- Передача гормонального сигнала через мембрану

Содержание
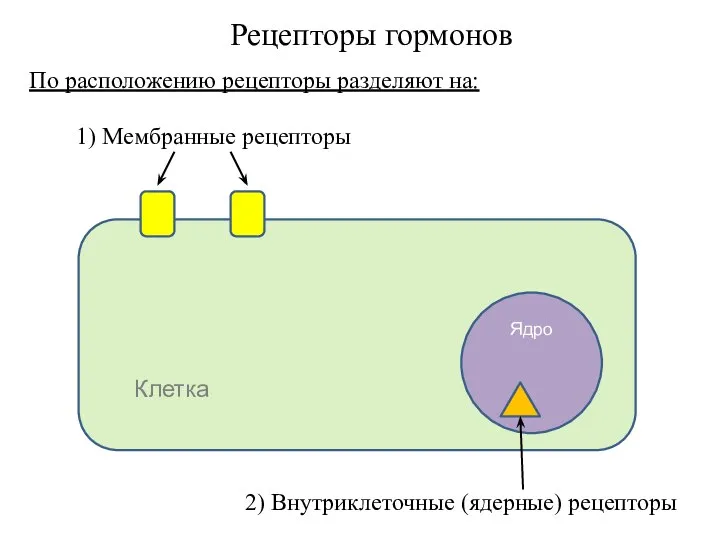
- 2. Рецепторы гормонов 1) Мембранные рецепторы Клетка Ядро 2) Внутриклеточные (ядерные) рецепторы По расположению рецепторы разделяют на:
- 3. I. Мембранные рецепторы гормонов Мембранные рецепторы составляют большинство рецепторов. Молекула лиганда в данном случае не проникает
- 4. I. Мембранные рецепторы гормонов Эту группу рецепторов можно разделить на: 1) Рецепторы, не обладающие ферментативной активностью:
- 5. 1. Рецепторы без ферментативной активности Рецепторы без ферментативной активности – подтип трансмембранных рецепторов, которым для воздействия
- 6. 1.1.Рецепторы, ассоциированные с G-белками Рецепторы, ассоциированные с G-белками – один из наиболее изученных классов мембранных рецепторов.
- 8. 1.1.Рецепторы, ассоциированные с G-белками С такими рецепторами связан ассоциированный с мембраной адапторный белок с гуанозинтрифосфатазной (ГТФазной)
- 9. 1.1.Рецепторы, ассоциированные с G-белками Ферментативной (ГТФазной) активностью обладает только α-субъединица G-белка. В неактивной форме она связана
