- Пищеварение в тонкой кишке

Содержание
- 2. Пищеварение в тонкой кишке Ферменты пристеночного пищеварения более специализированы, это: альфа-глюкозидза, бэтта-галактозидаза, инвертаза, несколько пептидаз, липаза,
- 3. Пищеварение в тонкой кишке Идея мембранного пищеварения была высказана Уголевым А.М. в 1963 г. Проведя опыты
- 4. Взаимоотношения внутриполостного и мембранного пищеварения А — схема последовательной деполимеризации пищевых субстратов в полости и на
- 5. ОСОБЕННОСТИ МЕМБРАННОГО ПИЩЕВАРЕНИЯ Ферменты мембранного пищеварения концентрированы, структурированы, пространственно ориентированы и работают дольше, чем в полости
- 6. Панкреатические ферменты в пристеночном пищеварении
- 7. Ферменты мембраны энтероцитов
- 8. Изолированная петля кишки по Тири-Веллу
- 9. Регуляция секреции кишечного сока Секреторные клетки желез слизистой оболочки тонкой кишки возбуждаются при местном воздействии, в
- 10. Регуляция секреции кишечного сока Выработка пепетидаз не претерпевает существенных изменений даже при резком недостатке белка в
- 11. Стимулирующее действие на либеркюновы и бруннеровы железы оказывают, выделяющиеся из слизистой кишечника энтерокринин и дуокринин. Деятельность
- 13. Физиология всасывания Физиология всасывания физиология всасывания
- 14. У взрослого человека число всасывающих кишечных клеток достигает 1010, а число соматических – 1015, т.е. один
- 15. Характерной особенностью сосудистой системы кишечных ворсинок является высокая степень фенестрирования эндотелия капилляров и большой размер фенестр
- 17. В слизистой тонкого кишечника постоянно осуществляются 2 процесса - секреция - переход веществ из кровеносных капилляров
- 18. Всасывание – это физиологический процесс переноса веществ из просвета ЖКТ во внутреннею среду организма, является конечной
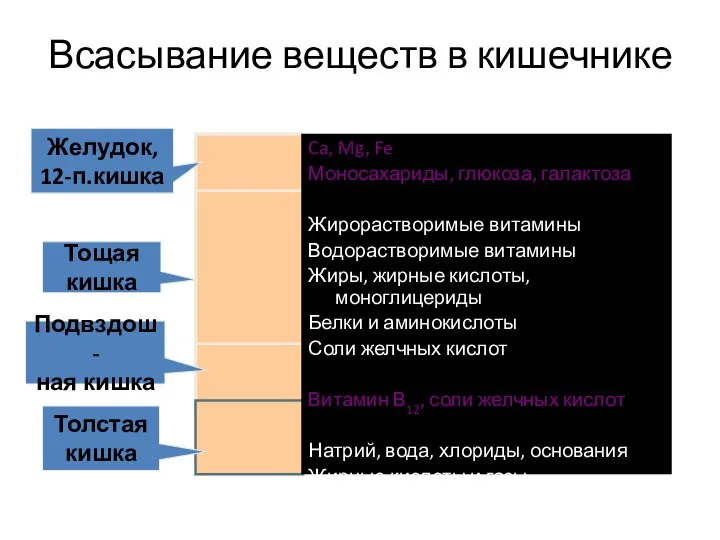
- 19. Топография всасывания веществ из пищеварительного тракта
- 20. Относительный объем всасывания жиров, белков и углеводов в различных отделах тонкой кишки человека 1 — желудок,
- 21. Всасывание осуществляется путём активного и пассивного транспорта. Пассивный транспорт: - диффузия - осмос - фильтрация -
- 22. ОБЩИЕ МЕХАНИЗМЫ ВСАСЫВАНИЯ ПАССИВНЫЙ ТРАНСПОРТ - ПЕРЕНОС БЕЗ ЗАТРАТ ЭНЕРГИИ - - ПЕРЕНОС ПО ГРАДИЕНТАМ ФИЛЬТРАЦИЯ
- 23. Соседние клетки образуют вместе с плотным контактом и межклеточным пространством функциональную единицу. Форма эпителиальных клеток и
- 24. Пассивный перенос через эпителий Зависит от размера пор плотных контактов, который уменьшается в направлении от проксимальных
- 25. Переваривание белков происходит за счет панкреатических протеаз (гидролиз на короткие пептидные фрагменты и АК), расщепление до
- 26. Всасывание белков Нейтральные АК всасываются посредством вторично-активного транспорта с натрием. Na+-независимые переносчики осуществляют перенос части нейтральных
