- Обмен липидов. Анаболизм липидов

Содержание
- 2. АНАБОЛИЗМ ЛИПИДОВ СИНТЕЗ ЖИРНЫХ КИСЛОТ СИНТЕЗ ХОЛЕСТЕРОЛА СИНТЕЗ ТРИАЦИЛГЛИЦЕРОЛОВ (НЕЙТРАЛЬНЫХ ЖИРОВ) СИНТЕЗ ФОСФОЛИПИДОВ (ГЛИЦЕРОФОСФОЛИПИДОВ и СФИНГОФОСФОЛИПИДОВ)
- 3. СИНТЕЗ ЖИРНЫХ КИСЛОТ
- 4. СИНТЕЗ ЖИРНЫХ КИСЛОТ ЛОКАЛИЗАЦИЯ В КЛЕТКЕ: -ЦИТОПЛАЗМА (пальмитоилсинтаза синтезирует насыщенные ЖК до С16) -МИТОХОНДРИИ (элонгаза синтезирует
- 5. СИНТЕЗ ЖИРНЫХ КИСЛОТ Источник углерода для синтеза жирных кислот - ацетил-КоА, образующийся при окислении глюкозы в
- 6. СИНТЕЗ ЖИРНЫХ КИСЛОТ Синтез жирных кислот требует: источник углеродного скелета: ацетил-КоА источник энергии: АТФ дополнительный источник
- 7. СИНТЕЗ ЖИРНЫХ КИСЛОТ Активность фермента ацетил-КоА-карбоксилазы определяет скорость всех последующих реакций синтеза жирных кислот Карбоксилирование ацетил-КоА
- 8. СИНТЕЗ ЖИРНЫХ КИСЛОТ Строение мульферментного комплекса- ПАЛЬМИТОИЛСИНТАЗЫ Пальмитоилсинтаза - димер из двух полипептидных цепей. Каждая субъединица
- 9. СИНТЕЗ ЖИРНЫХ КИСЛОТ 1. Перенос ацетильной группы ацетил-КоА на тиоловую группу цистеина
- 10. СИНТЕЗ ЖИРНЫХ КИСЛОТ 2. Остаток малонила от малонил-КоА переносится на сульфгидрильную группу ацилпереносящего белка
- 11. 3. Ацетильная группа конденсируется с остатком малонила по месту отделившегося СО2 СИНТЕЗ ЖИРНЫХ КИСЛОТ
- 12. СИНТЕЗ ЖИРНЫХ КИСЛОТ 4. Кетоацильный остаток восстанавливается, донор атомов водорода - НАДФН2
- 13. СИНТЕЗ ЖИРНЫХ КИСЛОТ 5. Дегидратация β-гидроксиацильного остатка
- 14. СИНТЕЗ ЖИРНЫХ КИСЛОТ 6. Транс-еноильный остаток восстанавливается, донор атомов водорода - НАДФН2
- 15. СИНТЕЗ ЖИРНЫХ КИСЛОТ 7. Завершается первый этап синтеза перемещением радикала бутирила на свободную SH-группу цистеина Затем
- 16. СИНТЕЗ ЖИРНЫХ КИСЛОТ Радикал пальмитиновой кислоты гидролитически отделяется от ферментного комплекса, превращаясь в свободную пальмитиновую кислоту
- 17. ЭЛОНГАЦИЯ ЖИРНЫХ КИСЛОТ Элонгаза использует малонил-КоА в качестве донора углеродных атомов, а НАДФН2- в качестве восстановителя.
- 18. ДЕСАТУРАЦИЯ ЖИРНЫХ КИСЛОТ Десатурация- включение двойных связей в радикалы жирных кислот. Основные жирные кислоты, образующиеся в
- 19. РЕГУЛЯЦИЯ СИНТЕЗА ЖИРНЫХ КИСЛОТ Регулируется пищевым режимом: - тормозит голодание - активирует богатая углеводами пища 2.
- 20. СИНТЕЗ ХОЛЕСТЕРИНА Субстрат для синтеза холестерола- ацетил-КоА Активация синтеза холестерина происходит при поступлении в организм пищи,
- 21. СИНТЕЗ ХОЛЕСТЕРИНА ОСНОВНЫЕ СТАДИИ СИНТЕЗА ХОЛЕСТЕРИНА: • синтез мевалоната (С6) • синтез сквалена из мевалоната (С30)
- 22. СИНТЕЗ ХОЛЕСТЕРИНА: ОБРАЗОВАНИЕ МЕВАЛОНАТА Регуляторная реакция синтеза холестерина
- 23. СИНТЕЗ ХОЛЕСТЕРИНА: ОБРАЗОВАНИЕ СКВАЛЕНА
- 24. СИНТЕЗ ХОЛЕСТЕРИНА: ЦИКЛИЗАЦИЯ СКВАЛЕНА
- 25. РЕГУЛЯЦИЯ СИНТЕЗА ХОЛЕСТЕРИНА 1. Регулируется пищевым режимом: • тормозит голодание • активирует богатая углеводами пища 2.