- 10. Схема действия G-белка
- 11. 1.1.Рецепторы, ассоциированные с G-белками Наиболее хорошо изученные пути передачи сигнала с помощью ассоциированных с G-белком рецепторов:
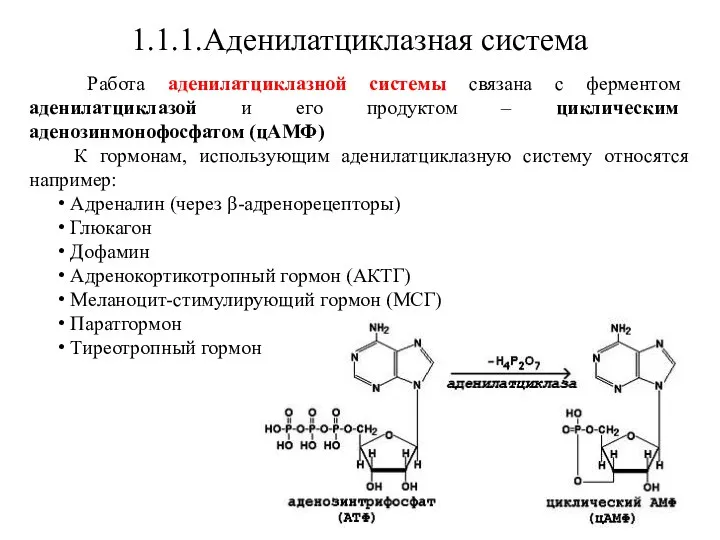
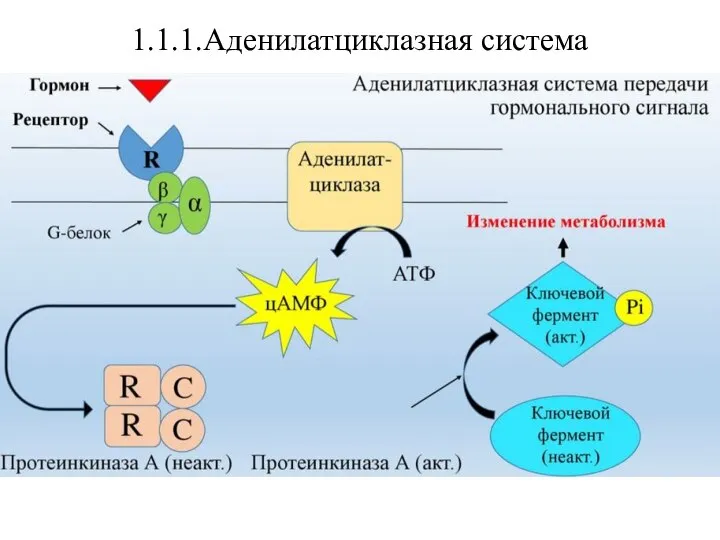
- 12. 1.1.1.Аденилатциклазная система Работа аденилатциклазной системы связана с ферментом аденилатциклазой и его продуктом – циклическим аденозинмонофосфатом (цАМФ)
- 13. 1.1.1.Аденилатциклазная система
- 14. 1.1.1.Аденилатциклазная система
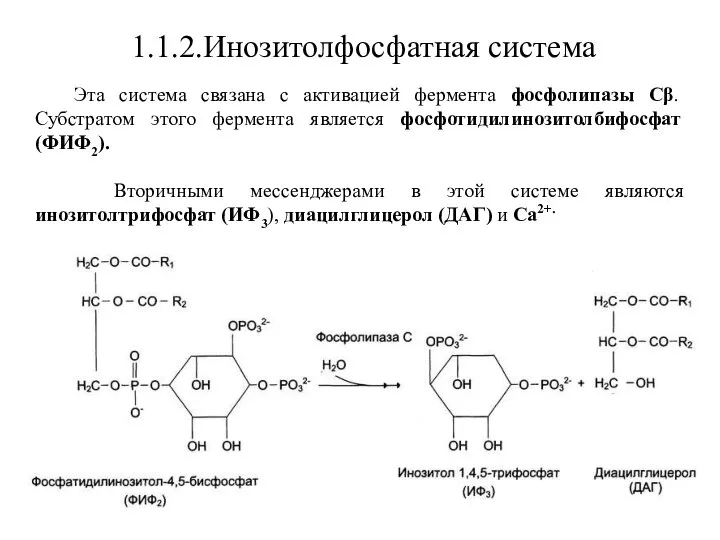
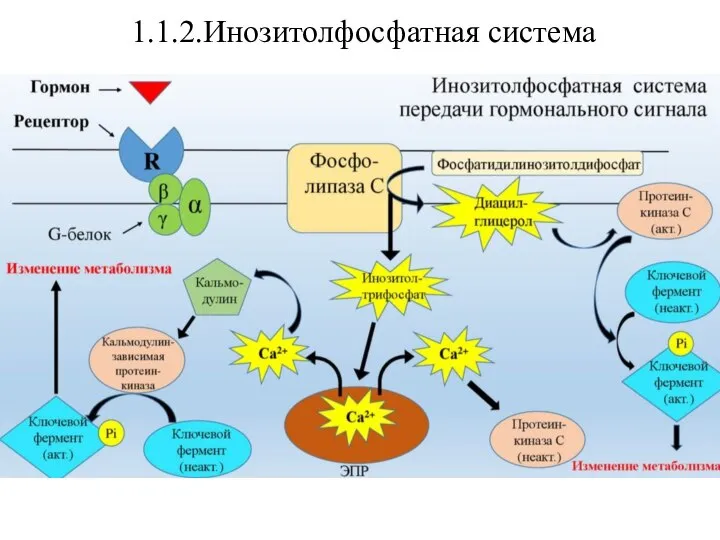
- 15. 1.1.2.Инозитолфосфатная система Эта система связана с активацией фермента фосфолипазы Сβ. Субстратом этого фермента является фосфотидилинозитолбифосфат(ФИФ2). Вторичными
- 16. 1.1.2.Инозитолфосфатная система Гормоны, использующие инозитолфосфатную систему: Адреналин (через α-адренорецепторы) Окситоцин Вазопрессин (через V1-рецепторы) Ацетилхолин (через М1-рецепторы)
- 17. 1.1.2.Инозитолфосфатная система
- 18. 1.1.2.Инозитолфосфатная система