- 27. Предполагают наличие различных транспортных систем для всасывания аминокислот: - нейтральных - основных иминокислот дикарбоновых
- 28. Симпорт аминокислот и Na+
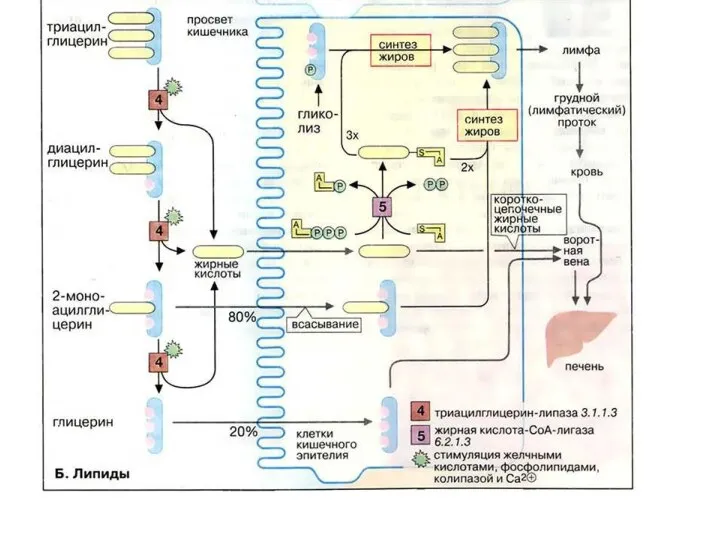
- 29. Переваривание и всасывание жиров 2. Жирные кислоты, содержащие более 10–12 углеродных атомов, в энтероцитах превращаются в
- 30. Строение смешанной мицеллы Эмульгирующее действие желчных кислот на жиры основано в первую очередь на их способности
- 34. Переваривание и всасывание сахаров Связанные с гликокаликсом щеточной каёмки дисахаридазы расщепляют сахара до моносахаридов (главным образом,
- 35. Переваривание и всасывание сахаров всасываются энтероцитами с последующим поступлением в капилляры.
- 36. Всасывание глюкозы
- 37. Схема работы переносчика глюкозы
- 38. Симпорт натрия и глюкозы Глюкоза всасывается в энтероцит путем симпорта совместно с натрием. Последний затем попадает
- 39. Симпорт сахарозы и протонов Сахароза всасывается с помощью симпорта с протонами, выводящимися затем из энтероцитов за
- 40. Всасывание воды Из общего количества жидкости, поступающей в ЖКТ с пищей (2 л) и эндогенными секретами
- 41. A—in vivo, активный транспорт натрия, Б — модель трех компартментов; 1 — полость кишки, 2 —
- 42. Механизм экскреции воды Гипертоничность химуса вызывает движение воды из плазмы в химус посредством осмоса. Каёмчатые клетки
- 43. Механизм всасывания воды Клетки ворсинок «накачивают» Na+ в межклеточное пространство и таким образом компенсируют перемещение Na+
- 44. Всасывание ионов 1) электрогенный транспорт Na+, 2) электрогенный транспорт Na+, сопряженный с транспортом растворимых органических веществ
- 45. Всасывание витаминов Водорастворимые витамины всасываются очень быстро. Всасывание жирорастворимых витаминов A, D, E и K зависит
- 46. Регуляция всасывания Осуществляется за счет изменения процессов кровотока через слизистую кишечника и желудка, лимфотока, а также
- 47. Регуляция всасывания При усилении функциональной активности ЖКТ он может возрастать в 8-10 раз. Это способствует не
- 48. Регуляция всасывания Интенсивность кровотока и особенно лимфотока может также регулироваться за счет сократительной активности ворсинок: имеющиеся
- 50. Регуляция всасывания На активность всасывания оказывает непосредственное влияние гладкая мускулатура кишечника: способствуя перемешиванию химуса и создавая
- 51. Регуляция всасывания Следует отметить, что гормоны, меняющие процесс реабсорбции данного вещества в кишечнике, одновременно, и в
- 52. Пищеварение в толстом кишечнике
- 53. Функции толстого кишечника 1. Поддержание водного и электролитного баланса 2. Участие в углеводном обмене 3. Окончательный
- 54. Сок толстого кишечника Состоит из плотной и жидкой частей и имеет щелочную реакцию (рН 8,5 –
- 55. Реабсорция Na через Na-каналы (проксимальный отдел толстого кишечника)
- 56. Реабсорция ионов Na, Cl и глюкозы в кишечнике.
