- Обмен_АМИНОК_2020_МУДЛ (1)

Содержание
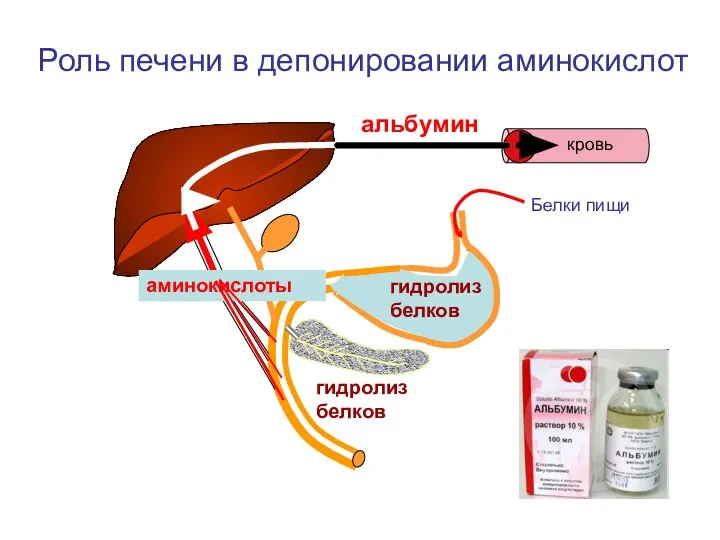
- 2. аминокислоты альбумин Роль печени в депонировании аминокислот гидролиз белков Белки пищи кровь гидролиз белков
- 3. Превращения аминокислот в клетке
- 4. Реакции поликонденсации (биосинтез белков) Реакции трансаминирования Реакции декарбоксилирования Реакции окислительного дезаминирования Все превращения аминокислот можно суммировать
- 5. Биосинтез белка
- 6. Стадии синтеза белка Образование инициирующего комплекса; Элонгация (удлинение полипептидной цепи); Терминация (завершение синтеза); Процессинг (окончательное достраивание
- 7. Для синтеза белка нужна и-РНК кодон кодон кодон кодон кодон -А-Ц-Г-А-У-Г-А-У-Ц-Г-А-У-А-Ц-Г - и-РНК
- 8. Для синтеза белка нужна рибосома Большая субъединица рибосомы Малая субъединица рибосомы
- 9. Для синтеза белка нужны т-РНК ЦЦА -аминокислота У-А-Ц антикодон в т-РНК, соответствует кодону АУГ на И-РНК
- 10. Все аминокислоты кодируются в виде троек нуклеотидов (кодонов или триплетов). Одна аминокислота может кодироваться одним триплетом
- 11. РНК - аминокислотный код
- 12. Активация аминокислот АТФ + НООС-СН-NН2 R O АМФ - C - CH - NH2 R аминоациладенилат
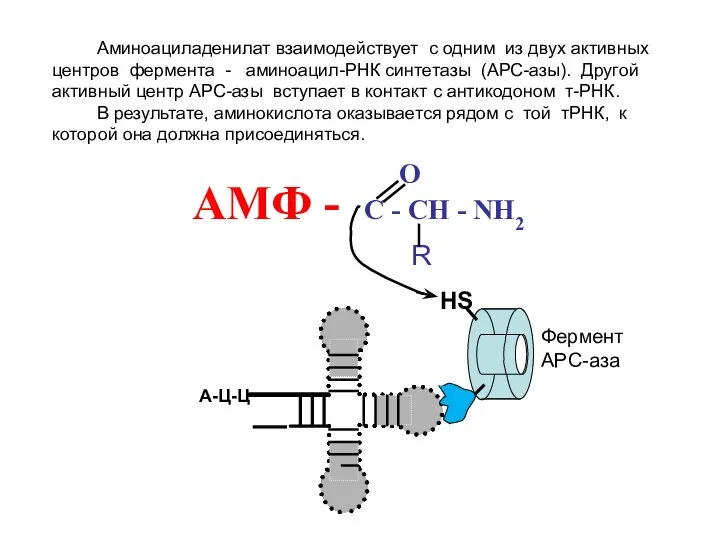
- 13. НS O АМФ - C - CH - NH2 R А-Ц-Ц Фермент АРС-аза Аминоациладенилат взаимодействует с
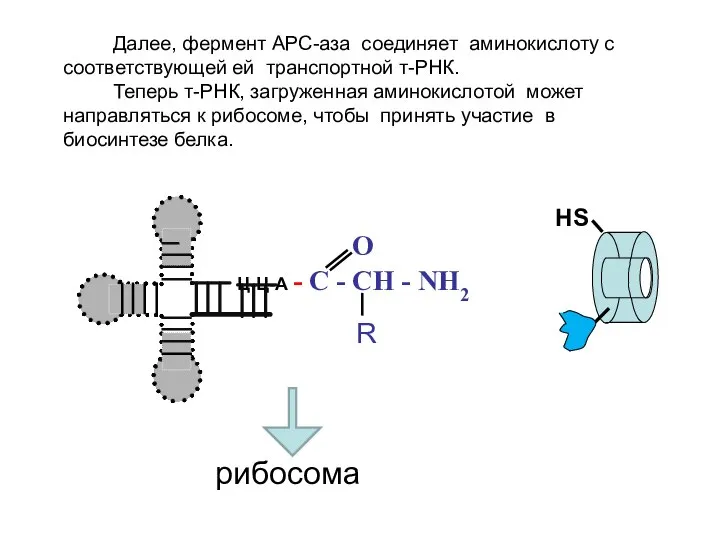
- 14. НS Ц-Ц-А O - C - CH - NH2 R Далее, фермент АРС-аза соединяет аминокислоту с
- 15. Образование инициирующего комплекса.
- 16. кодон кодон кодон кодон кодон ЦЦА -аминокислота т-РНК -А-Ц-Г-А-У-Г-А-У-Ц-Г-А-У-А-Ц-Г - и-РНК У-А-Ц Инициирующий комплекс состоит из
- 17. ЦЦА ЦЦА и-РНК Вторая т-РНК занимает место в аминоацильном центре (А) рибосомы согласно правилу комплементарности антикодона
- 18. ЦЦА ЦЦА и-РНК Оказавшись рядом с первой т-РНК, вторая т-РНК переносит к своей аминокислоте аминокислоту первой
- 19. СAP и-РНК ЦЦА ЦЦА После этого рибосома передвигается на один кодон в сторону. При этом в
- 20. и-РНК ЦЦА ЦЦА К освободившемуся месту на аминоацильном центре рибосомы устремляется третья т-РНК. Согласно кодону она
- 21. Синтез белка продолжается до тех пор, пока в аминоацильный центр не заходит особый кодон и-РНК, сигнализирующий
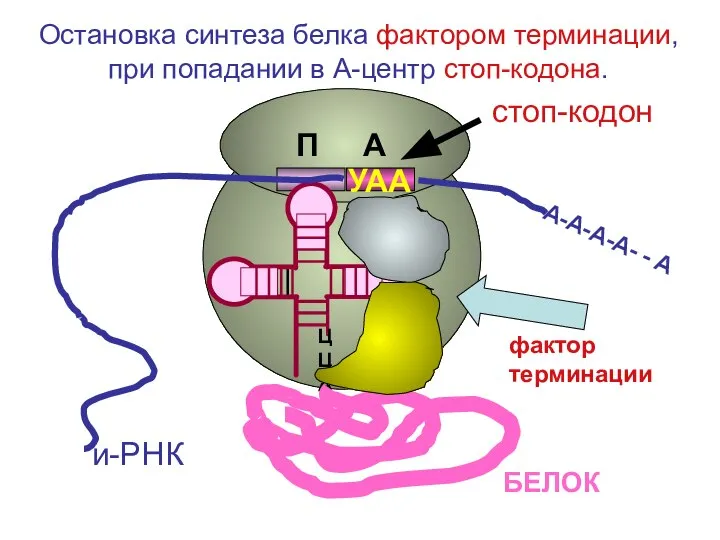
- 22. ЦЦА УАА А-А-А-А- - А и-РНК фактор терминации БЕЛОК стоп-кодон Остановка синтеза белка фактором терминации, при
- 23. Биосинтез белка (мультик)
- 24. ЦЦА Образование инициирующего комплекса и-РНК СAP П А аминокислота
- 25. ЦЦА Сборка рибосомы и-РНК СAP Большая субъединица рибосомы П А
- 26. ЦЦА Сборка рибосомы и-РНК СAP П А
- 27. ЦЦА Сборка рибосомы и-РНК СAP П А
- 28. ЦЦА Сборка рибосомы и-РНК СAP П А
- 29. ЦЦА Сборка рибосомы и-РНК СAP П А
- 30. ЦЦА ЦЦА и-РНК Начало синтеза белка П А
- 31. ЦЦА и-РНК Начало синтеза белка П А
- 32. ЦЦА и-РНК Начало синтеза белка П А
- 33. ЦЦА и-РНК Начало синтеза белка П А
- 34. ЦЦА и-РНК Начало синтеза белка П А
- 35. ЦЦА ЦЦА и-РНК Перенос первой аминокислоты на вторую П А
- 36. ЦЦА ЦЦА и-РНК Перенос первой аминокислоты на вторую П А
- 37. ЦЦА ЦЦА и-РНК Образование дипептида П А
- 38. ЦЦА ЦЦА и-РНК П А Перемещение рибосомы вдоль и-РНК на один триплет (кодон)
- 39. ЦЦА ЦЦА и-РНК П А Перемещение рибосомы вдоль и-РНК на один триплет (кодон)
- 40. и-РНК П А Перемещение рибосомы вдоль и-РНК на один триплет (кодон)
- 41. СAP и-РНК ЦЦА Удаление 1-й тРНК
- 42. СAP и-РНК ЦЦА Удаление 1-й тРНК
- 43. СAP и-РНК ЦЦА ЦЦА Удаление 1-й тРНК
- 44. и-РНК ЦЦА ЦЦА Повторение цикла элонгации. Присоединение третьей т-РНК П А
- 45. и-РНК ЦЦА Повторение цикла элонгации. Присоединение третьей т-РНК П А
- 46. и-РНК ЦЦА Повторение цикла элонгации. Присоединение третьей т-РНК П А
- 47. и-РНК ЦЦА Повторение цикла элонгации. Присоединение третьей т-РНК П А
- 48. ЦЦА ЦЦА и-РНК П А Перенос дипептида на третью аминокислоту
- 49. ЦЦА ЦЦА и-РНК П А Перенос дипептида на третью аминокислоту
- 50. ЦЦА ЦЦА и-РНК П А Перенос дипептида на третью аминокислоту
- 51. ЦЦА ЦЦА и-РНК П А Образование трипептида
- 52. ЦЦА ЦЦА и-РНК П А Перемещение рибосомы вдоль и-РНК на один триплет (кодон)
- 53. ЦЦА ЦЦА и-РНК П А Перемещение рибосомы вдоль и-РНК на один триплет (кодон)
- 54. ЦЦА ЦЦА и-РНК П А Перемещение рибосомы вдоль и-РНК на один триплет (кодон)
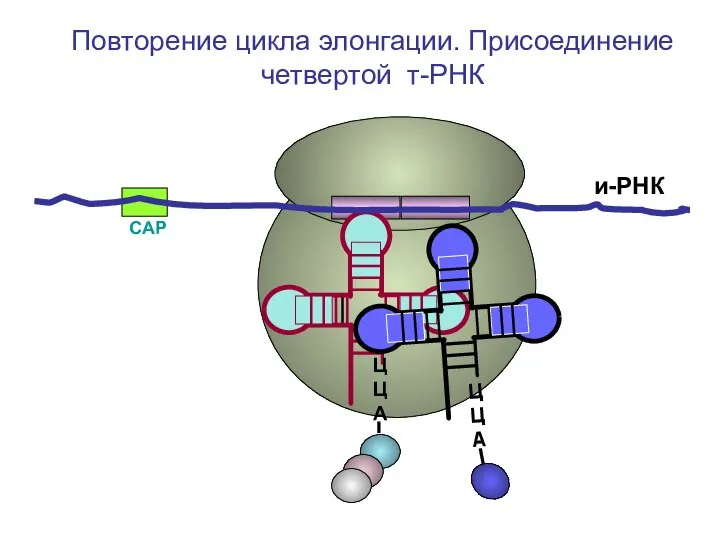
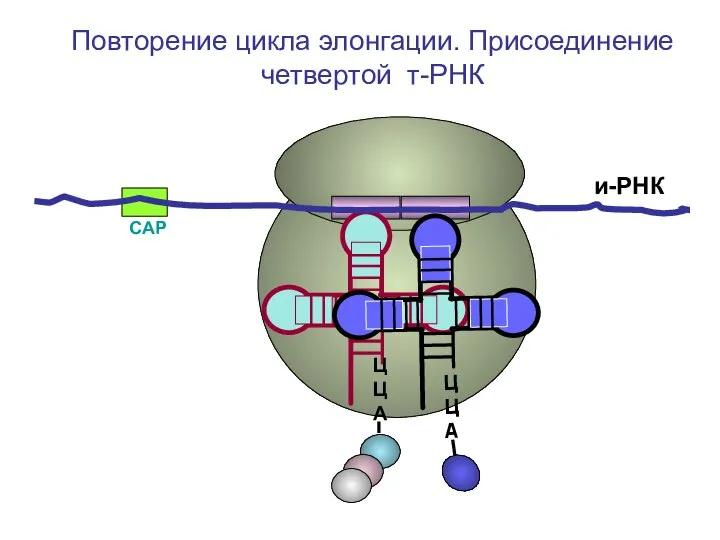
- 55. СAP и-РНК ЦЦА ЦЦА ЦЦА Повторение цикла элонгации. Присоединение четвертой т-РНК
- 56. СAP и-РНК ЦЦА ЦЦА Повторение цикла элонгации. Присоединение четвертой т-РНК
- 57. СAP и-РНК ЦЦА Повторение цикла элонгации. Присоединение четвертой т-РНК
- 58. СAP и-РНК ЦЦА Повторение цикла элонгации. Присоединение четвертой т-РНК
- 59. СAP и-РНК ЦЦА Повторение цикла элонгации. Присоединение четвертой т-РНК
- 60. СAP и-РНК ЦЦА Повторение цикла элонгации. Присоединение четвертой т-РНК
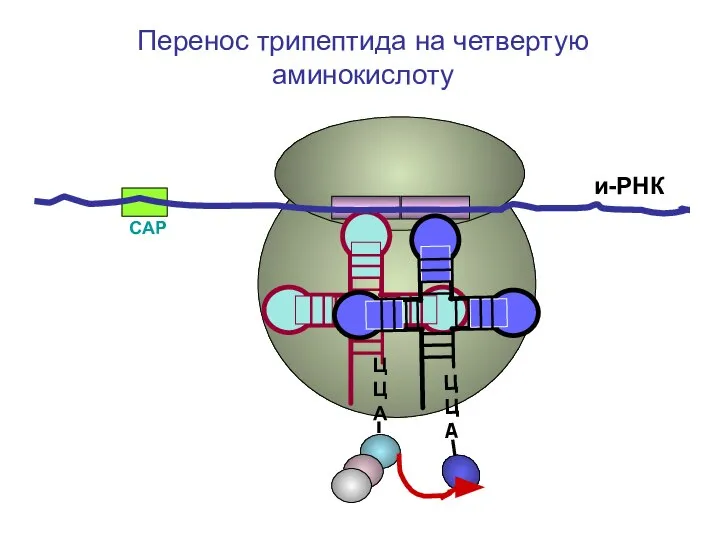
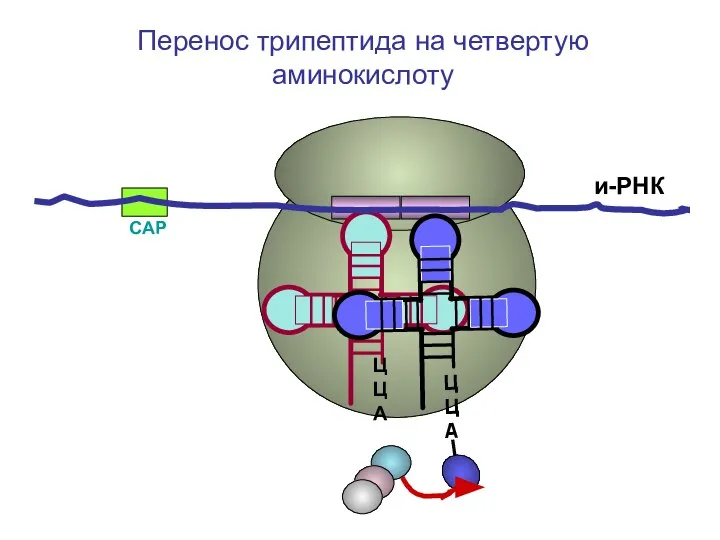
- 61. СAP и-РНК ЦЦА Перенос трипептида на четвертую аминокислоту
- 62. СAP и-РНК ЦЦА Перенос трипептида на четвертую аминокислоту
- 63. СAP и-РНК ЦЦА Перенос трипептида на четвертую аминокислоту
- 64. СAP и-РНК ЦЦА Перенос трипептида на четвертую аминокислоту
- 65. СAP и-РНК ЦЦА Перемещение рибосомы вдоль и-РНК на один триплет (кодон)
- 66. СAP и-РНК Перемещение рибосомы вдоль и-РНК на один триплет (кодон)
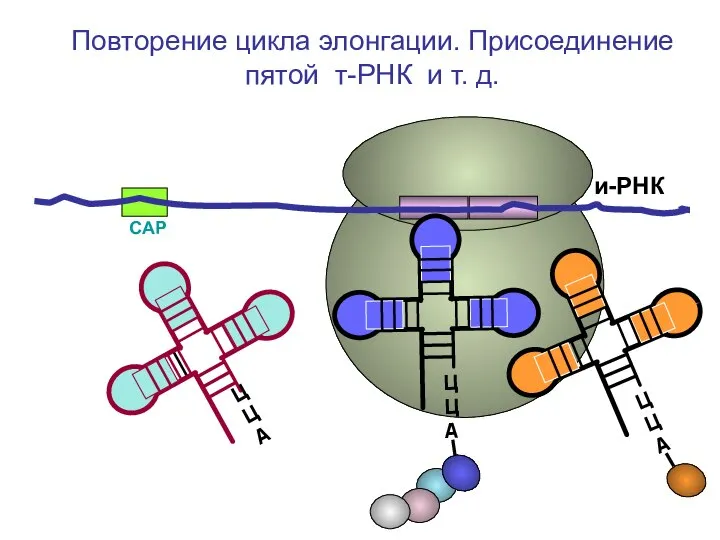
- 67. СAP и-РНК ЦЦА Повторение цикла элонгации. Присоединение пятой т-РНК и т. д.
- 68. Стадия терминации
- 69. Образовавшийся белок, потоком жидкости в эндоплазматическом ретикулуме (состоящим из трубочек), направляется в Аппарат Гольджи. В этом
- 70. В аппарат Гольджи Многие белки требуют достраивания.
- 71. Присоединение коферментов Присоединение углеводов Обьединение с металлами Фосфорили-рование Удаление “лишних” аминокислот или участков белка Образование четвер-тичной
- 72. Регуляция обмена белков
- 73. Гормональная регуляция скорости синтеза белка Соматотропин, половые гормоны (индукторы транскрипции и биосинтеза белка); 2. Тироксин (активатор
- 74. Влияние некоторых факторов на концентрацию и состав белков организма Доступность в белковой диете; 2. Наличие полноценных
- 75. Влияние антибиотиков на биосинтез белков
- 76. Трансамини-рование аминокислот
- 77. Поступающий с пищей набор аминокислот, редко соответствует их требуемому соотношению для использования клетками. Часто одних аминокислот
- 78. СН3 СООН СН3 СООН СНNH2 + C=О С=О + СНNH2 СООН СН2 СООН СН2 СН2 СН2
- 79. С СН2ОН НО Н3С N Н О пиридоксаль Витамин В6 (неактивная форма витамина)
- 80. С СН2О-РО3Н2 НО Н3С N Н О Фосфопиридоксаль (активная форма витамина)
- 81. Н О С СН2О-РО3Н2 НО Н3С N СН2-NH2 СН2О-РО3Н2 НО Н3С N пиридоксамин-фосфат Витамин как акцептор
- 82. Н О С СН2О-РО3Н2 НО Н3С N СН2-NH2 СН2О-РО3Н2 НО Н3С N Витамин как донор аминогруппы
- 83. В клинической практике активность АлАТ или АсАТ часто измеряют в сыворотке крови для обнаружения цитолиза клеток
- 84. Дни болезни Активность ферментов 1 2 3 4 5 6 7 8 9 10 11 12
- 85. Декарбоксилирование аминокислот (образование биологически-активных аминов)
- 86. Часть аминокислот используется организмом для получения биологически активных молекул, которые участвуют в регуляции обменных процессов. Путем
- 87. H2N – CН – CООН H2N – CН2 + СО2 R R Схема декарбоксилирования аминокислот Биологически
- 88. N -CH2-CH-COOH NH2 NH N -CH2-CH2 NH2 NH декарбоксилаза СО2 Образование гистамина. Участвует в воспалительных реакциях.
- 89. -CH2-CH-COOH NH2 NH НО- -CH2-CH2 NH2 NH НО 5-гидрокситриптамин (серотонин) декарбоксилаза СО2 Гидроксилирование и декарбоксилирование триптофана
- 90. СOOH CH-NH2 CH2 CH2 COOH CH2-NH2 CH2 CH2 COOH декарбоксилаза CO2 глутаминовая кислота гамма-аминомасляная кислота (ГАМК)
- 91. CH2-CH-COOH NH2 ОН ОН CH2-CH2 NH2 ОН ОН СО2 декарбоксилаза Гидроксилирование и декарбоксилирование фенилаланина, с образованием
- 92. CH2-CH2 NH2 ОН ОН Синтез норадреналина из дофамина. CH-CH2 ОН NH2 ОН ОН НАДФН2; О2; р450
- 93. CH-CH2 ОН NH2 ОН ОН норадреналин CH-CH2 ОН NH – СН3 ОН ОН адреналин метил-фолиевая кислота;
- 94. Биологически активные амины, используемые как лекарственные средства. Гистамин Триптамин Серотонин Гамма-аминомасляная кислота (ГАМК) Дофамин Норадреналин Адреналин
- 95. Использование аминокислот в качестве исходных субстратов для выработки энергии с помощью окислительного дезаминирования.
- 96. Иногда в клетках возникает необходимость в более интенсивном образовании энергии. При этом, кроме углеводов и липидов,
- 97. Реакция окислительного дезаминирования глутаминовой кислоты СООН СН2 СН2 CH-NH2 COOH HАД HАДН2 +Н2О СООН СН2 СН2
- 98. В результате окислительного дезаминирования глутаминовой, или аспарагиновой кислот образуется одна молекула НАДН2 , которая используется митохондриями
- 99. NH3 дезаминирова-ние аминокислот дезаминирова-ние нуклеотидов окисление аминов дезаминирование аминосахаров метаболические источники аммиака в организме
- 100. синтез мочевины
- 101. • • Местом обезвреживания аммиака в организме ( 20 г в сутки) является печень
- 102. В первой реакции аммиак с помощью угольной кислоты и энергии АТФ преобразуется в карбамоилфосфат. глутаминовая кислота
- 103. + NH2 CH2 CH2 CH2 CHNH2 COOH орнитин трансфераза H3PO4 NH2 C=O NH CH2 CH2 CH2
- 104. NH2 C=NH NH CH2 CH2 CH2 CHNH2 COOH фумаровая к-та + АТФ аргининсукцинатсинтетаза АМФ H4P2O7 аргинин
- 105. NH2 CH2 CH2 CH2 CHNH2 COOH C=O NH2 NH2 мочевина NH2 C=NH NH CH2 CH2 CH2
- 106. NH3 + CO2 + 2 АТФ NH2-C-O~P=O O OH OH орнитин цитруллин аспарагино-вая кислота аргинин фумаро-вая
- 107. Содержание мочевины в крови 2,5 – 8,3 ммоль/л сыворотки
- 108. Диагностическая значимость определения концентрации мочевины в крови. Понижение ниже нормы наблюдается при: тяжелом заболевании печени; длительном
- 109. Вспомогательный, быстрый механизм связывания аммиака внутри клеток
- 110. Этот пример удаления аммиака в клетках происходит в тех органах, где мочевина не может синтезироваться. Например,
- 111. Образование глутамина СООН СН2 СН2 CH-NH2 COOH NH3 + СОNH2 СН2 СН2 CH-NH2 COOH АТФ +
- 112. С током крови глутамин поступает в печень или в почки. В печени происходит гидролиз глутамина, образуется
- 113. СООН СН2 СН2 CH-NH2 COOH NH3 СОNH2 СН2 СН2 CH-NH2 COOH + H2O глутамино-вая кислота глутамин
- 115. Скачать презентацию
Слайд 2аминокислоты
альбумин
Роль печени в депонировании аминокислот
гидролиз белков
Белки пищи
кровь
гидролиз белков
аминокислоты
альбумин
Роль печени в депонировании аминокислот
гидролиз белков
Белки пищи
кровь
гидролиз белков
Слайд 3Превращения аминокислот в клетке
Превращения аминокислот в клетке

