- Отдел Lycopodiophyta

Содержание
- 2. Отдел Lycopodiophyta Возникли 400 млн. лет назад. Самые древние из нынеживущих растений спорофитной линии эволюции.
- 3. Ландшафт среднего Девона. Раннедевонские ( 408 - 387 млн лет назад) Cooksonia, Zosterophyllum, Aglaophyton. Среднедевонский Psilophyton
- 4. Одиночные пазушные спорангии эвспорангиатного типа, нет приспособлений для вскрывания
- 5. Появление листьев и корней
- 6. Возникновение разноспоровости
- 7. Класс Lycopodiopsida Нынеживщие и ископаемые равноспоровые плауны
- 8. Средний Девон 390-370 млн лет назад
- 10. Особенности строения нынеживущих равноспоровых плаунов
- 11. АМП с единственной АК АМП с множественными АИ У растений существует три структурных типа АМП. АИ
- 12. Lycopodium sp. Зачаток листа Поверхностные инициали Подповерхностные инициали Прокамбий
- 13. Huperzia selago
- 14. Строение апикальной меристемы Huperzia selago Прокамбий Группа поверхностных апикальных инициалей Периферическая зона Подповерхностные инициали Зачаток листа
- 15. Заложение листа в апикальной меристеме Huperzia selago Зачаток листа
- 16. Внутренне строение побегов и корней
- 18. Плектостела на поперечном срезе корневища Lycopodium,
- 20. Плектостела
- 21. Актиностела на поперечном срезе корня Lycopodium sp.
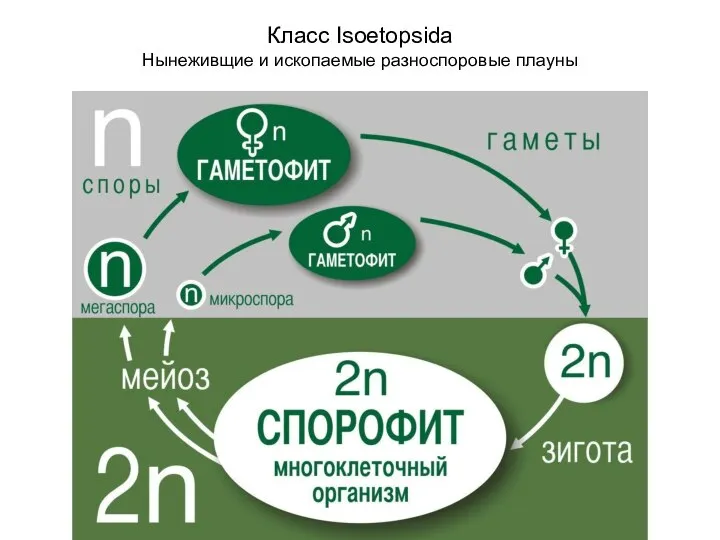
- 22. Класс Isoetopsida Нынеживщие и ископаемые разноспоровые плауны
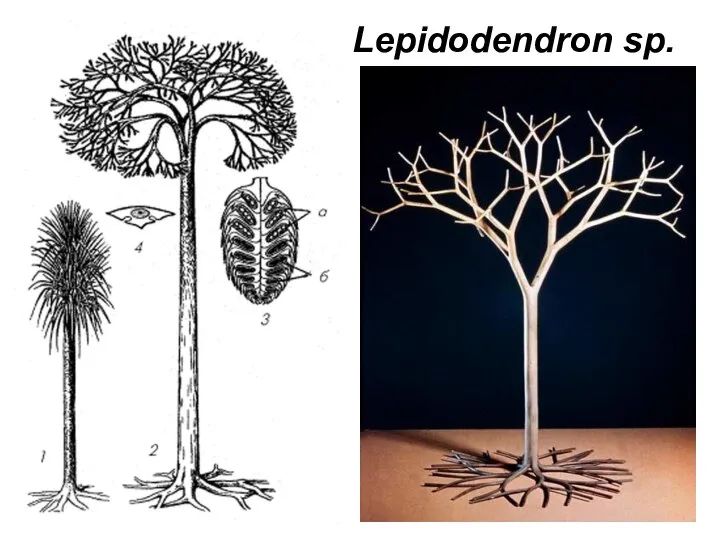
- 24. Lepidodendron sp.
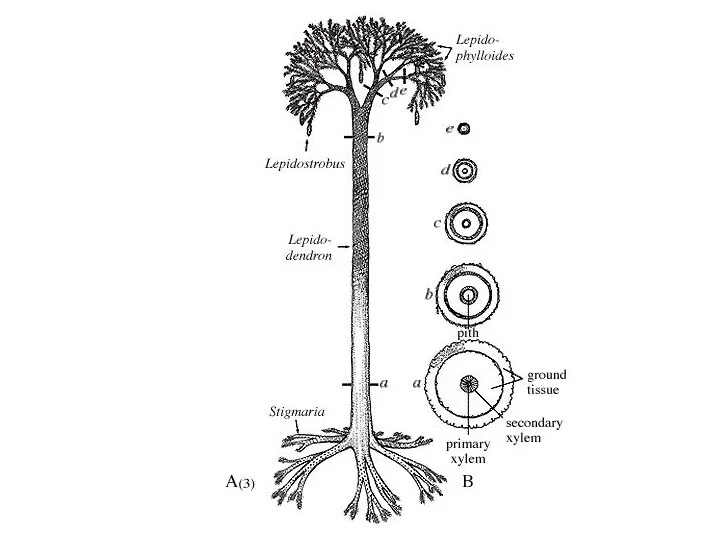
- 25. Lepidodendron реконструкция олиственной ветви с листьями до 70 см длиной и стробилами микроспорангиев
- 26. “Окаменевшая роща” Victoria Park, Glasgow, Scotland
- 27. Окаменевшие ризофоры (род Stigmaria)
- 29. Stigmaria – слепок, на котором видны места прикрепления корней
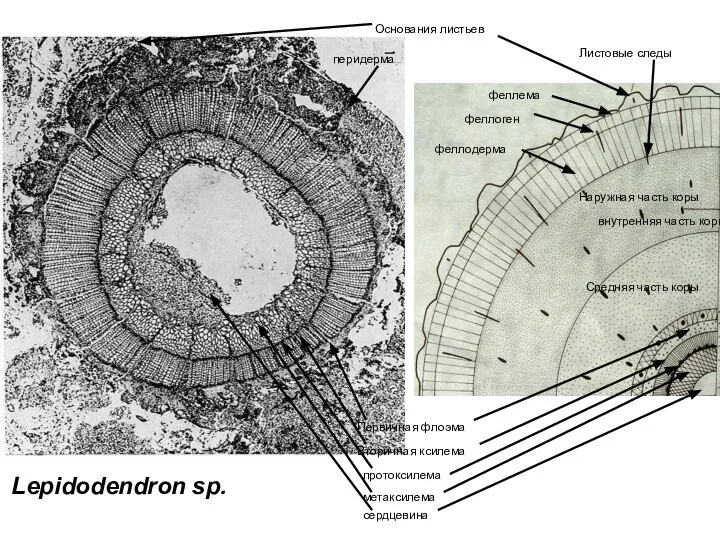
- 31. перидерма феллема феллоген феллодерма Наружная часть коры Листовые следы Средняя часть коры внутренняя часть коры Первичная
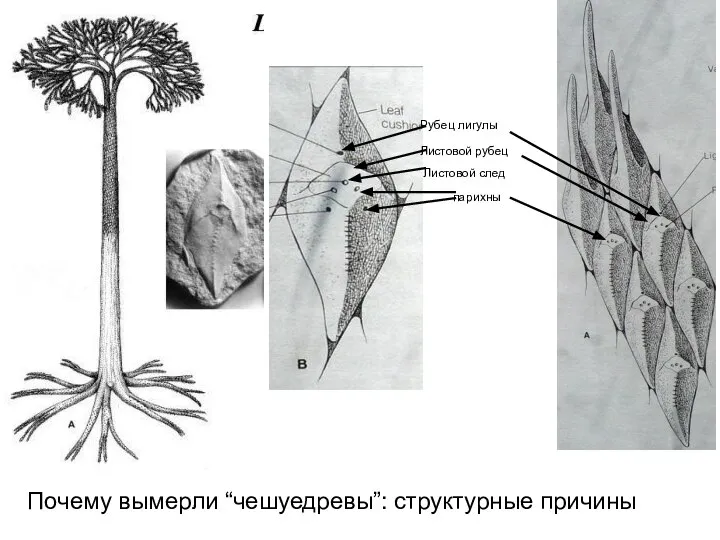
- 32. Листовой рубец Рубец лигулы Листовой след парихны Почему вымерли “чешуедревы”: структурные причины
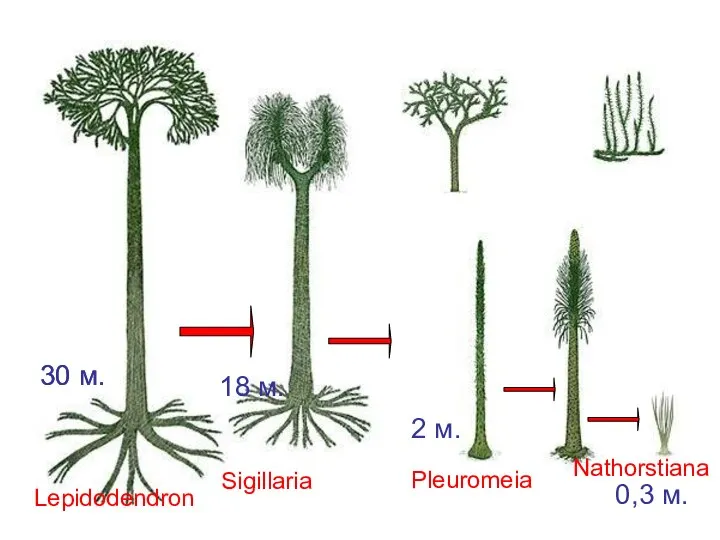
- 33. Становление низкорослых форм у Lepidodendrales Lepidodendron Sigillaria Pleuromeia Nathorstiana 30 м. 30 м. 18 м. 2
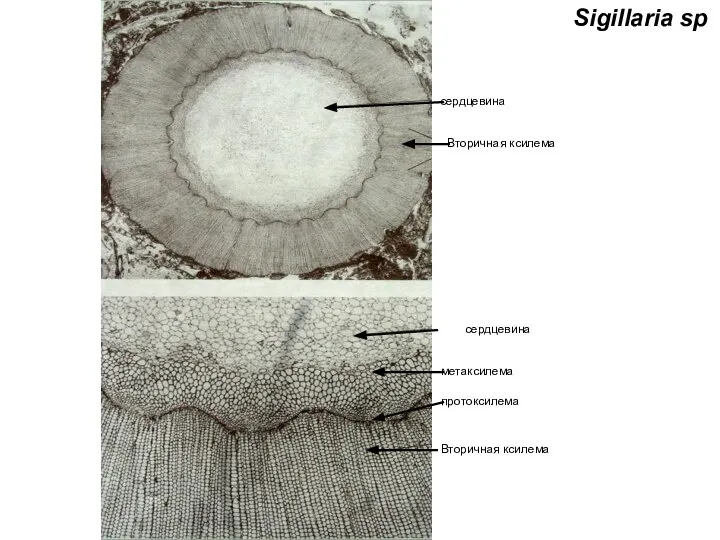
- 35. Sigillaria sp Вторичная ксилема метаксилема сердцевина протоксилема Вторичная ксилема сердцевина
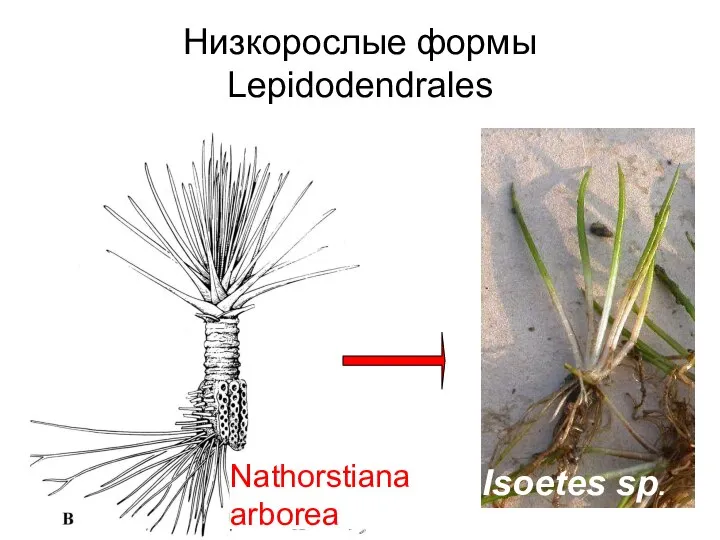
- 36. Низкорослые формы Lepidodendrales Nathorstiana arborea Isoetes sp.