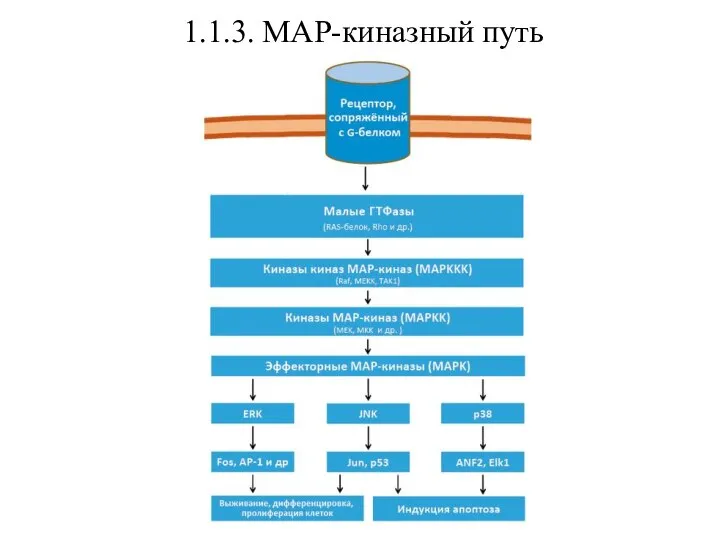
- 19. 1.1.3. МАР-киназный путь Помимо указанных выше, для рецепторов, сопряженных с G-белками, существует важный дополнительный сигнальный путь
- 20. 1.1.3. МАР-киназный путь
- 21. 1.2. Рецепторы, ассоциированные с JAK-киназами Принципиально отличным классом рецепторных молекул являются рецепторы, сопряженные с тирозинкиназами класса
- 22. 1.2. Рецепторы, ассоциированные с JAK-киназами К рецепторам, сопряжённым с JAK-киназами относятся три семейства: Семейство рецепторов группы
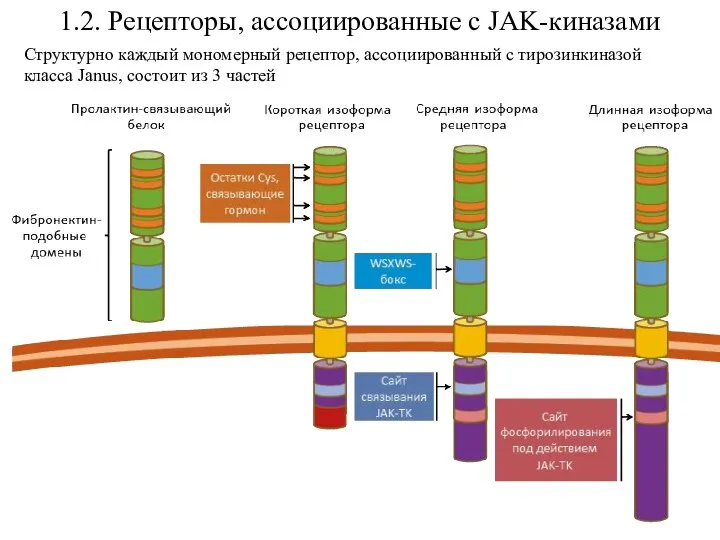
- 23. 1.2. Рецепторы, ассоциированные с JAK-киназами Структурно каждый мономерный рецептор, ассоциированный с тирозинкиназой класса Janus, состоит из
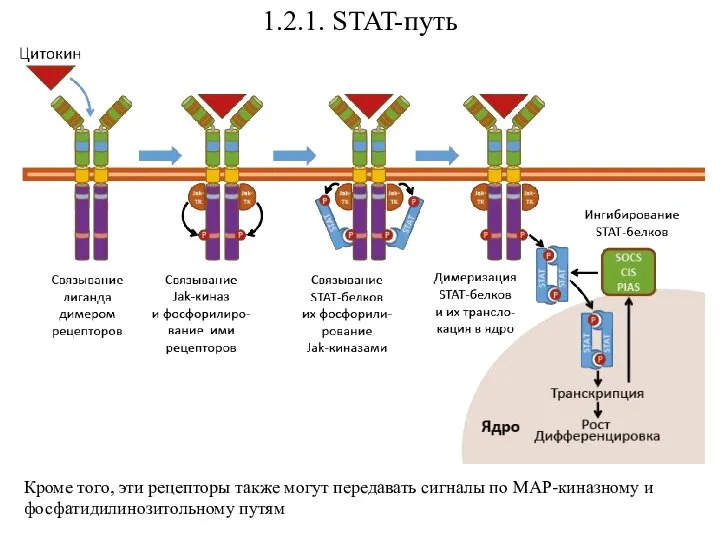
- 24. 1.2.1. STAT-путь Кроме того, эти рецепторы также могут передавать сигналы по МАР-киназному и фосфатидилинозитольному путям
- 25. 2. Рецепторы с ферментативной активностью Рецепторы, обладающие ферментативной активностью – при взаимодействии лиганда с рецептором активируется
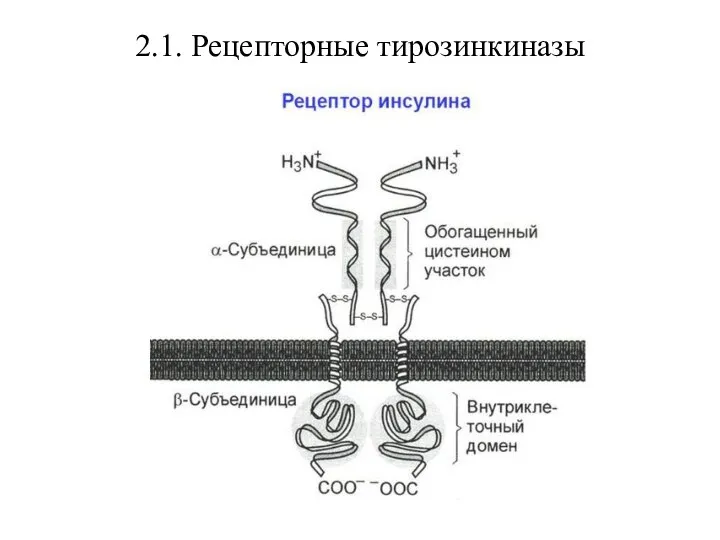