Слайд 4Реакции поликонденсации (биосинтез белков)
Реакции трансаминирования
Реакции декарбоксилирования
Реакции окислительного дезаминирования
Все превращения аминокислот можно суммировать
Реакции поликонденсации (биосинтез белков)
Реакции трансаминирования
Реакции декарбоксилирования
Реакции окислительного дезаминирования
Все превращения аминокислот можно суммировать

Слайд 5Биосинтез белка
Биосинтез белка

Слайд 6 Стадии синтеза белка
Образование инициирующего комплекса;
Элонгация (удлинение полипептидной цепи);
Терминация (завершение синтеза);
Процессинг (окончательное
Стадии синтеза белка
Образование инициирующего комплекса;
Элонгация (удлинение полипептидной цепи);
Терминация (завершение синтеза);
Процессинг (окончательное

Слайд 7Для синтеза белка нужна и-РНК
кодон кодон кодон кодон кодон
-А-Ц-Г-А-У-Г-А-У-Ц-Г-А-У-А-Ц-Г -
Для синтеза белка нужна и-РНК
кодон кодон кодон кодон кодон
-А-Ц-Г-А-У-Г-А-У-Ц-Г-А-У-А-Ц-Г -

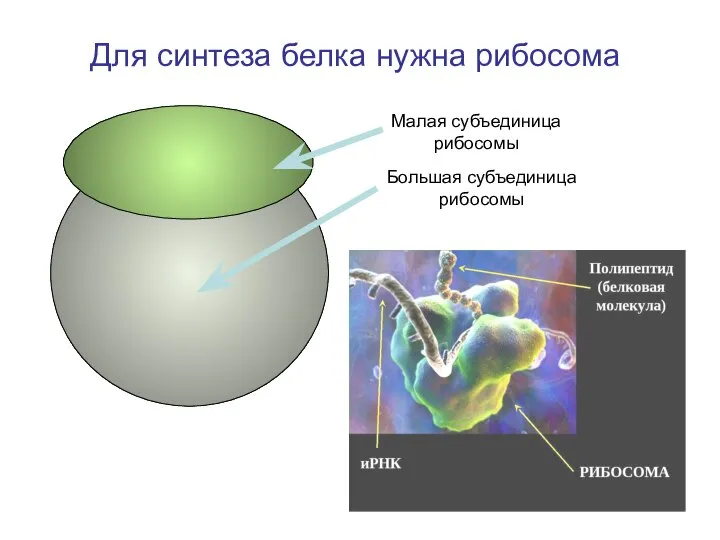
Слайд 8Для синтеза белка нужна рибосома
Большая субъединица рибосомы
Малая субъединица рибосомы
Для синтеза белка нужна рибосома
Большая субъединица рибосомы
Малая субъединица рибосомы
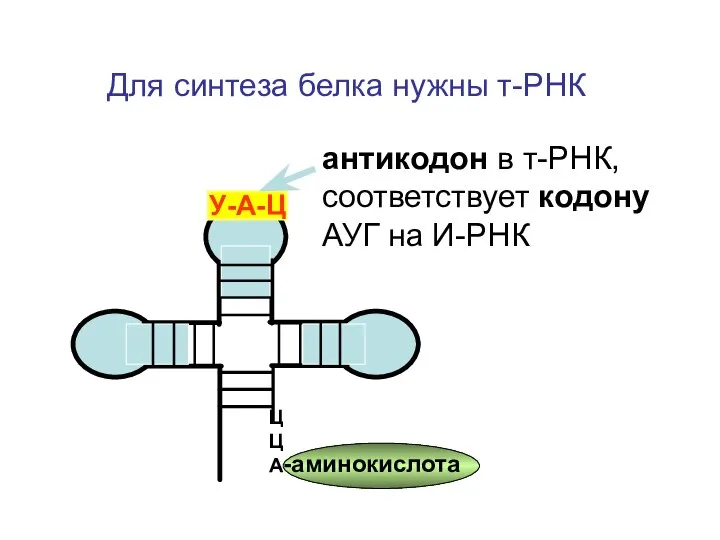
Слайд 9Для синтеза белка нужны т-РНК
ЦЦА
-аминокислота
У-А-Ц
антикодон в т-РНК, соответствует кодону АУГ
Для синтеза белка нужны т-РНК
ЦЦА
-аминокислота
У-А-Ц
антикодон в т-РНК, соответствует кодону АУГ
Слайд 10 Все аминокислоты кодируются в виде троек нуклеотидов (кодонов или триплетов). Одна
Все аминокислоты кодируются в виде троек нуклеотидов (кодонов или триплетов). Одна

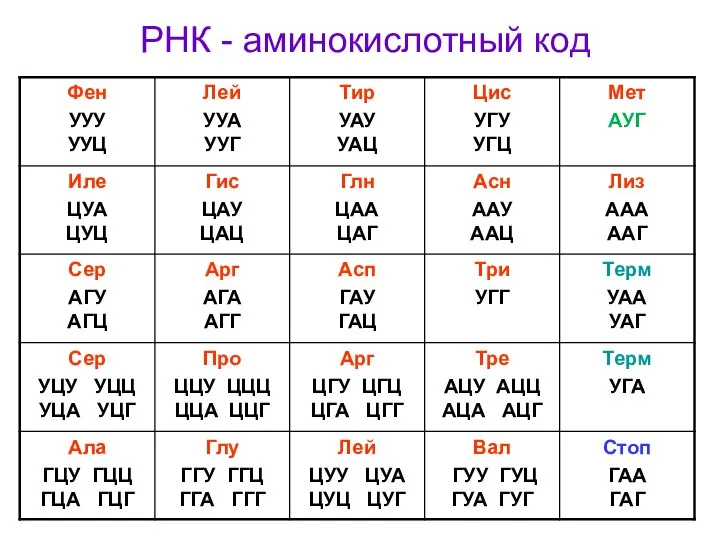
Слайд 11РНК - аминокислотный код
РНК - аминокислотный код
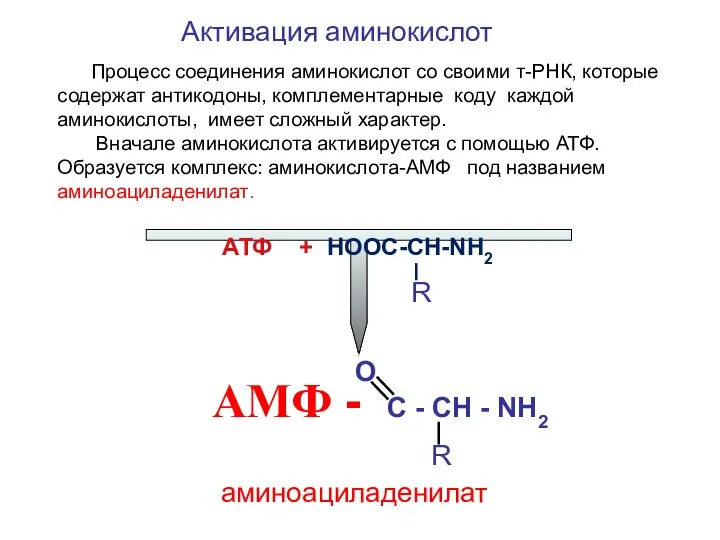
Слайд 12Активация аминокислот
АТФ + НООС-СН-NН2
R
O
АМФ - C
Активация аминокислот
АТФ + НООС-СН-NН2
R
O
АМФ - C
Слайд 13 НS
O
АМФ - C - CH -
НS
O
АМФ - C - CH -
Слайд 14 НS
Ц-Ц-А
O
- C - CH - NH2
R
Далее, фермент
НS
Ц-Ц-А
O
- C - CH - NH2
R
Далее, фермент
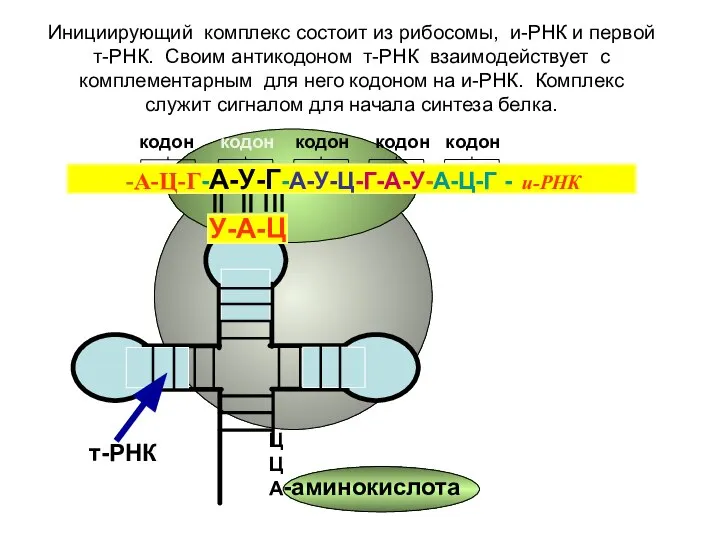
Слайд 15Образование инициирующего комплекса.
Образование инициирующего комплекса.

