- Общая физиология центральной нервной системы

Содержание
- 2. План лекции: Основные физиологические свойства нервных центров. Особенности распространения возбуждения в ЦНС Торможение в ЦНС. Природа
- 3. – это совокупность нейронов, необходимых для осуществления определенного рефлекса или регуляции определенной функции (центр дыхания, сосудодвигательный
- 4. СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ Суммация возбуждения – сочетание двух или нескольких подпороговых раздражителей вызывает ответ, в то
- 5. Временная суммация Последовательная или временная суммация (имеет место при взаимодействии подпороговых раздражителей приходящих за короткий промежуток
- 6. Пространственная суммация Пространственная суммация – если два или более подпороговых раздражителей действуют одновременно на разные рецепторы
- 7. ТРАНСФОРМАЦИЯ РИТМА Частота импульсов от ЦНС к рабочему органу относительно независимо от частоты раздражения, т.е. в
- 8. ТРАНСФОРМАЦИЯ РИТМА Трансформация ритма возможна и в сторону уменьшения. Это объясняется снижением возбудимости нервного цента за
- 9. Посттетаническая потенциация Раздражая стимулами редкой частоты афферентный нерв, можно получить некоторый рефлекс определенной интенсивности. Если затем
- 10. Посттетаническая потенциация Са2+ Са2+
- 11. Инерционность – способность длительно удерживать возбуждение после окончания действия стимула. Низкая лабильность и высокая утомляемость НЦ.
- 12. Избирательная чувствительность к химическим веществам. По сравнению с другими тканями нейроны обладают высокой избирательной чувствительностью ко
- 13. Тонус НЦ. Тонус, или наличие определенной фоновой активности НЦ, определяется тем, что в покое в отсутствие
- 14. Пластичность — это способность НЦ изменять свое прямое функциональное назначение и расширять свои функциональные возможности. Это
- 15. Пластичность синапса Это функциональная и морфологическая перестройка синапса: Увеличение пластичности: облегчение (пресинаптическая природа, Са++), потенциация (постсинаптическая
- 16. Долговременные формы пластичности Долговременная потенциация – длительное усиление синаптической передачи на выскочастотное раздражение, может продолжаться дни
- 17. Особенности распространения возбуждения в ЦНС
- 18. 1. Дивергенция (мультипликация) — возможность нейрона устанавливать многочисленные синаптические связи с другими нейронами, т.е. одна клетка
- 19. 3. Одностороннее проведение возбуждения. Через НЦ передача возбуждения возможна от рецепторного нейрона к вставочному или эфферентному,
- 20. Время рефлекса Центральное время рефлекса Время рефлекса (латентный период рефлекса) – это время от момента нанесения
- 21. Центральное время рефлекса– это промежуток времени, за который нервный импульс проводится по структурам мозга. В случае
- 22. Коленный рефлекс Моносинаптический. В результате резкого растяжения проприорецепторов четырехглавой мышцы происходит разгибание голени (- оборонительный проприорецептивный
- 23. Реверберация. Рефлекторные акты заканчиваются не всегда прекращением действия возбуждения, а через некоторое время, что связано с
- 24. Р Е В Е Р Б Е Р А Ц И Я (циркуляция)
- 25. Торможение – активный самостоятельный физиологический процесс, вызываемый возбуждением и направленный на ослабление, прекращение или предотвращение другого
- 26. Т о р м о ж е н и е Торможение нервных клеток, центров – равный
- 27. Функции торможения Ограничивает распространение возбуждения в ЦНС (иррадиацию, реверберацию, мультипликацию и др. ) и способствует его
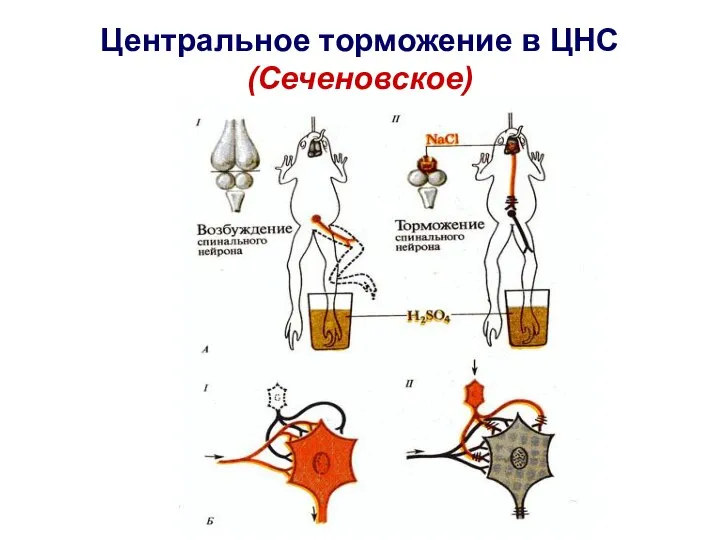
- 28. Центральное торможение открыл И.М. Сеченов в 1863 г
- 29. У лягушки Сеченов И.М. удалял большие полушария на уровне зрительных бугров. После чего раздражал заднюю лапку
- 30. Центральное торможение в ЦНС (Сеченовское)
- 31. Классификация торможения в ЦНС Различают: Первичное торможение. Не связанно с первоначальным процессом возбуждения, развивается в результате
- 32. Классификация торможения в ЦНС По электрическому состоянию постсинаптической мембраны: гиперполяризационное деполяризационное По отношению к синапсу: постсинаптическое
- 33. Биоэлектрическая активность нейрона
- 34. Т о р м о з н ы е медиаторы - Возникновение ТПСП в ответ на
- 35. Ионная природа постсинаптического торможения Постсинаптическое торможение. Оно развивается в ответ на действие тормозного медиатора в синапсе
- 36. ПОСТСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ ГАМК ТПСП Cl К Если МП постсинаптической мембраны менее отрицательный(50-60 мВ), то Cl входит