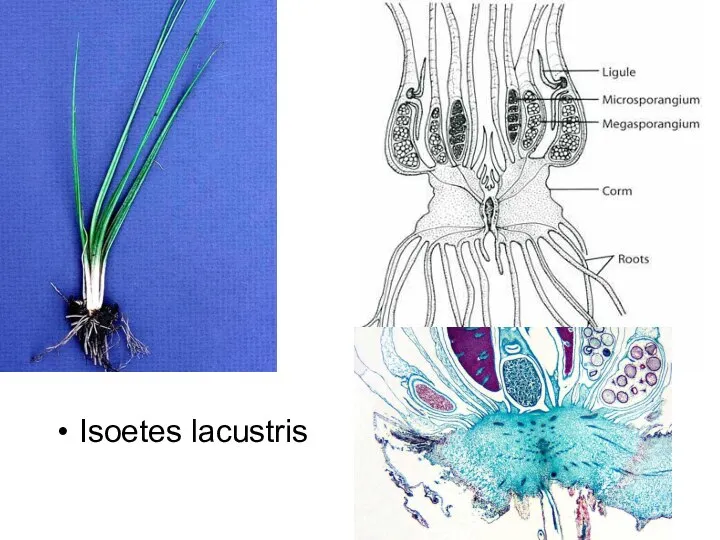
- 37. Isoetes lacustris
- 38. Призматический слой Базальная меристема Зачатки корней Листовой след Вторичная кора
- 39. Основания листьев Листовой след Апекс побега Призматический слой Базальная меристема Зачатки корней Масса ксилемы Листовой след
- 40. Монархный корень Isoetes с воздухоносной полостью
- 41. Разноспоровые плауны Род Selaginella – 700 видов
- 42. Selaginella kraussiana Микрофилльные листья , дихотомически ветвящиеся побеги, органы неясной морфологической природы ризофоры
- 43. Органы двойственной морфологической природы ризофоры закономерно возникают из угловой меристемы (экзогенно, предположительно из “старой” апикальной клетки)
- 44. Анизодихоподиальное нарастание. Переход в генеративную фазу коррелирует с терминацией морфогенеза.
- 45. Как побеги, ризофоры образуются экзогенно, не имеют корневого чехлика, при определенных условиях они могут развиться в
- 46. АК Апикальная меристема с единственной инициалью; зональность формируется за счет неравных периклинальных делений в мерофитах ПИ
- 47. Заложение листа в апикальной меристеме Selaginella kraussiana
- 48. Дихотомическое ветвление со сменой апикальных инициалей.
- 49. Проводящая система - стела из двух меристел с экзархной ксилемой (разновидность актиностелы), окруженная воздухоносной полостью. Корень
- 50. Нет общепринятого мнения относительно происхождения и эволюционных гомологий листьев у высших растений Листья всех растений имеют
- 51. Энации Nothia (Zosterophyllophyta)
- 52. Protolepidodendron (Lycopodiophyta) Редукция теломов
- 53. Drepanophycus (Lycopodiophyta) Asteroxylon (Lycopodiophyta) Стерилизация спорангиев
- 54. Huperzia selago ЦЗ ПЗ ПЗ Один из критериев энационного листа - возникновение ниже апикальной меристемы.
- 55. Отд. Lycopodiophyta Появление листьев. 3 версии происхождения: (1) Энации (микрофилльная линия), (2) редукция теломов (листья всех
- 56. Однополярный рост спорофита. Из апикальной меристемы образуются только листья и спорангии. Ветвление верхушечное (дихотомическое) со сменой
- 57. Эвспорангии одиночные, расположены в пазухах листьев. Появление разноспоровости. Все эволюционные преобразования (появление листьев, корней, разноспоровости) у
- 59. Скачать презентацию
Слайд 2Отдел
Lycopodiophyta
Возникли 400 млн. лет назад.
Самые древние из нынеживущих растений спорофитной линии
Отдел
Lycopodiophyta
Возникли 400 млн. лет назад.
Самые древние из нынеживущих растений спорофитной линии