- 57. Трансэпителиальная секреция NaCl требует активного накопления анионов Сl в клетке.
- 58. Трансэпителиальная секреция NaCl
- 59. Вверху: активное поглощение Na+ (стимулируется альдостероном) и пассивная диффузия К+ в противоположном направлении. Внизу: обмен ионов
- 60. Трансэпителиальная секреция NaHCO3
- 61. Трансэпителиальная секреция КCl
- 62. Стимуляторы секреции 1. Эндогенные стимуляторы 2. Бактериальные энтеротоксины 3. Гормоны (VIP, ADG и др.) 4. Местные
- 63. Микрофлора толстого кишечника Дистальный отдел пищеварительного канала является местом самого обильного размножения микроорганизмов. В химусе толстой
- 64. По подсчётам исследователей из Института Вейцманна : «среднее» человеческое тело весом 70 кг содержит около 30
- 65. Нормальная микрофлора толстого кишечника (эубиоз) делится на три группы: 1-я – (главная) включает бифидобактерии и бактероиды,
- 66. Микроорганизмы, связанные со слизистой кишечника, относятся к мукозной микрофлоре – М-микрофлоре, а локализованные в полости кишки
- 67. защитная функция инактивация тонкокишечных ферментов расщепление компонентов пищеварительных секретов синтез витаминов и др. биологических веществ участие
- 68. Защитная функция Состоит в том, что кишечная микрофлора в организме хозяина действует как постоянный стимул, обусловливающий
- 69. Так называемая, нормальная микрофлора заселяет не только ЖКТ, но и органы дыхания, мочеполовую и репродуктивную системы,
- 70. Рост и развитие бактериальной флоры находится под контролем иммунной системы (иммуноглобулины, лейкоциты слизистой ), влияние т.ж.
- 71. Роль кишечной флоры в регуляции моторной функции толстого кишечника Метаболическая активность кишечной флоры может регулировать моторную
- 72. Роль кишечной флоры в регуляции моторной функции толстого кишечника Микрофлора кишечника синтезирует оксид азота (NO) из
- 73. Детские особенности. Становление кишечной микрофлоры Внутриутробно ребенок развивается со стерильным желудочно-кишечным трактом. Первая фаза формирования его
- 74. Детские особенности. Становление кишечной микрофлоры Во вторую фазу (длительностью 2-4 дня) происходит активное заселение ЖКТ микроорганизмами.
- 75. Детские особенности. Становление кишечной микрофлоры Заселение кишечника новорожденных бифидофлорой тесно связано с грудным вскармливанием. Сначала, в
- 76. Детские особенности. Становление кишечной микрофлоры Третья фаза становления характеризуется стабилизацией микрофлоры, при этом основной становится бифидофлора.
- 77. Для сохранения бифидофлоры имеет значение количество получаемого ребенком женского молока: если оно составляет не менее 1/3
- 78. Детские особенности. Становление кишечной микрофлоры Переход на дефинитивное питание существенно изменяет микрофлору кишечника, появляются новые виды
- 79. МОТОРНАЯ ФУНКЦИЯ ЖКТ МОТОРНАЯ ФУНКЦИЯ ЖКТ МОТОРНАЯ ФУНКЦИЯ ЖКТ
- 80. Оказывает влияние на все этапы процесса пищеварения. Она обеспечивает: механическую обработку пищи, смешивание ее с ферментами,
- 81. Моторная функция желудка Обеспечивается тремя слоями гладкомышечной ткани (продольным, циркулярным и косым). Обеспечивает перемешивание поверхностных слоев
- 82. МОТОРИКА ЖЕЛУДКА
- 83. Рефлекторные дуги рефлексов проксимального отдела желудка: рефлекторная релаксация и рефлекс расширения при наполнении.
