- Обмен липидов

Содержание
- 2. Лекция 11 Липогенез
- 3. Актуальность темы Липогенез – совокупность метаболических путей обмена липидов, связанных с синтезом высших жирных кислот (ВЖК)
- 4. План лекции Синтез ТАГ в тканях Транспорт эндогенных ТАГ из печени в ткани Нарушения транспорта эндогенных
- 5. Цель лекции Знать: химико-биологическую сущность процессов липогенеза, протекающих в организме человека Использовать знания о липогенезе для
- 6. План основной характеристики метаболических процессов Когда преимущественно идет процесс в норме (абсорбтивный, постабсорбтивный период, голодание, физическая
- 7. План характеристики гормонов-регуляторов метаболических процессов Химическая природа гормона (белково-пептидный, стероидный, производное аминокислоты) Место синтеза (органы, ткани)
- 8. СИНТЕЗ ТАГ В ТКАНЯХ Происходит в абсорбтивный период (после приема пищи, содержащей углеводы и липиды) Основное
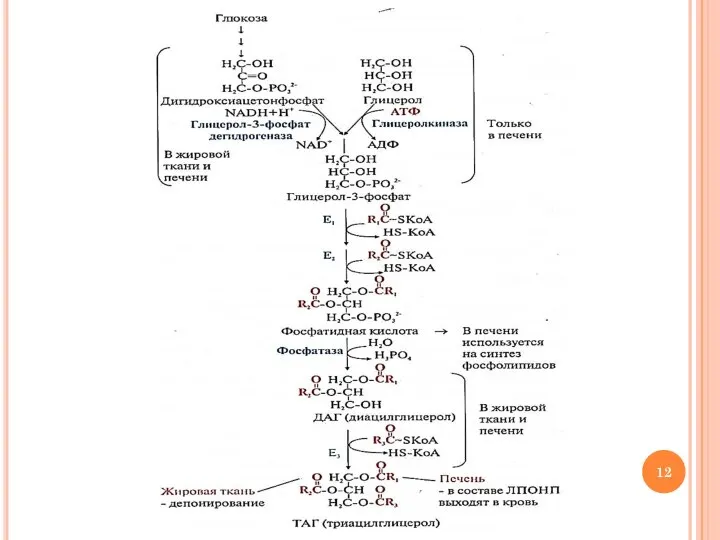
- 9. Синтез ТАГ: этап 1 Образование активных форм субстратов Образование активной формы ВЖК RCOOH + HS-КoA +
- 10. Синтез ТАГ: этап 1 Образование активных форм субстратов Образование активной формы глицерола глицерол + АТФ →
- 11. Синтез ТАГ: этап 2 Перенос ацильных остатков 1) глицерол-3-Р + 2 ацил-КоА → 1,2-ДАГ-3-Р (фосфатидная кислота)
- 13. Метаболизм ЛПОНП, сформированных в печени и транспортирующих ТАГ в ткани ЛПОНП формируются в печени, в кровотоке
- 14. Нарушение транспорта эндогенных ТАГ: жировое перерождение печени Избыточное накопление жира (ТАГ) в печени при нарушении транспорта
- 15. СИНТЕЗ ВЖК В ТКАНЯХ Происходит в абсорбтивный период Значение: трансформация избытка углеводов и аккумулирование их энергии
- 16. Синтез ВЖК в тканях (ПРОДОЛЖЕНИЕ) Источники NADPН: ПФП окисления глюкозы (глюкозо-6-фосфат дегидрогеназа, 6-фосфоглюконатдегидрогеназа) Окислительное декарбоксилирование малата
- 17. Синтез ВЖК в тканях Основные этапы: перенос ацетильных групп ацетил-КоА из митохондрий в цитоплазму в составе
- 18. Синтез ВЖК: этап 1. Перенос ацетильных групп из митохондрий в цитоплазму Митохондрии ацетил-КоА + ЩУК +
- 19. Использование ЩУК в цитоплазме ЩУК + NADН+Н+ → малат + NAD+ фермент: NAD-малатдегидрогеназа Малат + NADP+
- 20. Синтез ЖК: этап 2 Образование малонил-КоА Ключевая реакция синтеза ВЖК ацетил-КоА + СО2 + АТФ →
- 21. Синтез ВЖК: этап 3 Удлинение углеродной цепи Ацетил-КоА – источник С15 и С16 атомов пальмитиновой кислоты
- 22. Удлинение углеродной цепи Мультиферментный комплекс - пальмитоилсинтаза Структура комплекса: димер, состоящий из 2-х идентичных полипептидных мономеров