- 26. СИНТЕЗ ФОСФАТИДНОЙ КИСЛОТЫ
- 27. СИНТЕЗ ТРИАЦИЛГЛИЦЕРОЛОВ 1. Биосинтез из фосфатидной кислоты (в печени и жировой ткани) 2. Ресинтез триацилглицеролов в
- 28. СИНТЕЗ ГЛИЦЕРОФОСФОЛИПИДОВ
- 30. Скачать презентацию
Слайд 2АНАБОЛИЗМ ЛИПИДОВ
СИНТЕЗ ЖИРНЫХ КИСЛОТ
СИНТЕЗ ХОЛЕСТЕРОЛА
СИНТЕЗ ТРИАЦИЛГЛИЦЕРОЛОВ (НЕЙТРАЛЬНЫХ ЖИРОВ)
СИНТЕЗ ФОСФОЛИПИДОВ
(ГЛИЦЕРОФОСФОЛИПИДОВ и
АНАБОЛИЗМ ЛИПИДОВ
СИНТЕЗ ЖИРНЫХ КИСЛОТ
СИНТЕЗ ХОЛЕСТЕРОЛА
СИНТЕЗ ТРИАЦИЛГЛИЦЕРОЛОВ (НЕЙТРАЛЬНЫХ ЖИРОВ)
СИНТЕЗ ФОСФОЛИПИДОВ
(ГЛИЦЕРОФОСФОЛИПИДОВ и

Слайд 3СИНТЕЗ ЖИРНЫХ КИСЛОТ
СИНТЕЗ ЖИРНЫХ КИСЛОТ

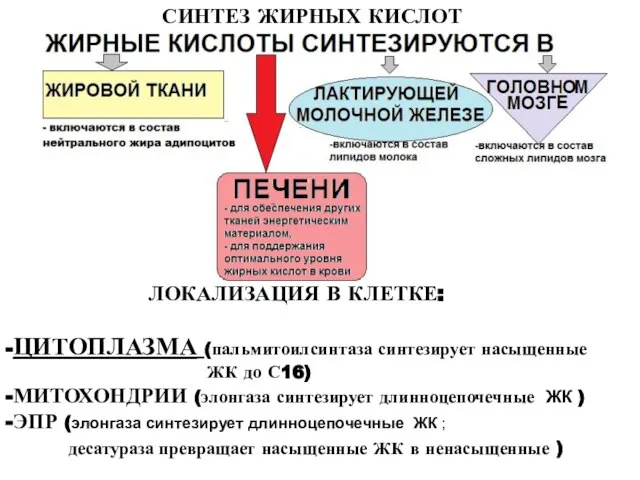
Слайд 4СИНТЕЗ ЖИРНЫХ КИСЛОТ
ЛОКАЛИЗАЦИЯ В КЛЕТКЕ:
-ЦИТОПЛАЗМА (пальмитоилсинтаза синтезирует насыщенные
ЖК до
СИНТЕЗ ЖИРНЫХ КИСЛОТ
ЛОКАЛИЗАЦИЯ В КЛЕТКЕ:
-ЦИТОПЛАЗМА (пальмитоилсинтаза синтезирует насыщенные
ЖК до
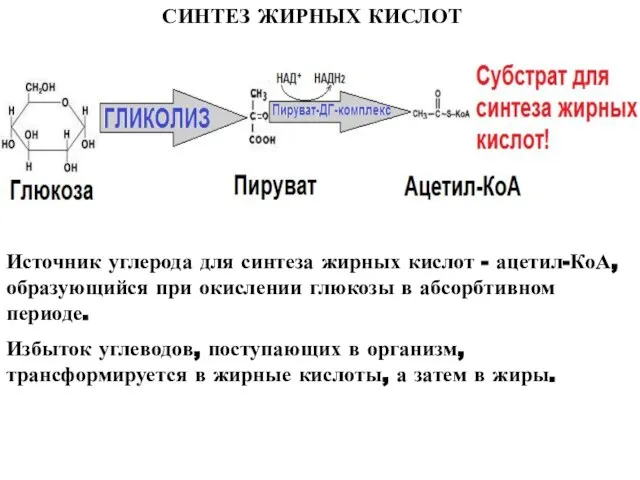
Слайд 5СИНТЕЗ ЖИРНЫХ КИСЛОТ
Источник углерода для синтеза жирных кислот - ацетил-КоА, образующийся при
СИНТЕЗ ЖИРНЫХ КИСЛОТ
Источник углерода для синтеза жирных кислот - ацетил-КоА, образующийся при