- 84. МОТОРИКА РАЗНЫХ ОТДЕЛОВ ЖЕЛУДКА Проксимальный отдел Нет базальной электрической активности, медленные тонические сокращения, высокая растяжимость, основной
- 85. Регистрация моторики желудка у собаки
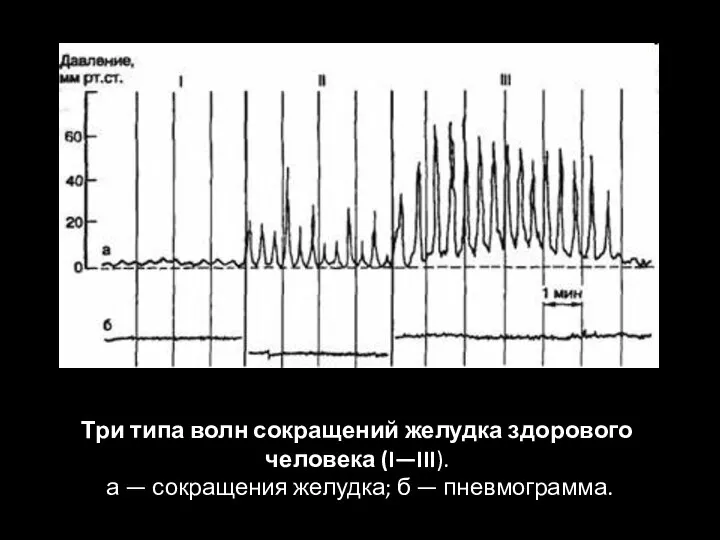
- 86. Три типа волн сокращений желудка здорового человека (I—III). а — сокращения желудка; б — пневмограмма.
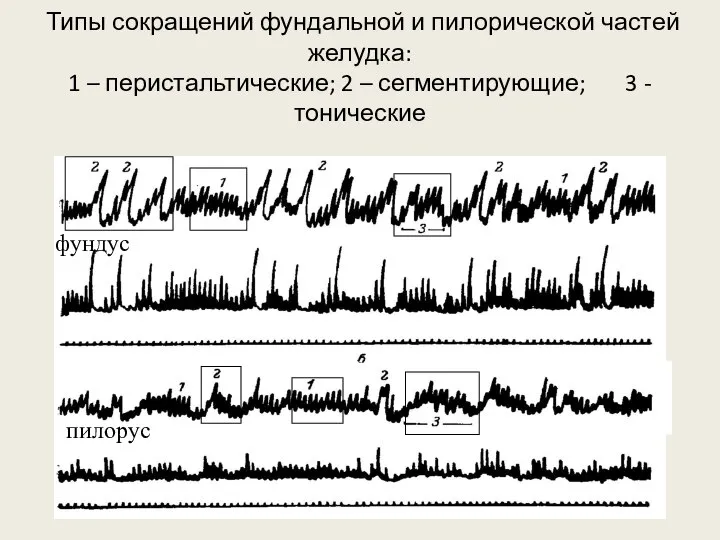
- 87. Типы сокращений фундальной и пилорической частей желудка: 1 – перистальтические; 2 – сегментирующие; 3 - тонические
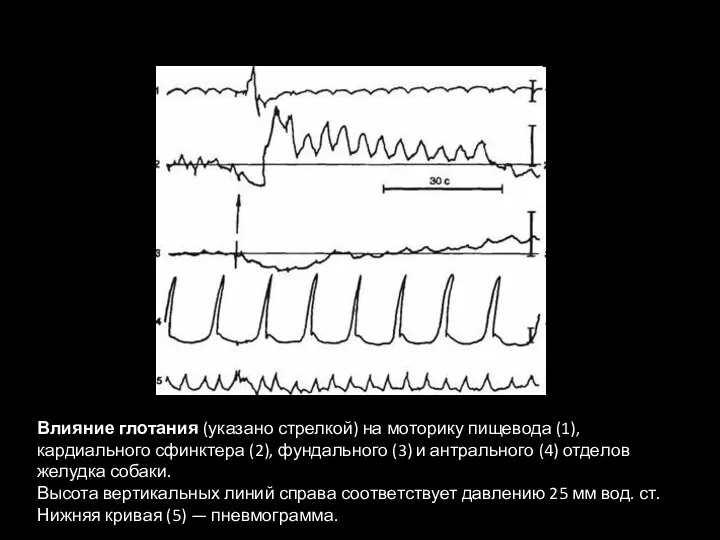
- 88. Влияние глотания (указано стрелкой) на моторику пищевода (1), кардиального сфинктера (2), фундального (3) и антрального (4)
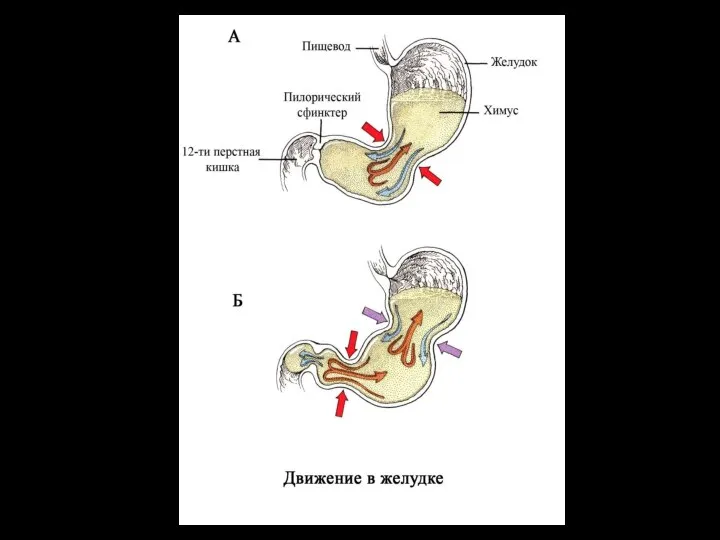
- 91. Факторы, определяющие переход пищи из желудка в двенадцатиперстную кишку: моторика желудка (увеличивает давление в желудке и
- 92. Виды моторики тонкого кишечника 1. Ритмическая сегментация (8-10 в мин) 2. Перистальтика (1-20 см/сек) 3. Маятникообразные
- 93. Моторика кишечника Маятникообразные движения (ритмическая сегментация) Перистальтика
- 94. Виды перистальтики желудочно–кишечного тракта и их функциональное значение
- 95. Взаимосвязь между медленными и быстрыми электрическими волнами и изменением тонуса гладкой мускулатуры тонкой кишки. а —