- 23. Структура мономеров пальмитоилсинтазного комплекса 7 доменов ацилпереносящий белок (АПБ), содержащий витамин В5 - пантотеновую кислоту в
- 24. кетоацил - синтаза трансацилаза Граница между функциональными единицами Граница между мономерами Еноилт- редуктаза гидратаза кетоацил-редуктаза ацил-переносящий
- 25. Этапы синтеза пальмитиновой кислоты Перенос ацетильного и малонильного остатков на активные центры пальмитоилсинтазы (реакции 1,2) (трансацилаза)
- 27. Суммарное уравнение синтеза пальмитиновой кислоты на пальмитоилсинтазном комплексе ацетил-КоА + 7 малонил-КоА + 14 NADPН +
- 28. Синтез ненасыщенных жирных кислот – реакции десатурации (образование двойных связей) Синтез пальмитоолеиновой кислоты С16:1, 9, ώ7
- 29. Взаимосвязь углеводного обмена и липогенеза
- 30. «Точки соприкосновения» липогенеза и обмена глюкозы Гликолиз – источник АТФ для реакций синтеза ВЖК и ТАГ
- 31. Регуляция липогенеза Синтез ВЖК «запускается» инсулином Вспомните механизм передачи сигнала инсулина в клетки-мишени! Механизмы регуляции стимулирование
- 32. Механизмы регуляции липогенеза инсулином (продолжение) индукция синтеза ферментов в печени и жировой ткани липидного обмена: ЛП-липазы,
- 33. Аллостерическая регуляция липогенеза При избыточном потреблении легкоусвояемых углеводов и активации гликолиза, реакций ОПК энергетический статус гепатоцитов
- 34. Регуляция активности ацетил-КоА карбоксилазы – ключевого фермента синтеза ВЖК Способы регуляции индукция синтеза (инсулин) ассоциация /диссоциация
- 35. Схема регуляции активности ацетил-КоА карбоксилазы
- 36. абсорбтивный период После приема пищи, содержащей липиды и углеводы в крови повышается Уровень хиломикронов (ТАГ) Концентрация
- 37. Ожирение В норме у человека с массой тела 70 кг количество жира в депо - 10-11
- 38. Первичное ожирение: причины Алиментарный дисбаланс – избыточная калорийность питания по сравнению с расходами энергии Генетические факторы
- 39. Первичное ожирение: причины (продолжение) Генетические факторы ожирения Например: мутация гена белка адипоцитов – лептина лептин действует
- 40. Вторичное ожирение: причины Вторичное ожирение развивается в результате какого-либо заболевания (чаще эндокринного, например, гипотиреоза) Чем опасно
- 41. Задание для самостоятельной работы Используя интернет-ресурсы, найдите информацию на тему: Жировая ткань как эндокринный орган: адипокины
- 42. Заключение Процессы липогенеза (синтез ВЖК и ТАГ) являются источником образования в организме резервного «топлива» с большой
- 44. Скачать презентацию