Слайд 6СИНТЕЗ ЖИРНЫХ КИСЛОТ
Синтез жирных кислот требует:
источник углеродного скелета: ацетил-КоА
источник энергии: АТФ
дополнительный
СИНТЕЗ ЖИРНЫХ КИСЛОТ
Синтез жирных кислот требует:
источник углеродного скелета: ацетил-КоА
источник энергии: АТФ
дополнительный

Слайд 7СИНТЕЗ ЖИРНЫХ КИСЛОТ
Активность фермента ацетил-КоА-карбоксилазы определяет скорость всех последующих реакций синтеза жирных
СИНТЕЗ ЖИРНЫХ КИСЛОТ
Активность фермента ацетил-КоА-карбоксилазы определяет скорость всех последующих реакций синтеза жирных
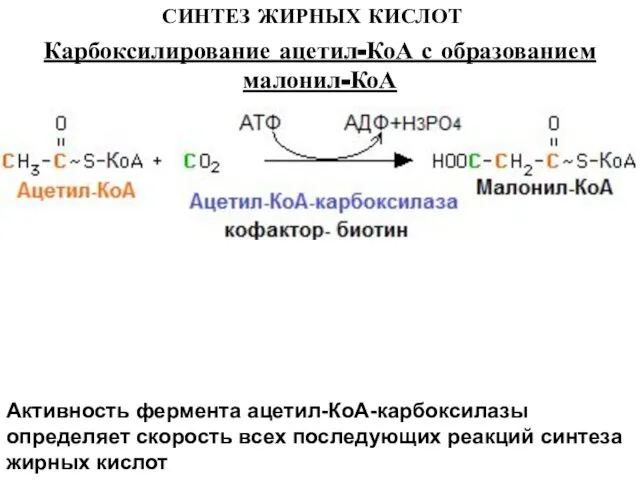
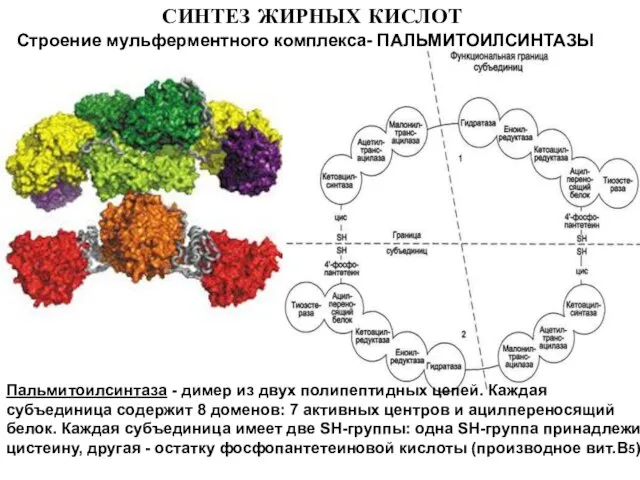
Слайд 8СИНТЕЗ ЖИРНЫХ КИСЛОТ
Строение мульферментного комплекса- ПАЛЬМИТОИЛСИНТАЗЫ
Пальмитоилсинтаза - димер из двух полипептидных цепей.
СИНТЕЗ ЖИРНЫХ КИСЛОТ
Строение мульферментного комплекса- ПАЛЬМИТОИЛСИНТАЗЫ
Пальмитоилсинтаза - димер из двух полипептидных цепей.
Слайд 9СИНТЕЗ ЖИРНЫХ КИСЛОТ
1. Перенос ацетильной группы ацетил-КоА на тиоловую группу цистеина
СИНТЕЗ ЖИРНЫХ КИСЛОТ
1. Перенос ацетильной группы ацетил-КоА на тиоловую группу цистеина