- 96. Мембранный ПП и ПД ГМК ЖКТ.
- 97. Частота колебаний МП клеток ГМ тонкого кишечника изменяется скачкообразно в проксимально-дистальном направлении.
- 98. Тонические волны (а) и сегментация (б) подвздошной кишки кошки а
- 99. Схема интрамурального перистальтического рефлекса
- 100. Нервная регуляция моторики кишечника симпатикус вагус
- 101. ГУМОРАЛЬНАЯ РЕГУЛЯЦИЯ МОТОРИКИ ТОНКОЙ КИШКИ АМПЛИТУДУ СОКРАЩЕНИЙ УМЕНЬШАЮТ УВЕЛИЧИВАЮТ СЕКРЕТИН ГАСТРИН ГЛЮКАГОН ХОЛЕЦИСТОКИНИН МОТИЛИН ИНСУЛИН
- 102. Зависимость функции илеоцекального сфинктера (баугиниевой заслонки) от давления. При повышении давления в подвздошной кишке тонус илеоцекальной
- 103. Моторика толстого кишечника Время пребывания химуса в толстой кишке исчисляется днями, в целом транзит содержимого по
- 104. Моторика толстого кишечника Только для толстой кишки свойственны масс - сокращения, которые захватывают большую часть кишки
- 105. Моторика толстого кишечника По сравнению с восходящей ободочной кишкой частота сокращений поперечной и нисходящей кишки выше.
- 106. РЕГУЛЯЦИЯ МОТОРИКИ ТОЛСТОГО КИШЕЧНИКА Базальный тонус кишечника задается авторитмической активностью гладкомышечных клеток (циркулярный и продольный слои).