Слайд 3Актуальность темы
Липогенез – совокупность метаболических путей обмена липидов, связанных с синтезом высших
Актуальность темы
Липогенез – совокупность метаболических путей обмена липидов, связанных с синтезом высших

Слайд 4План лекции
Синтез ТАГ в тканях
Транспорт эндогенных ТАГ из печени в ткани
Нарушения транспорта
План лекции
Синтез ТАГ в тканях
Транспорт эндогенных ТАГ из печени в ткани
Нарушения транспорта

Слайд 5Цель лекции
Знать: химико-биологическую сущность процессов липогенеза, протекающих в организме человека
Использовать знания о
Цель лекции
Знать: химико-биологическую сущность процессов липогенеза, протекающих в организме человека
Использовать знания о

Слайд 6План основной характеристики метаболических процессов
Когда преимущественно идет процесс в норме (абсорбтивный, постабсорбтивный
План основной характеристики метаболических процессов
Когда преимущественно идет процесс в норме (абсорбтивный, постабсорбтивный

Слайд 7План характеристики гормонов-регуляторов метаболических процессов
Химическая природа гормона (белково-пептидный, стероидный, производное аминокислоты)
Место синтеза
План характеристики гормонов-регуляторов метаболических процессов
Химическая природа гормона (белково-пептидный, стероидный, производное аминокислоты)
Место синтеза

Слайд 8СИНТЕЗ ТАГ В ТКАНЯХ
Происходит в абсорбтивный период (после приема пищи, содержащей углеводы
СИНТЕЗ ТАГ В ТКАНЯХ
Происходит в абсорбтивный период (после приема пищи, содержащей углеводы

Слайд 9Синтез ТАГ: этап 1
Образование активных форм субстратов
Образование активной формы ВЖК
RCOOH + HS-КoA
Синтез ТАГ: этап 1
Образование активных форм субстратов
Образование активной формы ВЖК
RCOOH + HS-КoA

Слайд 10Синтез ТАГ: этап 1
Образование активных форм субстратов
Образование активной формы глицерола
глицерол + АТФ
Синтез ТАГ: этап 1
Образование активных форм субстратов
Образование активной формы глицерола
глицерол + АТФ

Слайд 11Синтез ТАГ: этап 2
Перенос ацильных остатков
1) глицерол-3-Р + 2 ацил-КоА → 1,2-ДАГ-3-Р
Синтез ТАГ: этап 2
Перенос ацильных остатков
1) глицерол-3-Р + 2 ацил-КоА → 1,2-ДАГ-3-Р

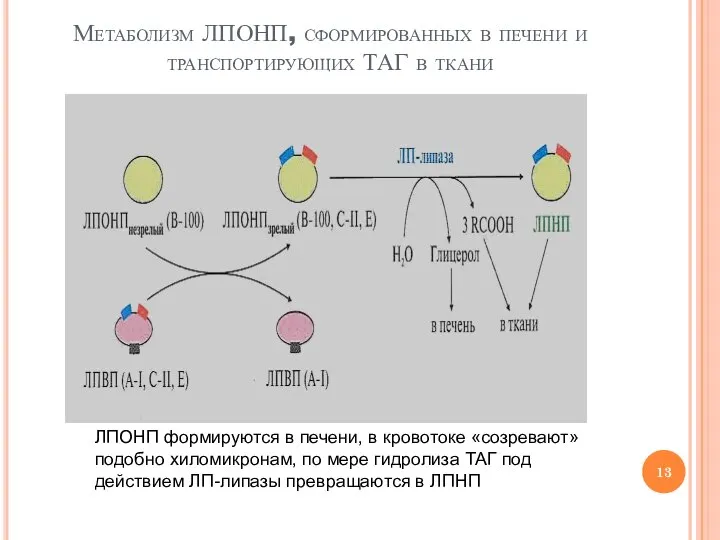
Слайд 13Метаболизм ЛПОНП, сформированных в печени и транспортирующих ТАГ в ткани
ЛПОНП формируются в
Метаболизм ЛПОНП, сформированных в печени и транспортирующих ТАГ в ткани
ЛПОНП формируются в
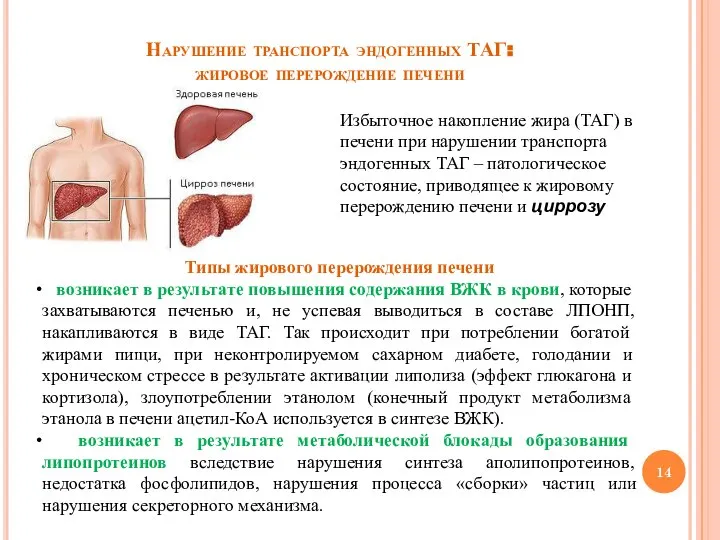
Слайд 14Нарушение транспорта эндогенных ТАГ:
жировое перерождение печени
Избыточное накопление жира (ТАГ) в печени при
Нарушение транспорта эндогенных ТАГ:
жировое перерождение печени
Избыточное накопление жира (ТАГ) в печени при
Слайд 15СИНТЕЗ ВЖК В ТКАНЯХ
Происходит в абсорбтивный период
Значение: трансформация избытка углеводов и аккумулирование
СИНТЕЗ ВЖК В ТКАНЯХ
Происходит в абсорбтивный период
Значение: трансформация избытка углеводов и аккумулирование