Слайд 16 кодон кодон кодон кодон кодон
ЦЦА
-аминокислота
т-РНК
-А-Ц-Г-А-У-Г-А-У-Ц-Г-А-У-А-Ц-Г - и-РНК
У-А-Ц
Инициирующий
кодон кодон кодон кодон кодон
ЦЦА
-аминокислота
т-РНК
-А-Ц-Г-А-У-Г-А-У-Ц-Г-А-У-А-Ц-Г - и-РНК
У-А-Ц
Инициирующий
Слайд 17ЦЦА
ЦЦА
и-РНК
Вторая т-РНК занимает место в аминоацильном центре
ЦЦА
ЦЦА
и-РНК
Вторая т-РНК занимает место в аминоацильном центре
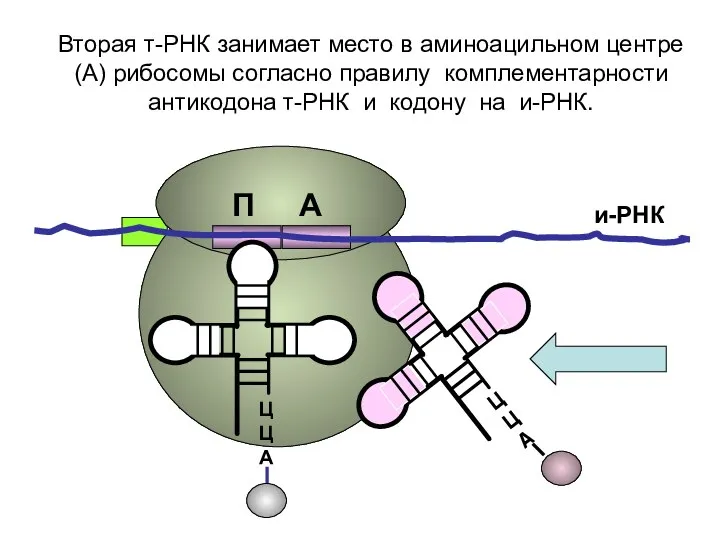
Слайд 18ЦЦА
ЦЦА
и-РНК
Оказавшись рядом с первой т-РНК, вторая т-РНК переносит
ЦЦА
ЦЦА
и-РНК
Оказавшись рядом с первой т-РНК, вторая т-РНК переносит
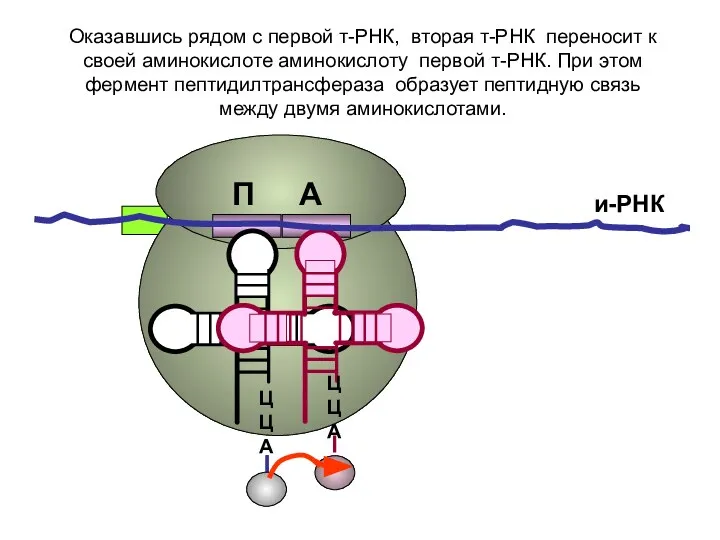
Слайд 19СAP
и-РНК
ЦЦА
ЦЦА
После этого рибосома передвигается на один
СAP
и-РНК
ЦЦА
ЦЦА
После этого рибосома передвигается на один
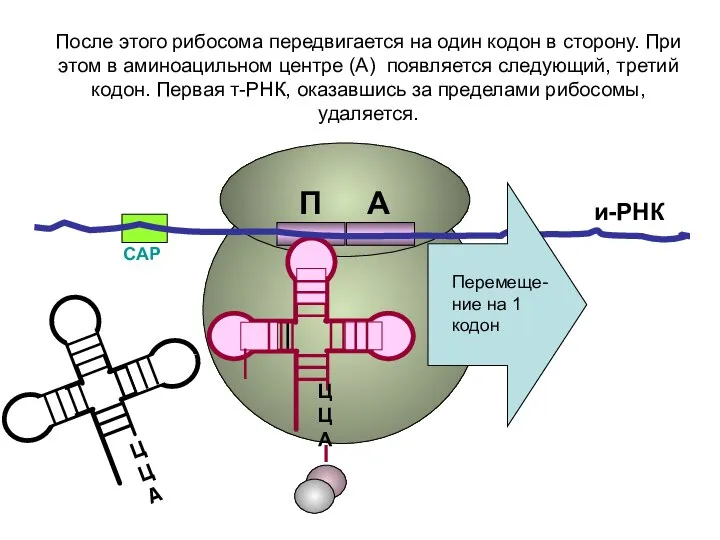
Слайд 20и-РНК
ЦЦА
ЦЦА
К освободившемуся месту на аминоацильном центре рибосомы устремляется третья т-РНК. Согласно
и-РНК
ЦЦА
ЦЦА
К освободившемуся месту на аминоацильном центре рибосомы устремляется третья т-РНК. Согласно
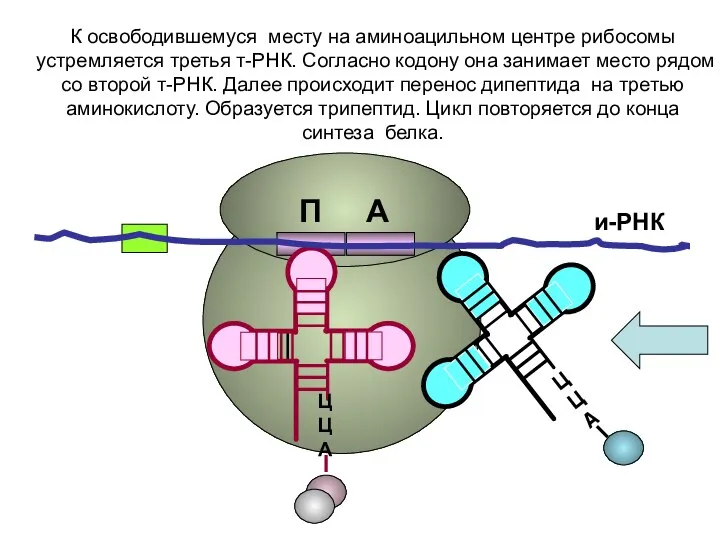
Слайд 21 Синтез белка продолжается до тех пор, пока в аминоацильный центр не
Синтез белка продолжается до тех пор, пока в аминоацильный центр не

Слайд 22ЦЦА
УАА
А-А-А-А- - А
и-РНК
фактор терминации
БЕЛОК
стоп-кодон
Остановка синтеза белка фактором терминации, при попадании
ЦЦА
УАА
А-А-А-А- - А
и-РНК
фактор терминации
БЕЛОК
стоп-кодон
Остановка синтеза белка фактором терминации, при попадании
Слайд 23Биосинтез белка
(мультик)
Биосинтез белка
(мультик)