Слайд 3Ландшафт среднего Девона. Раннедевонские ( 408 - 387 млн лет назад) Cooksonia,
Ландшафт среднего Девона. Раннедевонские ( 408 - 387 млн лет назад) Cooksonia,

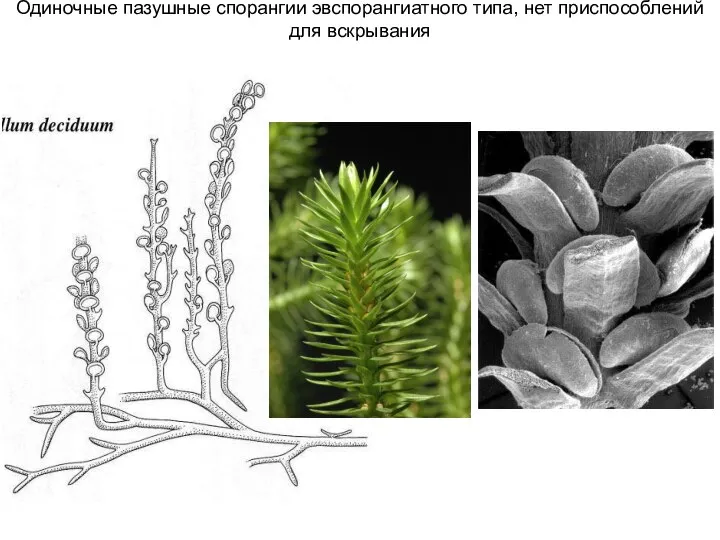
Слайд 4Одиночные пазушные спорангии эвспорангиатного типа, нет приспособлений для вскрывания
Одиночные пазушные спорангии эвспорангиатного типа, нет приспособлений для вскрывания
Слайд 5Появление листьев и корней
Появление листьев и корней

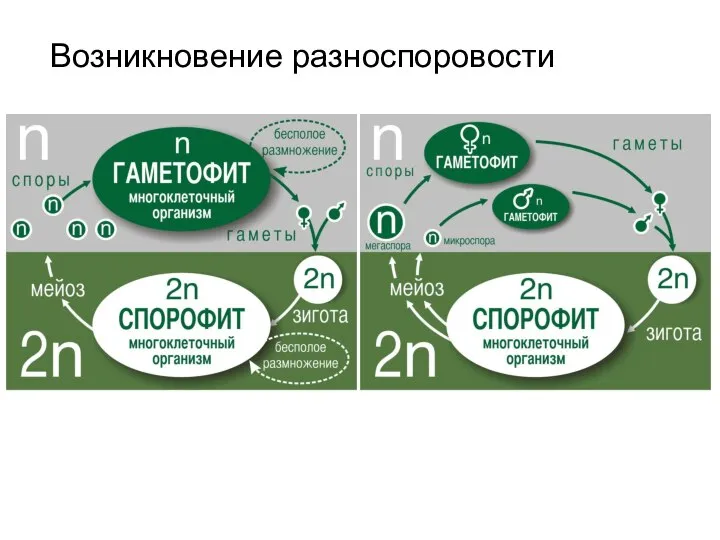
Слайд 6Возникновение разноспоровости
Возникновение разноспоровости
Слайд 7Класс Lycopodiopsida
Нынеживщие и ископаемые равноспоровые плауны
Класс Lycopodiopsida
Нынеживщие и ископаемые равноспоровые плауны

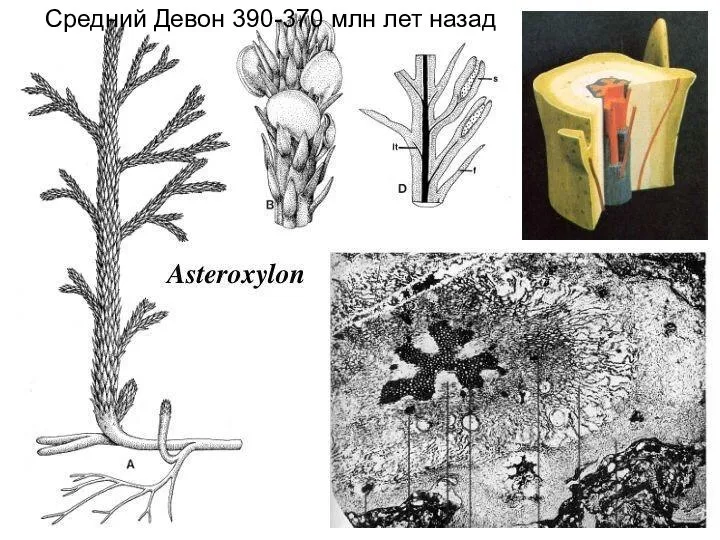
Слайд 8Средний Девон 390-370 млн лет назад
Средний Девон 390-370 млн лет назад

Слайд 10Особенности строения нынеживущих равноспоровых плаунов
Особенности строения нынеживущих равноспоровых плаунов

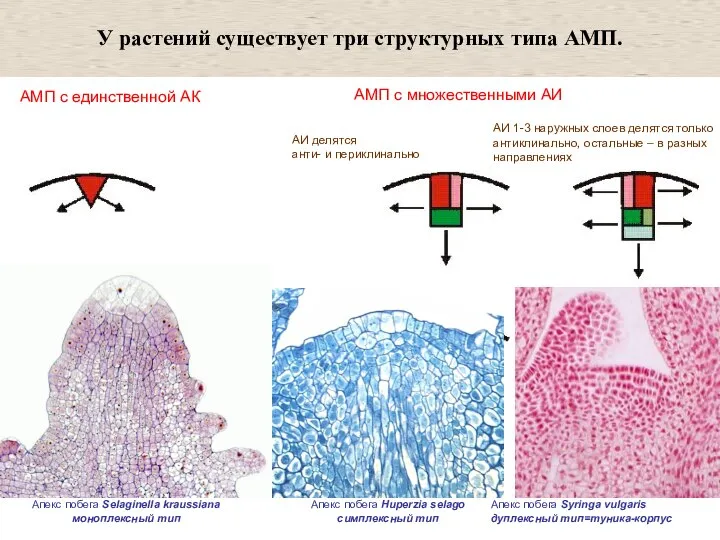
Слайд 11АМП с единственной АК
АМП с множественными АИ
У растений существует три структурных типа
АМП с единственной АК
АМП с множественными АИ
У растений существует три структурных типа
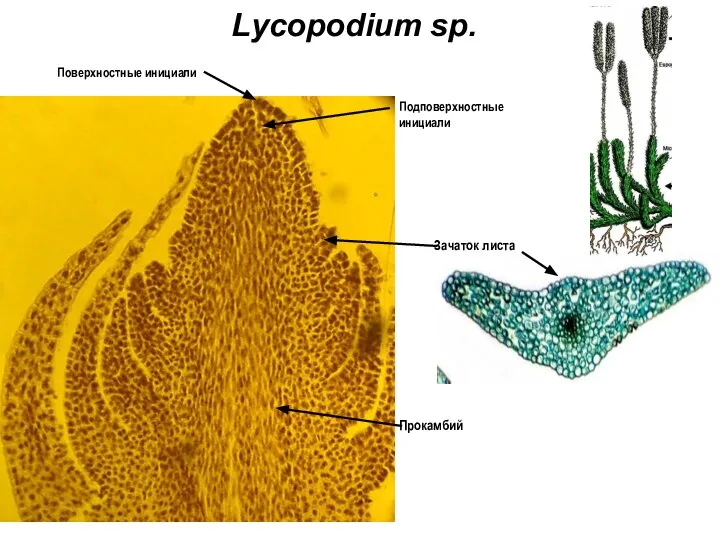
Слайд 12Lycopodium sp.
Зачаток листа
Поверхностные инициали
Подповерхностные
инициали
Прокамбий
Lycopodium sp.
Зачаток листа
Поверхностные инициали
Подповерхностные
инициали
Прокамбий
Слайд 13Huperzia selago
Huperzia selago