- 107. РЕГУЛЯЦИЯ МОТОРИКИ ТОЛСТОГО КИШЕЧНИКА Экстрамуральная иннервация осуществляется симпатическим и парасимпатическим отделами нервной системы: симпатический отдел -
- 108. РЕГУЛЯЦИЯ МОТОРИКИ ТОЛСТОГО КИШЕЧНИКА Существенную роль в регуляции моторики играют рефлекторные связи толстой кишки с другими
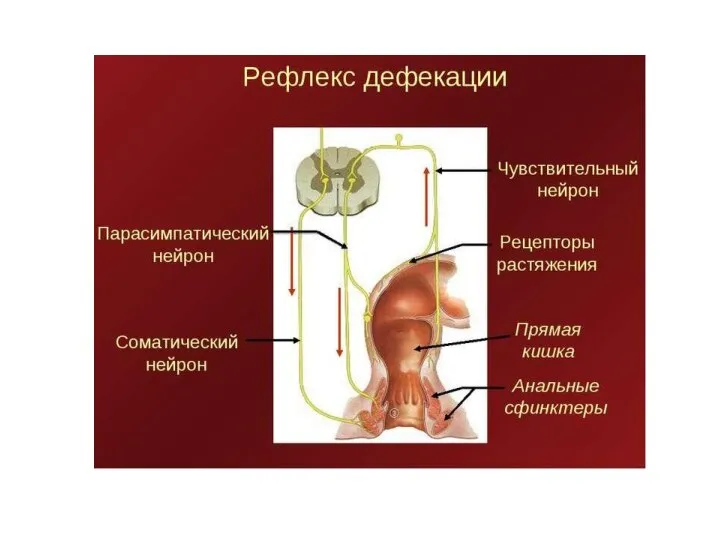
- 109. Схема рефлекторного обеспечения дефекации Р — рецепторы; 1 — внутренний сфинктер заднего прохода, 2 — наружный
- 110. Наличие сфинктеров ЖКТ: 1-7- сфинктеры в полости рта и ротоглотки; 8-перстнеглоточная мышца; 9-верхний сфинктер пищевода; 10-нижний
- 111. РОЛЬ ИНТЕРСТИЦИАЛЬНЫХ КЛЕТОК КАХАЛЯ В МЕХНАНИЗМЕ ЭЛЕКТРОМЕХАНИЧЕСКОГО РИТМОГЕНЕЗА ЖЕЛУДОЧНО-КИШЕЧНОГО ТРАКТА.
- 112. Сантья́го Рамо́н-и-Каха́ль (исп. Santiago Ramón y Cajal) -испанский врач и гистолог, лауреат Нобелевской премии по физиологии
- 113. Keith A. An account of six specimens of the great bowel removed by operation: with some
- 114. Имеется четыре основных популяции ICC в тонком кишечнике мышей. Ультраструктурные особенности ICC, как одного из фенотипов