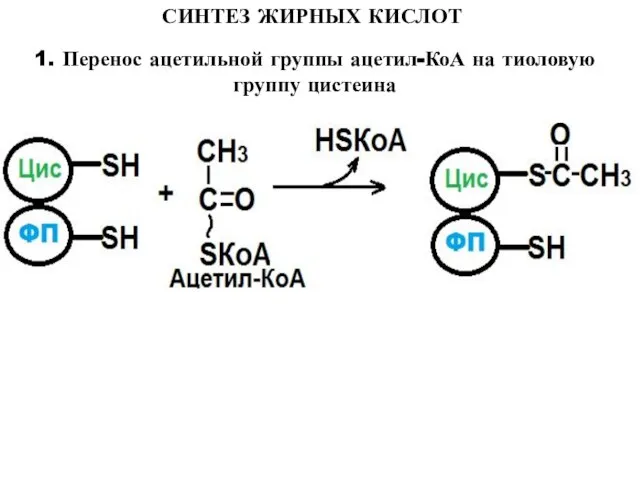
Слайд 10СИНТЕЗ ЖИРНЫХ КИСЛОТ
2. Остаток малонила от малонил-КоА переносится на сульфгидрильную группу ацилпереносящего
СИНТЕЗ ЖИРНЫХ КИСЛОТ
2. Остаток малонила от малонил-КоА переносится на сульфгидрильную группу ацилпереносящего
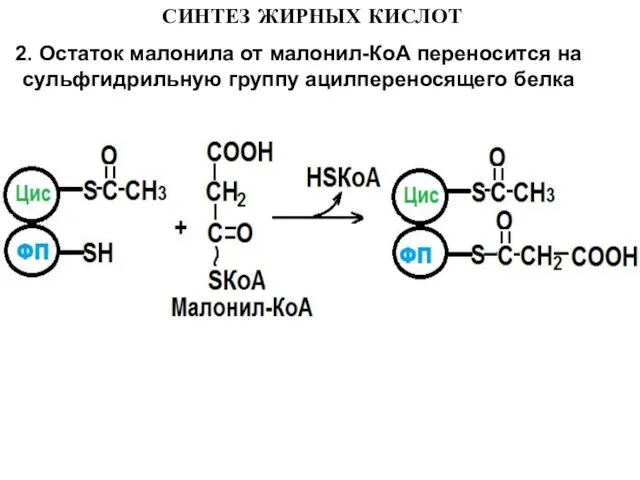
Слайд 113. Ацетильная группа конденсируется с остатком малонила по месту отделившегося СО2
СИНТЕЗ
3. Ацетильная группа конденсируется с остатком малонила по месту отделившегося СО2
СИНТЕЗ
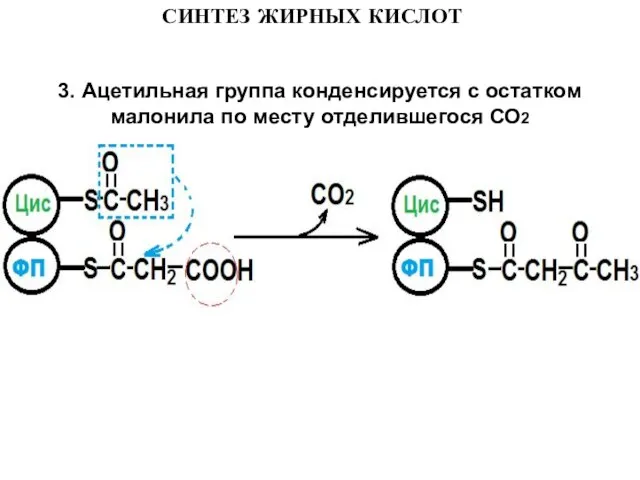
Слайд 12СИНТЕЗ ЖИРНЫХ КИСЛОТ
4. Кетоацильный остаток восстанавливается, донор атомов водорода - НАДФН2
СИНТЕЗ ЖИРНЫХ КИСЛОТ
4. Кетоацильный остаток восстанавливается, донор атомов водорода - НАДФН2
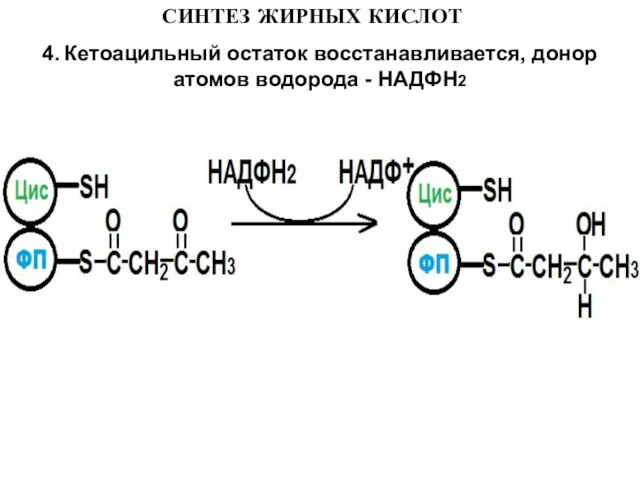
Слайд 13СИНТЕЗ ЖИРНЫХ КИСЛОТ
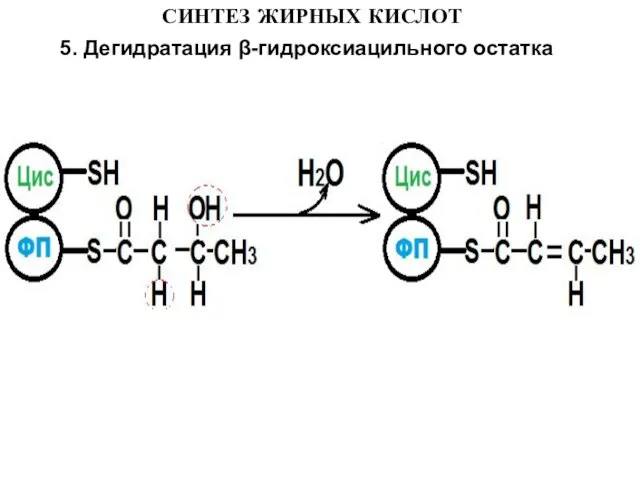
5. Дегидратация β-гидроксиацильного остатка
СИНТЕЗ ЖИРНЫХ КИСЛОТ
5. Дегидратация β-гидроксиацильного остатка
Слайд 14СИНТЕЗ ЖИРНЫХ КИСЛОТ
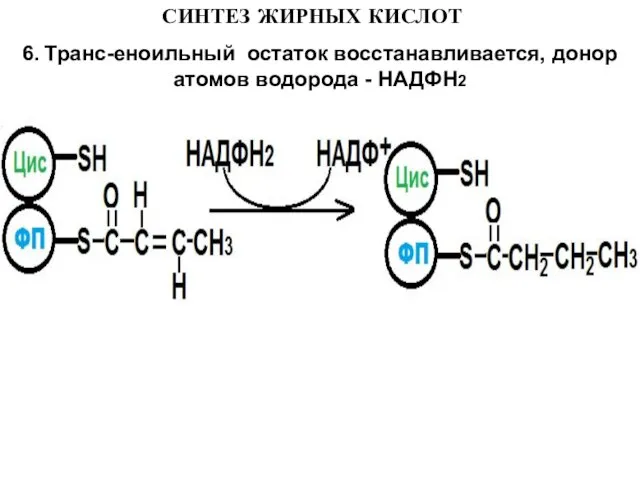