- 37. Пресинаптическое торможение Осуществляется посредством специальных тормозных интернейронов. Его структурной основой являются аксо-аксональные синапсы, образованные терминалиями аксонов
- 38. Пресинаптическое торможение Тормозной аксон высвобождает тормозящий медиатор, который увеличивает проницаемость мембраны возбуждающего аксона для ионов хлора.
- 39. Афферентные импульсы от мышцы – сгибателя с помощью клеток Реншоу вызывают пресинаптическое торможение на афферентном нерве,
- 40. Классификация видов Первичное постсинаптическое торможение: а) Центральное (Сеченовское) торможение. б) Корковое в) Возвратное торможение г) Реципрокное
- 41. Возвратное (антидромное) торможение 1. Антидромное (возвратное) импульсы от мотонейрона одновременно с активацией мышцы, через коллатералии аксона,
- 42. Реципрокное торможение – на уровне сегментов спинного мозга Основано на том, что одни и те же
- 43. Латеральное торможение Вставочные нейроны способны влиять не только на возбуждающую клетку, но и на соседние, где
- 44. В - возбуждение Т - торможение ТОРМОЖЕНИЕ В ЦНС Латеральное торможение Возвратное(антидромное) торможение по Реншоу По
- 45. ВТОРИЧНОЕ ТОРМОЖЕНИЕ Запредельное торможение или охранительное. возникает на раздражение превышающее предел работоспособности нейронов. Оно препятствует истощению
- 46. Примеры нарушения торможения в ЦНС НАРУШЕНИЕ ПОСТСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯ: СТРИХНИН - блокада рецепторов тормозных синапсов. СТОЛБНЯЧНЫЙ ТОКСИН
- 47. Постсинаптическое возвратное торможение.. Блокируется стрихнином.
- 48. Пресинаптическое торможение. Блокируется пикротоксином
- 49. ПРИНЦИПЫ КООРДИНАЦИИ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ
- 50. Осуществление каждого рефлекса зависит от: – состояния ЦНС в данный момент времени – совокупности межцентральных соотношений
- 51. Принцип реципрокности. В опыте на животных он стимулировал определенные точки двигательной зоны коры и установил, что
- 52. Шагательный рефлекс Примеры рефлекторных дуг А. непрерывное возбуждение двигательных центров ЦНС разбивается на поочередные акты возбуждение
- 53. Принцип реципрокной (взаимосочетанной) иннервации
- 54. Индукция. Индукцией называется свойство основного нервного процесса (возбуждения и торможения) вызывать вокруг себя и после себя
- 55. Взаимотормозящие влияния рефлексов, основанные на реципрокном торможении, называются одновременной отрицательной индукцией. Сильное и длительное раздражение возбуждает
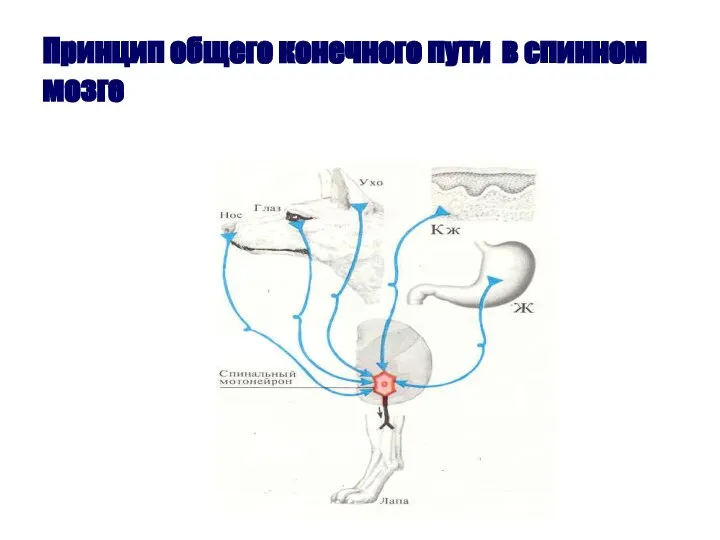
- 56. Принцип общего «конечного пути». Открыт английским физиологом Ч. Шеррингтоном (1906). Один и тот же рефлекс (например,
- 57. ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ КОРА ПОДКОРКА СТВОЛ СПИННОЙ МОЗГ
- 58. Принцип общего конечного пути в спинном мозге
- 59. Принцип обратной связи. Любой рефлекторный акт контролируется благодаря обратной связи с центром. Обратная связь состоит во
- 60. Различают обратную связь положительную (усиливающую рефлекс, который является источником обратной афферентации) и отрицательную, когда рефлекс, ее
- 61. ПРИНЦИП ОБРАТНОЙ АФФЕРЕНТАЦИИ ДВИГАТЕЛЬНАЯ КОМАНДА ОБРАТНАЯ АФФЕРЕНТАЦИЯ
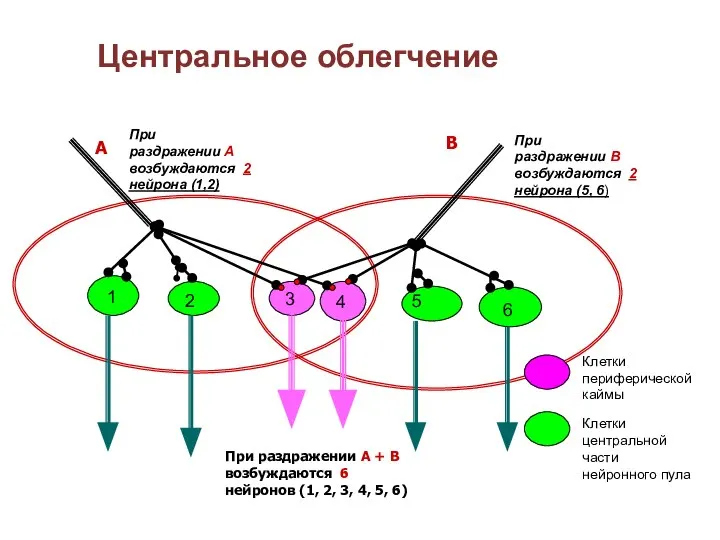
- 62. По окончании реакции на раздражитель возбуждение некоторое время остается повышенным и поэтому порог раздражения понижается, а
- 63. Центральное облегчение 1 2 3 4 5 6 А В При раздражении А возбуждаются 2 нейрона
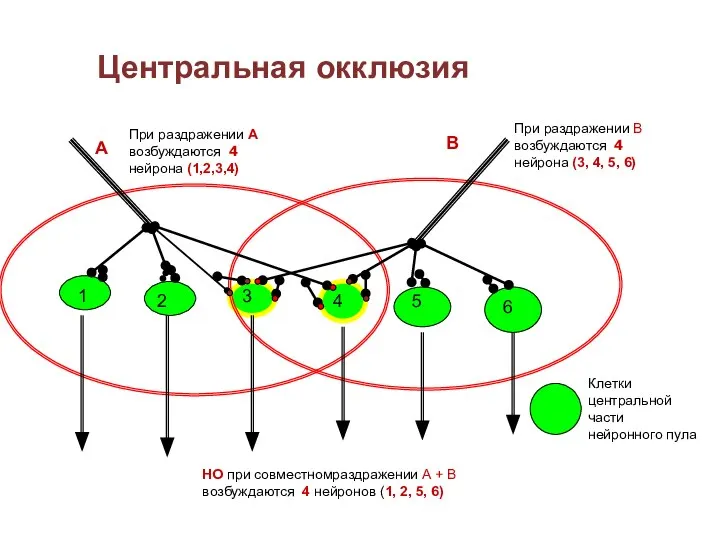
- 64. Окклюзия. Если одновременно раздражать две группы афферентов, каждая из которых дает сильный сократительный рефлекс мышцы, то
- 65. Центральная окклюзия 1 2 3 4 5 6 А В При раздражении А возбуждаются 4 нейрона
- 66. Принцип Доминанты Алексей Алексеевич Ухтомский профессор СПбГУ, сформулировал этот принцип
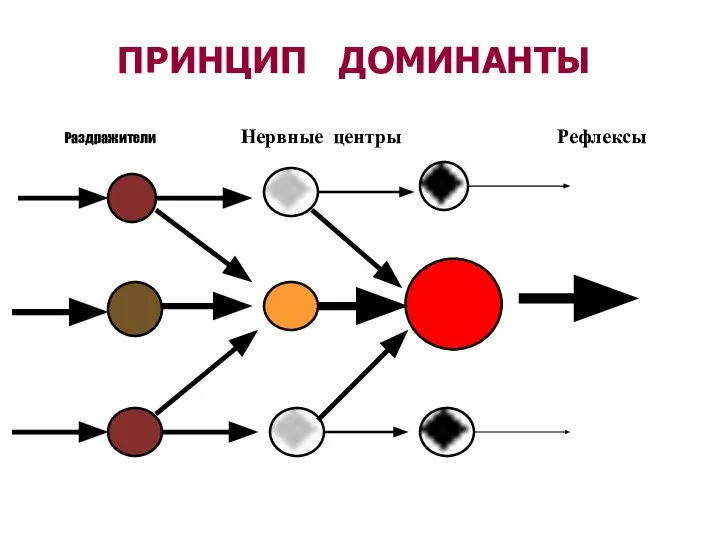
- 67. ОПРЕДЕЛЕНИЕ ДОМИНАНТЫ ( ПО А.А.Ухтомскому, 1931) ДОМИНАНТА - временно господствующий очаг возбуждения в центральной нервной системе,
- 68. Появление доминанты ведет к тому, что раздражение любых рецепторов будет вызывать ответ характерный для данной доминанты.
- 69. ПРИНЦИП ДОМИНАНТЫ Раздражители Нервные центры Рефлексы
- 70. Основные признаки доминанты ( по А.А.Ухтомскому) 1. Повышенная возбудимость доминантного центра 2. Стойкость возбуждения в доминантном
- 71. ПРИНЦИП СУБОРДИНАЦИИ НЕРВНЫХ ЦЕНТРОВ Принцип субординации нервных центров (принцип подчинения) проявляется в виде регулирующего влияния выше
- 73. Скачать презентацию
Слайд 2План лекции:
Основные физиологические свойства нервных центров.
Особенности распространения возбуждения в ЦНС
Торможение в
План лекции:
Основные физиологические свойства нервных центров.
Особенности распространения возбуждения в ЦНС
Торможение в