Слайд 24ЦЦА
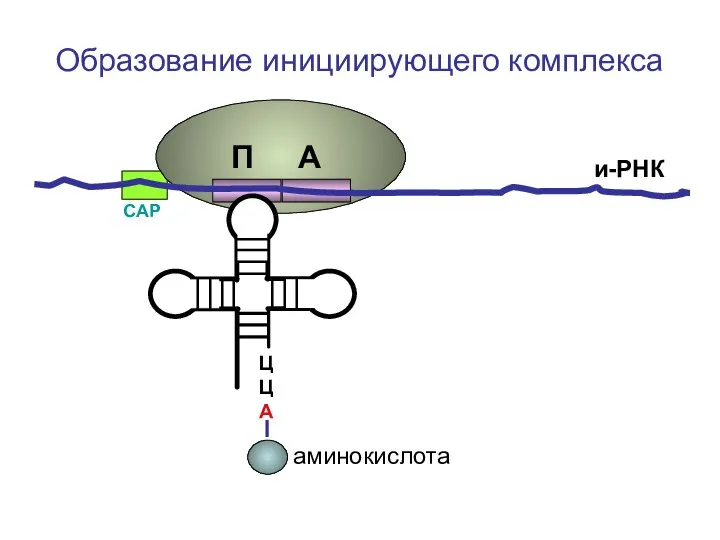
Образование инициирующего комплекса
и-РНК
СAP
П А
аминокислота
ЦЦА
Образование инициирующего комплекса
и-РНК
СAP
П А
аминокислота
Слайд 25ЦЦА
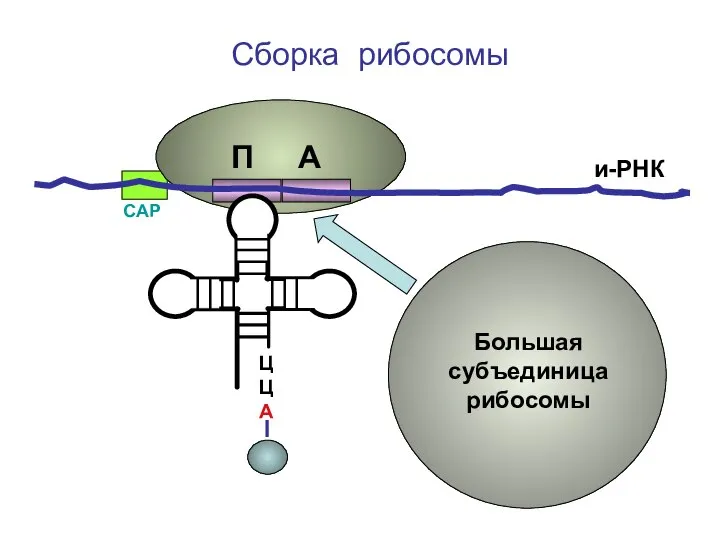
Сборка рибосомы
и-РНК
СAP
Большая субъединица рибосомы
П А
ЦЦА
Сборка рибосомы
и-РНК
СAP
Большая субъединица рибосомы
П А
Слайд 26ЦЦА
Сборка рибосомы
и-РНК
СAP
П А
ЦЦА
Сборка рибосомы
и-РНК
СAP
П А
Слайд 27ЦЦА
Сборка рибосомы
и-РНК
СAP
П А
ЦЦА
Сборка рибосомы
и-РНК
СAP
П А
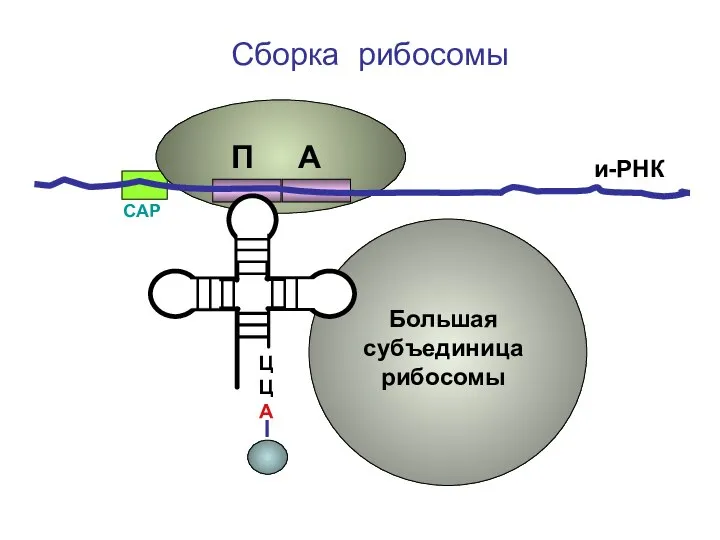
Слайд 28ЦЦА
Сборка рибосомы
и-РНК
СAP
П А
ЦЦА
Сборка рибосомы
и-РНК
СAP
П А
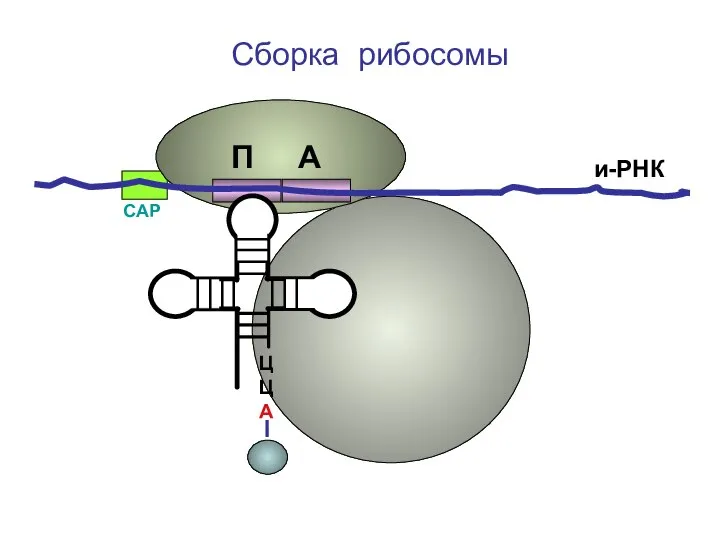
Слайд 29ЦЦА
Сборка рибосомы
и-РНК
СAP
П А
ЦЦА
Сборка рибосомы
и-РНК
СAP
П А
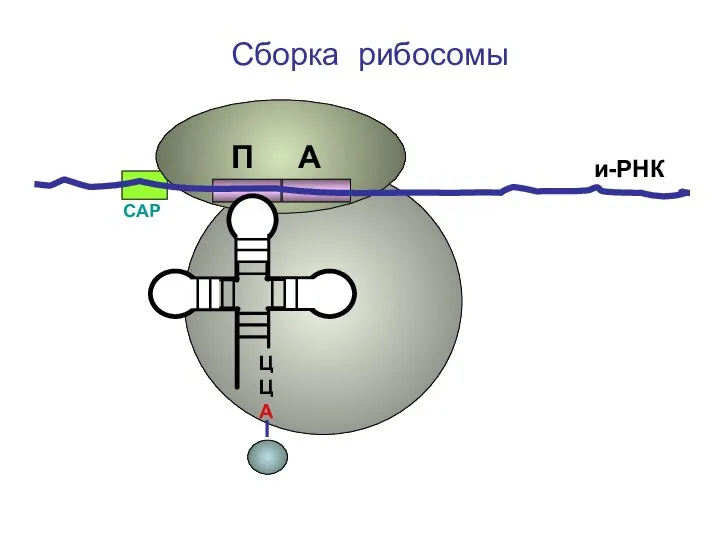
Слайд 30ЦЦА
ЦЦА
и-РНК
Начало синтеза белка
П А
ЦЦА
ЦЦА
и-РНК
Начало синтеза белка
П А
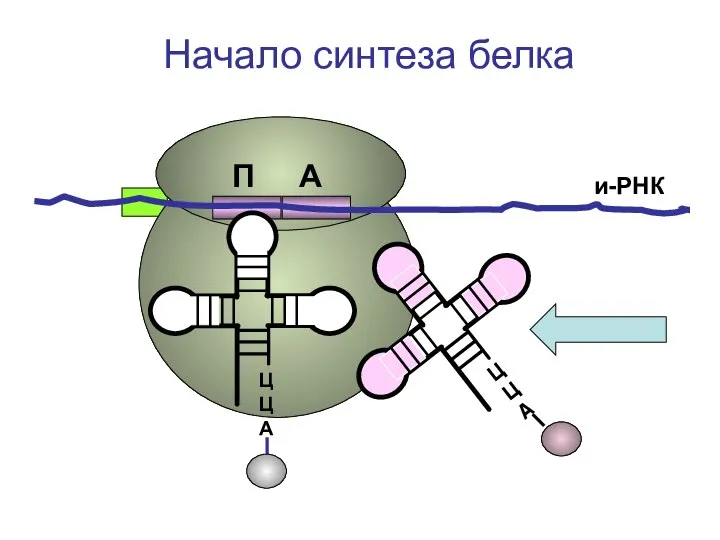
Слайд 31ЦЦА
и-РНК
Начало синтеза белка
П А
ЦЦА
и-РНК
Начало синтеза белка
П А
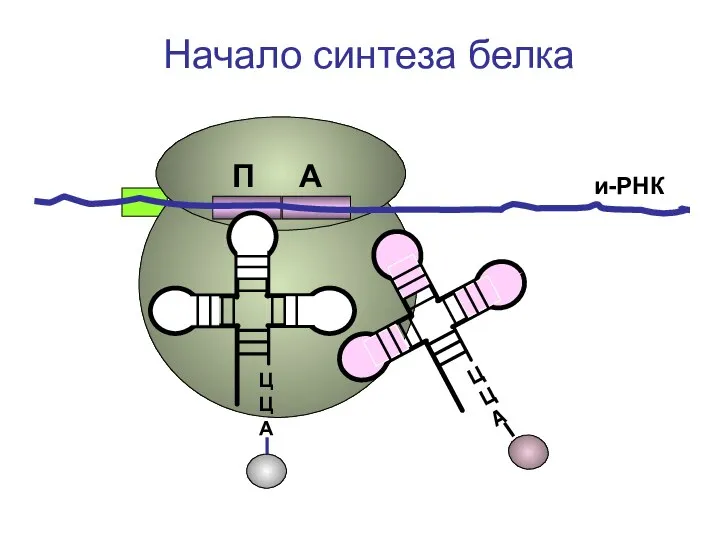
Слайд 32ЦЦА
и-РНК
Начало синтеза белка
П А
ЦЦА
и-РНК
Начало синтеза белка
П А
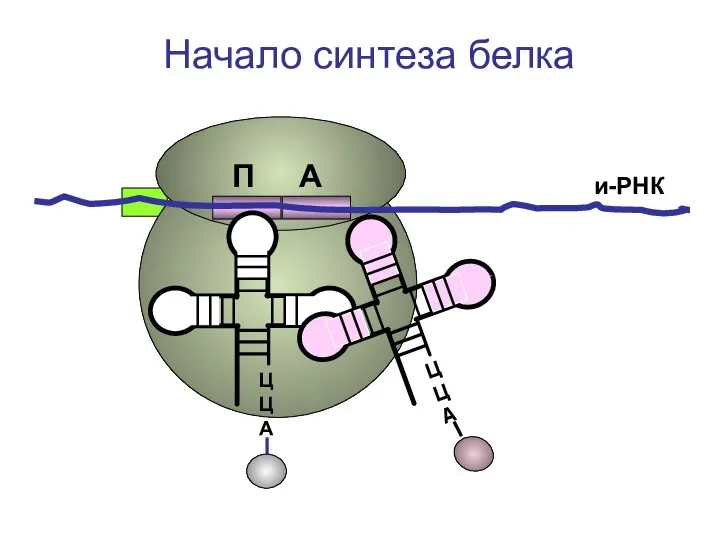
Слайд 33ЦЦА
и-РНК
Начало синтеза белка
П А
ЦЦА
и-РНК
Начало синтеза белка
П А
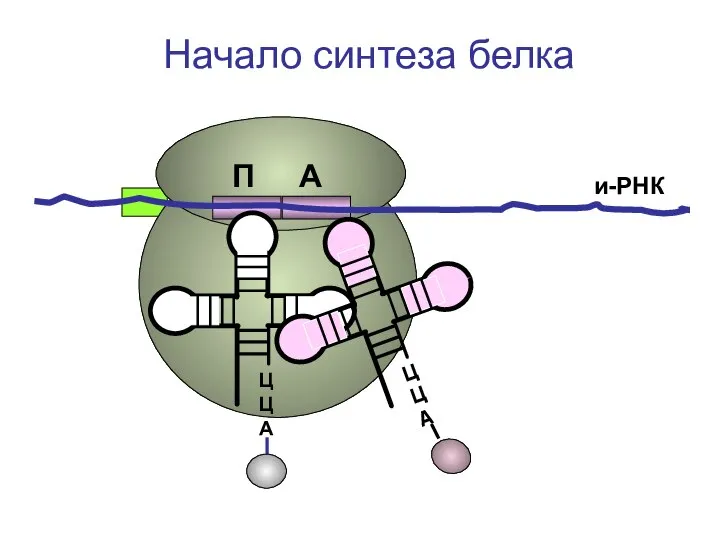
Слайд 34ЦЦА
и-РНК
Начало синтеза белка
П А
ЦЦА
и-РНК
Начало синтеза белка
П А
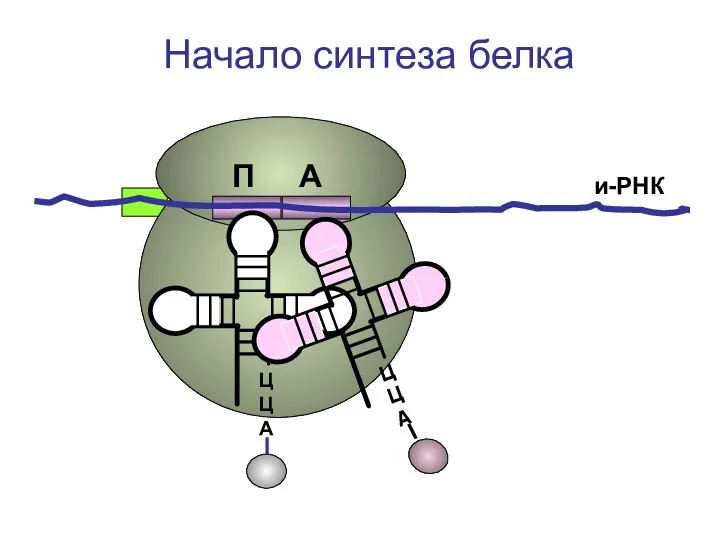
Слайд 35ЦЦА
ЦЦА
и-РНК
Перенос первой аминокислоты на вторую
П А
ЦЦА
ЦЦА
и-РНК
Перенос первой аминокислоты на вторую
П А
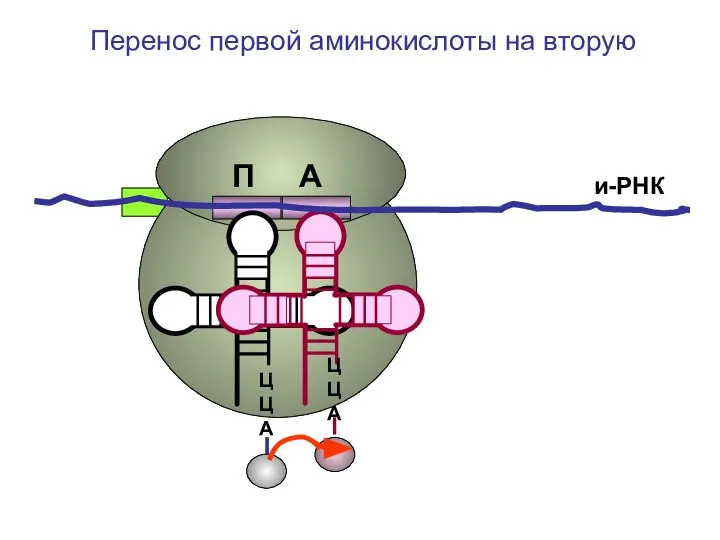
Слайд 36ЦЦА
ЦЦА
и-РНК
Перенос первой аминокислоты на вторую
П А
ЦЦА
ЦЦА
и-РНК
Перенос первой аминокислоты на вторую
П А
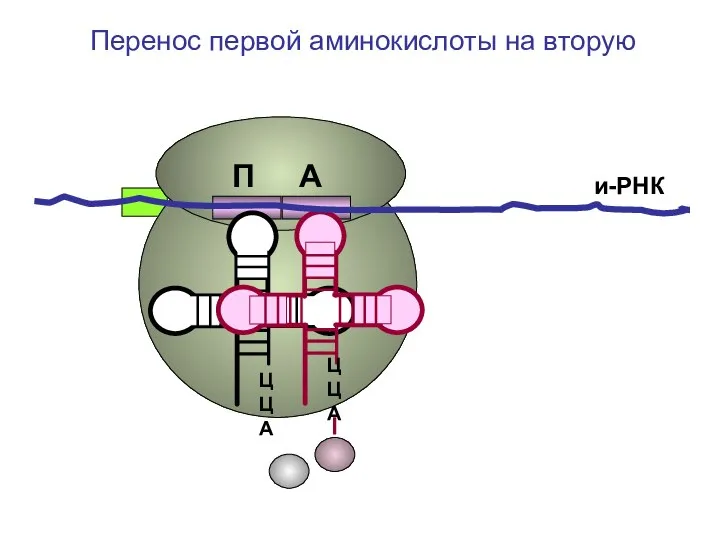
Слайд 37ЦЦА
ЦЦА
и-РНК
Образование дипептида
П А
ЦЦА
ЦЦА
и-РНК
Образование дипептида
П А
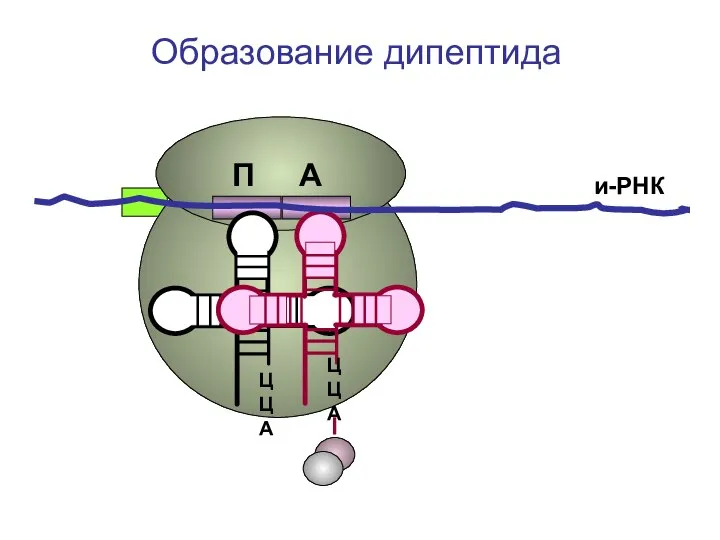
Слайд 38ЦЦА
ЦЦА
и-РНК
П А
Перемещение рибосомы вдоль и-РНК на один триплет
ЦЦА
ЦЦА
и-РНК
П А
Перемещение рибосомы вдоль и-РНК на один триплет
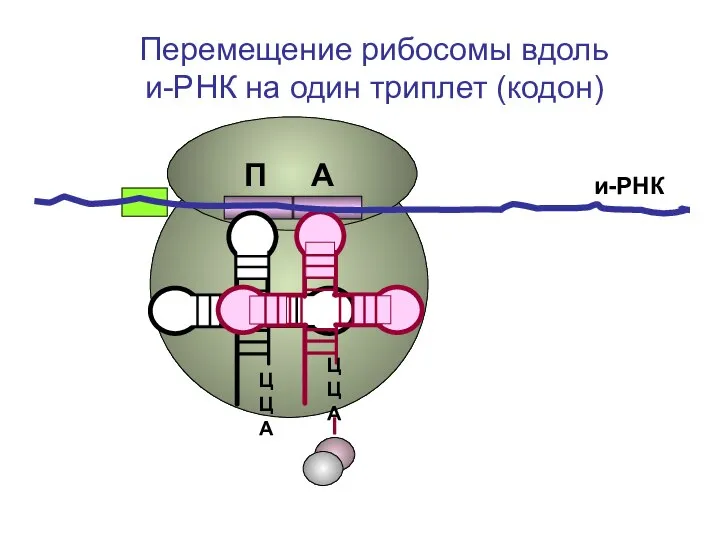
Слайд 39ЦЦА
ЦЦА
и-РНК
П А
Перемещение рибосомы вдоль и-РНК на один триплет
ЦЦА
ЦЦА
и-РНК
П А
Перемещение рибосомы вдоль и-РНК на один триплет
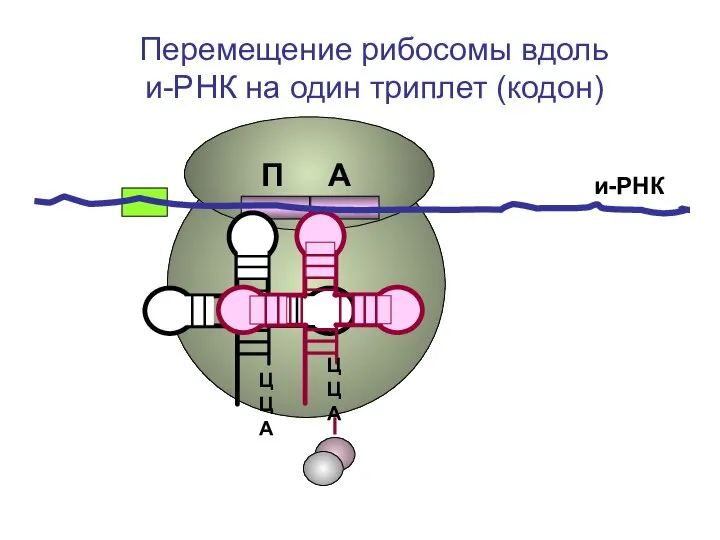
Слайд 40и-РНК
П А
Перемещение рибосомы вдоль и-РНК на один триплет (кодон)
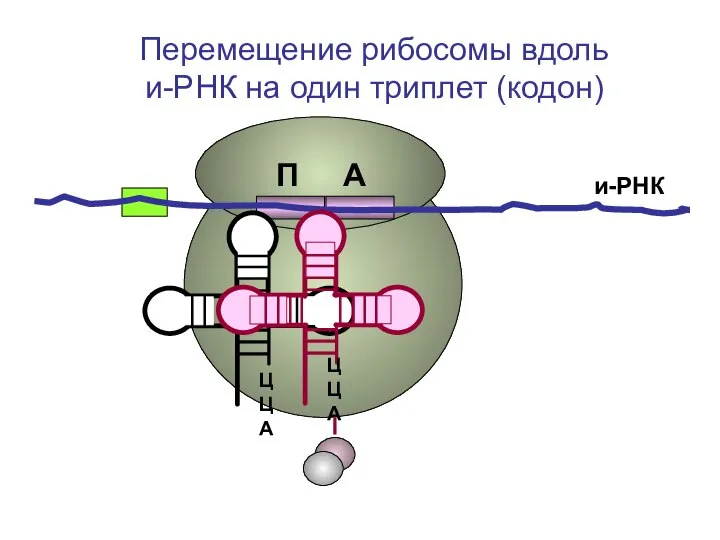
и-РНК
П А
Перемещение рибосомы вдоль и-РНК на один триплет (кодон)
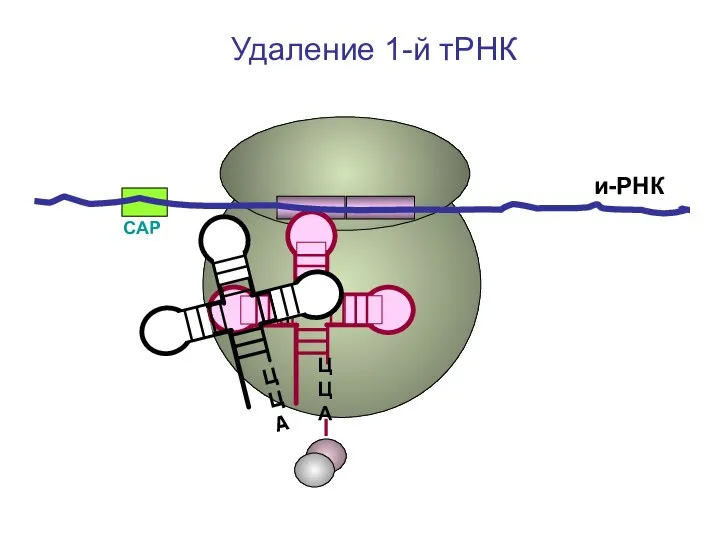
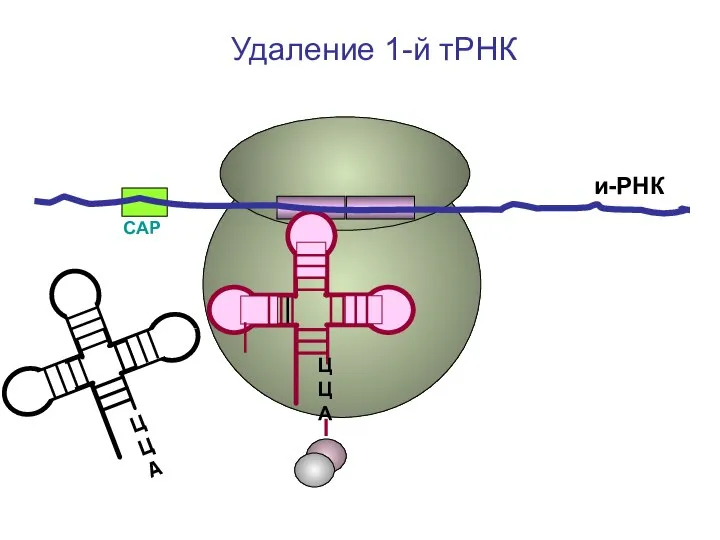
Слайд 41СAP
и-РНК
ЦЦА
Удаление 1-й тРНК
СAP
и-РНК
ЦЦА
Удаление 1-й тРНК
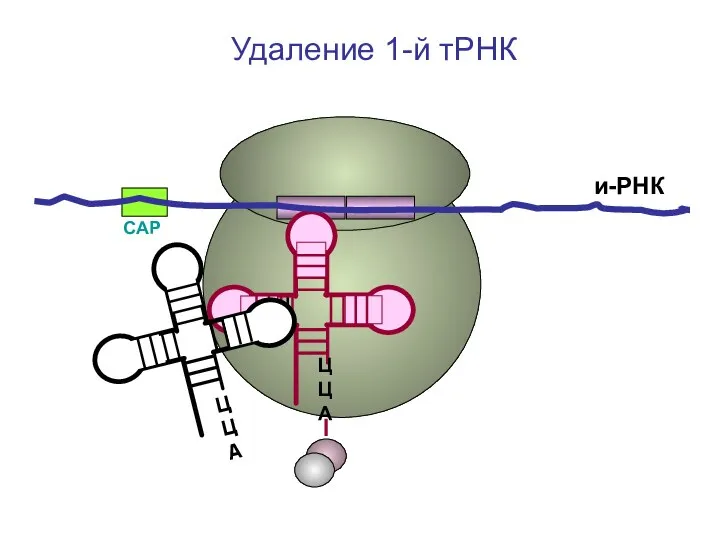
Слайд 42СAP
и-РНК
ЦЦА
Удаление 1-й тРНК
СAP
и-РНК
ЦЦА
Удаление 1-й тРНК
Слайд 43СAP
и-РНК
ЦЦА
ЦЦА
Удаление 1-й тРНК
СAP
и-РНК
ЦЦА
ЦЦА
Удаление 1-й тРНК
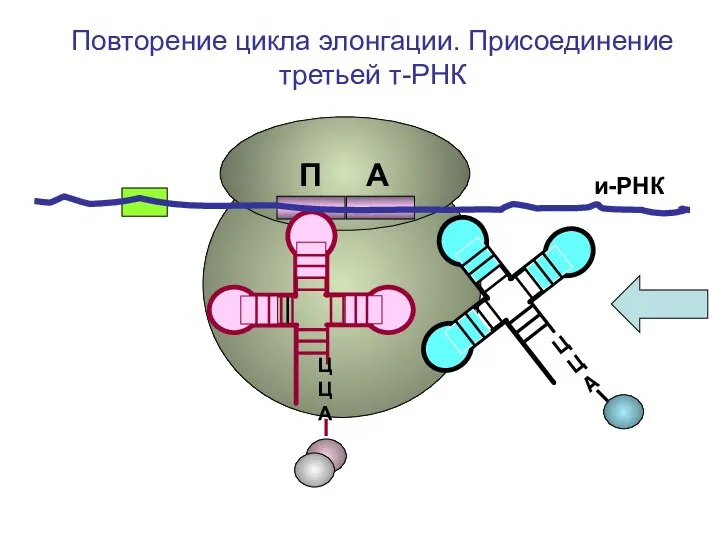
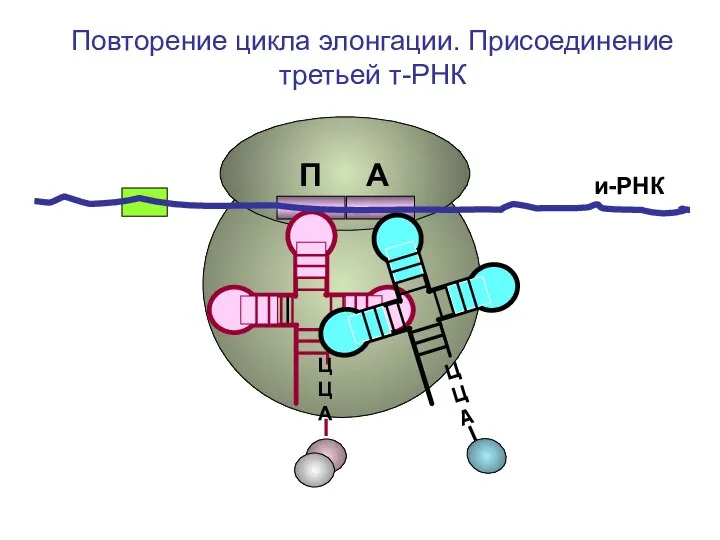
Слайд 44и-РНК
ЦЦА
ЦЦА
Повторение цикла элонгации. Присоединение третьей т-РНК
П А
и-РНК
ЦЦА
ЦЦА
Повторение цикла элонгации. Присоединение третьей т-РНК
П А
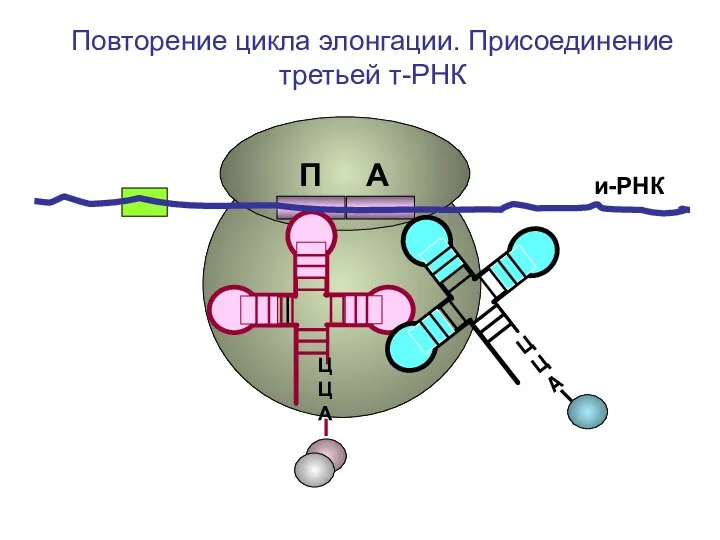
Слайд 45и-РНК
ЦЦА
Повторение цикла элонгации. Присоединение третьей т-РНК
П А
и-РНК
ЦЦА
Повторение цикла элонгации. Присоединение третьей т-РНК
П А
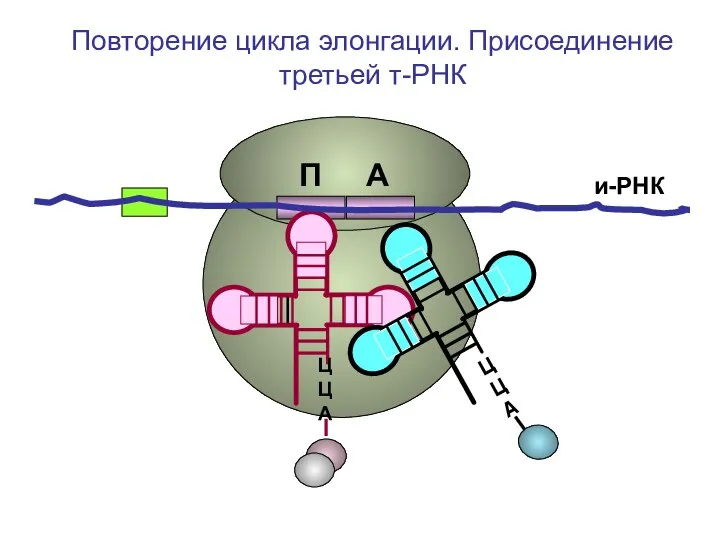
Слайд 46и-РНК
ЦЦА
Повторение цикла элонгации. Присоединение третьей т-РНК
П А
и-РНК
ЦЦА
Повторение цикла элонгации. Присоединение третьей т-РНК
П А
Слайд 47и-РНК
ЦЦА
Повторение цикла элонгации. Присоединение третьей т-РНК
П А
и-РНК
ЦЦА
Повторение цикла элонгации. Присоединение третьей т-РНК