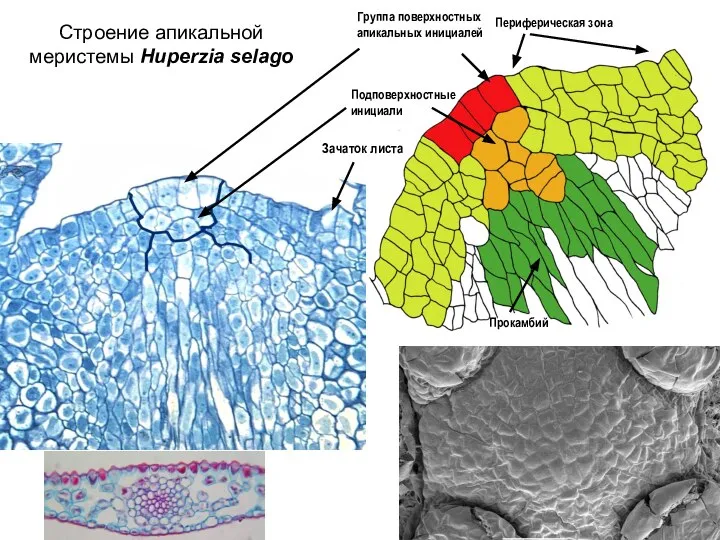
Слайд 14Строение апикальной меристемы Huperzia selago
Прокамбий
Группа поверхностных
апикальных инициалей
Периферическая зона
Подповерхностные
инициали
Зачаток листа
Строение апикальной меристемы Huperzia selago
Прокамбий
Группа поверхностных
апикальных инициалей
Периферическая зона
Подповерхностные
инициали
Зачаток листа
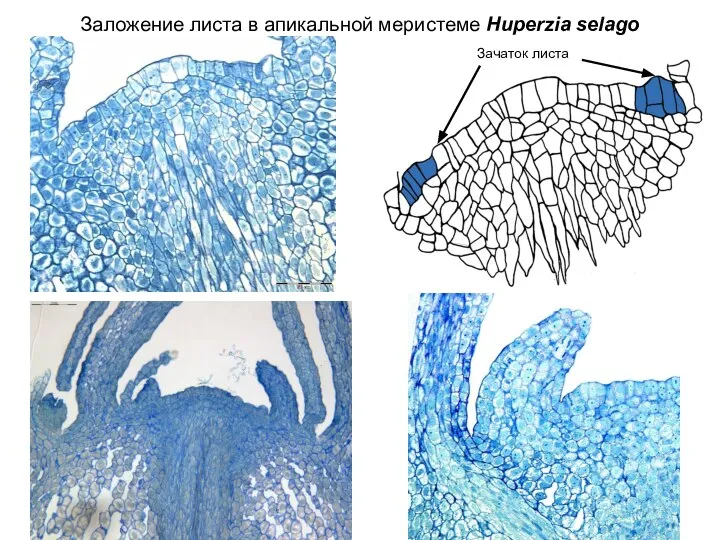
Слайд 15Заложение листа в апикальной меристеме Huperzia selago
Зачаток листа
Заложение листа в апикальной меристеме Huperzia selago
Зачаток листа
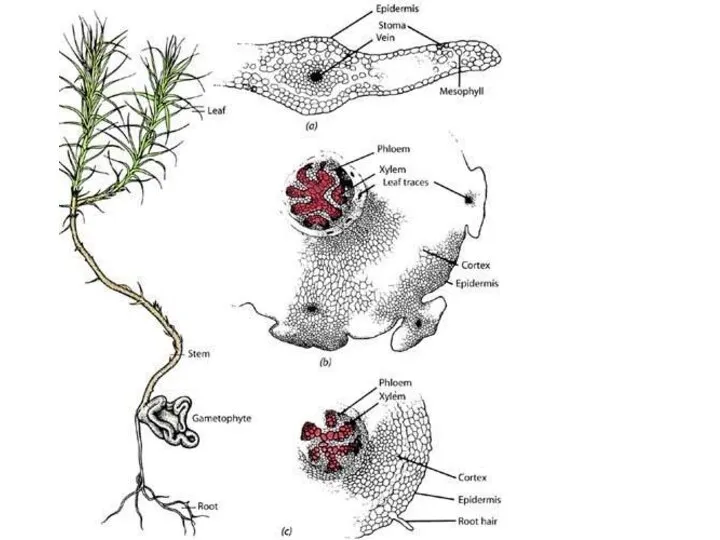
Слайд 16Внутренне строение побегов и корней
Внутренне строение побегов и корней