Слайд 3– это совокупность нейронов, необходимых для осуществления определенного рефлекса или регуляции определенной
– это совокупность нейронов, необходимых для осуществления определенного рефлекса или регуляции определенной

Слайд 4СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
Суммация возбуждения – сочетание двух или нескольких подпороговых раздражителей вызывает
СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
Суммация возбуждения – сочетание двух или нескольких подпороговых раздражителей вызывает

Слайд 5Временная суммация
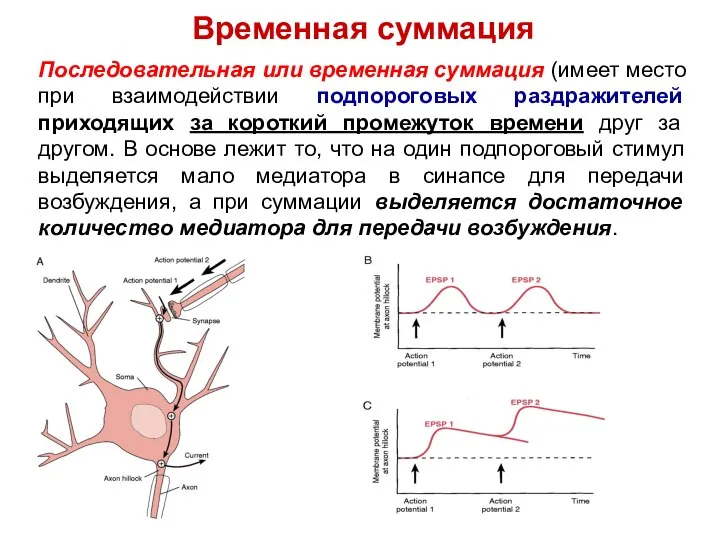
Последовательная или временная суммация (имеет место при взаимодействии подпороговых раздражителей приходящих
Временная суммация
Последовательная или временная суммация (имеет место при взаимодействии подпороговых раздражителей приходящих
Слайд 6Пространственная суммация
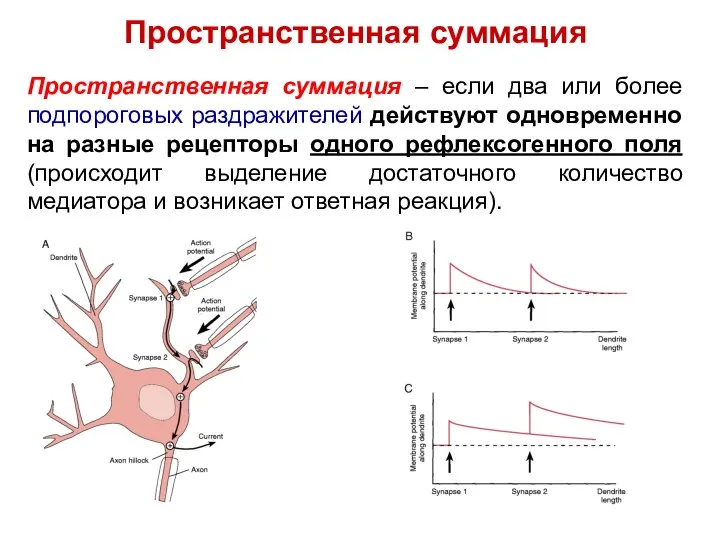
Пространственная суммация – если два или более подпороговых раздражителей действуют одновременно
Пространственная суммация
Пространственная суммация – если два или более подпороговых раздражителей действуют одновременно
Слайд 7ТРАНСФОРМАЦИЯ РИТМА
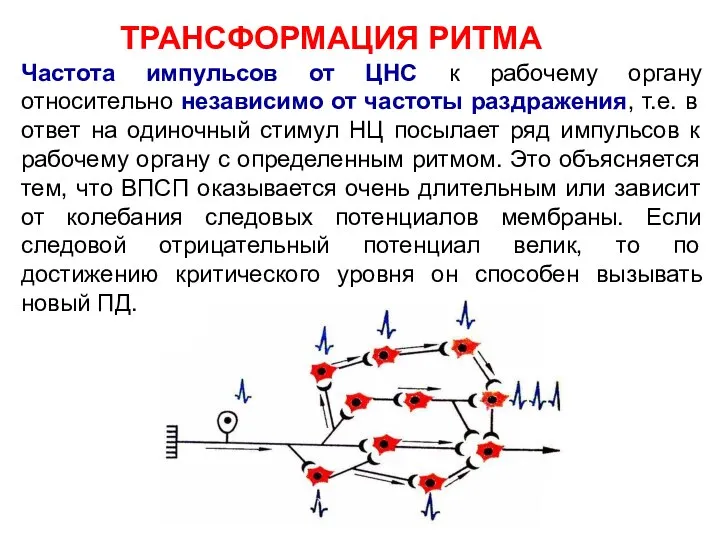
Частота импульсов от ЦНС к рабочему органу относительно независимо от частоты
ТРАНСФОРМАЦИЯ РИТМА
Частота импульсов от ЦНС к рабочему органу относительно независимо от частоты
Слайд 8ТРАНСФОРМАЦИЯ РИТМА
Трансформация ритма возможна и в сторону уменьшения. Это объясняется снижением возбудимости
ТРАНСФОРМАЦИЯ РИТМА
Трансформация ритма возможна и в сторону уменьшения. Это объясняется снижением возбудимости

Слайд 9Посттетаническая потенциация
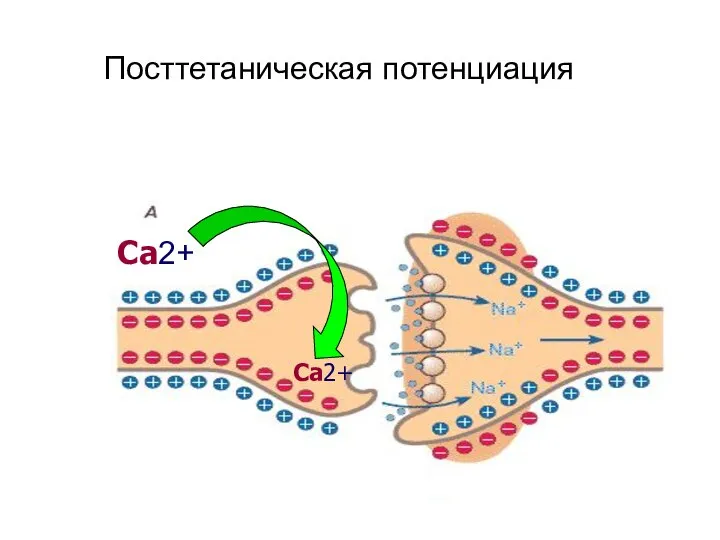
Раздражая стимулами редкой частоты афферентный нерв, можно получить некоторый рефлекс определенной
Посттетаническая потенциация
Раздражая стимулами редкой частоты афферентный нерв, можно получить некоторый рефлекс определенной

Слайд 10Посттетаническая потенциация
Са2+
Са2+
Посттетаническая потенциация
Са2+
Са2+
Слайд 11Инерционность – способность длительно удерживать возбуждение после окончания действия стимула.
Низкая лабильность и
Инерционность – способность длительно удерживать возбуждение после окончания действия стимула.
Низкая лабильность и

Слайд 12Избирательная чувствительность к химическим веществам. По сравнению с другими тканями нейроны обладают
Избирательная чувствительность к химическим веществам. По сравнению с другими тканями нейроны обладают

Слайд 13Тонус НЦ. Тонус, или наличие определенной фоновой активности НЦ, определяется тем, что
Тонус НЦ. Тонус, или наличие определенной фоновой активности НЦ, определяется тем, что

Слайд 14Пластичность — это способность НЦ изменять свое прямое функциональное назначение и расширять
Пластичность — это способность НЦ изменять свое прямое функциональное назначение и расширять

Слайд 15Пластичность синапса
Это функциональная и морфологическая перестройка синапса:
Увеличение пластичности: облегчение (пресинаптическая природа,
Пластичность синапса
Это функциональная и морфологическая перестройка синапса:
Увеличение пластичности: облегчение (пресинаптическая природа,

Слайд 16Долговременные формы пластичности
Долговременная потенциация – длительное усиление синаптической передачи на выскочастотное раздражение,
Долговременные формы пластичности
Долговременная потенциация – длительное усиление синаптической передачи на выскочастотное раздражение,