6. Транс-еноильный остаток восстанавливается, донор атомов водорода - НАДФН2
СИНТЕЗ ЖИРНЫХ КИСЛОТ
6. Транс-еноильный остаток восстанавливается, донор атомов водорода - НАДФН2
Слайд 15СИНТЕЗ ЖИРНЫХ КИСЛОТ
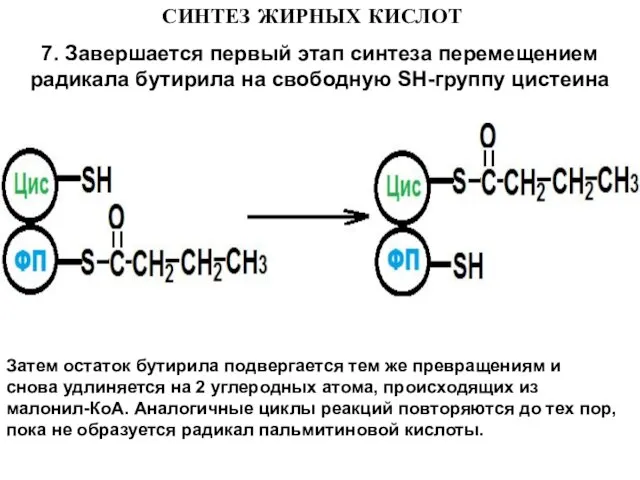
7. Завершается первый этап синтеза перемещением радикала бутирила на свободную
СИНТЕЗ ЖИРНЫХ КИСЛОТ
7. Завершается первый этап синтеза перемещением радикала бутирила на свободную
Слайд 16СИНТЕЗ ЖИРНЫХ КИСЛОТ
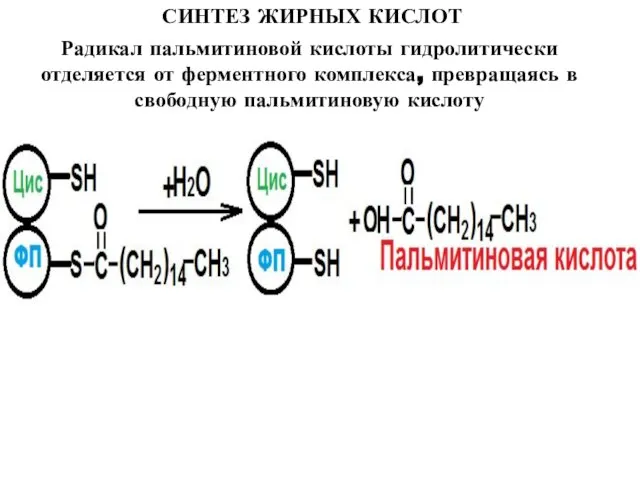
Радикал пальмитиновой кислоты гидролитически отделяется от ферментного комплекса, превращаясь в
СИНТЕЗ ЖИРНЫХ КИСЛОТ
Радикал пальмитиновой кислоты гидролитически отделяется от ферментного комплекса, превращаясь в
Слайд 17ЭЛОНГАЦИЯ ЖИРНЫХ КИСЛОТ
Элонгаза использует малонил-КоА в качестве донора углеродных атомов, а НАДФН2-
ЭЛОНГАЦИЯ ЖИРНЫХ КИСЛОТ
Элонгаза использует малонил-КоА в качестве донора углеродных атомов, а НАДФН2-

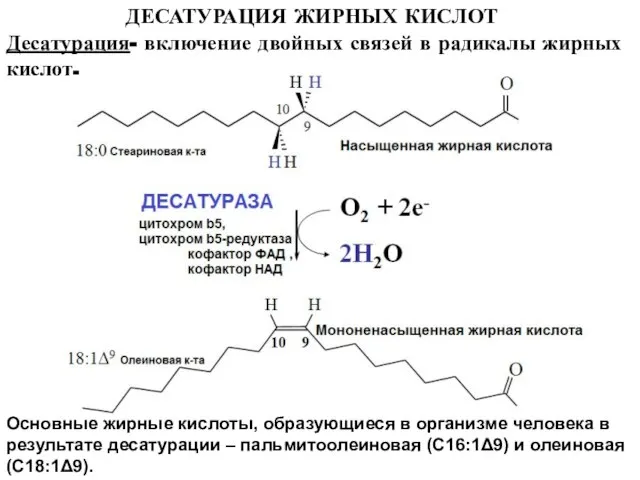
Слайд 18ДЕСАТУРАЦИЯ ЖИРНЫХ КИСЛОТ
Десатурация- включение двойных связей в радикалы жирных кислот.