Слайд 16Синтез ВЖК в тканях (ПРОДОЛЖЕНИЕ)
Источники NADPН:
ПФП окисления глюкозы (глюкозо-6-фосфат дегидрогеназа, 6-фосфоглюконатдегидрогеназа)
Окислительное декарбоксилирование
Синтез ВЖК в тканях (ПРОДОЛЖЕНИЕ)
Источники NADPН:
ПФП окисления глюкозы (глюкозо-6-фосфат дегидрогеназа, 6-фосфоглюконатдегидрогеназа)
Окислительное декарбоксилирование

Слайд 17Синтез ВЖК в тканях
Основные этапы:
перенос ацетильных групп ацетил-КоА из митохондрий в цитоплазму
Синтез ВЖК в тканях
Основные этапы:
перенос ацетильных групп ацетил-КоА из митохондрий в цитоплазму

Слайд 18Синтез ВЖК: этап 1. Перенос ацетильных групп из митохондрий в цитоплазму
Митохондрии
ацетил-КоА +
Синтез ВЖК: этап 1. Перенос ацетильных групп из митохондрий в цитоплазму
Митохондрии
ацетил-КоА +

Слайд 19Использование ЩУК в цитоплазме
ЩУК + NADН+Н+ → малат + NAD+
фермент: NAD-малатдегидрогеназа
Малат
Использование ЩУК в цитоплазме
ЩУК + NADН+Н+ → малат + NAD+
фермент: NAD-малатдегидрогеназа
Малат

Слайд 20Синтез ЖК: этап 2
Образование малонил-КоА
Ключевая реакция синтеза ВЖК
ацетил-КоА + СО2 + АТФ
Синтез ЖК: этап 2
Образование малонил-КоА
Ключевая реакция синтеза ВЖК
ацетил-КоА + СО2 + АТФ

Слайд 21Синтез ВЖК: этап 3
Удлинение углеродной цепи
Ацетил-КоА – источник С15 и С16 атомов
Синтез ВЖК: этап 3
Удлинение углеродной цепи
Ацетил-КоА – источник С15 и С16 атомов

Слайд 22Удлинение углеродной цепи
Мультиферментный комплекс - пальмитоилсинтаза
Структура комплекса: димер, состоящий из 2-х
Удлинение углеродной цепи
Мультиферментный комплекс - пальмитоилсинтаза
Структура комплекса: димер, состоящий из 2-х

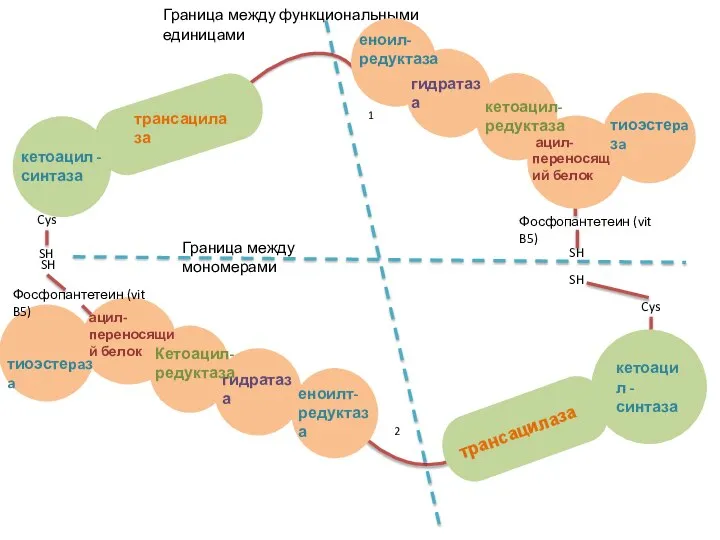
Слайд 23Структура мономеров пальмитоилсинтазного комплекса
7 доменов
ацилпереносящий белок (АПБ), содержащий витамин В5 - пантотеновую
Структура мономеров пальмитоилсинтазного комплекса
7 доменов
ацилпереносящий белок (АПБ), содержащий витамин В5 - пантотеновую