Слайд 17Особенности распространения возбуждения в ЦНС
Особенности распространения возбуждения в ЦНС

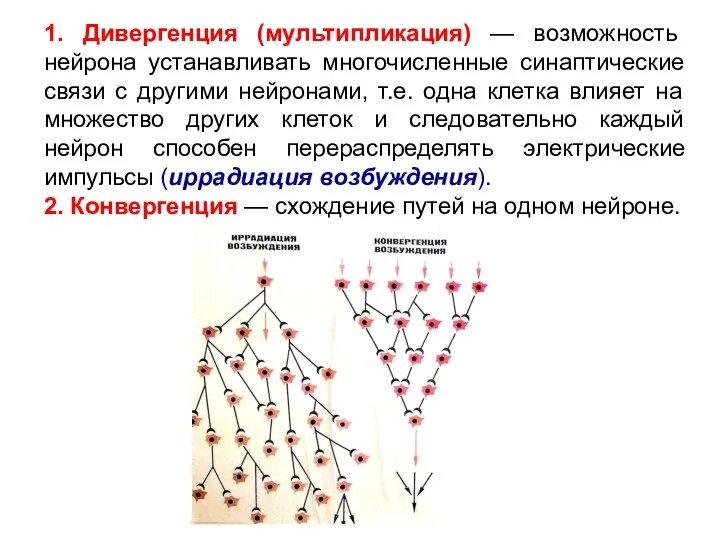
Слайд 181. Дивергенция (мультипликация) — возможность нейрона устанавливать многочисленные синаптические связи с другими
1. Дивергенция (мультипликация) — возможность нейрона устанавливать многочисленные синаптические связи с другими
Слайд 193. Одностороннее проведение возбуждения. Через НЦ передача возбуждения возможна от рецепторного нейрона
3. Одностороннее проведение возбуждения. Через НЦ передача возбуждения возможна от рецепторного нейрона

Слайд 20Время рефлекса
Центральное время рефлекса
Время рефлекса (латентный период рефлекса) – это
Время рефлекса
Центральное время рефлекса
Время рефлекса (латентный период рефлекса) – это

Слайд 21Центральное время рефлекса– это промежуток времени, за который нервный импульс проводится по
Центральное время рефлекса– это промежуток времени, за который нервный импульс проводится по

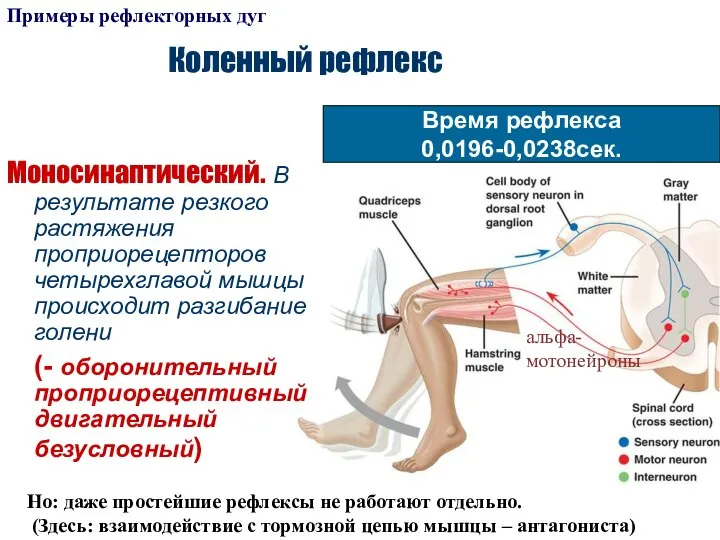
Слайд 22Коленный рефлекс
Моносинаптический. В результате резкого растяжения проприорецепторов четырехглавой мышцы происходит разгибание голени
(-
Коленный рефлекс
Моносинаптический. В результате резкого растяжения проприорецепторов четырехглавой мышцы происходит разгибание голени
(-
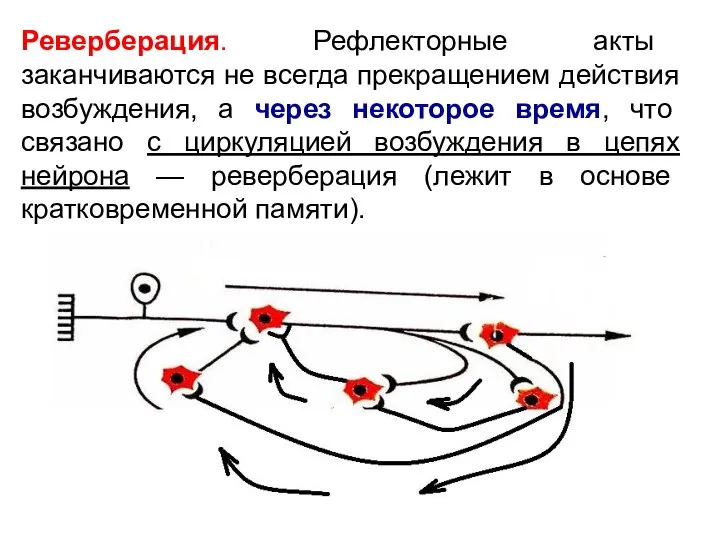
Слайд 23Реверберация. Рефлекторные акты заканчиваются не всегда прекращением действия возбуждения, а через некоторое
Реверберация. Рефлекторные акты заканчиваются не всегда прекращением действия возбуждения, а через некоторое
Слайд 24Р Е В Е Р Б Е Р А Ц И Я
(циркуляция)
Р Е В Е Р Б Е Р А Ц И Я
(циркуляция)
Слайд 25 Торможение – активный самостоятельный физиологический процесс, вызываемый возбуждением и направленный на
Торможение – активный самостоятельный физиологический процесс, вызываемый возбуждением и направленный на

Слайд 26Т о р м о ж е н и е
Торможение нервных клеток,
Т о р м о ж е н и е
Торможение нервных клеток,

Слайд 27Функции торможения
Ограничивает распространение возбуждения в ЦНС (иррадиацию, реверберацию, мультипликацию и др. )
Функции торможения
Ограничивает распространение возбуждения в ЦНС (иррадиацию, реверберацию, мультипликацию и др. )

Слайд 28Центральное торможение
открыл
И.М. Сеченов
в 1863 г
Центральное торможение
открыл
И.М. Сеченов
в 1863 г

Слайд 29У лягушки Сеченов И.М. удалял большие полушария на уровне зрительных бугров. После
У лягушки Сеченов И.М. удалял большие полушария на уровне зрительных бугров. После

Слайд 30Центральное торможение в ЦНС (Сеченовское)
Центральное торможение в ЦНС (Сеченовское)
Слайд 31Классификация торможения в ЦНС
Различают:
Первичное торможение. Не связанно с первоначальным процессом возбуждения, развивается
Классификация торможения в ЦНС
Различают:
Первичное торможение. Не связанно с первоначальным процессом возбуждения, развивается

Слайд 32Классификация
торможения в ЦНС
По электрическому состоянию постсинаптической мембраны: гиперполяризационное
деполяризационное
По отношению
Классификация
торможения в ЦНС
По электрическому состоянию постсинаптической мембраны: гиперполяризационное
деполяризационное
По отношению

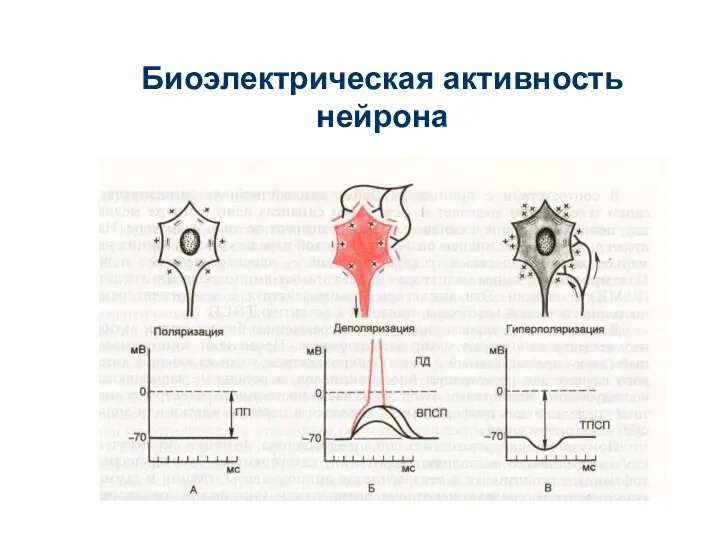
Слайд 33Биоэлектрическая активность нейрона
Биоэлектрическая активность нейрона
Слайд 34Т о р м о з н ы е медиаторы -
Т о р м о з н ы е медиаторы -