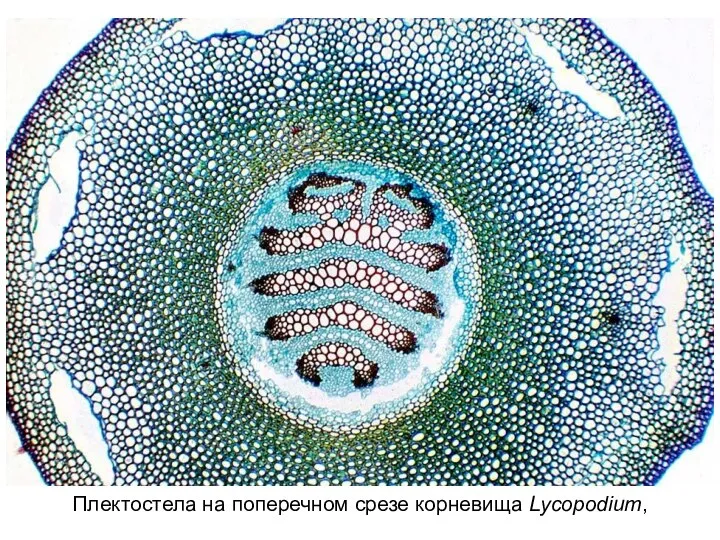
Слайд 18Плектостела на поперечном срезе корневища Lycopodium,
Плектостела на поперечном срезе корневища Lycopodium,
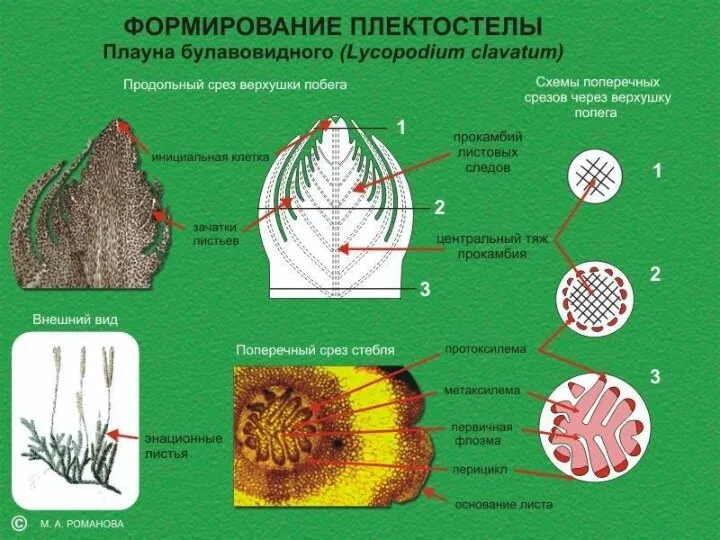
Слайд 20Плектостела
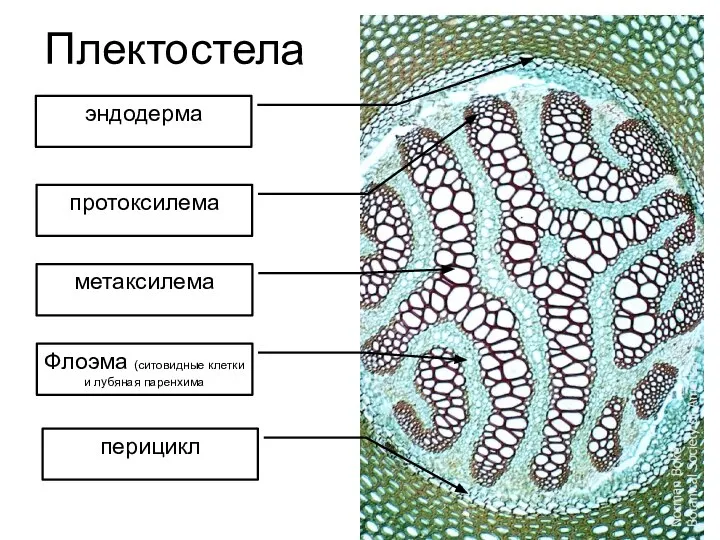
Плектостела
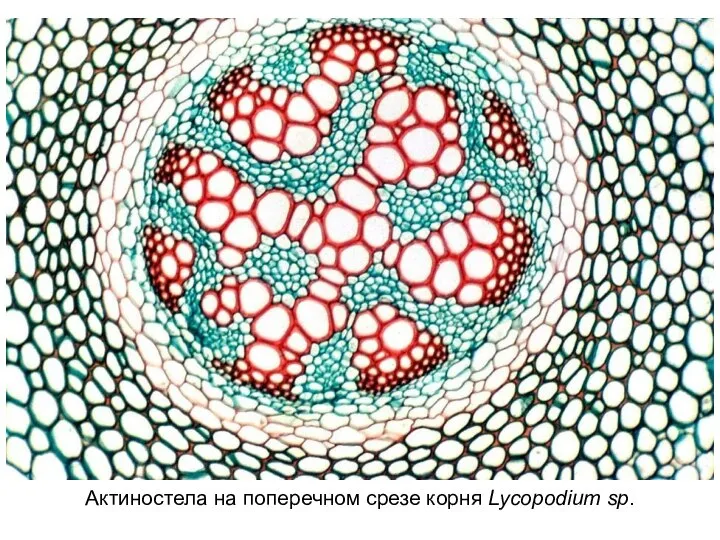
Слайд 21Актиностела на поперечном срезе корня Lycopodium sp.
Актиностела на поперечном срезе корня Lycopodium sp.
Слайд 22Класс Isoetopsida
Нынеживщие и ископаемые разноспоровые плауны
Класс Isoetopsida
Нынеживщие и ископаемые разноспоровые плауны

Слайд 24Lepidodendron sp.
Lepidodendron sp.
Слайд 25Lepidodendron реконструкция олиственной ветви с листьями до 70 см длиной и стробилами
Lepidodendron реконструкция олиственной ветви с листьями до 70 см длиной и стробилами

Слайд 26“Окаменевшая роща” Victoria Park, Glasgow, Scotland
“Окаменевшая роща” Victoria Park, Glasgow, Scotland

Слайд 27Окаменевшие ризофоры (род Stigmaria)
Окаменевшие ризофоры (род Stigmaria)


Слайд 29Stigmaria – слепок, на котором видны места прикрепления корней
Stigmaria – слепок, на котором видны места прикрепления корней

Слайд 31перидерма
феллема
феллоген
феллодерма
Наружная часть коры
Листовые следы
Средняя часть коры
внутренняя часть коры
Первичная флоэма
Вторичная ксилема
Основания листьев
метаксилема
сердцевина
протоксилема
Lepidodendron
перидерма
феллема
феллоген
феллодерма
Наружная часть коры
Листовые следы
Средняя часть коры
внутренняя часть коры
Первичная флоэма
Вторичная ксилема
Основания листьев
метаксилема
сердцевина
протоксилема
Lepidodendron
Слайд 32Листовой рубец
Рубец лигулы
Листовой след
парихны
Почему вымерли “чешуедревы”: структурные причины
Листовой рубец
Рубец лигулы
Листовой след
парихны
Почему вымерли “чешуедревы”: структурные причины
Слайд 33Становление низкорослых форм у Lepidodendrales
Lepidodendron
Sigillaria
Pleuromeia
Nathorstiana
30 м.
30 м.
18 м.
2 м.
0,3 м.
Становление низкорослых форм у Lepidodendrales
Lepidodendron
Sigillaria
Pleuromeia
Nathorstiana
30 м.
30 м.
18 м.
2 м.
0,3 м.

Слайд 35Sigillaria sp
Вторичная ксилема
метаксилема
сердцевина
протоксилема
Вторичная ксилема
сердцевина
Sigillaria sp
Вторичная ксилема
метаксилема
сердцевина
протоксилема
Вторичная ксилема
сердцевина
Слайд 36Низкорослые формы Lepidodendrales
Nathorstiana arborea
Isoetes sp.
Низкорослые формы Lepidodendrales
Nathorstiana arborea
Isoetes sp.
Слайд 37Isoetes lacustris
Isoetes lacustris
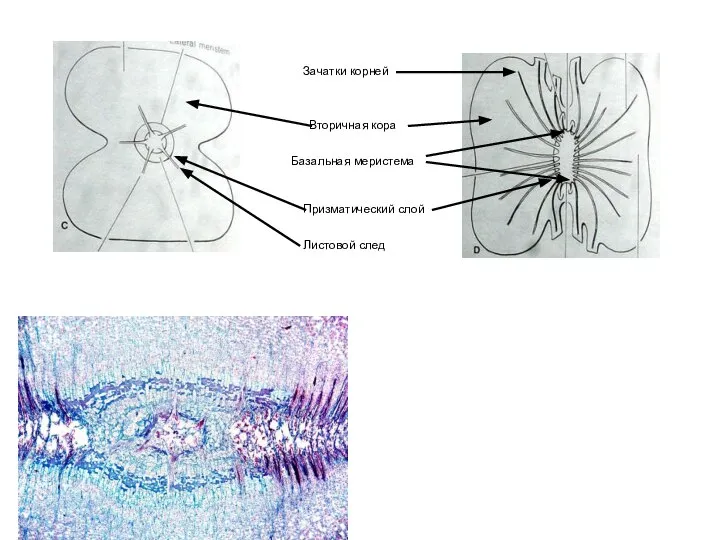
Слайд 38Призматический слой
Базальная меристема
Зачатки корней
Листовой след
Вторичная кора
Призматический слой
Базальная меристема
Зачатки корней
Листовой след
Вторичная кора
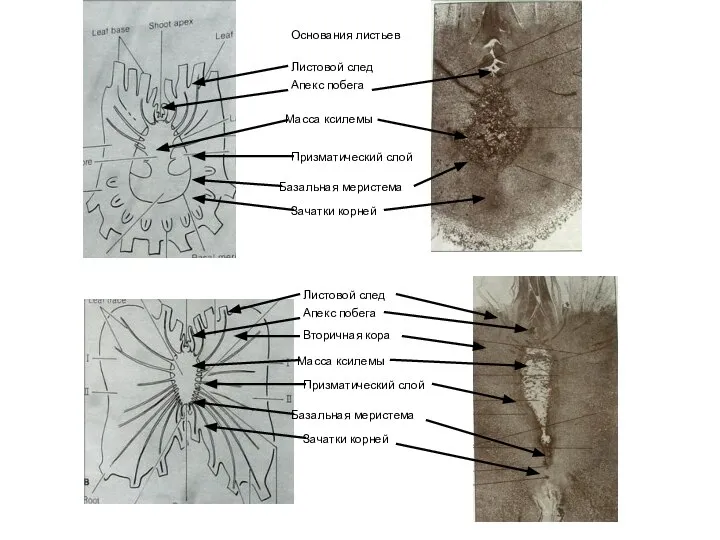
Слайд 39Основания листьев
Листовой след
Апекс побега
Призматический слой
Базальная меристема
Зачатки корней
Масса ксилемы
Листовой след
Апекс побега
Призматический слой
Базальная меристема
Зачатки
Основания листьев
Листовой след
Апекс побега
Призматический слой
Базальная меристема
Зачатки корней
Масса ксилемы
Листовой след
Апекс побега
Призматический слой
Базальная меристема
Зачатки
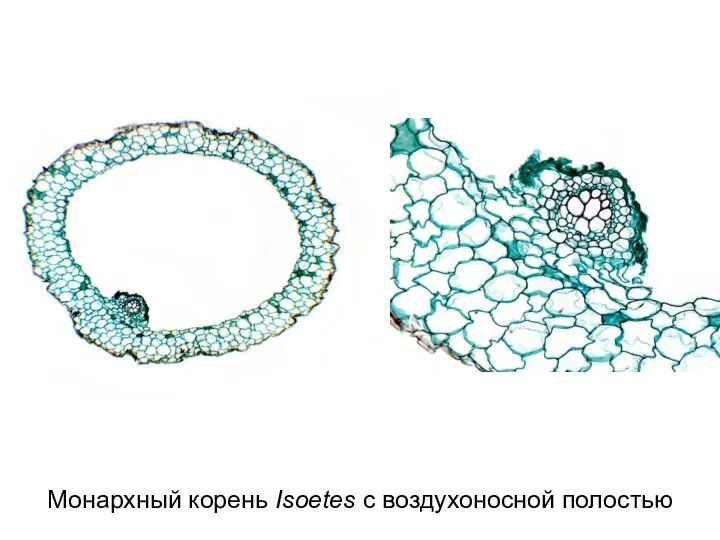
Слайд 40Монархный корень Isoetes с воздухоносной полостью
Монархный корень Isoetes с воздухоносной полостью
Слайд 41Разноспоровые плауны
Род Selaginella – 700 видов
Разноспоровые плауны
Род Selaginella – 700 видов