Основные жирные
ДЕСАТУРАЦИЯ ЖИРНЫХ КИСЛОТ
Десатурация- включение двойных связей в радикалы жирных кислот.
Основные жирные
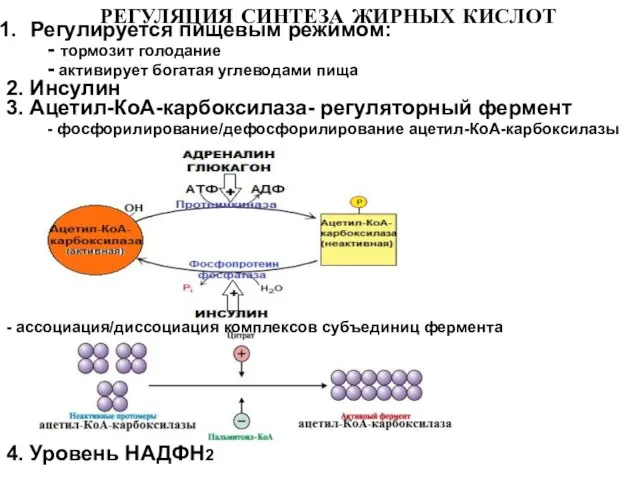
Слайд 19РЕГУЛЯЦИЯ СИНТЕЗА ЖИРНЫХ КИСЛОТ
Регулируется пищевым режимом:
- тормозит голодание
- активирует
РЕГУЛЯЦИЯ СИНТЕЗА ЖИРНЫХ КИСЛОТ
Регулируется пищевым режимом:
- тормозит голодание
- активирует
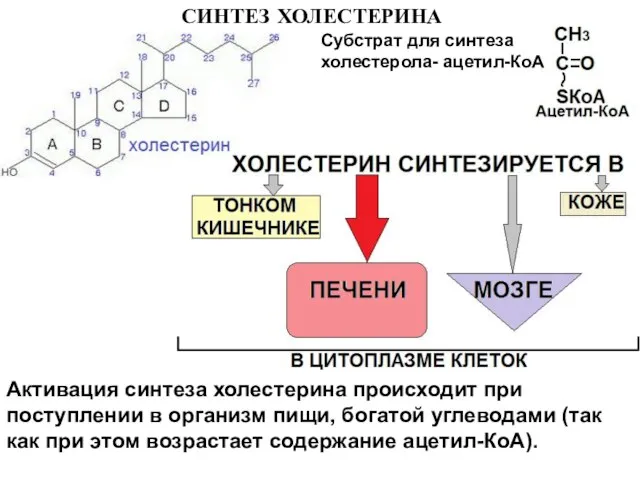
Слайд 20СИНТЕЗ ХОЛЕСТЕРИНА
Субстрат для синтеза холестерола- ацетил-КоА
Активация синтеза холестерина происходит при поступлении в
СИНТЕЗ ХОЛЕСТЕРИНА
Субстрат для синтеза холестерола- ацетил-КоА
Активация синтеза холестерина происходит при поступлении в
Слайд 21СИНТЕЗ ХОЛЕСТЕРИНА
ОСНОВНЫЕ СТАДИИ СИНТЕЗА ХОЛЕСТЕРИНА:
• синтез мевалоната (С6)
• синтез сквалена из мевалоната
СИНТЕЗ ХОЛЕСТЕРИНА
ОСНОВНЫЕ СТАДИИ СИНТЕЗА ХОЛЕСТЕРИНА:
• синтез мевалоната (С6)
• синтез сквалена из мевалоната

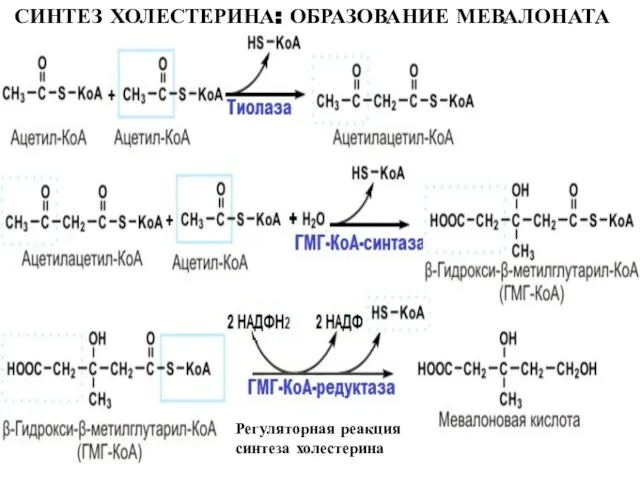
Слайд 22СИНТЕЗ ХОЛЕСТЕРИНА: ОБРАЗОВАНИЕ МЕВАЛОНАТА
Регуляторная реакция синтеза холестерина
СИНТЕЗ ХОЛЕСТЕРИНА: ОБРАЗОВАНИЕ МЕВАЛОНАТА
Регуляторная реакция синтеза холестерина
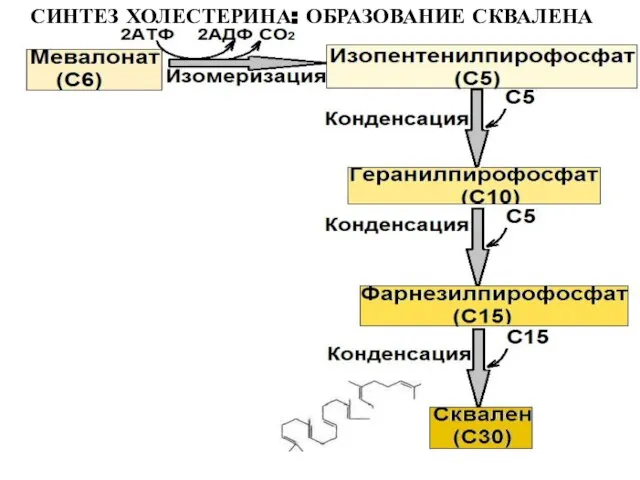
Слайд 23СИНТЕЗ ХОЛЕСТЕРИНА: ОБРАЗОВАНИЕ СКВАЛЕНА
СИНТЕЗ ХОЛЕСТЕРИНА: ОБРАЗОВАНИЕ СКВАЛЕНА
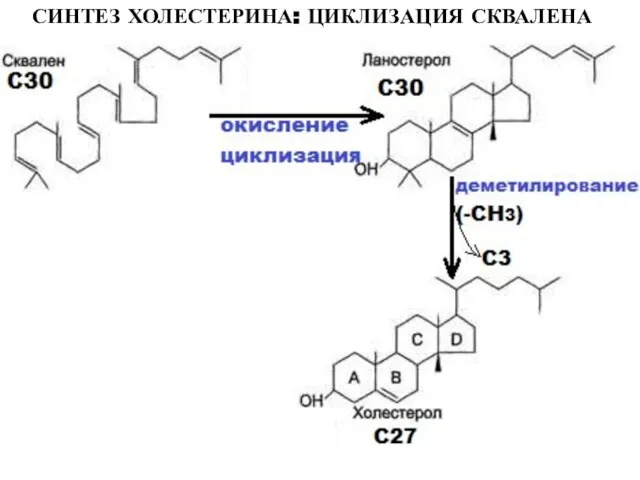
Слайд 24СИНТЕЗ ХОЛЕСТЕРИНА: ЦИКЛИЗАЦИЯ СКВАЛЕНА