П А
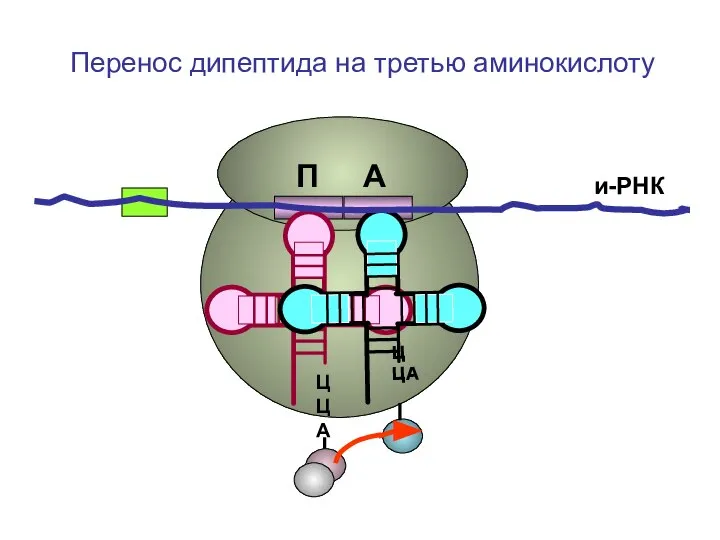
Слайд 48ЦЦА
ЦЦА
и-РНК
П А
Перенос дипептида на третью аминокислоту
ЦЦА
ЦЦА
и-РНК
П А
Перенос дипептида на третью аминокислоту
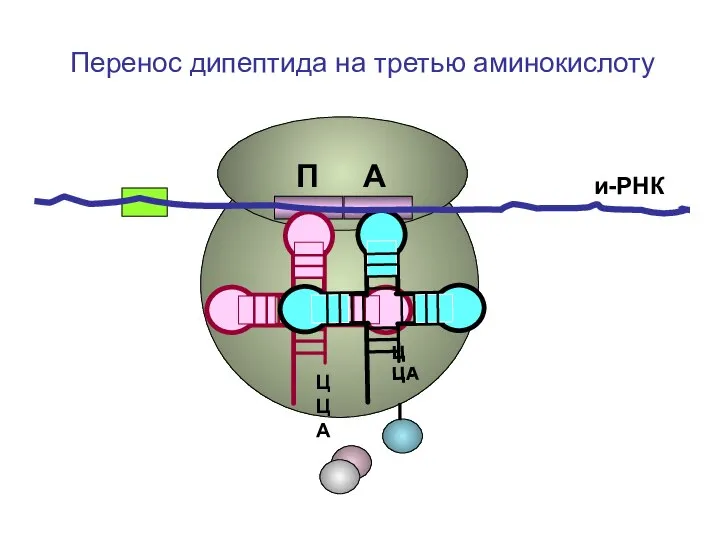
Слайд 49ЦЦА
ЦЦА
и-РНК
П А
Перенос дипептида на третью аминокислоту
ЦЦА
ЦЦА
и-РНК
П А
Перенос дипептида на третью аминокислоту
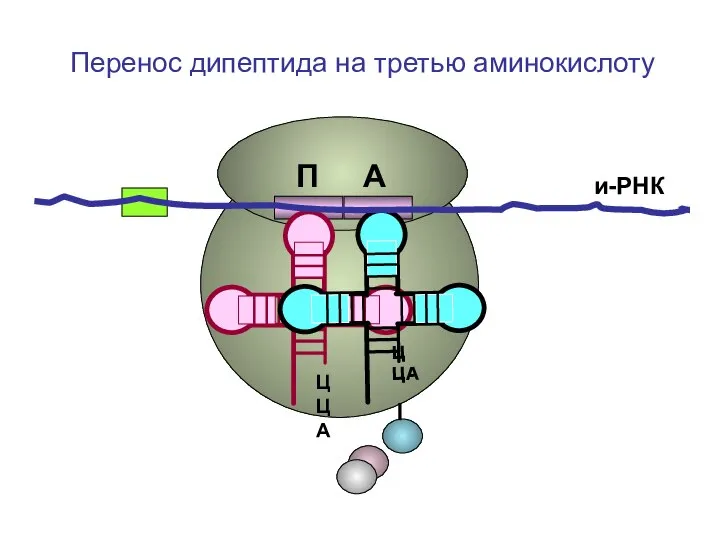
Слайд 50ЦЦА
ЦЦА
и-РНК
П А
Перенос дипептида на третью аминокислоту
ЦЦА
ЦЦА
и-РНК
П А
Перенос дипептида на третью аминокислоту
Слайд 51ЦЦА
ЦЦА
и-РНК
П А
Образование трипептида
ЦЦА
ЦЦА
и-РНК
П А
Образование трипептида
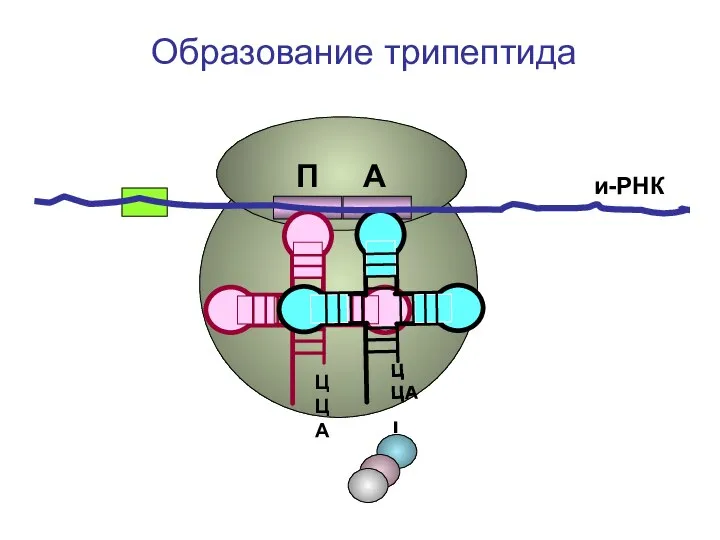
Слайд 52ЦЦА
ЦЦА
и-РНК
П А
Перемещение рибосомы вдоль и-РНК на один триплет (кодон)
ЦЦА
ЦЦА
и-РНК
П А
Перемещение рибосомы вдоль и-РНК на один триплет (кодон)
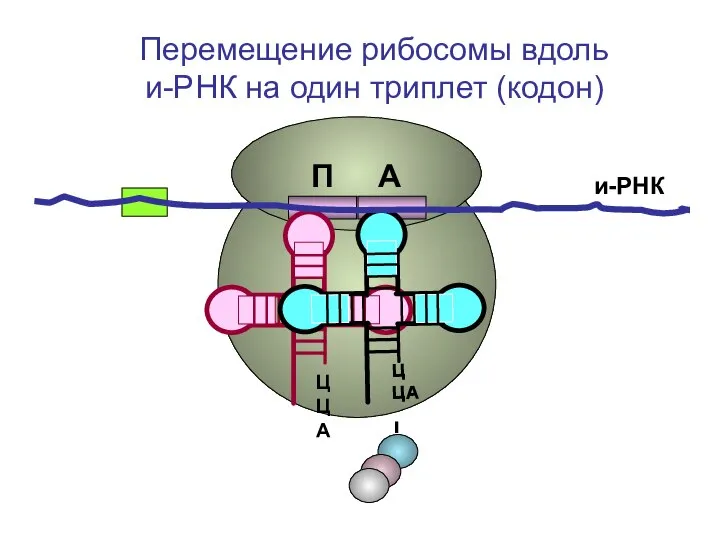
Слайд 53ЦЦА
ЦЦА
и-РНК
П А
Перемещение рибосомы вдоль и-РНК на один триплет (кодон)
ЦЦА
ЦЦА
и-РНК
П А
Перемещение рибосомы вдоль и-РНК на один триплет (кодон)
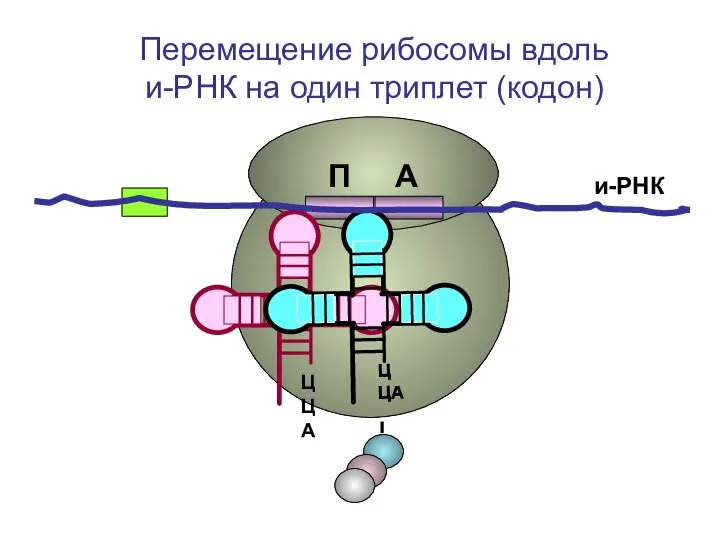
Слайд 54ЦЦА
ЦЦА
и-РНК
П А
Перемещение рибосомы вдоль и-РНК на один триплет (кодон)
ЦЦА
ЦЦА
и-РНК
П А
Перемещение рибосомы вдоль и-РНК на один триплет (кодон)
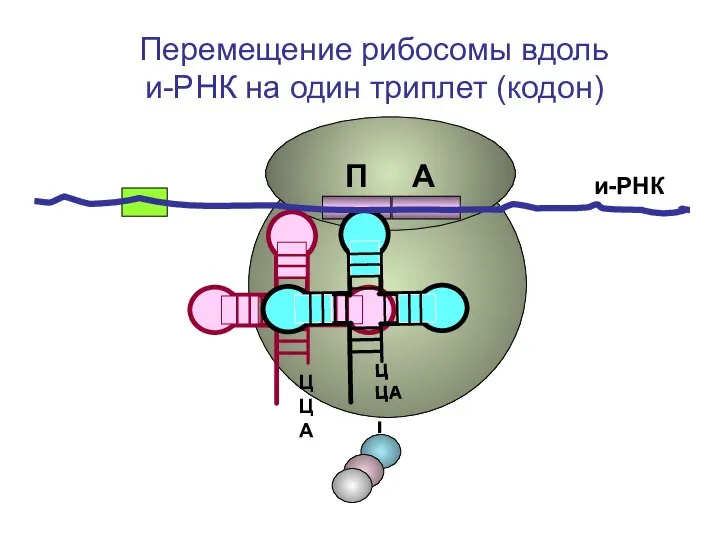
Слайд 55СAP
и-РНК
ЦЦА
ЦЦА
ЦЦА
Повторение цикла элонгации. Присоединение четвертой
СAP
и-РНК
ЦЦА
ЦЦА
ЦЦА
Повторение цикла элонгации. Присоединение четвертой
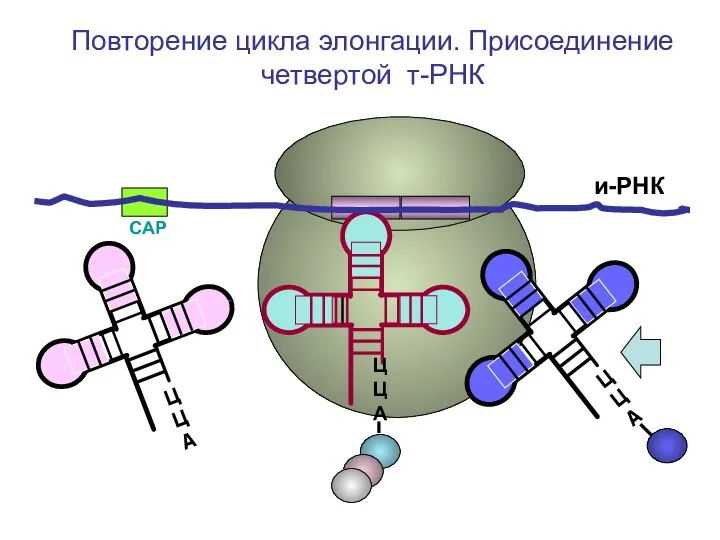
Слайд 56СAP
и-РНК
ЦЦА
ЦЦА
Повторение цикла элонгации. Присоединение четвертой т-РНК
СAP
и-РНК
ЦЦА
ЦЦА
Повторение цикла элонгации. Присоединение четвертой т-РНК
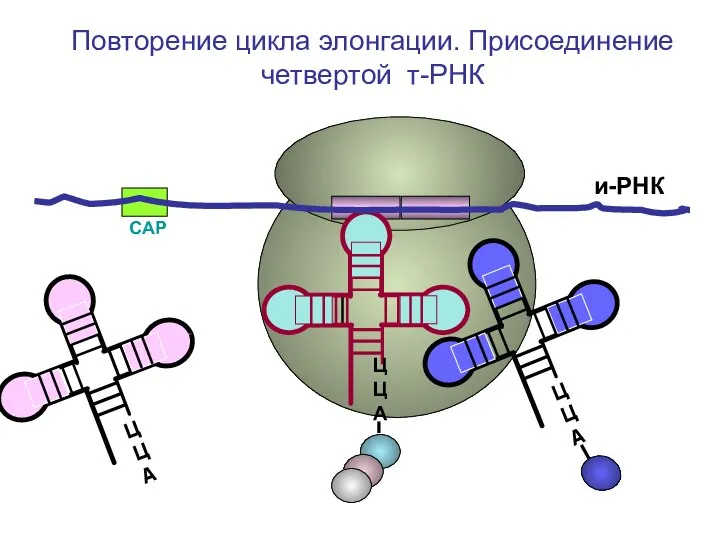
Слайд 57СAP
и-РНК
ЦЦА
Повторение цикла элонгации. Присоединение четвертой т-РНК
СAP
и-РНК
ЦЦА
Повторение цикла элонгации. Присоединение четвертой т-РНК
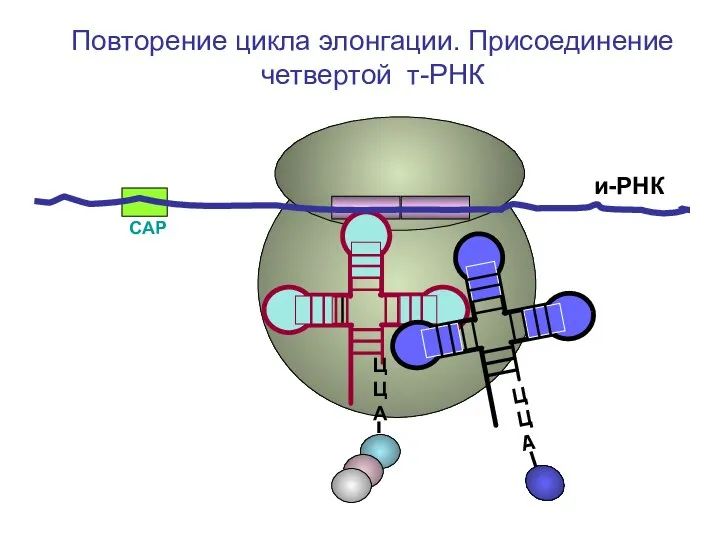
Слайд 58СAP
и-РНК
ЦЦА
Повторение цикла элонгации. Присоединение четвертой т-РНК
СAP
и-РНК
ЦЦА
Повторение цикла элонгации. Присоединение четвертой т-РНК
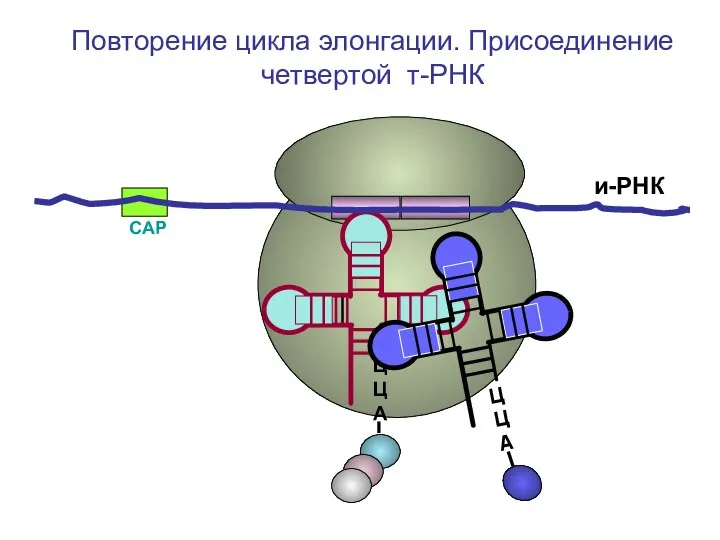
Слайд 59СAP
и-РНК
ЦЦА
Повторение цикла элонгации. Присоединение четвертой т-РНК
СAP
и-РНК
ЦЦА
Повторение цикла элонгации. Присоединение четвертой т-РНК
Слайд 60СAP
и-РНК
ЦЦА
Повторение цикла элонгации. Присоединение четвертой т-РНК
СAP
и-РНК
ЦЦА
Повторение цикла элонгации. Присоединение четвертой т-РНК
Слайд 61СAP
и-РНК
ЦЦА
Перенос трипептида на четвертую аминокислоту
СAP
и-РНК
ЦЦА
Перенос трипептида на четвертую аминокислоту
Слайд 62СAP
и-РНК
ЦЦА
Перенос трипептида на четвертую аминокислоту
СAP
и-РНК
ЦЦА
Перенос трипептида на четвертую аминокислоту
Слайд 63СAP
и-РНК
ЦЦА
Перенос трипептида на четвертую аминокислоту
СAP
и-РНК
ЦЦА
Перенос трипептида на четвертую аминокислоту
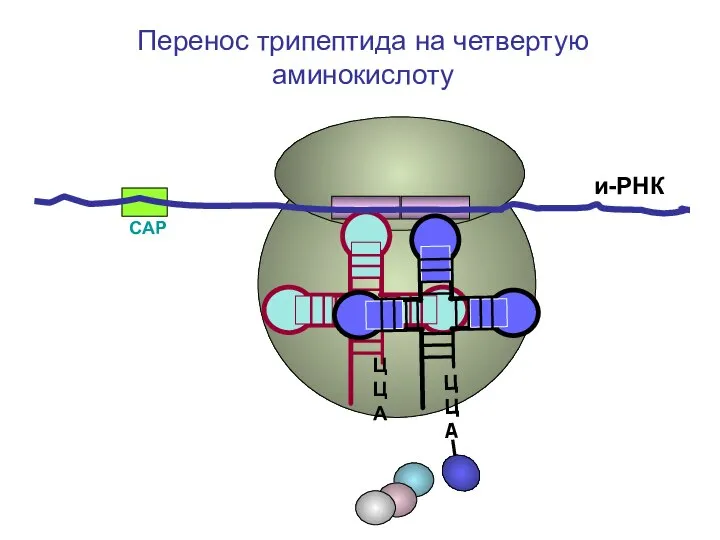
Слайд 64СAP
и-РНК
ЦЦА
Перенос трипептида на четвертую аминокислоту
СAP
и-РНК
ЦЦА
Перенос трипептида на четвертую аминокислоту
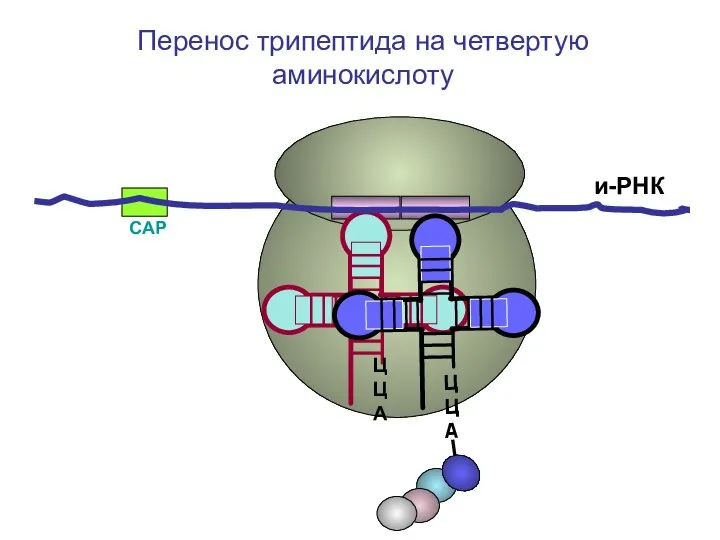
Слайд 65СAP
и-РНК
ЦЦА
Перемещение рибосомы вдоль и-РНК на один триплет (кодон)
СAP
и-РНК
ЦЦА
Перемещение рибосомы вдоль и-РНК на один триплет (кодон)
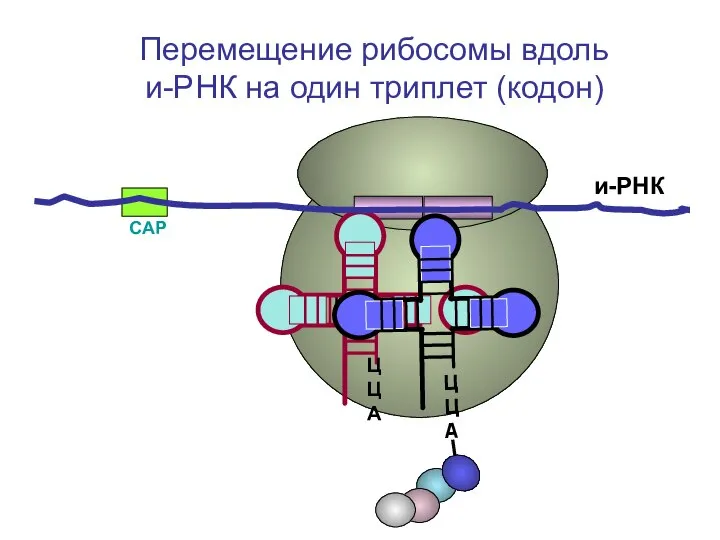
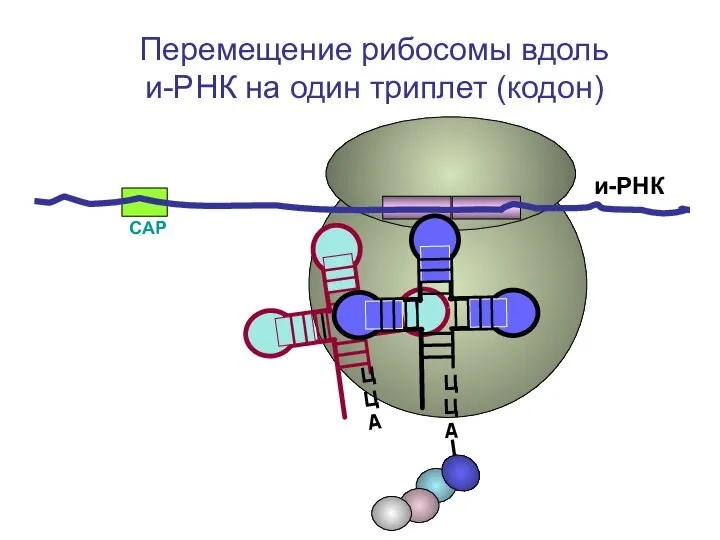
Слайд 66СAP
и-РНК
Перемещение рибосомы вдоль и-РНК на один триплет (кодон)
СAP
и-РНК
Перемещение рибосомы вдоль и-РНК на один триплет (кодон)
Слайд 67СAP
и-РНК
ЦЦА
Повторение цикла элонгации. Присоединение пятой т-РНК и т. д.
СAP
и-РНК
ЦЦА
Повторение цикла элонгации. Присоединение пятой т-РНК и т. д.
Слайд 68Стадия терминации
Стадия терминации