- 26. 2.1. Рецепторные тирозинкиназы Рецепторные тирозинкиназы – трансмембранные белки с ферментативной активностью и одним трансмембранным доменом, связывающие
- 27. 2.1. Рецепторные тирозинкиназы
- 28. 2.1. Рецепторные тирозинкиназы Передача сигнала рецепторными тирозинкиназами осуществляется по фосфоинозитидному (за счёт активации фосфолипазы Сγ) и
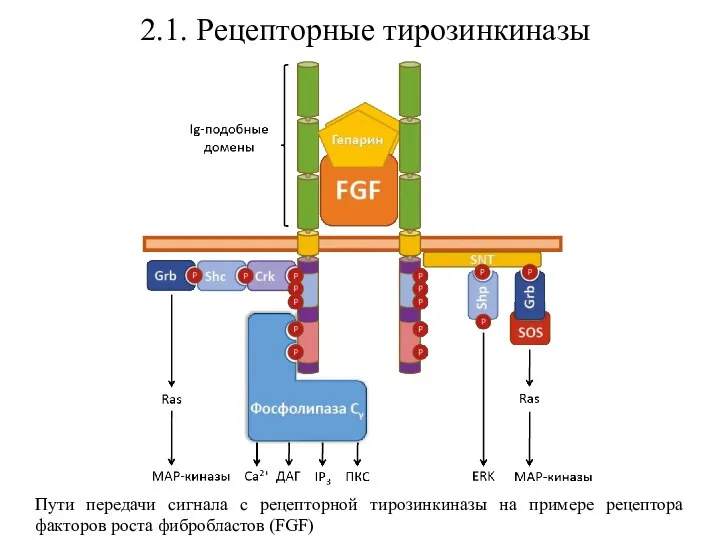
- 29. 2.1. Рецепторные тирозинкиназы Пути передачи сигнала с рецепторной тирозинкиназы на примере рецептора факторов роста фибробластов (FGF)
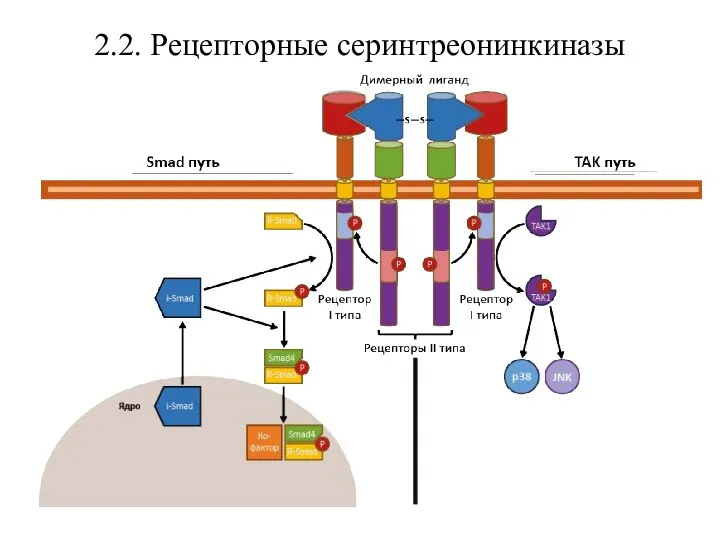
- 30. 2.2. Рецепторные серинтреонинкиназы Рецепторные серинтреонинкиназы – трансмембранные белки с ферментативной активностью, связывающие димерные лиганды (активины, ингибины,
- 31. 2.2.1. Smad-путь После взаимодействия с димером сигнальной молекулы рецепторы II типа активируют рецепторы I типа, позволяя
- 32. 2.2. Рецепторные серинтреонинкиназы
- 33. 2.3. Рецепторные гуанилатциклазы Рецепторные гуанилатциклазы – трансмембранные белки, способные индуцировать синтез цГМФ, связывающие натрийуретические пептиды, гуанилины