Слайд 24кетоацил - синтаза
трансацилаза
Граница между функциональными единицами
Граница между мономерами
Еноилт- редуктаза
гидратаза
кетоацил-редуктаза
ацил-переносящий белок
тиоэстеpaзa
кетоацил -
кетоацил - синтаза
трансацилаза
Граница между функциональными единицами
Граница между мономерами
Еноилт- редуктаза
гидратаза
кетоацил-редуктаза
ацил-переносящий белок
тиоэстеpaзa
кетоацил -
Слайд 25Этапы синтеза пальмитиновой кислоты
Перенос ацетильного и малонильного остатков на активные центры пальмитоилсинтазы
Этапы синтеза пальмитиновой кислоты
Перенос ацетильного и малонильного остатков на активные центры пальмитоилсинтазы

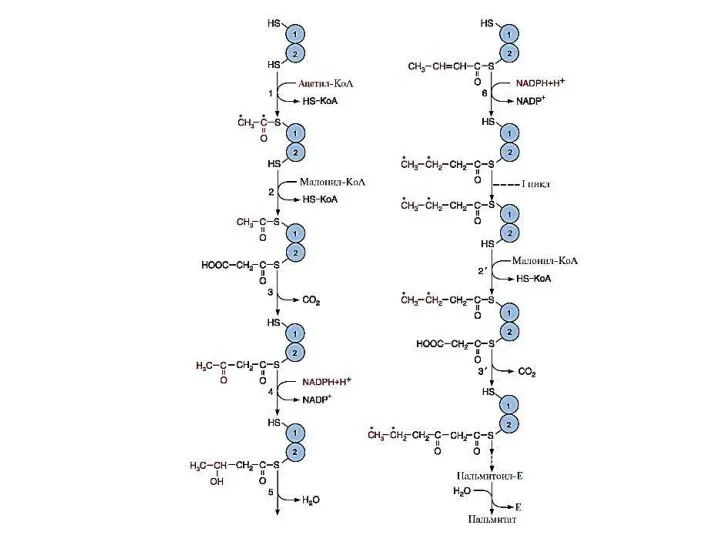
Слайд 27Суммарное уравнение синтеза пальмитиновой кислоты на пальмитоилсинтазном комплексе
ацетил-КоА + 7 малонил-КоА +
Суммарное уравнение синтеза пальмитиновой кислоты на пальмитоилсинтазном комплексе
ацетил-КоА + 7 малонил-КоА +

Слайд 28Синтез ненасыщенных жирных кислот – реакции десатурации (образование двойных связей)
Синтез пальмитоолеиновой кислоты
С16:1,
Синтез ненасыщенных жирных кислот – реакции десатурации (образование двойных связей)
Синтез пальмитоолеиновой кислоты
С16:1,

Слайд 29Взаимосвязь углеводного обмена и липогенеза
Взаимосвязь углеводного обмена и липогенеза
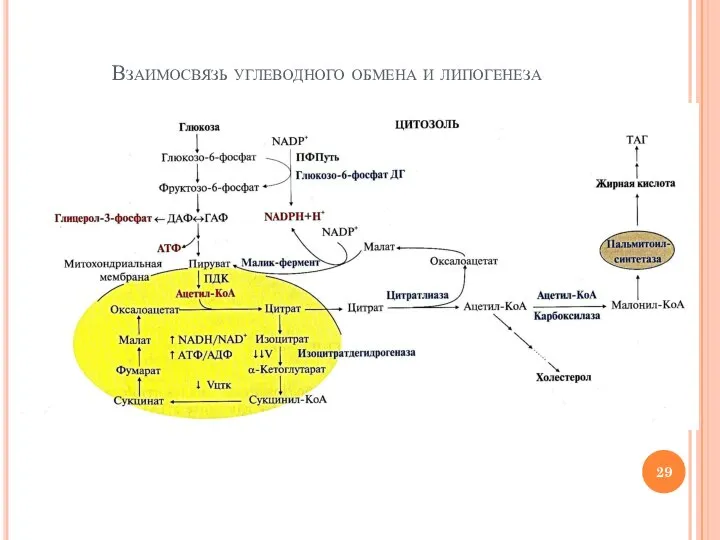
Слайд 30«Точки соприкосновения» липогенеза и обмена глюкозы
Гликолиз – источник АТФ для реакций синтеза
«Точки соприкосновения» липогенеза и обмена глюкозы
Гликолиз – источник АТФ для реакций синтеза