Слайд 69 Образовавшийся белок, потоком жидкости в эндоплазматическом ретикулуме (состоящим из трубочек), направляется
Образовавшийся белок, потоком жидкости в эндоплазматическом ретикулуме (состоящим из трубочек), направляется

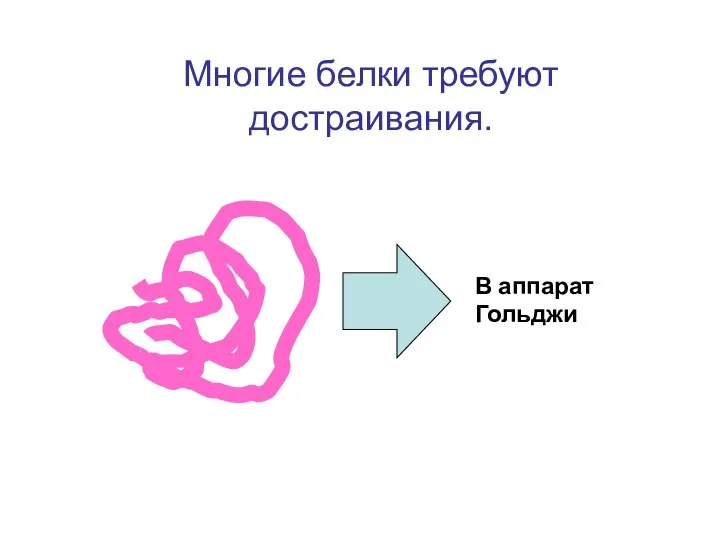
Слайд 70В аппарат Гольджи
Многие белки требуют достраивания.
В аппарат Гольджи
Многие белки требуют достраивания.
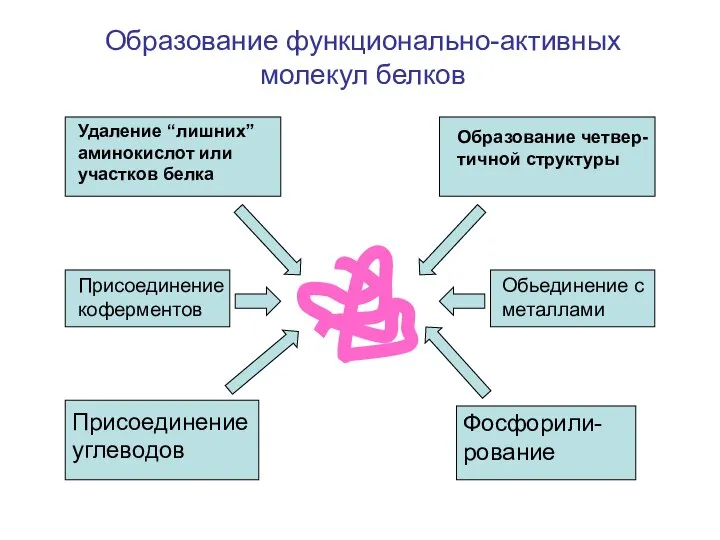
Слайд 71Присоединение коферментов
Присоединение углеводов
Обьединение с металлами
Фосфорили-рование
Удаление “лишних” аминокислот или участков белка
Образование четвер-тичной структуры
Образование
Присоединение коферментов
Присоединение углеводов
Обьединение с металлами
Фосфорили-рование
Удаление “лишних” аминокислот или участков белка
Образование четвер-тичной структуры
Образование
Слайд 72Регуляция обмена белков
Регуляция обмена белков

Слайд 73Гормональная регуляция скорости синтеза белка
Соматотропин, половые гормоны (индукторы транскрипции и биосинтеза белка);
2.
Гормональная регуляция скорости синтеза белка
Соматотропин, половые гормоны (индукторы транскрипции и биосинтеза белка);
2.

Слайд 74Влияние некоторых факторов на концентрацию и состав белков организма
Доступность в белковой диете;
Влияние некоторых факторов на концентрацию и состав белков организма
Доступность в белковой диете;

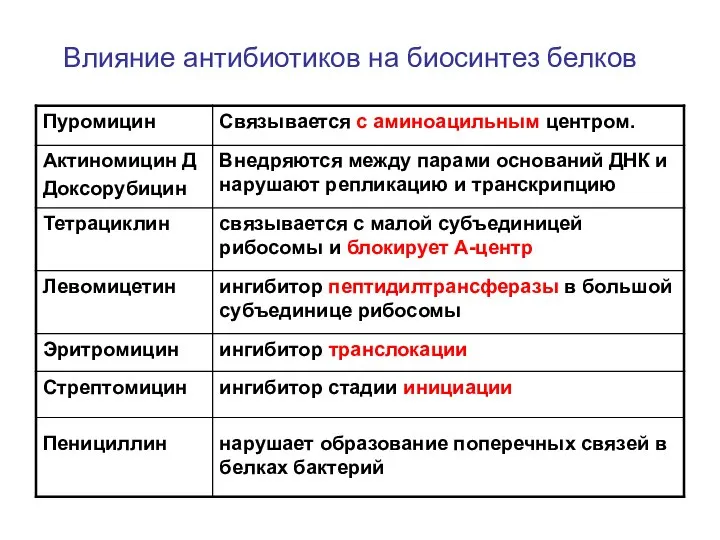
Слайд 75Влияние антибиотиков на биосинтез белков
Влияние антибиотиков на биосинтез белков
Слайд 76Трансамини-рование аминокислот
Трансамини-рование аминокислот

Слайд 77 Поступающий с пищей набор аминокислот, редко соответствует их требуемому соотношению для
Поступающий с пищей набор аминокислот, редко соответствует их требуемому соотношению для

Слайд 78СН3 СООН СН3 СООН
СНNH2 + C=О С=О + СНNH2
СООН СН2 СООН СН2
СНNH2 + C=О С=О + СНNH2
СООН СН2 СООН СН2
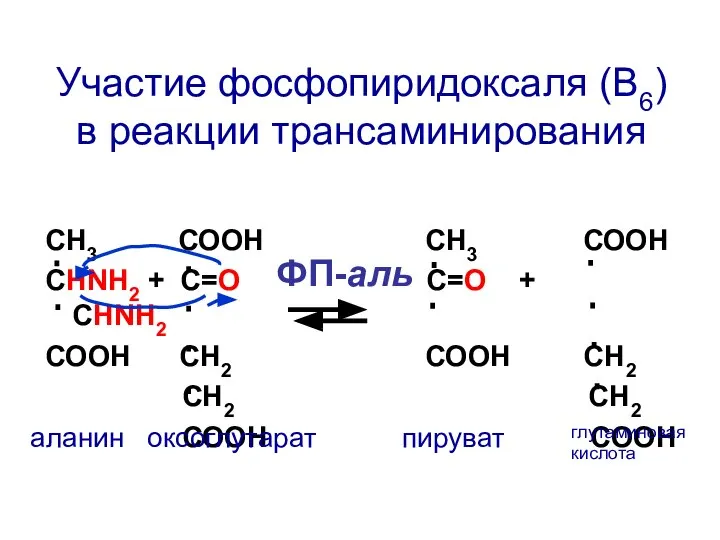
Слайд 79 С
СН2ОН
НО
Н3С
N
Н О
пиридоксаль
Витамин В6
(неактивная форма витамина)
С
СН2ОН
НО
Н3С
N
Н О
пиридоксаль
Витамин В6
(неактивная форма витамина)
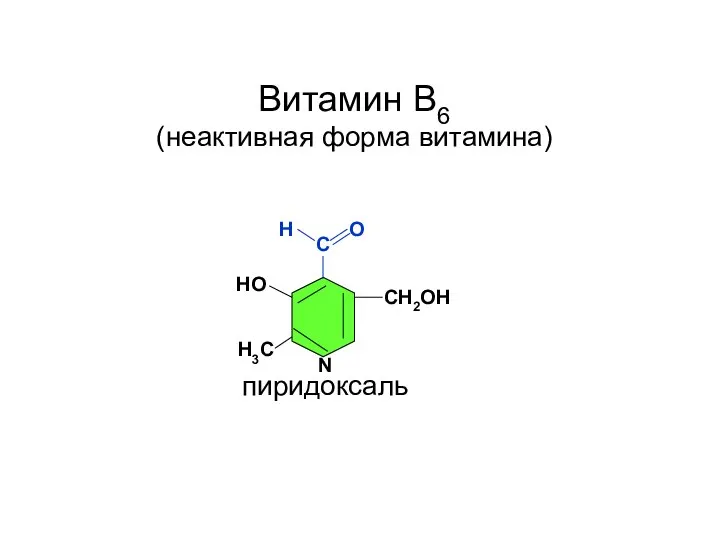
Слайд 80 С
СН2О-РО3Н2
НО
Н3С
N
Н О
Фосфопиридоксаль
(активная форма витамина)
С
СН2О-РО3Н2
НО
Н3С
N
Н О
Фосфопиридоксаль
(активная форма витамина)
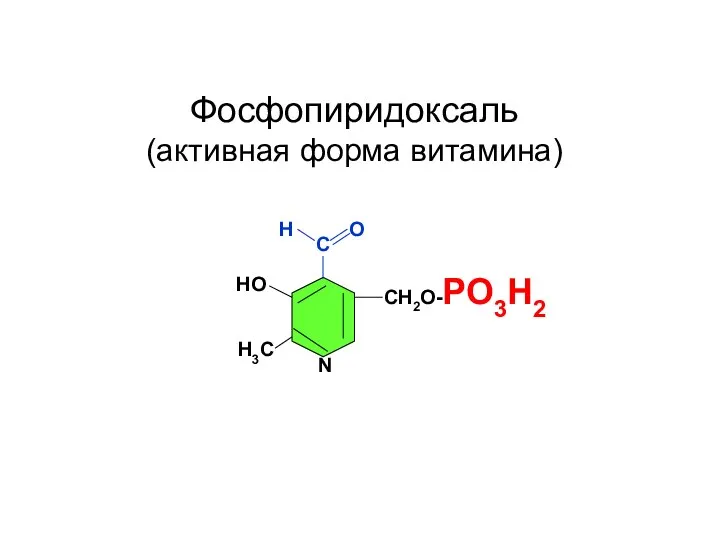
Слайд 81Н О
С
СН2О-РО3Н2
НО
Н3С
N
СН2-NH2
СН2О-РО3Н2
НО
Н3С
N
пиридоксамин-фосфат
Витамин как акцептор аминогруппы
пиридоксаль-фосфат
+
СН2
СН-NН2
СООН
СН2
С = О
СООН
+
Н О
С
СН2О-РО3Н2
НО
Н3С
N
СН2-NH2
СН2О-РО3Н2
НО
Н3С
N
пиридоксамин-фосфат
Витамин как акцептор аминогруппы
пиридоксаль-фосфат
+
СН2
СН-NН2
СООН
СН2
С = О
СООН
+
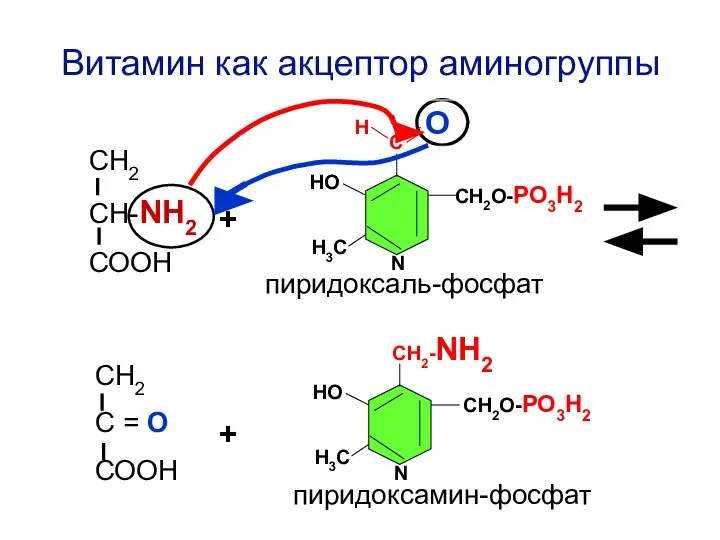
Слайд 82Н О
С
СН2О-РО3Н2
НО
Н3С
N
СН2-NH2
СН2О-РО3Н2
НО
Н3С
N
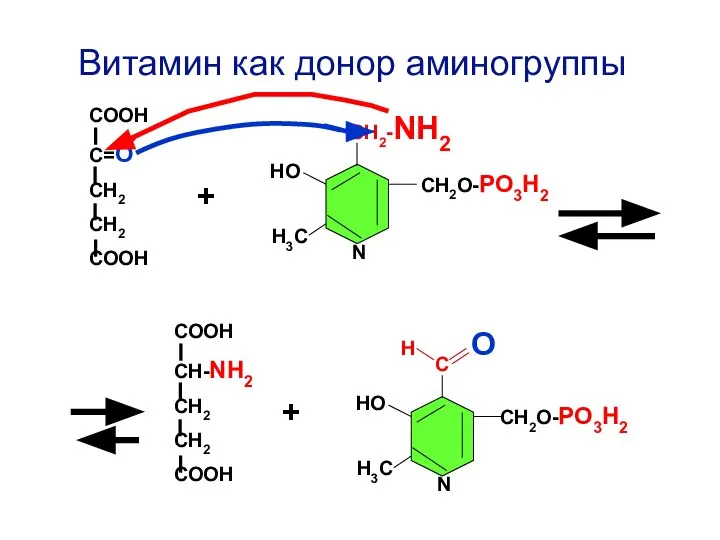
Витамин как донор аминогруппы
+
СООН
С=О
СН2
СН2
СООН
+
СООН
СН-NН2
СН2
СН2
СООН
Н О
С
СН2О-РО3Н2
НО
Н3С
N
СН2-NH2
СН2О-РО3Н2
НО
Н3С
N
Витамин как донор аминогруппы
+
СООН
С=О
СН2
СН2
СООН
+
СООН
СН-NН2
СН2
СН2
СООН
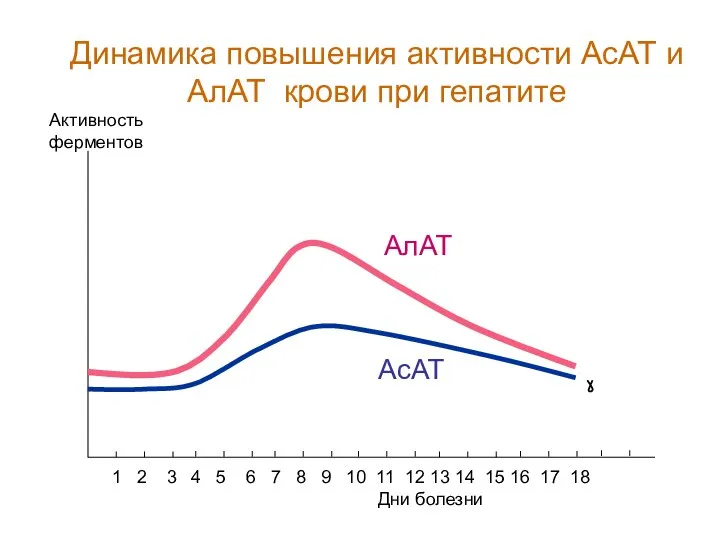
Слайд 83 В клинической практике активность АлАТ или АсАТ часто измеряют в сыворотке
В клинической практике активность АлАТ или АсАТ часто измеряют в сыворотке