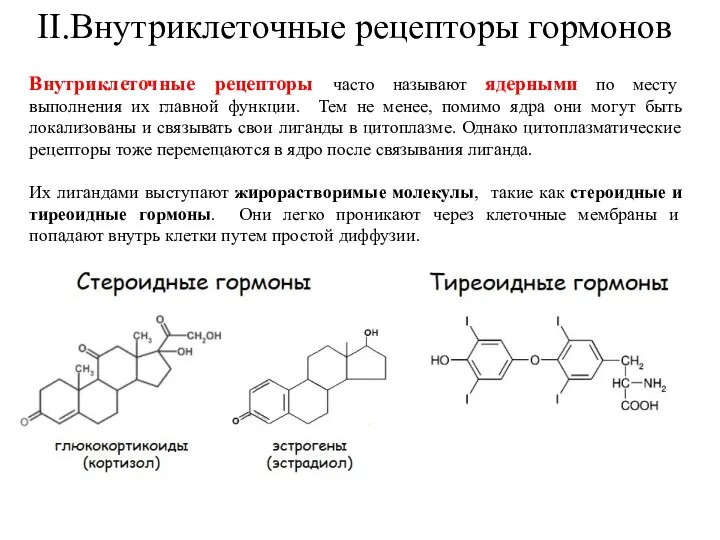
- 34. II.Внутриклеточные рецепторы гормонов Внутриклеточные рецепторы часто называют ядерными по месту выполнения их главной функции. Тем не
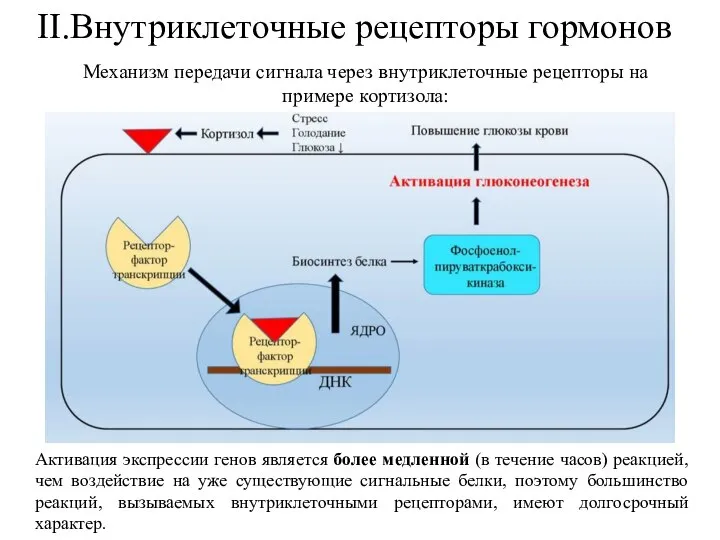
- 35. II.Внутриклеточные рецепторы гормонов Активация экспрессии генов является более медленной (в течение часов) реакцией, чем воздействие на
- 36. II.Внутриклеточные рецепторы гормонов Активированные внутриклеточные рецепторы контролируют экспрессию генов; они связываются со специальными последовательностями на ДНК,
- 38. Скачать презентацию
Слайд 2Рецепторы гормонов
1) Мембранные рецепторы
Клетка
Ядро
2) Внутриклеточные (ядерные) рецепторы
По расположению рецепторы разделяют на:
Рецепторы гормонов
1) Мембранные рецепторы
Клетка
Ядро
2) Внутриклеточные (ядерные) рецепторы
По расположению рецепторы разделяют на:
Слайд 3I. Мембранные рецепторы гормонов
Мембранные рецепторы составляют большинство рецепторов.
Молекула лиганда в данном
I. Мембранные рецепторы гормонов
Мембранные рецепторы составляют большинство рецепторов.
Молекула лиганда в данном