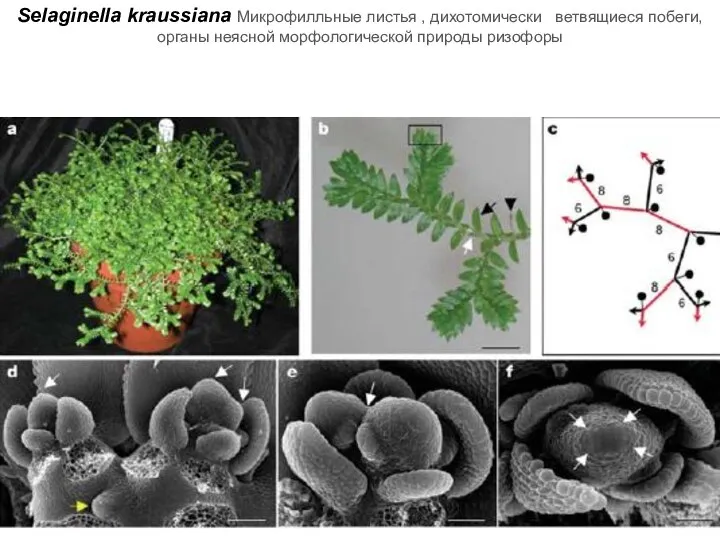
Слайд 42Selaginella kraussiana Микрофилльные листья , дихотомически ветвящиеся побеги, органы неясной морфологической природы
Selaginella kraussiana Микрофилльные листья , дихотомически ветвящиеся побеги, органы неясной морфологической природы
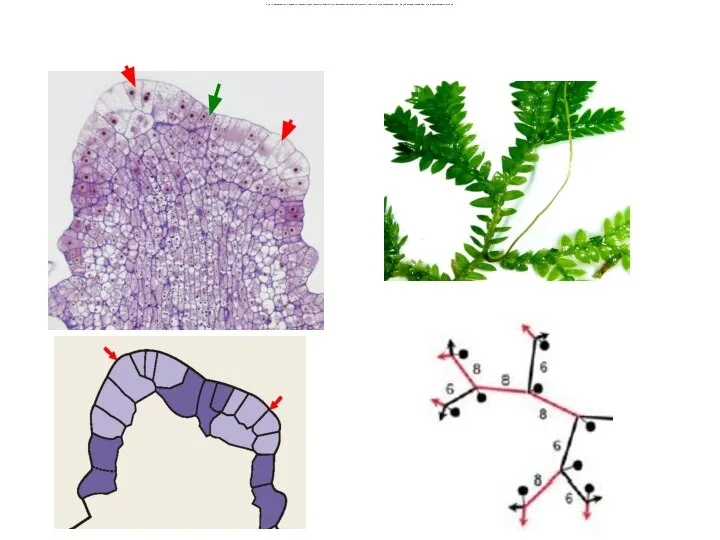
Слайд 43Органы двойственной морфологической природы ризофоры закономерно возникают из угловой меристемы (экзогенно, предположительно
Органы двойственной морфологической природы ризофоры закономерно возникают из угловой меристемы (экзогенно, предположительно
Слайд 44Анизодихоподиальное нарастание. Переход в генеративную фазу коррелирует с терминацией морфогенеза.
Анизодихоподиальное нарастание. Переход в генеративную фазу коррелирует с терминацией морфогенеза.

