адам тіршілігіне әсері