- 115. Возможности изучения роли ICC были значительно расширены, когда было показано, что в развитии и сохранении структуры
- 116. Кроме этого, в настоящее время для исследования роли ICC в регуляции моторики ЖКТ используются модели мутантных
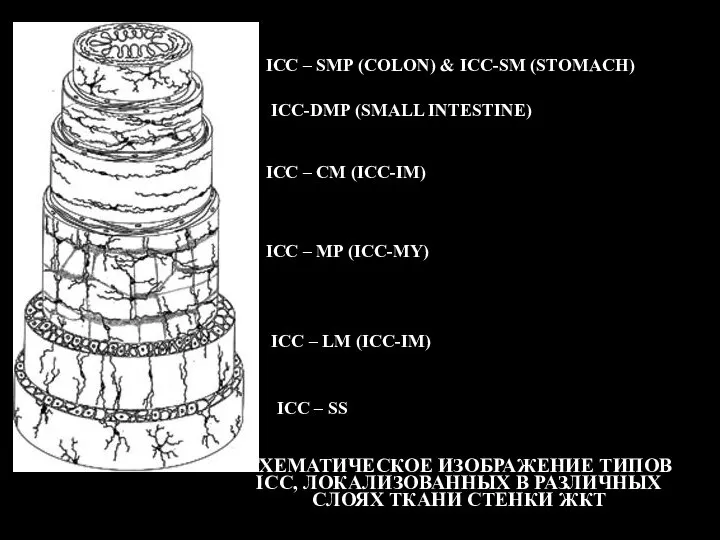
- 117. СХЕМАТИЧЕСКОЕ ИЗОБРАЖЕНИЕ ТИПОВ ICC, ЛОКАЛИЗОВАННЫХ В РАЗЛИЧНЫХ СЛОЯХ ТКАНИ СТЕНКИ ЖКТ ICC – SMP (COLON) &
- 118. Классификация ICC. ICC миэнтерического плексуса (ICC – MP – myenteric plexus). ICC –MP являются мультиполярными, имеющими
- 119. Сопряжение ICC и ГМК. Ультрамикроскопические исследования показывают, что отдельные типы ICC имеют плотные контакты с варикозными
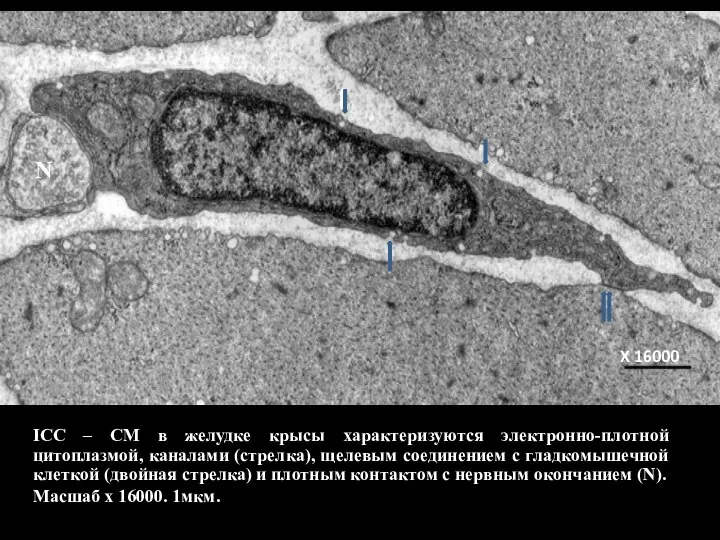
- 120. ICC – CM в желудке крысы характеризуются электронно-плотной цитоплазмой, каналами (стрелка), щелевым соединением с гладкомышечной клеткой
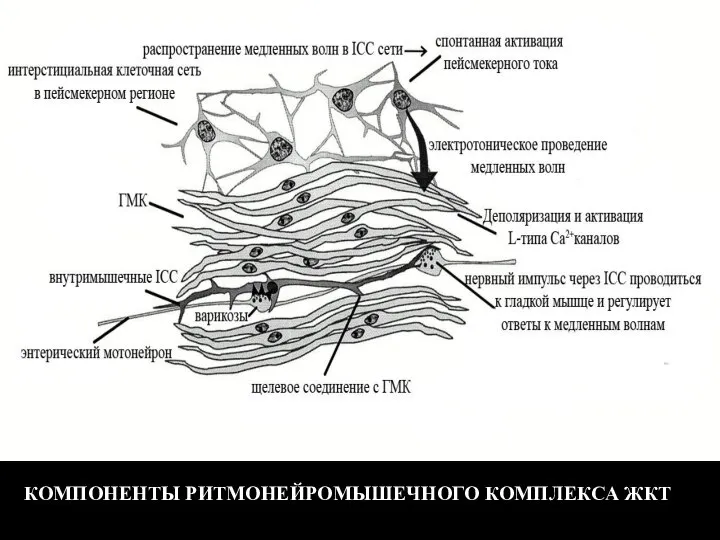
- 121. КОМПОНЕНТЫ РИТМОНЕЙРОМЫШЕЧНОГО КОМПЛЕКСА ЖКТ
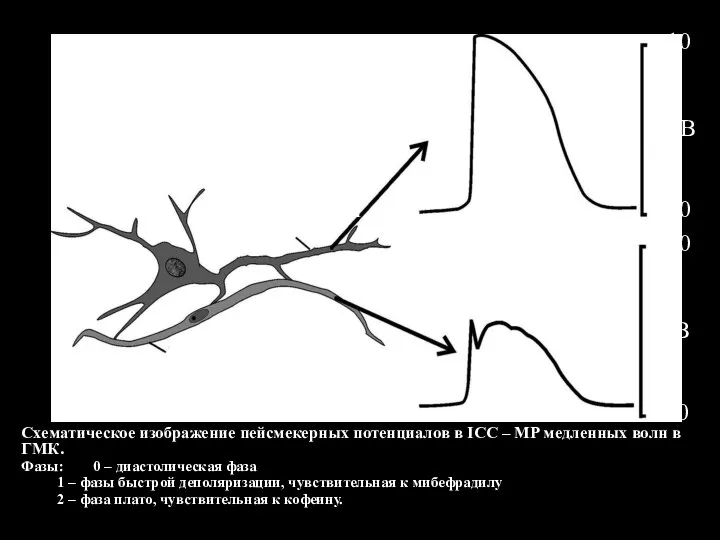
- 122. Схематическое изображение пейсмекерных потенциалов в ICC – MP медленных волн в ГМК. Фазы: 0 – диастолическая