СИНТЕЗ ХОЛЕСТЕРИНА: ЦИКЛИЗАЦИЯ СКВАЛЕНА
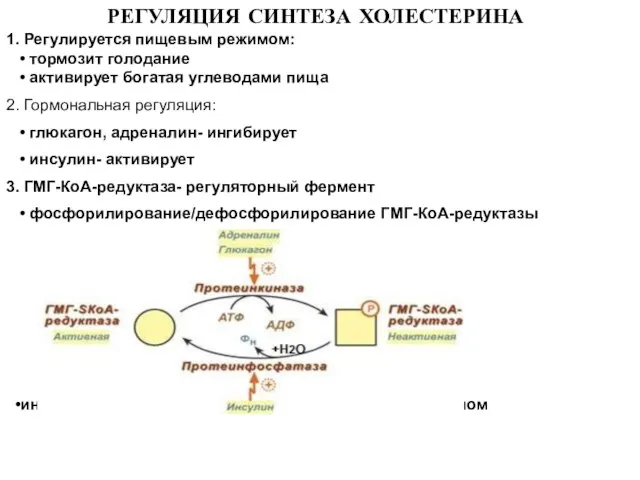
Слайд 25 РЕГУЛЯЦИЯ СИНТЕЗА ХОЛЕСТЕРИНА
1. Регулируется пищевым режимом:
• тормозит голодание
•
РЕГУЛЯЦИЯ СИНТЕЗА ХОЛЕСТЕРИНА
1. Регулируется пищевым режимом:
• тормозит голодание
•
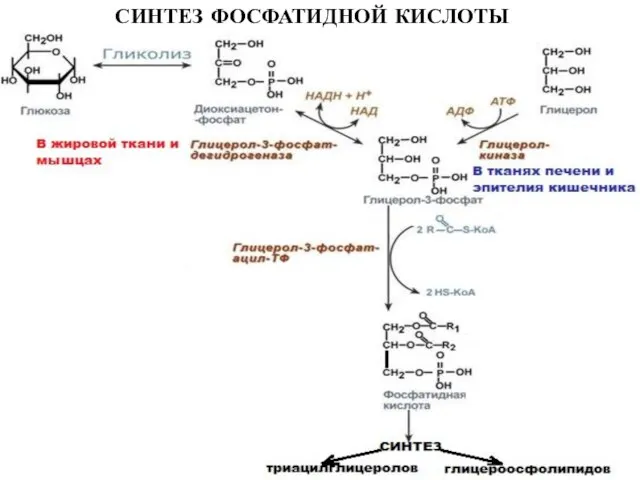
Слайд 26СИНТЕЗ ФОСФАТИДНОЙ КИСЛОТЫ
СИНТЕЗ ФОСФАТИДНОЙ КИСЛОТЫ
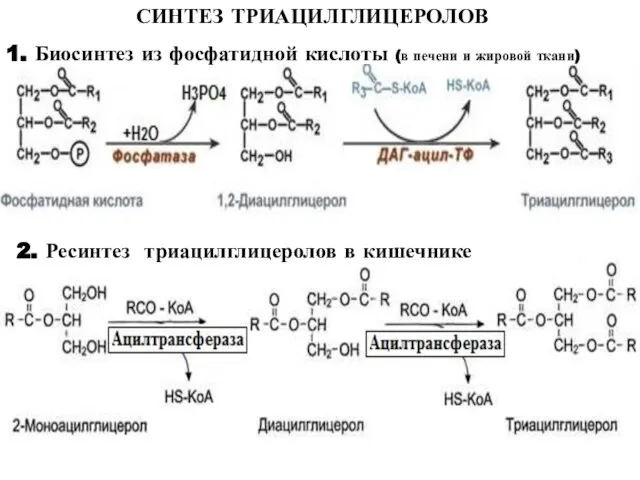
Слайд 27СИНТЕЗ ТРИАЦИЛГЛИЦЕРОЛОВ
1. Биосинтез из фосфатидной кислоты (в печени и жировой ткани)
2. Ресинтез
СИНТЕЗ ТРИАЦИЛГЛИЦЕРОЛОВ
1. Биосинтез из фосфатидной кислоты (в печени и жировой ткани)
2. Ресинтез
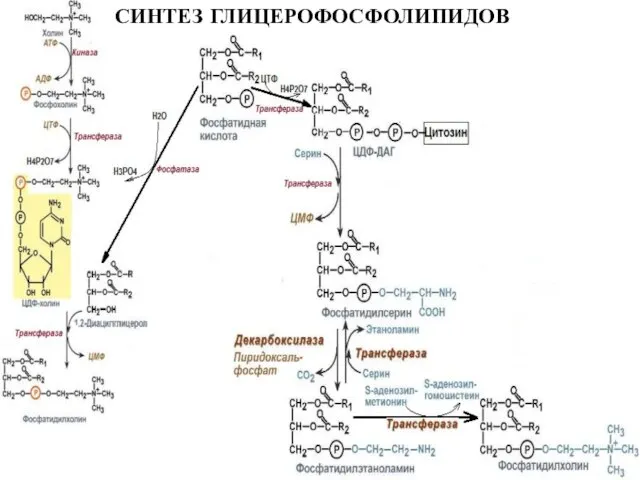
Слайд 28СИНТЕЗ ГЛИЦЕРОФОСФОЛИПИДОВ
СИНТЕЗ ГЛИЦЕРОФОСФОЛИПИДОВ
 Голосеменные
Голосеменные Презентация на тему Размножение споровых растений
Презентация на тему Размножение споровых растений  Функции крови
Функции крови Роль селекции в лесном хозяйстве. Лекция 1
Роль селекции в лесном хозяйстве. Лекция 1 Строение нервной системы
Строение нервной системы Условные и безусловные рефлексы
Условные и безусловные рефлексы Транскрипция. Молекулярная биотехнология. Часть 2
Транскрипция. Молекулярная биотехнология. Часть 2 Микроскоп. Части микроскопа
Микроскоп. Части микроскопа Презентация Растения
Презентация Растения Главные направления эволюции
Главные направления эволюции Стрекозы
Стрекозы Одомашнивание домашних животных (собак)