Слайд 31Регуляция липогенеза
Синтез ВЖК «запускается» инсулином
Вспомните механизм передачи сигнала инсулина в клетки-мишени!
Механизмы регуляции
стимулирование
Регуляция липогенеза
Синтез ВЖК «запускается» инсулином
Вспомните механизм передачи сигнала инсулина в клетки-мишени!
Механизмы регуляции
стимулирование

Слайд 32Механизмы регуляции липогенеза инсулином (продолжение)
индукция синтеза ферментов в печени и жировой ткани
липидного
Механизмы регуляции липогенеза инсулином (продолжение)
индукция синтеза ферментов в печени и жировой ткани
липидного

Слайд 33Аллостерическая регуляция липогенеза
При избыточном потреблении легкоусвояемых углеводов и активации гликолиза, реакций ОПК
Аллостерическая регуляция липогенеза
При избыточном потреблении легкоусвояемых углеводов и активации гликолиза, реакций ОПК

Слайд 34Регуляция активности ацетил-КоА карбоксилазы – ключевого фермента синтеза ВЖК
Способы регуляции
индукция синтеза (инсулин)
ассоциация
Регуляция активности ацетил-КоА карбоксилазы – ключевого фермента синтеза ВЖК
Способы регуляции
индукция синтеза (инсулин)
ассоциация

Слайд 35Схема регуляции активности
ацетил-КоА карбоксилазы
Схема регуляции активности
ацетил-КоА карбоксилазы
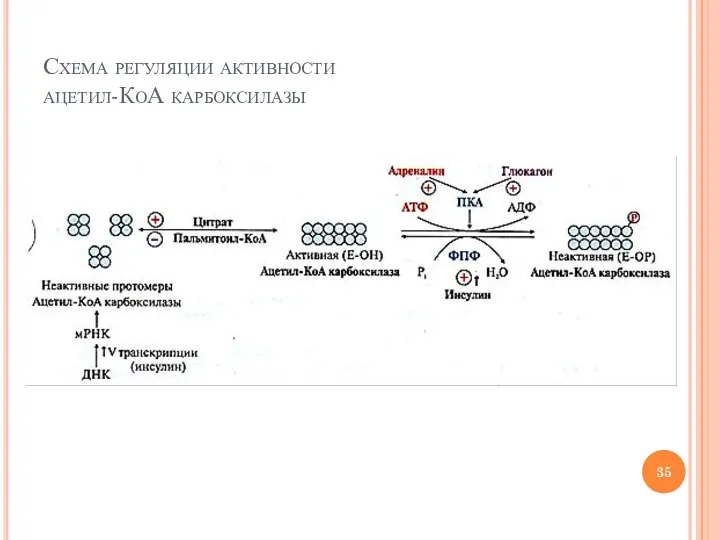
Слайд 36абсорбтивный период
После приема пищи, содержащей липиды и углеводы в крови повышается
Уровень хиломикронов
абсорбтивный период
После приема пищи, содержащей липиды и углеводы в крови повышается
Уровень хиломикронов

Слайд 37Ожирение
В норме у человека с массой тела 70 кг количество жира в
Ожирение
В норме у человека с массой тела 70 кг количество жира в

Слайд 38Первичное ожирение: причины
Алиментарный дисбаланс – избыточная калорийность питания по сравнению с расходами
Первичное ожирение: причины
Алиментарный дисбаланс – избыточная калорийность питания по сравнению с расходами

Слайд 39Первичное ожирение: причины (продолжение)
Генетические факторы ожирения
Например: мутация гена белка адипоцитов – лептина
лептин
Первичное ожирение: причины (продолжение)
Генетические факторы ожирения
Например: мутация гена белка адипоцитов – лептина
лептин