Слайд 84Дни болезни
Активность ферментов
1 2 3 4 5 6 7 8 9 10
Дни болезни
Активность ферментов
1 2 3 4 5 6 7 8 9 10
Слайд 85Декарбоксилирование аминокислот (образование биологически-активных аминов)
Декарбоксилирование аминокислот (образование биологически-активных аминов)

Слайд 86 Часть аминокислот используется организмом для получения биологически активных молекул, которые участвуют
Часть аминокислот используется организмом для получения биологически активных молекул, которые участвуют

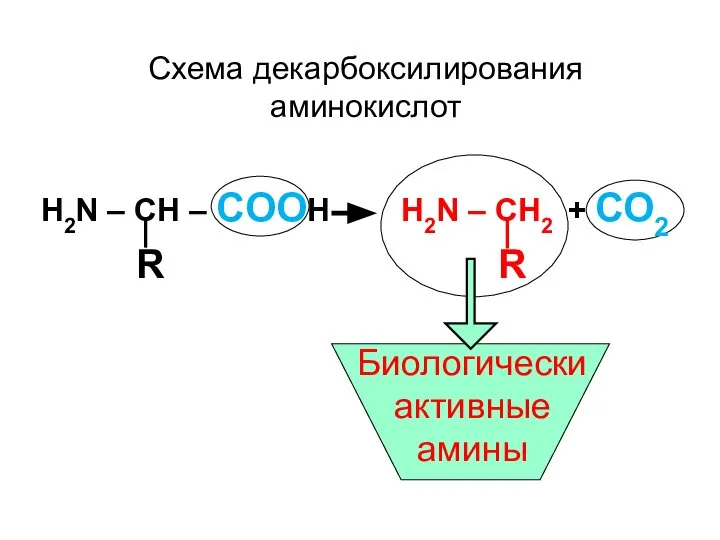
Слайд 87H2N – CН – CООН H2N – CН2 + СО2
R R
Схема декарбоксилирования
H2N – CН – CООН H2N – CН2 + СО2
R R
Схема декарбоксилирования
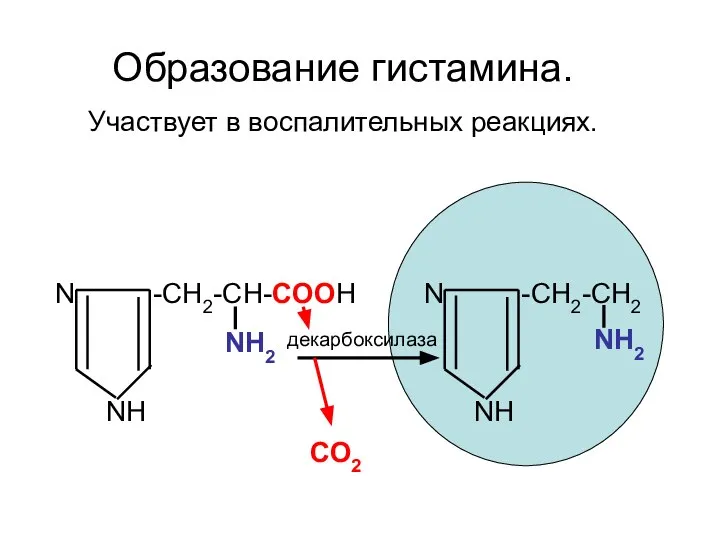
Слайд 88N -CH2-CH-COOH
NH2
NH
N -CH2-CH2
NH2
NH
декарбоксилаза
СО2
Образование гистамина.
Участвует в воспалительных реакциях.
N -CH2-CH-COOH
NH2
NH
N -CH2-CH2
NH2
NH
декарбоксилаза
СО2
Образование гистамина.
Участвует в воспалительных реакциях.
Слайд 89-CH2-CH-COOH
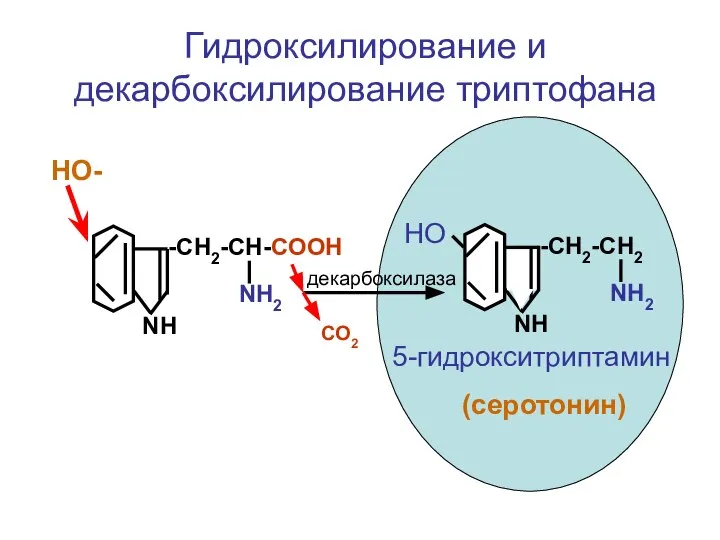
NH2
NH
НО-
-CH2-CH2
NH2
NH
НО
5-гидрокситриптамин
(серотонин)
декарбоксилаза
СО2
Гидроксилирование и декарбоксилирование триптофана
-CH2-CH-COOH
NH2
NH
НО-
-CH2-CH2
NH2
NH
НО
5-гидрокситриптамин
(серотонин)
декарбоксилаза
СО2
Гидроксилирование и декарбоксилирование триптофана
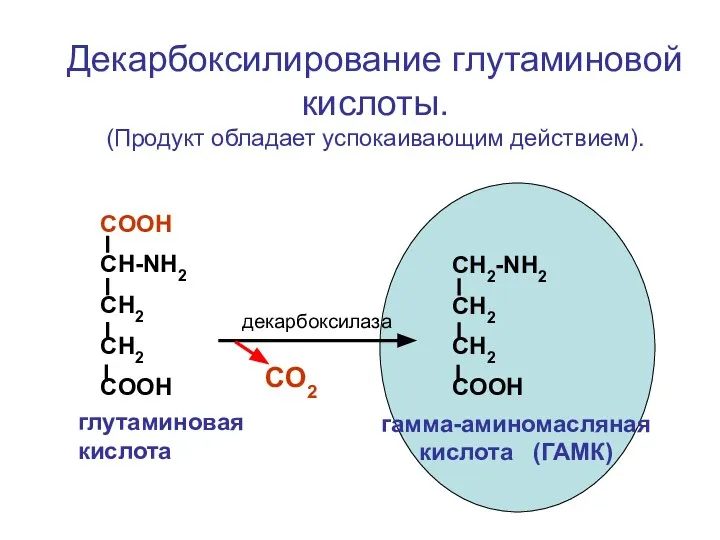
Слайд 90СOOH
CH-NH2
CH2
CH2
COOH
CH2-NH2
CH2
CH2
COOH
декарбоксилаза
CO2
глутаминовая кислота
гамма-аминомасляная кислота (ГАМК)
Декарбоксилирование глутаминовой кислоты.
(Продукт обладает успокаивающим действием).
СOOH
CH-NH2
CH2
CH2
COOH
CH2-NH2
CH2
CH2
COOH
декарбоксилаза
CO2
глутаминовая кислота
гамма-аминомасляная кислота (ГАМК)
Декарбоксилирование глутаминовой кислоты.
(Продукт обладает успокаивающим действием).
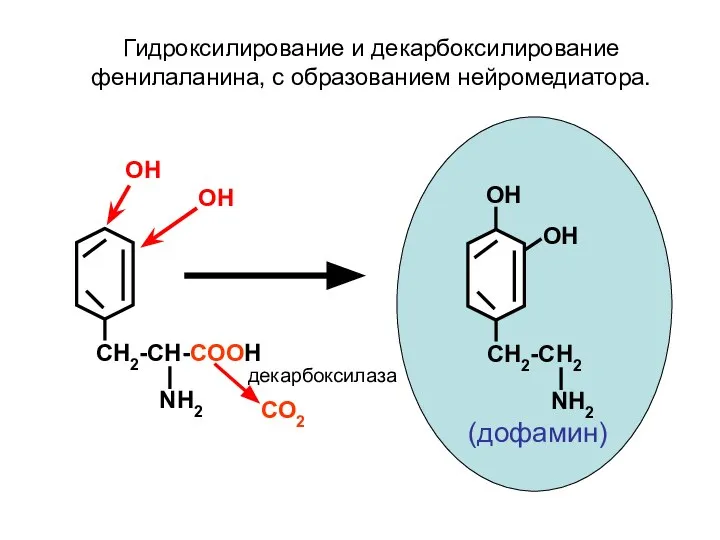
Слайд 91 CH2-CH-COOH
NH2
ОН
ОН
CH2-CH2
NH2
ОН
ОН
СО2
декарбоксилаза
Гидроксилирование и декарбоксилирование фенилаланина, с образованием нейромедиатора.
(дофамин)
CH2-CH-COOH
NH2
ОН
ОН
CH2-CH2
NH2
ОН
ОН
СО2
декарбоксилаза
Гидроксилирование и декарбоксилирование фенилаланина, с образованием нейромедиатора.
(дофамин)
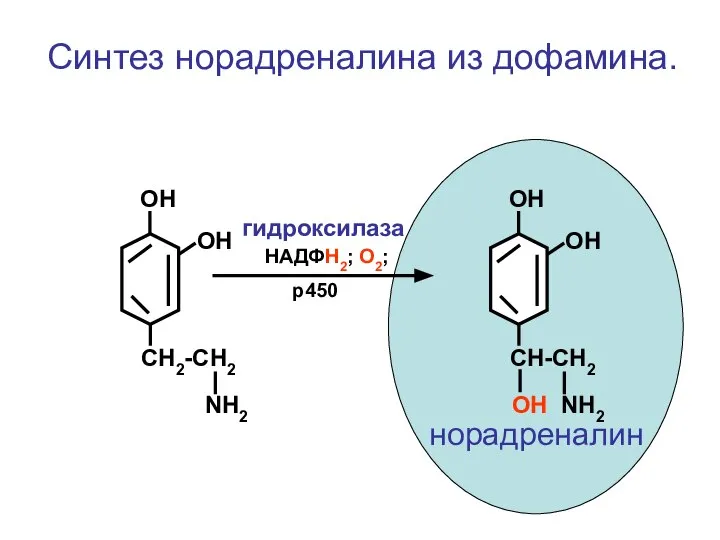
Слайд 92 CH2-CH2
NH2
ОН
ОН
Синтез норадреналина из дофамина.
CH-CH2
ОН NH2
ОН
ОН
НАДФН2; О2;
р450
гидроксилаза
норадреналин
CH2-CH2
NH2
ОН
ОН
Синтез норадреналина из дофамина.
CH-CH2
ОН NH2
ОН
ОН
НАДФН2; О2;
р450
гидроксилаза
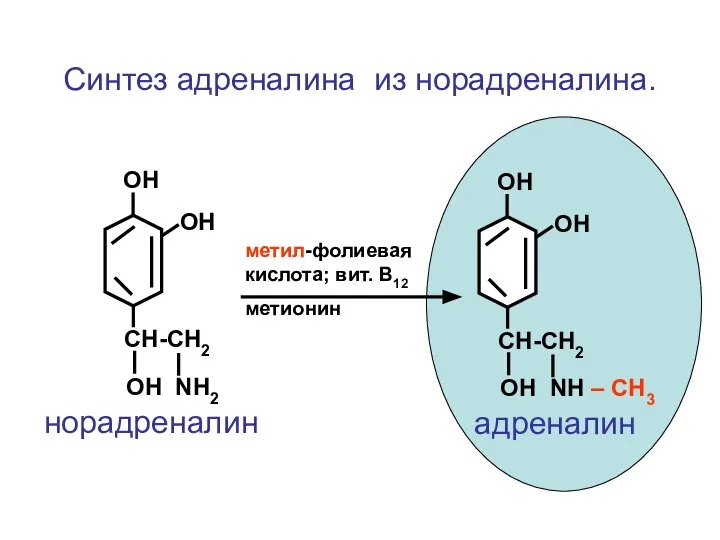
норадреналин
Слайд 93 CH-CH2
ОН NH2
ОН
ОН
норадреналин
CH-CH2
ОН NH – СН3
ОН
ОН
адреналин
метил-фолиевая кислота; вит. В12
метионин
Синтез адреналина из
CH-CH2
ОН NH2
ОН
ОН
норадреналин
CH-CH2
ОН NH – СН3
ОН
ОН
адреналин
метил-фолиевая кислота; вит. В12
метионин
Синтез адреналина из
Слайд 94Биологически активные амины, используемые как лекарственные средства.
Гистамин
Триптамин
Серотонин
Гамма-аминомасляная кислота (ГАМК)
Дофамин
Норадреналин
Адреналин
Биологически активные амины, используемые как лекарственные средства.
Гистамин
Триптамин
Серотонин
Гамма-аминомасляная кислота (ГАМК)
Дофамин
Норадреналин
Адреналин
Слайд 95Использование аминокислот в качестве исходных субстратов для выработки энергии с помощью окислительного
Использование аминокислот в качестве исходных субстратов для выработки энергии с помощью окислительного

Слайд 96 Иногда в клетках возникает необходимость в более интенсивном образовании энергии. При
Иногда в клетках возникает необходимость в более интенсивном образовании энергии. При

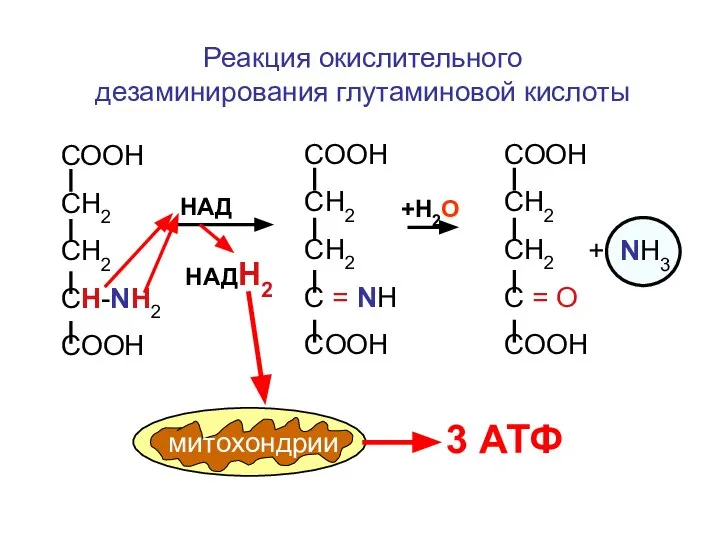
Слайд 97Реакция окислительного дезаминирования глутаминовой кислоты
СООН
СН2
СН2
CH-NH2
COOH
HАД
HАДН2
+Н2О
СООН
СН2
СН2
C = NH
COOH
СООН
СН2
СН2
C = О
COOH
+ NH3
митохондрии
3 АТФ
Реакция окислительного дезаминирования глутаминовой кислоты
СООН
СН2
СН2
CH-NH2
COOH
HАД
HАДН2
+Н2О
СООН
СН2
СН2
C = NH
COOH
СООН
СН2
СН2
C = О
COOH
+ NH3
митохондрии
3 АТФ
Слайд 98 В результате окислительного дезаминирования глутаминовой, или аспарагиновой кислот образуется одна молекула
В результате окислительного дезаминирования глутаминовой, или аспарагиновой кислот образуется одна молекула