Одомашнивание домашних животных (собак) Значение пищеварения в жизнедеятельности организма. Процессы пищеварения. Строение пищеварительной системы
Значение пищеварения в жизнедеятельности организма. Процессы пищеварения. Строение пищеварительной системы Физиология человека: динамические показатели нервных процессов
Физиология человека: динамические показатели нервных процессов Недостатки рефракции глаза и их коррекция
Недостатки рефракции глаза и их коррекция Что такое ферменты
Что такое ферменты Цепи питания
Цепи питания Основы морфологии и методы изучения микроорганизмов
Основы морфологии и методы изучения микроорганизмов Фотосинтез, его значение
Фотосинтез, его значение Дәнді дақылдар
Дәнді дақылдар Бурый медведь
Бурый медведь Оплодотворение и его значение. 9 класс
Оплодотворение и его значение. 9 класс Распределение микроорганизмов в окружающей среде
Распределение микроорганизмов в окружающей среде Общие вопросы анатомии и физиологии опорно-двигательного аппарата
Общие вопросы анатомии и физиологии опорно-двигательного аппарата Органы. Системы органов
Органы. Системы органов Биологический мониторинг состояния растительного и животного мира Бурзянского района
Биологический мониторинг состояния растительного и животного мира Бурзянского района Класс рыбы
Класс рыбы Неполное доминирование
Неполное доминирование