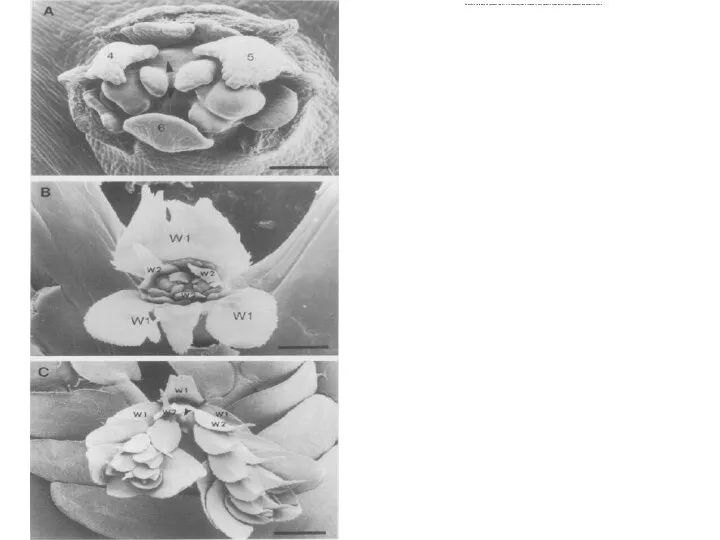
Слайд 45Как побеги, ризофоры образуются экзогенно, не имеют корневого чехлика, при определенных условиях
Как побеги, ризофоры образуются экзогенно, не имеют корневого чехлика, при определенных условиях
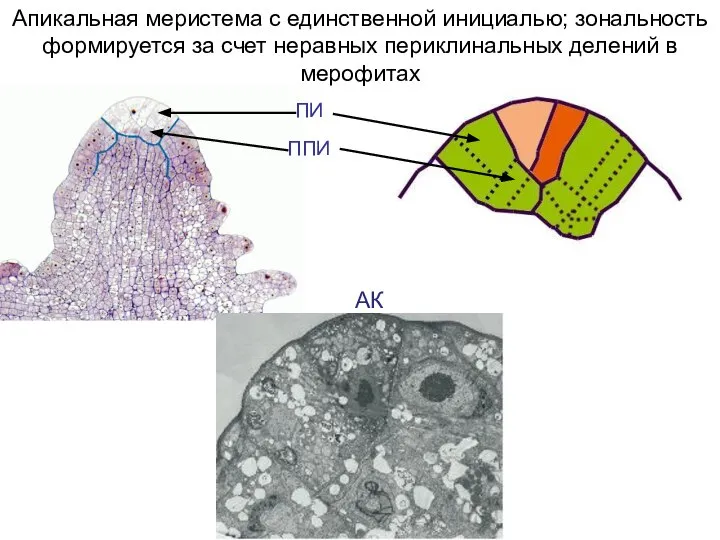
Слайд 46АК
Апикальная меристема с единственной инициалью; зональность формируется за счет неравных периклинальных делений
АК
Апикальная меристема с единственной инициалью; зональность формируется за счет неравных периклинальных делений
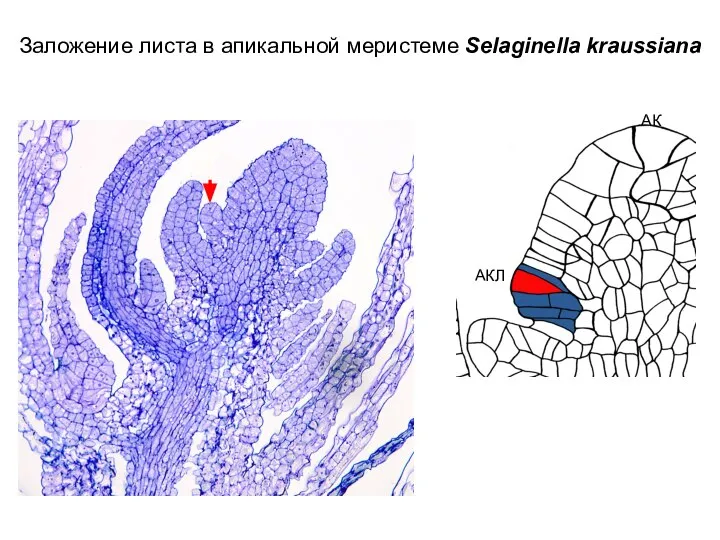
Слайд 47Заложение листа в апикальной меристеме Selaginella kraussiana
Заложение листа в апикальной меристеме Selaginella kraussiana
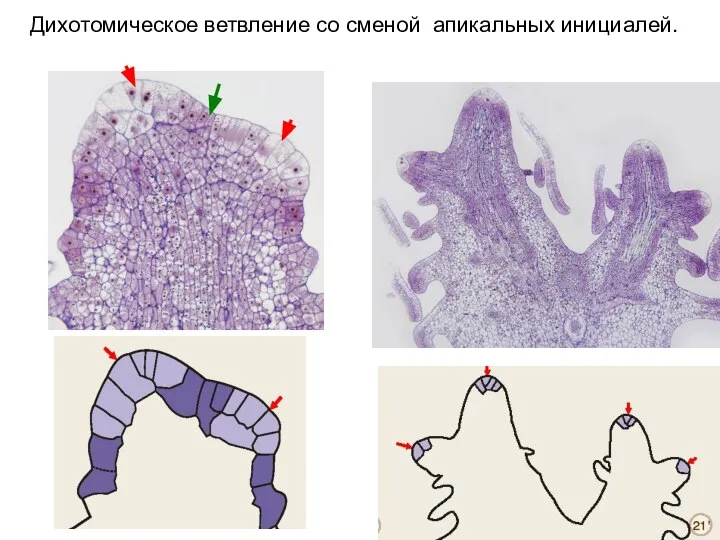
Слайд 48Дихотомическое ветвление со сменой апикальных инициалей.
Дихотомическое ветвление со сменой апикальных инициалей.
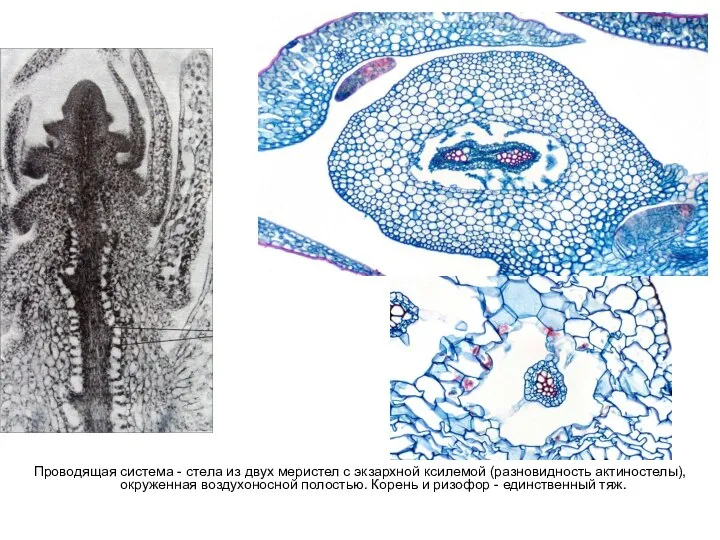
Слайд 49Проводящая система - стела из двух меристел с экзархной ксилемой (разновидность актиностелы),
Проводящая система - стела из двух меристел с экзархной ксилемой (разновидность актиностелы),
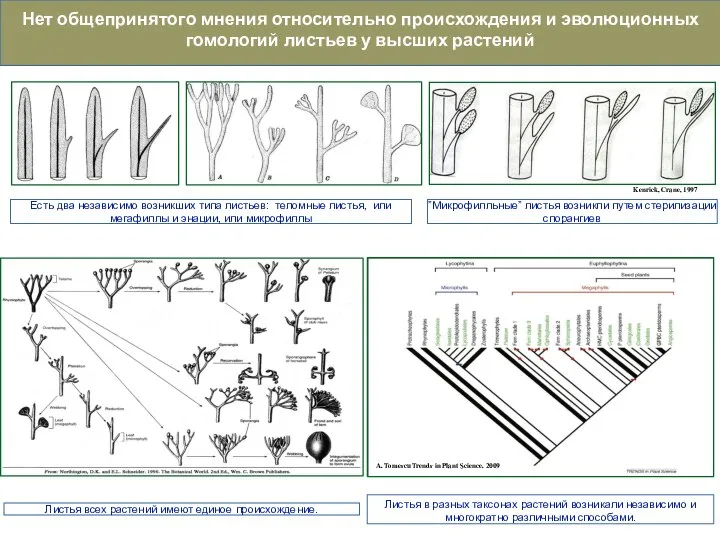
Слайд 50Нет общепринятого мнения относительно происхождения и эволюционных гомологий листьев у высших растений
Листья
Нет общепринятого мнения относительно происхождения и эволюционных гомологий листьев у высших растений
Листья
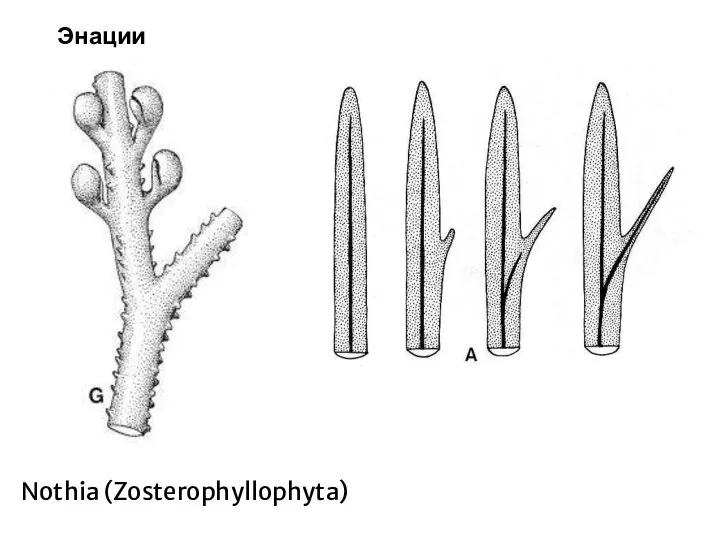
Слайд 51Энации
Nothia (Zosterophyllophyta)
Энации
Nothia (Zosterophyllophyta)
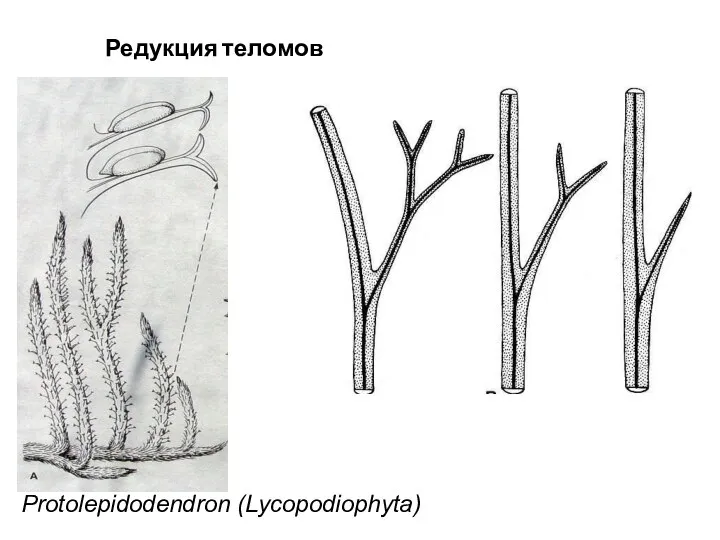
Слайд 52Protolepidodendron (Lycopodiophyta)
Редукция теломов
Protolepidodendron (Lycopodiophyta)
Редукция теломов
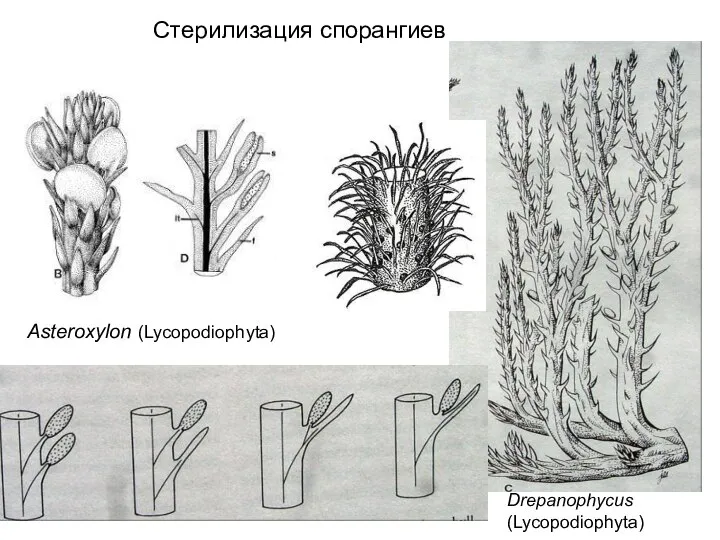
Слайд 53Drepanophycus (Lycopodiophyta)
Asteroxylon (Lycopodiophyta)
Стерилизация спорангиев
Drepanophycus (Lycopodiophyta)
Asteroxylon (Lycopodiophyta)
Стерилизация спорангиев
Слайд 54Huperzia selago
ЦЗ
ПЗ
ПЗ
Один из критериев энационного листа - возникновение ниже апикальной меристемы.
Huperzia selago
ЦЗ
ПЗ
ПЗ
Один из критериев энационного листа - возникновение ниже апикальной меристемы.