Слайд 4I. Мембранные рецепторы гормонов
Эту группу рецепторов можно разделить на:
1) Рецепторы, не обладающие
I. Мембранные рецепторы гормонов
Эту группу рецепторов можно разделить на:
1) Рецепторы, не обладающие

Слайд 51. Рецепторы без ферментативной активности
Рецепторы без ферментативной активности – подтип трансмембранных
1. Рецепторы без ферментативной активности
Рецепторы без ферментативной активности – подтип трансмембранных

Слайд 61.1.Рецепторы, ассоциированные с G-белками
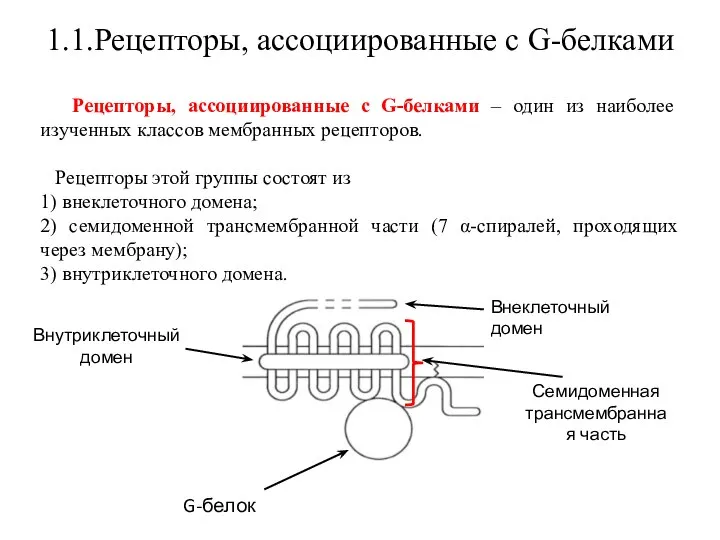
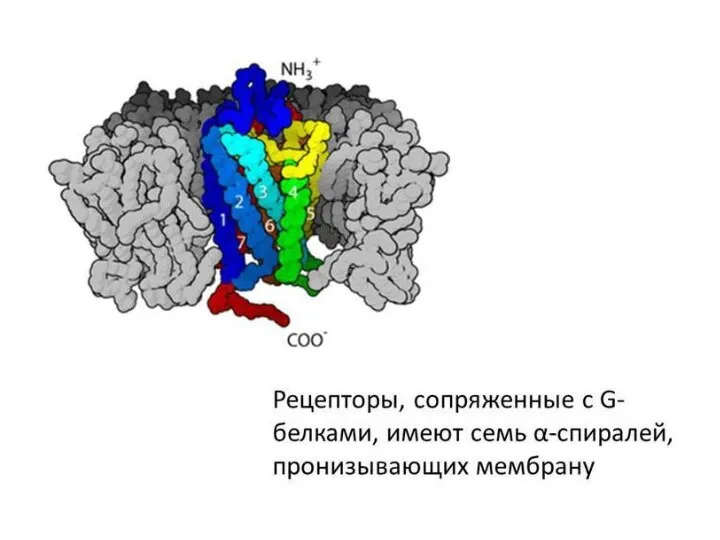
Рецепторы, ассоциированные с G-белками – один из наиболее
1.1.Рецепторы, ассоциированные с G-белками
Рецепторы, ассоциированные с G-белками – один из наиболее
Слайд 81.1.Рецепторы, ассоциированные с G-белками
С такими рецепторами связан ассоциированный с мембраной адапторный
1.1.Рецепторы, ассоциированные с G-белками
С такими рецепторами связан ассоциированный с мембраной адапторный

Слайд 91.1.Рецепторы, ассоциированные с G-белками
Ферментативной (ГТФазной) активностью обладает только
α-субъединица G-белка.
1.1.Рецепторы, ассоциированные с G-белками
Ферментативной (ГТФазной) активностью обладает только
α-субъединица G-белка.