Слайд 35
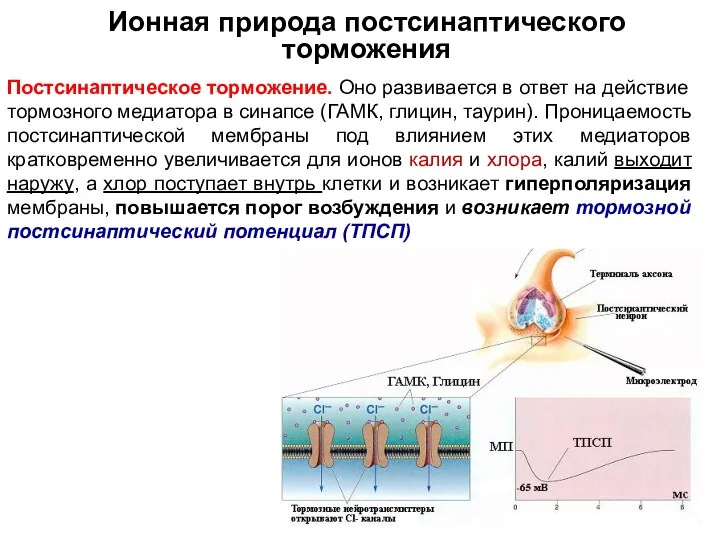
Ионная природа постсинаптического торможения
Постсинаптическое торможение. Оно развивается в ответ на действие тормозного
Ионная природа постсинаптического торможения
Постсинаптическое торможение. Оно развивается в ответ на действие тормозного
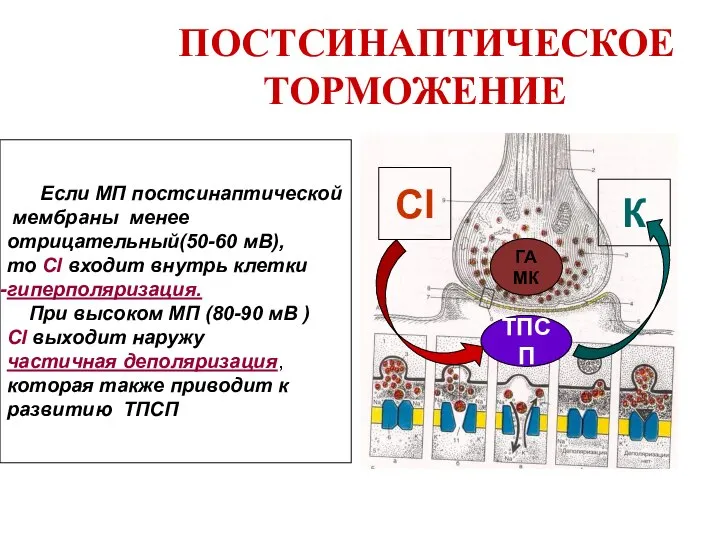
Слайд 36 ПОСТСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
ГАМК
ТПСП
Cl
К
Если МП постсинаптической
мембраны менее
отрицательный(50-60 мВ),
то Cl
ПОСТСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
ГАМК
ТПСП
Cl
К
Если МП постсинаптической
мембраны менее
отрицательный(50-60 мВ),
то Cl
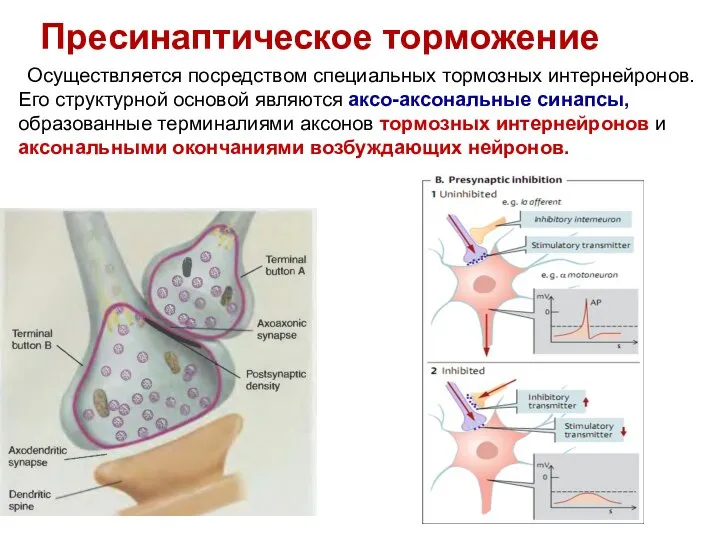
Слайд 37Пресинаптическое торможение
Осуществляется посредством специальных тормозных интернейронов.
Его структурной основой являются аксо-аксональные
Пресинаптическое торможение
Осуществляется посредством специальных тормозных интернейронов.
Его структурной основой являются аксо-аксональные
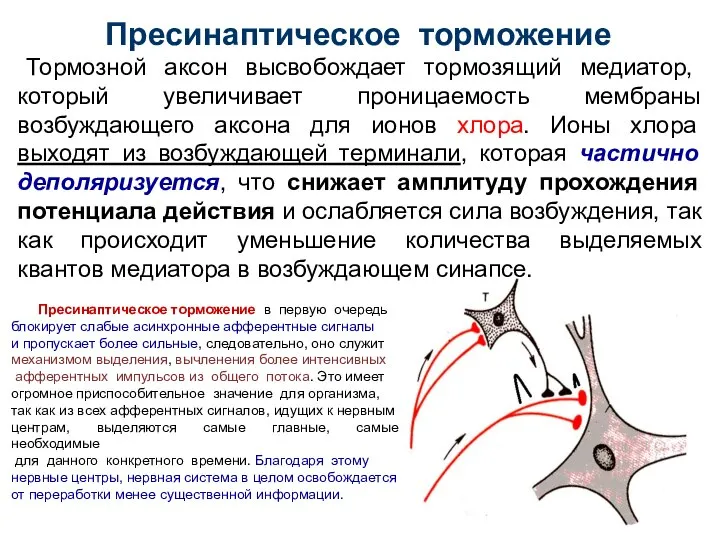
Слайд 38Пресинаптическое торможение
Тормозной аксон высвобождает тормозящий медиатор, который увеличивает проницаемость мембраны возбуждающего
Пресинаптическое торможение
Тормозной аксон высвобождает тормозящий медиатор, который увеличивает проницаемость мембраны возбуждающего
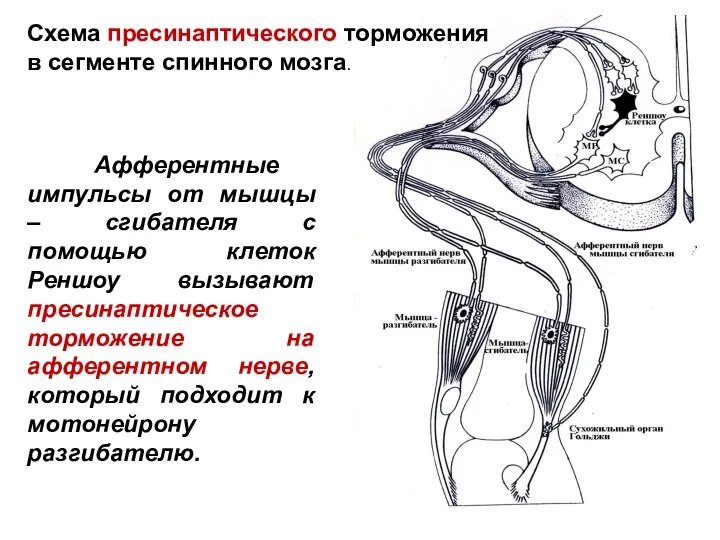
Слайд 39
Афферентные импульсы от мышцы – сгибателя с помощью клеток Реншоу вызывают
Афферентные импульсы от мышцы – сгибателя с помощью клеток Реншоу вызывают
Слайд 40Классификация видов
Первичное постсинаптическое торможение:
а) Центральное (Сеченовское) торможение.
б) Корковое
в) Возвратное торможение
г)
Классификация видов
Первичное постсинаптическое торможение:
а) Центральное (Сеченовское) торможение.
б) Корковое
в) Возвратное торможение
г)