Слайд 55Отд. Lycopodiophyta
Появление листьев. 3 версии происхождения: (1) Энации (микрофилльная линия), (2) редукция
Отд. Lycopodiophyta
Появление листьев. 3 версии происхождения: (1) Энации (микрофилльная линия), (2) редукция

Слайд 56Однополярный рост спорофита. Из апикальной меристемы образуются только листья и спорангии.
Ветвление
Однополярный рост спорофита. Из апикальной меристемы образуются только листья и спорангии.
Ветвление

Слайд 57Эвспорангии одиночные, расположены в пазухах листьев.
Появление разноспоровости.
Все эволюционные преобразования (появление листьев,
Эвспорангии одиночные, расположены в пазухах листьев.
Появление разноспоровости.
Все эволюционные преобразования (появление листьев,

 Мята перечная
Мята перечная Строение и функции головного мозга
Строение и функции головного мозга 4_Биология_7 класс_Органические вещества в продуктах питания_Презентация
4_Биология_7 класс_Органические вещества в продуктах питания_Презентация Репродуктивные органы растения Цветок Автор: Печенкина В.А. Учитель МОУ «Гимназия№10» Г.Пушкино Московской области
Репродуктивные органы растения Цветок Автор: Печенкина В.А. Учитель МОУ «Гимназия№10» Г.Пушкино Московской области Гистологические особенности женской репродуктивной системы
Гистологические особенности женской репродуктивной системы Презентация на тему Палеозой. Ородовик
Презентация на тему Палеозой. Ородовик  Презентация на тему Семейство Мальвовые
Презентация на тему Семейство Мальвовые  Взаимосвязь между структурой, свойствами и функциями клеточной мембраны
Взаимосвязь между структурой, свойствами и функциями клеточной мембраны Патология синапсов
Патология синапсов Значение дыхания. Органы дыхания
Значение дыхания. Органы дыхания Популяция - структурная единица вида
Популяция - структурная единица вида Экология лося
Экология лося Пшеница и рожь. Сравнение зерновых культур
Пшеница и рожь. Сравнение зерновых культур Цветок - орган полового размножения
Цветок - орган полового размножения Выделение. Мочевыделительная система
Выделение. Мочевыделительная система Лес - богатство Республики Коми
Лес - богатство Республики Коми Презентация на тему РЕЧЬ И МЫШЛЕНИЕ
Презентация на тему РЕЧЬ И МЫШЛЕНИЕ  Развитие биохимии и биофизики
Развитие биохимии и биофизики Корень. Биологический диктант
Корень. Биологический диктант Витамины в организме человека
Витамины в организме человека Марианская впадина
Марианская впадина Leishmania
Leishmania Hospodářská zvířata a drůbež
Hospodářská zvířata a drůbež Презентация на тему Патогенные бактерии
Презентация на тему Патогенные бактерии  Учение В.И. Вернадского о биосфере. Эволюция биосферы
Учение В.И. Вернадского о биосфере. Эволюция биосферы Презентация на тему Геном человека
Презентация на тему Геном человека  Разные насекомые
Разные насекомые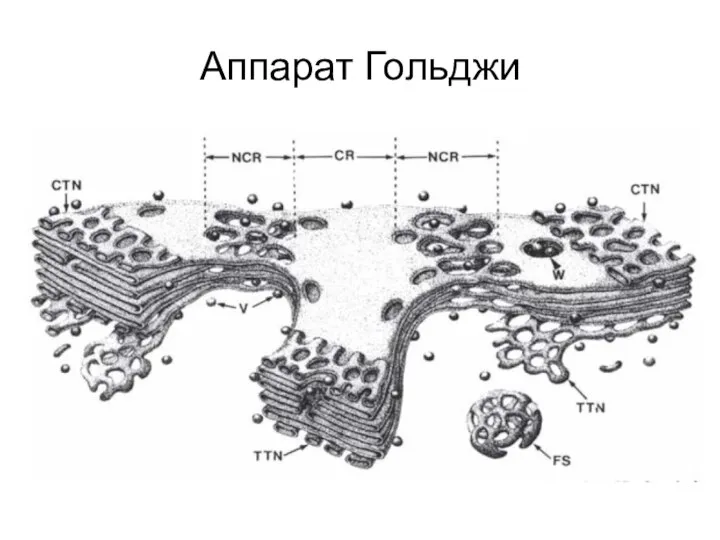 Аппарат Гольджи
Аппарат Гольджи