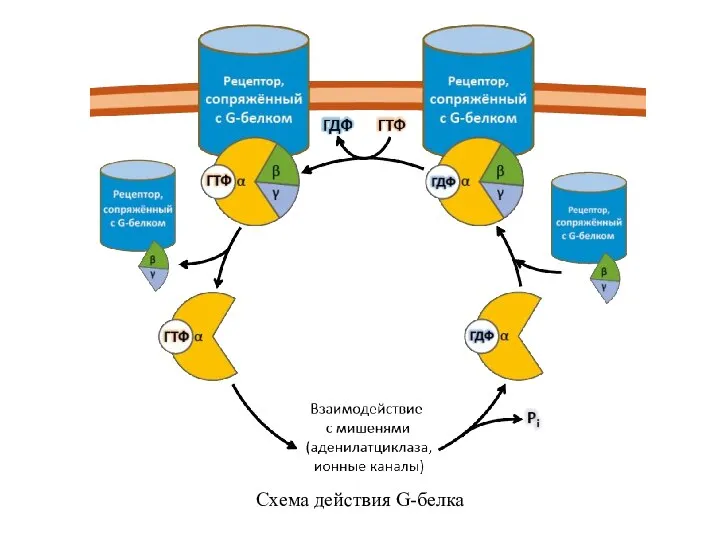
Слайд 10Схема действия G-белка
Схема действия G-белка
Слайд 111.1.Рецепторы, ассоциированные с G-белками
Наиболее хорошо изученные пути передачи сигнала с помощью ассоциированных
1.1.Рецепторы, ассоциированные с G-белками
Наиболее хорошо изученные пути передачи сигнала с помощью ассоциированных

Слайд 121.1.1.Аденилатциклазная система
Работа аденилатциклазной системы связана с ферментом аденилатциклазой и его
1.1.1.Аденилатциклазная система
Работа аденилатциклазной системы связана с ферментом аденилатциклазой и его
Слайд 131.1.1.Аденилатциклазная система
1.1.1.Аденилатциклазная система

Слайд 141.1.1.Аденилатциклазная система
1.1.1.Аденилатциклазная система
Слайд 151.1.2.Инозитолфосфатная система
Эта система связана с активацией фермента фосфолипазы Сβ. Субстратом
1.1.2.Инозитолфосфатная система
Эта система связана с активацией фермента фосфолипазы Сβ. Субстратом
Слайд 161.1.2.Инозитолфосфатная система
Гормоны, использующие инозитолфосфатную систему:
Адреналин (через α-адренорецепторы)
Окситоцин
Вазопрессин (через V1-рецепторы)
Ацетилхолин (через М1-рецепторы)
Гистамин
1.1.2.Инозитолфосфатная система
Гормоны, использующие инозитолфосфатную систему:
Адреналин (через α-адренорецепторы)
Окситоцин
Вазопрессин (через V1-рецепторы)
Ацетилхолин (через М1-рецепторы)
Гистамин

Слайд 171.1.2.Инозитолфосфатная система
1.1.2.Инозитолфосфатная система

Слайд 181.1.2.Инозитолфосфатная система
1.1.2.Инозитолфосфатная система
Слайд 191.1.3. МАР-киназный путь
Помимо указанных выше, для рецепторов, сопряженных с G-белками, существует
1.1.3. МАР-киназный путь
Помимо указанных выше, для рецепторов, сопряженных с G-белками, существует

Слайд 201.1.3. МАР-киназный путь
1.1.3. МАР-киназный путь
Слайд 211.2. Рецепторы, ассоциированные с JAK-киназами
Принципиально отличным классом рецепторных молекул являются рецепторы, сопряженные
1.2. Рецепторы, ассоциированные с JAK-киназами
Принципиально отличным классом рецепторных молекул являются рецепторы, сопряженные

Слайд 221.2. Рецепторы, ассоциированные с JAK-киназами
К рецепторам, сопряжённым с JAK-киназами относятся три семейства:
1.2. Рецепторы, ассоциированные с JAK-киназами
К рецепторам, сопряжённым с JAK-киназами относятся три семейства:

Слайд 231.2. Рецепторы, ассоциированные с JAK-киназами
Структурно каждый мономерный рецептор, ассоциированный с тирозинкиназой класса
1.2. Рецепторы, ассоциированные с JAK-киназами
Структурно каждый мономерный рецептор, ассоциированный с тирозинкиназой класса
Слайд 241.2.1. STAT-путь
Кроме того, эти рецепторы также могут передавать сигналы по МАР-киназному и
1.2.1. STAT-путь
Кроме того, эти рецепторы также могут передавать сигналы по МАР-киназному и
Слайд 252. Рецепторы с ферментативной активностью
Рецепторы, обладающие ферментативной активностью – при взаимодействии
2. Рецепторы с ферментативной активностью
Рецепторы, обладающие ферментативной активностью – при взаимодействии

Слайд 262.1. Рецепторные тирозинкиназы
Рецепторные тирозинкиназы – трансмембранные белки с ферментативной активностью и
2.1. Рецепторные тирозинкиназы
Рецепторные тирозинкиназы – трансмембранные белки с ферментативной активностью и