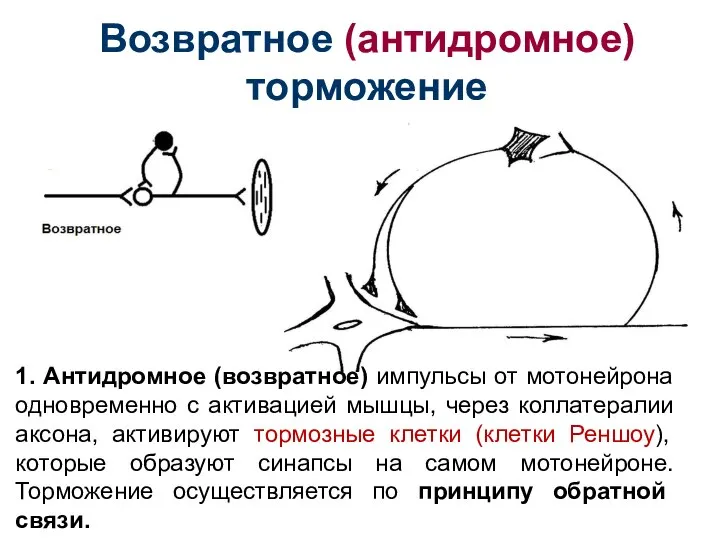
Слайд 41Возвратное (антидромное) торможение
1. Антидромное (возвратное) импульсы от мотонейрона одновременно с активацией мышцы,
Возвратное (антидромное) торможение
1. Антидромное (возвратное) импульсы от мотонейрона одновременно с активацией мышцы,
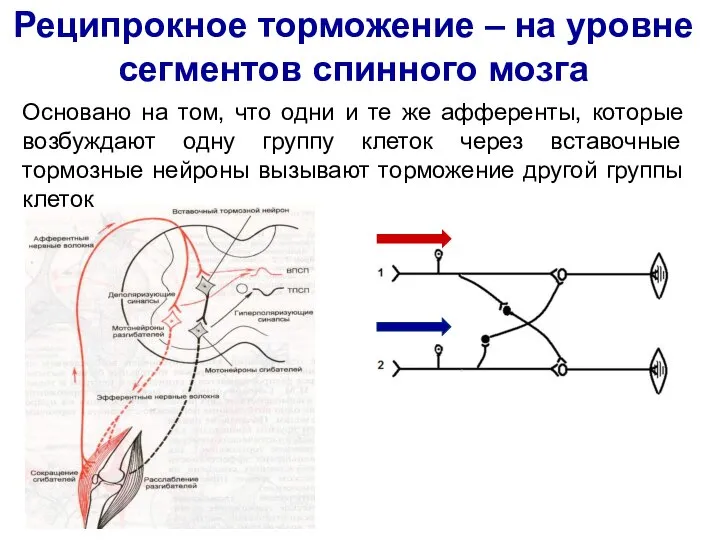
Слайд 42Реципрокное торможение – на уровне сегментов спинного мозга
Основано на том, что одни
Реципрокное торможение – на уровне сегментов спинного мозга
Основано на том, что одни
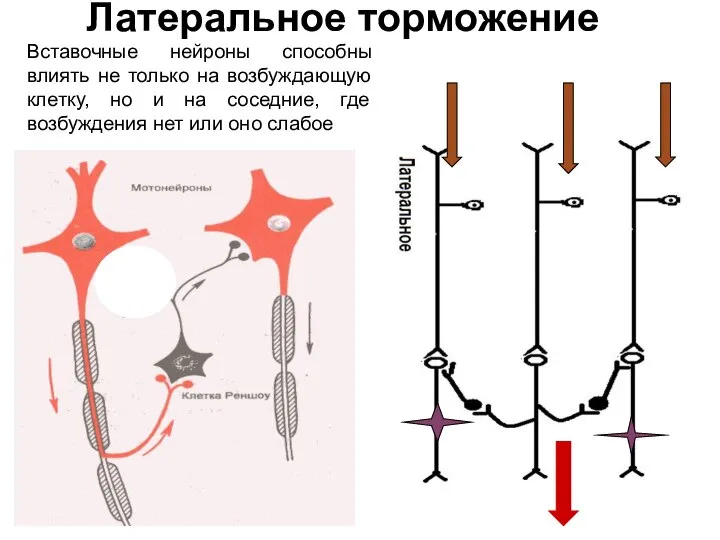
Слайд 43Латеральное торможение
Вставочные нейроны способны влиять не только на возбуждающую клетку, но и
Латеральное торможение
Вставочные нейроны способны влиять не только на возбуждающую клетку, но и
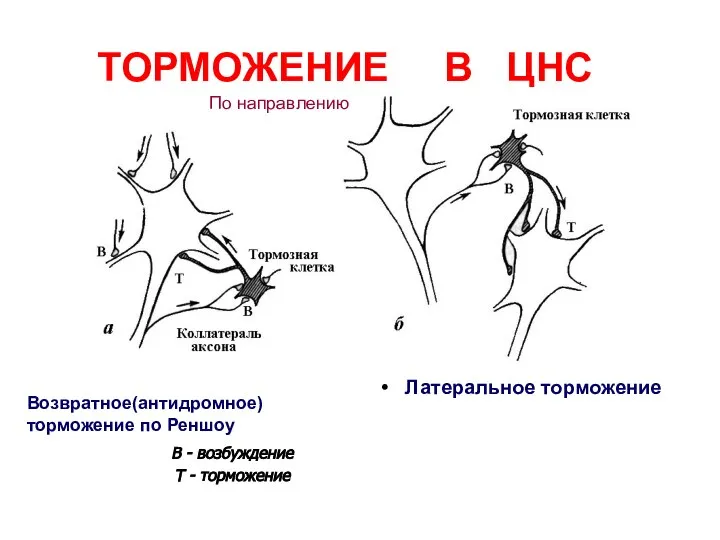
Слайд 44В - возбуждение
Т - торможение
ТОРМОЖЕНИЕ В ЦНС
Латеральное торможение
Возвратное(антидромное) торможение по Реншоу
По направлению
В - возбуждение
Т - торможение
ТОРМОЖЕНИЕ В ЦНС
Латеральное торможение
Возвратное(антидромное) торможение по Реншоу
По направлению
Слайд 45ВТОРИЧНОЕ ТОРМОЖЕНИЕ
Запредельное торможение или охранительное. возникает на раздражение превышающее предел работоспособности нейронов.
ВТОРИЧНОЕ ТОРМОЖЕНИЕ
Запредельное торможение или охранительное. возникает на раздражение превышающее предел работоспособности нейронов.