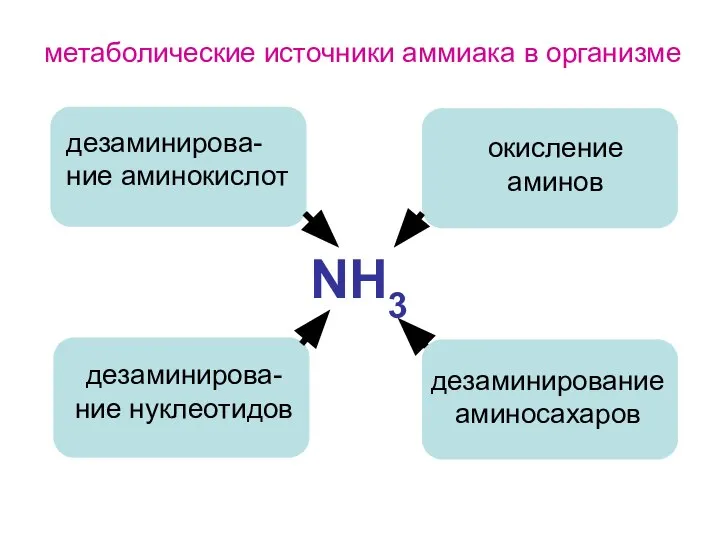
Слайд 99NH3
дезаминирова-ние аминокислот
дезаминирова-ние нуклеотидов
окисление аминов
дезаминирование аминосахаров
метаболические источники аммиака в организме
NH3
дезаминирова-ние аминокислот
дезаминирова-ние нуклеотидов
окисление аминов
дезаминирование аминосахаров
метаболические источники аммиака в организме
Слайд 100синтез мочевины
синтез мочевины

Слайд 101•
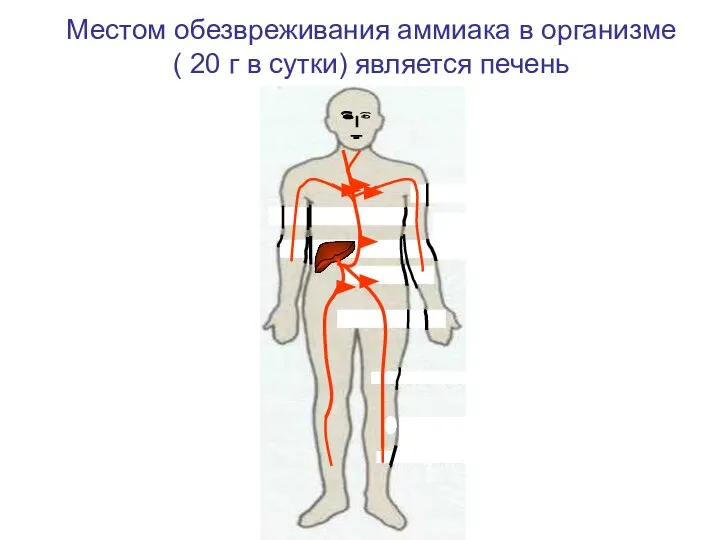
•
Местом обезвреживания аммиака в организме ( 20 г в сутки) является печень
•
•
Местом обезвреживания аммиака в организме ( 20 г в сутки) является печень
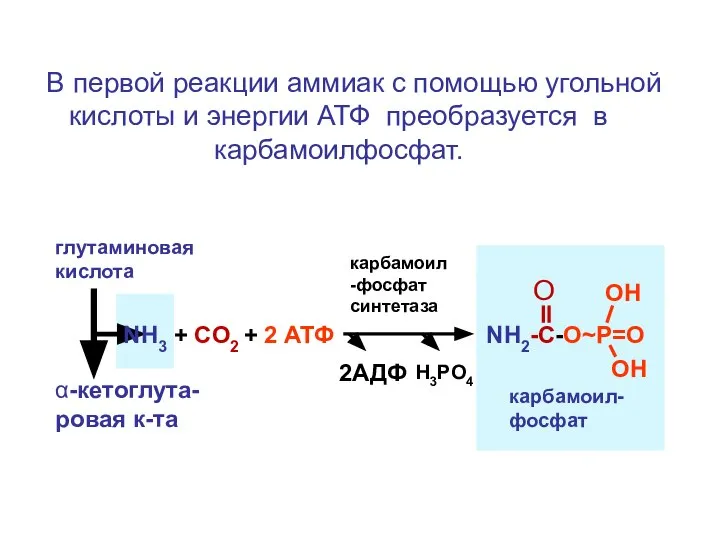
Слайд 102 В первой реакции аммиак с помощью угольной кислоты и энергии АТФ
В первой реакции аммиак с помощью угольной кислоты и энергии АТФ
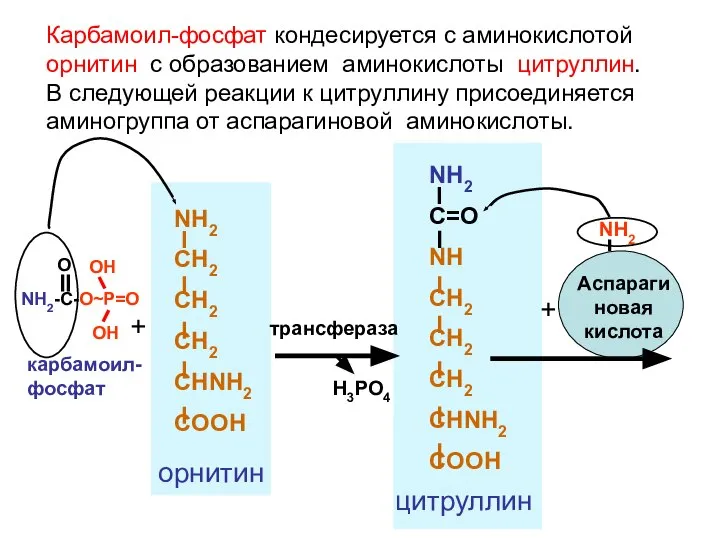
Слайд 103+
NH2
CH2
CH2
CH2
CHNH2
COOH
орнитин
трансфераза
H3PO4
NH2
C=O
NH
CH2
CH2
CH2
CHNH2
COOH
цитруллин
Аспарагиновая кислота
+
Карбамоил-фосфат кондесируется с аминокислотой орнитин с образованием аминокислоты цитруллин.
В следующей реакции
+
NH2
CH2
CH2
CH2
CHNH2
COOH
орнитин
трансфераза
H3PO4
NH2
C=O
NH
CH2
CH2
CH2
CHNH2
COOH
цитруллин
Аспарагиновая кислота
+
Карбамоил-фосфат кондесируется с аминокислотой орнитин с образованием аминокислоты цитруллин.
В следующей реакции
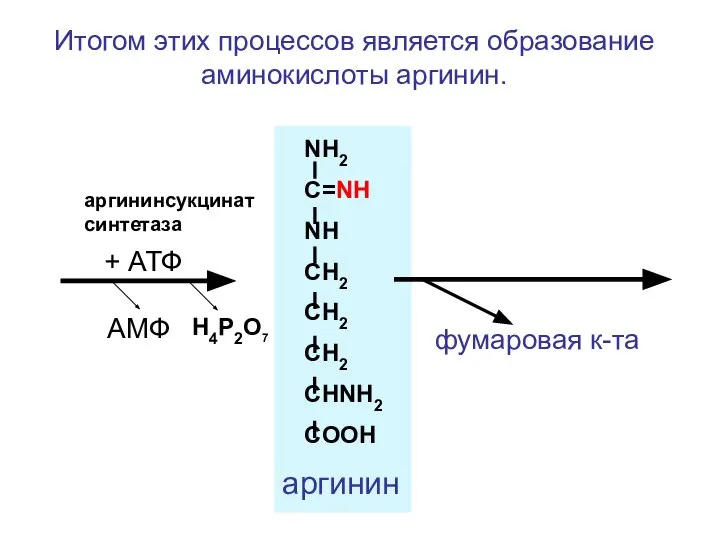
Слайд 104NH2
C=NH
NH
CH2
CH2
CH2
CHNH2
COOH
фумаровая к-та
+ АТФ
аргининсукцинатсинтетаза
АМФ
H4P2O7
аргинин
Итогом этих процессов является образование аминокислоты аргинин.
NH2
C=NH
NH
CH2
CH2
CH2
CHNH2
COOH
фумаровая к-та
+ АТФ
аргининсукцинатсинтетаза
АМФ
H4P2O7
аргинин
Итогом этих процессов является образование аминокислоты аргинин.
Слайд 105NH2
CH2
CH2
CH2
CHNH2
COOH
C=O
NH2
NH2
мочевина
NH2
C=NH
NH
CH2
CH2
CH2
CHNH2
COOH
аргинин
орнитин
аргиназа
+
Н2О
Из приведенной ниже схемы видно, что от аргинина путем гидролиза отщепляется
NH2
CH2
CH2
CH2
CHNH2
COOH
C=O
NH2
NH2
мочевина
NH2
C=NH
NH
CH2
CH2
CH2
CHNH2
COOH
аргинин
орнитин
аргиназа
+
Н2О
Из приведенной ниже схемы видно, что от аргинина путем гидролиза отщепляется
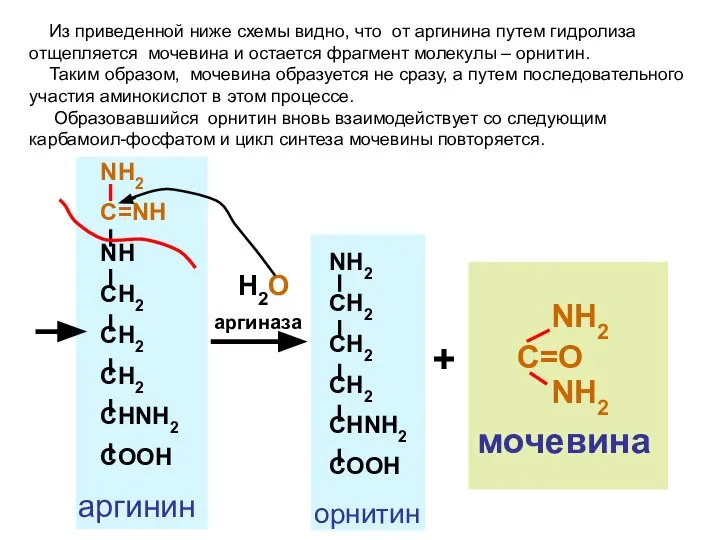
Слайд 106NH3 + CO2 + 2 АТФ
NH2-C-O~P=O
O
OH
OH
орнитин
цитруллин
аспарагино-вая кислота
аргинин
фумаро-вая к-та
мочевина
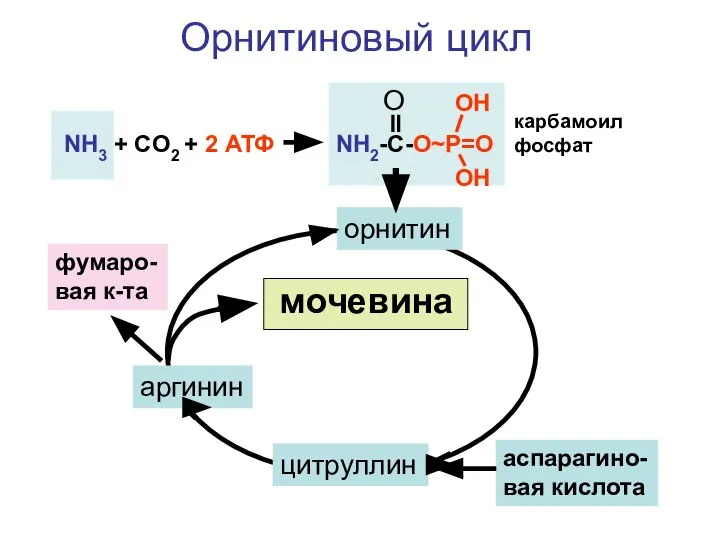
Орнитиновый цикл
карбамоилфосфат
NH3 + CO2 + 2 АТФ
NH2-C-O~P=O
O
OH
OH
орнитин
цитруллин
аспарагино-вая кислота
аргинин
фумаро-вая к-та
мочевина
Орнитиновый цикл
карбамоилфосфат
Слайд 107Содержание мочевины в крови
2,5 – 8,3 ммоль/л сыворотки
Содержание мочевины в крови
2,5 – 8,3 ммоль/л сыворотки

Слайд 108Диагностическая значимость определения концентрации мочевины в крови.
Понижение ниже нормы наблюдается при:
тяжелом
Диагностическая значимость определения концентрации мочевины в крови.
Понижение ниже нормы наблюдается при:
тяжелом

Слайд 109Вспомогательный, быстрый механизм связывания аммиака внутри клеток
Вспомогательный, быстрый механизм связывания аммиака внутри клеток

Слайд 110 Этот пример удаления аммиака в клетках происходит в тех органах, где
Этот пример удаления аммиака в клетках происходит в тех органах, где

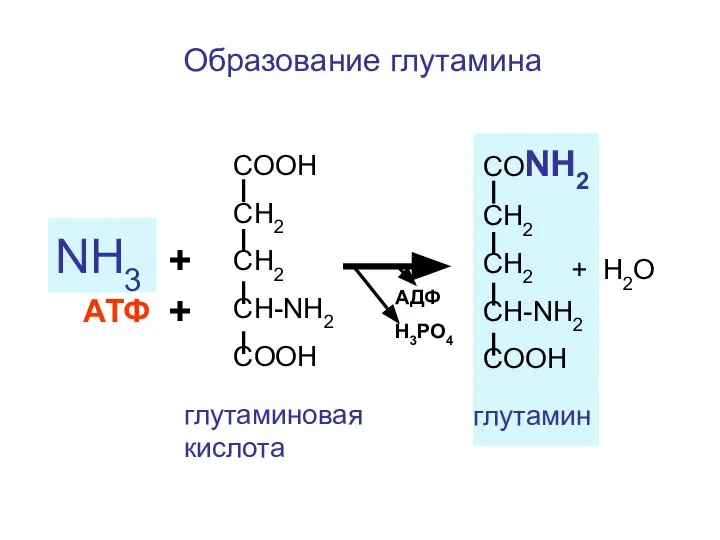
Слайд 111Образование глутамина
СООН
СН2
СН2
CH-NH2
COOH
NH3
+
СОNH2
СН2
СН2
CH-NH2
COOH
АТФ
+
+ H2O
глутаминовая кислота
АДФ
Н3РО4
глутамин
Образование глутамина
СООН
СН2
СН2
CH-NH2
COOH
NH3
+
СОNH2
СН2
СН2
CH-NH2
COOH
АТФ
+
+ H2O
глутаминовая кислота
АДФ
Н3РО4
глутамин
Слайд 112 С током крови глутамин поступает в печень или в почки. В
С током крови глутамин поступает в печень или в почки. В

Слайд 113СООН
СН2
СН2
CH-NH2
COOH
NH3
СОNH2
СН2
СН2
CH-NH2
COOH
+ H2O
глутамино-вая кислота
глутамин
глутаминаза
моча
возвраще-ние в кровь
в почки
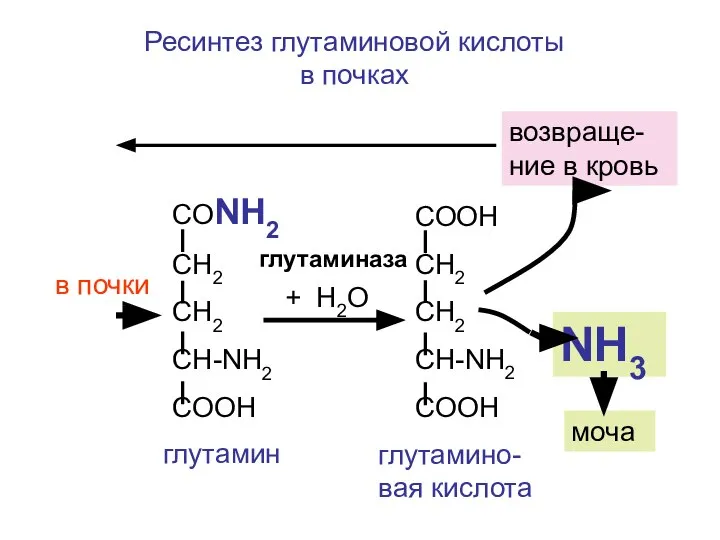
Ресинтез глутаминовой кислоты в почках
СООН
СН2
СН2
CH-NH2
COOH
NH3
СОNH2
СН2
СН2
CH-NH2
COOH
+ H2O
глутамино-вая кислота
глутамин
глутаминаза
моча
возвраще-ние в кровь
в почки
Ресинтез глутаминовой кислоты в почках
 Строение клетки
Строение клетки Теория катастроф Ж. Кювье
Теория катастроф Ж. Кювье Репликация ДНК
Репликация ДНК Жизнедеятельность бактерий. Питание
Жизнедеятельность бактерий. Питание Биология в естествознании
Биология в естествознании Ботаника. Введение. Корень
Ботаника. Введение. Корень 3. Урок - РНК и АТФ
3. Урок - РНК и АТФ Мутационная изменчивость
Мутационная изменчивость Урбопопуляции птиц
Урбопопуляции птиц Создание природного индикатора для определения pH-среды
Создание природного индикатора для определения pH-среды Изготовление кровяной муки
Изготовление кровяной муки Царство бактерии
Царство бактерии Кровь (физиология системы крови) - Презентация
Кровь (физиология системы крови) - Презентация Презентация на тему Класс Двудольные, семейство Паслёновые
Презентация на тему Класс Двудольные, семейство Паслёновые  Ткани растений. Виды тканей. Обмен веществ
Ткани растений. Виды тканей. Обмен веществ Женская половая система - 1. Яичник. Жёлтое тело
Женская половая система - 1. Яичник. Жёлтое тело Презентация на тему Віруси. Пріони. Бактерії
Презентация на тему Віруси. Пріони. Бактерії  Отдел Покрытосеменные Класс Двудольные
Отдел Покрытосеменные Класс Двудольные Элемент металлического происхождения марганец
Элемент металлического происхождения марганец Fiziologia_1
Fiziologia_1 Биоэлектрогенез
Биоэлектрогенез Высшая нервная деятельность
Высшая нервная деятельность Животные и растения степей
Животные и растения степей Презентация на тему Зона степей (4 класс)
Презентация на тему Зона степей (4 класс)  Класс млекопитающие или звери
Класс млекопитающие или звери Репарация
Репарация Презентация на тему Главные части и органоиды клеток
Презентация на тему Главные части и органоиды клеток  Лишайники. Строение и питание лишайников
Лишайники. Строение и питание лишайников