Слайд 40Вторичное ожирение: причины
Вторичное ожирение развивается в результате какого-либо заболевания (чаще эндокринного, например,
Вторичное ожирение: причины
Вторичное ожирение развивается в результате какого-либо заболевания (чаще эндокринного, например,

Слайд 41Задание для самостоятельной работы
Используя интернет-ресурсы, найдите информацию на тему:
Жировая ткань как эндокринный
Задание для самостоятельной работы
Используя интернет-ресурсы, найдите информацию на тему:
Жировая ткань как эндокринный

Слайд 42Заключение
Процессы липогенеза (синтез ВЖК и ТАГ) являются источником образования в организме резервного
Заключение
Процессы липогенеза (синтез ВЖК и ТАГ) являются источником образования в организме резервного

 Кровообращение. 8 класс
Кровообращение. 8 класс Дед и внук
Дед и внук Виды цветочного оформления
Виды цветочного оформления БИ6_ПОДГОТОВКА к ВПР_апрель 2019
БИ6_ПОДГОТОВКА к ВПР_апрель 2019 Презентация на тему Основные понятия генетики
Презентация на тему Основные понятия генетики  Мой друг - шотландский кот Вася
Мой друг - шотландский кот Вася Миксбордер. Хитрости и секреты миксбордеров
Миксбордер. Хитрости и секреты миксбордеров Изучение механизма наследственности
Изучение механизма наследственности Органы дыхания и газообмен
Органы дыхания и газообмен Куда растёт человечество
Куда растёт человечество Могут ли растения производить потомство без помощи семян?
Могут ли растения производить потомство без помощи семян? Monkeys. Mandrill
Monkeys. Mandrill Харофиты
Харофиты Промежуточный мозг (diencephalon)
Промежуточный мозг (diencephalon) Ретикулярная (сетчатая) формация
Ретикулярная (сетчатая) формация Sais-tu
Sais-tu Мегагаметофит. Лекция 1. О женском
Мегагаметофит. Лекция 1. О женском Симбиотические отношения среди морских организмов
Симбиотические отношения среди морских организмов Обмен веществ и энергии
Обмен веществ и энергии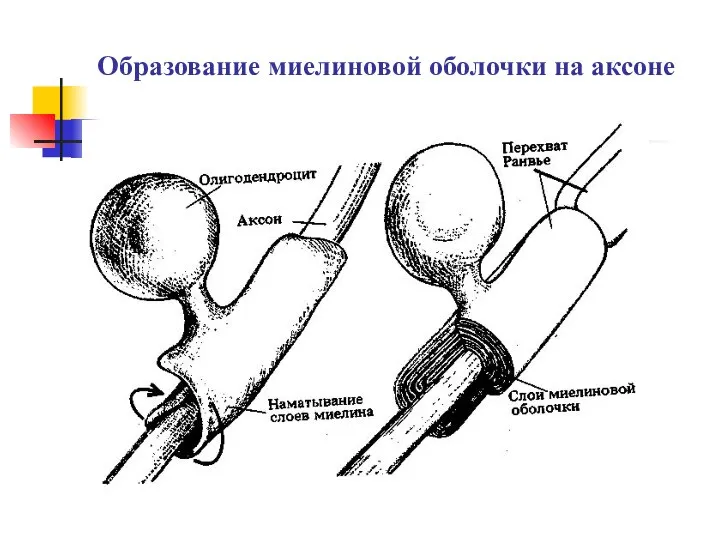 Образование миелиновой оболочки на аксоне
Образование миелиновой оболочки на аксоне «Общая характеристика надкласса Рыбы. Особенности внешнего строения рыб в связи с водной средой обитания».
«Общая характеристика надкласса Рыбы. Особенности внешнего строения рыб в связи с водной средой обитания». Внешнее строение птиц
Внешнее строение птиц Мышечная ткань
Мышечная ткань Мышцы
Мышцы Презентация на тему ЕГЭ по биологии
Презентация на тему ЕГЭ по биологии  Сравнительная характеристика микроскопа и Foldscope
Сравнительная характеристика микроскопа и Foldscope Вирусы- граница живой и неживой природы
Вирусы- граница живой и неживой природы Топыраққа биогенді элементтерді қолдану барысында олардың азаюы және жоғалу жағдайлары
Топыраққа биогенді элементтерді қолдану барысында олардың азаюы және жоғалу жағдайлары