Слайд 46Примеры нарушения
торможения в ЦНС
НАРУШЕНИЕ ПОСТСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯ:
СТРИХНИН - блокада рецепторов тормозных
Примеры нарушения
торможения в ЦНС
НАРУШЕНИЕ ПОСТСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯ:
СТРИХНИН - блокада рецепторов тормозных

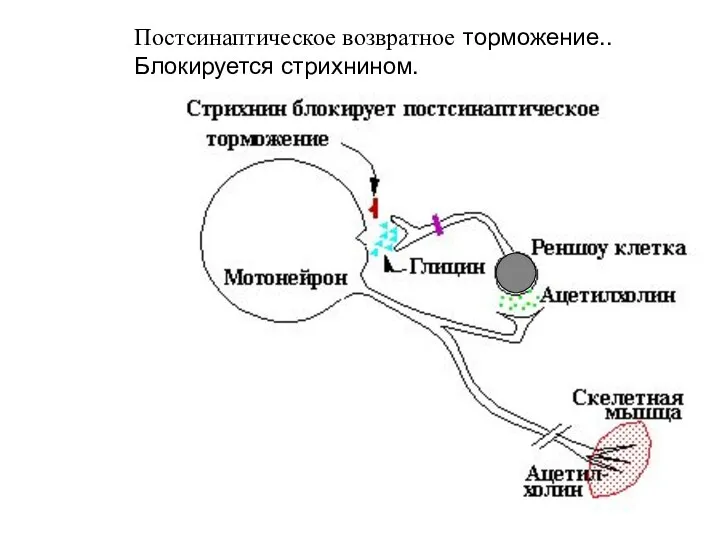
Слайд 47Постсинаптическое возвратное торможение.. Блокируется стрихнином.
Постсинаптическое возвратное торможение.. Блокируется стрихнином.
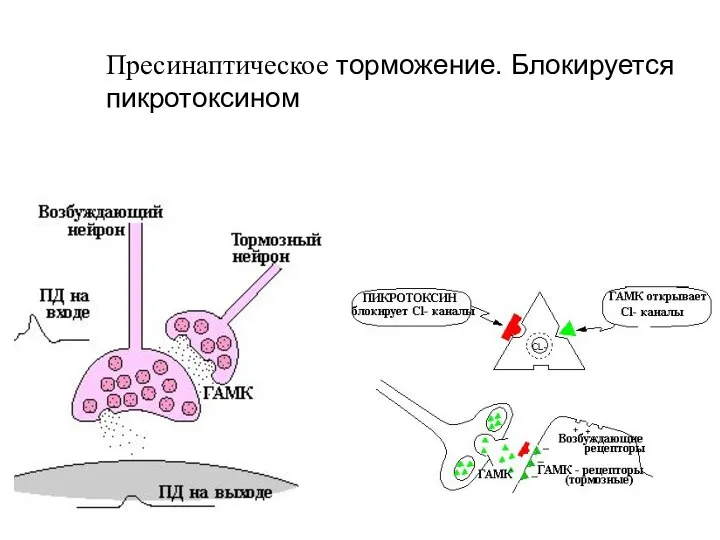
Слайд 48Пресинаптическое торможение. Блокируется пикротоксином
Пресинаптическое торможение. Блокируется пикротоксином
Слайд 49ПРИНЦИПЫ КООРДИНАЦИИ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ
ПРИНЦИПЫ КООРДИНАЦИИ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ

Слайд 50 Осуществление каждого рефлекса зависит от:
– состояния ЦНС в данный момент времени
Осуществление каждого рефлекса зависит от:
– состояния ЦНС в данный момент времени

Слайд 51Принцип реципрокности. В опыте на животных он стимулировал определенные точки двигательной зоны
Принцип реципрокности. В опыте на животных он стимулировал определенные точки двигательной зоны

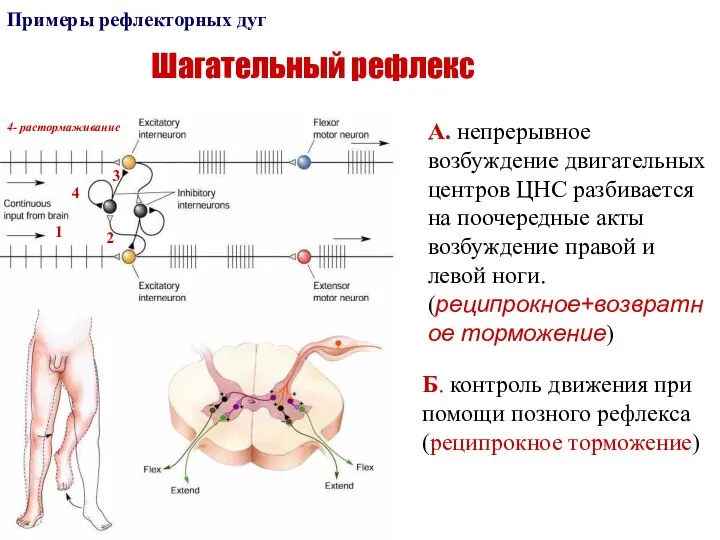
Слайд 52Шагательный рефлекс
Примеры рефлекторных дуг
А. непрерывное возбуждение двигательных центров ЦНС разбивается на поочередные
Шагательный рефлекс
Примеры рефлекторных дуг
А. непрерывное возбуждение двигательных центров ЦНС разбивается на поочередные
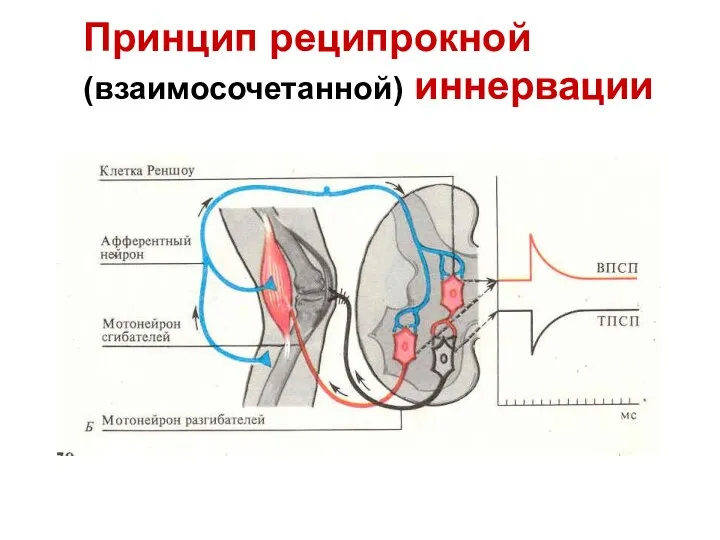
Слайд 53Принцип реципрокной (взаимосочетанной) иннервации
Принцип реципрокной (взаимосочетанной) иннервации
Слайд 54Индукция. Индукцией называется свойство основного нервного процесса (возбуждения и торможения) вызывать вокруг
Индукция. Индукцией называется свойство основного нервного процесса (возбуждения и торможения) вызывать вокруг

Слайд 55Взаимотормозящие влияния рефлексов, основанные на реципрокном торможении, называются одновременной отрицательной индукцией.
Сильное и
Взаимотормозящие влияния рефлексов, основанные на реципрокном торможении, называются одновременной отрицательной индукцией.
Сильное и

Слайд 56Принцип общего «конечного пути». Открыт английским физиологом Ч. Шеррингтоном (1906). Один и
Принцип общего «конечного пути». Открыт английским физиологом Ч. Шеррингтоном (1906). Один и

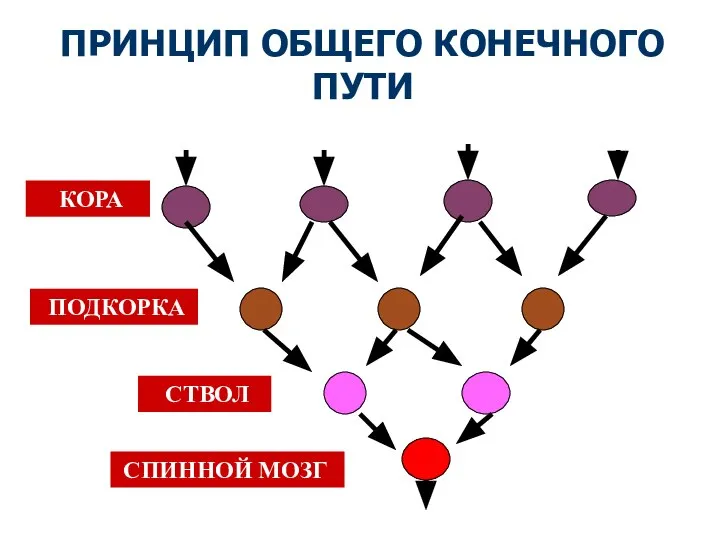
Слайд 57ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ
КОРА
ПОДКОРКА
СТВОЛ
СПИННОЙ МОЗГ
ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ
КОРА
ПОДКОРКА
СТВОЛ
СПИННОЙ МОЗГ
Слайд 58Принцип общего конечного пути в спинном мозге
Принцип общего конечного пути в спинном мозге
Слайд 59Принцип обратной связи. Любой рефлекторный акт контролируется благодаря обратной связи с центром.
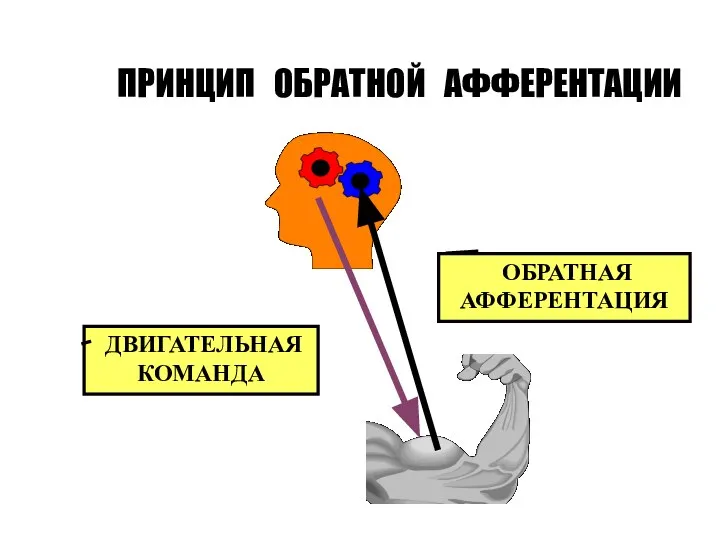
Принцип обратной связи. Любой рефлекторный акт контролируется благодаря обратной связи с центром.