Слайд 272.1. Рецепторные тирозинкиназы
2.1. Рецепторные тирозинкиназы
Слайд 282.1. Рецепторные тирозинкиназы
Передача сигнала рецепторными тирозинкиназами осуществляется по фосфоинозитидному (за счёт
2.1. Рецепторные тирозинкиназы
Передача сигнала рецепторными тирозинкиназами осуществляется по фосфоинозитидному (за счёт

Слайд 292.1. Рецепторные тирозинкиназы
Пути передачи сигнала с рецепторной тирозинкиназы на примере рецептора факторов
2.1. Рецепторные тирозинкиназы
Пути передачи сигнала с рецепторной тирозинкиназы на примере рецептора факторов
Слайд 302.2. Рецепторные серинтреонинкиназы
Рецепторные серинтреонинкиназы – трансмембранные белки с ферментативной активностью, связывающие димерные
2.2. Рецепторные серинтреонинкиназы
Рецепторные серинтреонинкиназы – трансмембранные белки с ферментативной активностью, связывающие димерные

Слайд 312.2.1. Smad-путь
После взаимодействия с димером сигнальной молекулы рецепторы II типа активируют рецепторы
2.2.1. Smad-путь
После взаимодействия с димером сигнальной молекулы рецепторы II типа активируют рецепторы

Слайд 322.2. Рецепторные серинтреонинкиназы
2.2. Рецепторные серинтреонинкиназы
Слайд 332.3. Рецепторные гуанилатциклазы
Рецепторные гуанилатциклазы – трансмембранные белки, способные индуцировать синтез цГМФ, связывающие
2.3. Рецепторные гуанилатциклазы
Рецепторные гуанилатциклазы – трансмембранные белки, способные индуцировать синтез цГМФ, связывающие

Слайд 34II.Внутриклеточные рецепторы гормонов
Внутриклеточные рецепторы часто называют ядерными по месту выполнения их главной
II.Внутриклеточные рецепторы гормонов
Внутриклеточные рецепторы часто называют ядерными по месту выполнения их главной
Слайд 35II.Внутриклеточные рецепторы гормонов
Активация экспрессии генов является более медленной (в течение часов) реакцией,
II.Внутриклеточные рецепторы гормонов
Активация экспрессии генов является более медленной (в течение часов) реакцией,
Слайд 36II.Внутриклеточные рецепторы гормонов
Активированные внутриклеточные рецепторы контролируют экспрессию генов; они связываются со специальными
II.Внутриклеточные рецепторы гормонов
Активированные внутриклеточные рецепторы контролируют экспрессию генов; они связываются со специальными

 Снег и лед в жизни растений, животных, людей
Снег и лед в жизни растений, животных, людей Мақалдар сәйкестігі. Табиғаты
Мақалдар сәйкестігі. Табиғаты Почемучка
Почемучка Питание и пищеварение
Питание и пищеварение Цепи питания. Поток энергии
Цепи питания. Поток энергии Хромосомная теория наследственности
Хромосомная теория наследственности Приспособляемость живых организмов к суровым условиям Антарктиды
Приспособляемость живых организмов к суровым условиям Антарктиды Нервная система человека
Нервная система человека Трипаносома. Лямблия
Трипаносома. Лямблия Колорад қоңызы
Колорад қоңызы Понятие инноваций и их роль в развитии предприятия
Понятие инноваций и их роль в развитии предприятия Фотосинтез
Фотосинтез Успехи селекции микроорганизмов. Биотехнология и генная инженерия
Успехи селекции микроорганизмов. Биотехнология и генная инженерия Цветок. Строение и значение цветка. 6 класс. Часть 2
Цветок. Строение и значение цветка. 6 класс. Часть 2 Пищеварительная система
Пищеварительная система Растения. Распространение растений
Растения. Распространение растений Почему эволюция - это факт
Почему эволюция - это факт Применения информационных технологий в изучении влияния фитогормонов на рост орхидей
Применения информационных технологий в изучении влияния фитогормонов на рост орхидей Такие большие, маленькие насекомые
Такие большие, маленькие насекомые Генотип и фенотип
Генотип и фенотип Класс Млекопитающие. Отряд Приматы
Класс Млекопитающие. Отряд Приматы Осенние цветы
Осенние цветы Египетская гладкошерстная кошка
Египетская гладкошерстная кошка Презентация на тему ОРГАНЫ И СИСТЕМЫ ОРГАНОВ
Презентация на тему ОРГАНЫ И СИСТЕМЫ ОРГАНОВ  Обмен веществ
Обмен веществ Комнатные растения (часть 1)
Комнатные растения (часть 1) Тонкий кишечник
Тонкий кишечник City farming - the technology of the 3rd millenium
City farming - the technology of the 3rd millenium