Слайд 60Различают обратную связь положительную (усиливающую рефлекс, который является источником обратной афферентации) и
Различают обратную связь положительную (усиливающую рефлекс, который является источником обратной афферентации) и

Слайд 61ПРИНЦИП ОБРАТНОЙ АФФЕРЕНТАЦИИ
ДВИГАТЕЛЬНАЯ КОМАНДА
ОБРАТНАЯ
АФФЕРЕНТАЦИЯ
ПРИНЦИП ОБРАТНОЙ АФФЕРЕНТАЦИИ
ДВИГАТЕЛЬНАЯ КОМАНДА
ОБРАТНАЯ
АФФЕРЕНТАЦИЯ
Слайд 62По окончании реакции на раздражитель возбуждение некоторое время остается повышенным и поэтому
По окончании реакции на раздражитель возбуждение некоторое время остается повышенным и поэтому

Слайд 63Центральное облегчение
1
2
3
4
5
6
А
В
При раздражении А возбуждаются 2 нейрона (1,2)
При раздражении В возбуждаются 2
Центральное облегчение
1
2
3
4
5
6
А
В
При раздражении А возбуждаются 2 нейрона (1,2)
При раздражении В возбуждаются 2
Слайд 64Окклюзия. Если одновременно раздражать две группы афферентов, каждая из которых дает сильный
Окклюзия. Если одновременно раздражать две группы афферентов, каждая из которых дает сильный

Слайд 65Центральная окклюзия
1
2
3
4
5
6
А
В
При раздражении А возбуждаются 4 нейрона (1,2,3,4)
При раздражении В возбуждаются 4
Центральная окклюзия
1
2
3
4
5
6
А
В
При раздражении А возбуждаются 4 нейрона (1,2,3,4)
При раздражении В возбуждаются 4
Слайд 66Принцип
Доминанты
Алексей Алексеевич
Ухтомский
профессор СПбГУ,
сформулировал
этот принцип
Принцип
Доминанты
Алексей Алексеевич
Ухтомский
профессор СПбГУ,
сформулировал
этот принцип

Слайд 67ОПРЕДЕЛЕНИЕ ДОМИНАНТЫ
( ПО А.А.Ухтомскому, 1931)
ДОМИНАНТА - временно господствующий очаг возбуждения в центральной
ОПРЕДЕЛЕНИЕ ДОМИНАНТЫ
( ПО А.А.Ухтомскому, 1931)
ДОМИНАНТА - временно господствующий очаг возбуждения в центральной

Слайд 68Появление доминанты ведет к тому, что раздражение любых рецепторов будет вызывать ответ
Появление доминанты ведет к тому, что раздражение любых рецепторов будет вызывать ответ

Слайд 69ПРИНЦИП ДОМИНАНТЫ
Раздражители Нервные центры Рефлексы
ПРИНЦИП ДОМИНАНТЫ
Раздражители Нервные центры Рефлексы
Слайд 70Основные признаки доминанты
( по А.А.Ухтомскому)
1. Повышенная возбудимость доминантного центра
2. Стойкость возбуждения в
Основные признаки доминанты
( по А.А.Ухтомскому)
1. Повышенная возбудимость доминантного центра
2. Стойкость возбуждения в

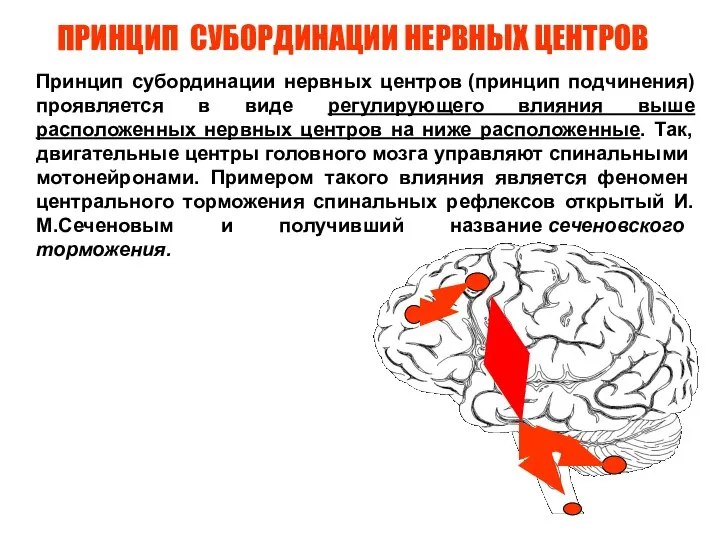
Слайд 71ПРИНЦИП СУБОРДИНАЦИИ НЕРВНЫХ ЦЕНТРОВ
Принцип субординации нервных центров (принцип подчинения) проявляется в виде регулирующего
ПРИНЦИП СУБОРДИНАЦИИ НЕРВНЫХ ЦЕНТРОВ
Принцип субординации нервных центров (принцип подчинения) проявляется в виде регулирующего
 Жизнь птиц в наше время
Жизнь птиц в наше время Разнообразие беспозвоночных животных. Викторина
Разнообразие беспозвоночных животных. Викторина Презентация на тему Третий закон Менделя
Презентация на тему Третий закон Менделя  Состав крови. Клетки крови. 8 класс
Состав крови. Клетки крови. 8 класс Растительный мир Песчанокопского района
Растительный мир Песчанокопского района Водный гиацинт. Влияние на него различных факторов загрязнения
Водный гиацинт. Влияние на него различных факторов загрязнения Вирусы
Вирусы Фотодыхание
Фотодыхание Головной мозг: строение и функции. Часть 1
Головной мозг: строение и функции. Часть 1 Сердце
Сердце Подборка учебной литературы по ветеринарии
Подборка учебной литературы по ветеринарии Презентация на тему Прибрежно-водные растения
Презентация на тему Прибрежно-водные растения  Загальна будова дихальної системи
Загальна будова дихальної системи Биология, как наука
Биология, как наука Буквы-витамины
Буквы-витамины Синтез систем лабораторного типа
Синтез систем лабораторного типа Закономерности и виды изменчивости
Закономерности и виды изменчивости Цветок толстянка. Наблюдение за прорастанием корня
Цветок толстянка. Наблюдение за прорастанием корня Царство бактерии
Царство бактерии Огород на подоконнике. Формирование представления детей об окружающем мире и прививать трудовые навыки
Огород на подоконнике. Формирование представления детей об окружающем мире и прививать трудовые навыки Внешнее строение млекопитающих
Внешнее строение млекопитающих Жеміс және Цитрус дақылдары зиянкестерінің энтомофагтары
Жеміс және Цитрус дақылдары зиянкестерінің энтомофагтары Опорно-двигательная система
Опорно-двигательная система Лес – наше богатство. Занятие №1
Лес – наше богатство. Занятие №1 Мейоз. Фазы мейоза
Мейоз. Фазы мейоза Презентация на тему Витамин А
Презентация на тему Витамин А  Класс птицы
Класс птицы Ядро. Занятие 4
Ядро. Занятие 4