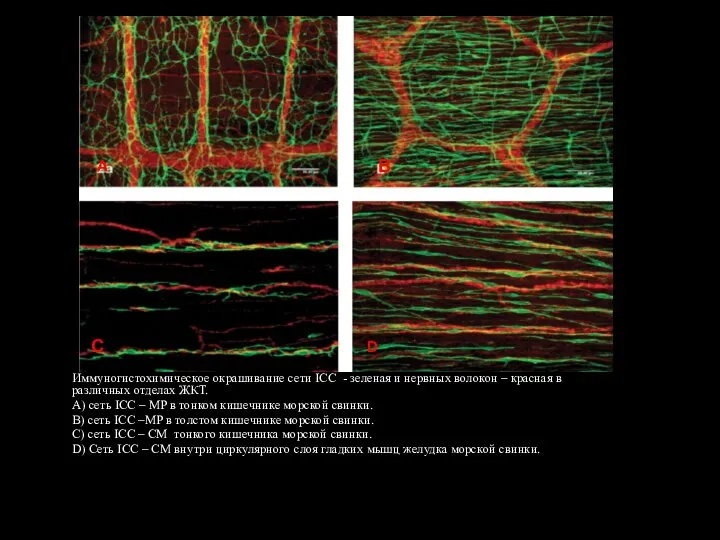
- 123. Иммуногистохимическое окрашивание сети ICC - зеленая и нервных волокон – красная в различных отделах ЖКТ. А)
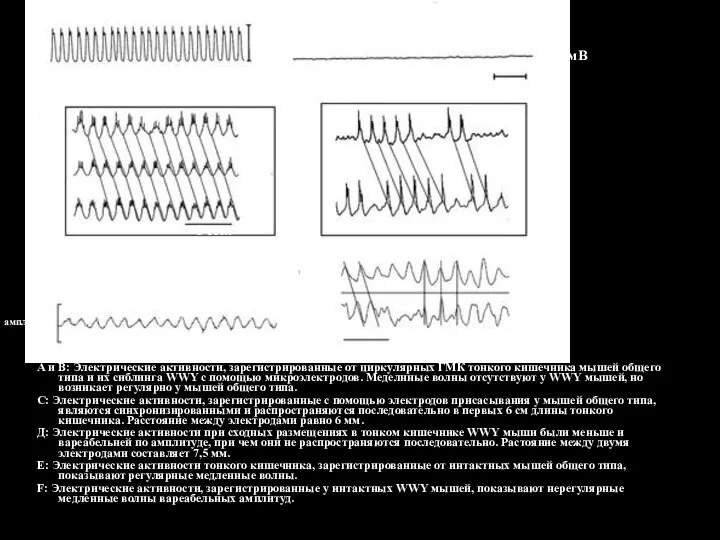
- 124. А и В: Электрические активности, зарегистрированные от циркулярных ГМК тонкого кишечника мышей общего типа и их
- 125. Модель СПР-митохондриального механизма пейсмекерной активности ICC.
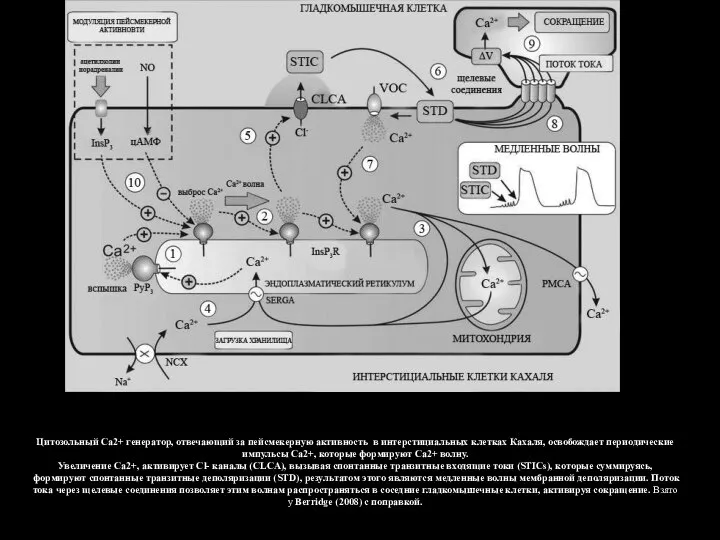
- 126. Цитозольный Са2+ генератор, отвечающий за пейсмекерную активность в интерстициальных клетках Кахаля, освобождает периодические импульсы Са2+, которые
- 127. СПАСИБО ЗА ВНИМАНИЕ
- 128. Всасывание веществ в кишечнике
- 130. Механизмы транспорта. Всасывание веществ осуществляется с участием различных видов транспорта. Пассивный транспорт жидкостей и растворенных в
- 131. Активный транспорт веществ является однонаправленным и связан с затратами энергии, источником которой служат макроэргические фосфаты. При
- 132. Облегченная диффузия, как и простая диффузи, осуществляется без затрат энергии, по градиенту концентрации. Вместе с тем
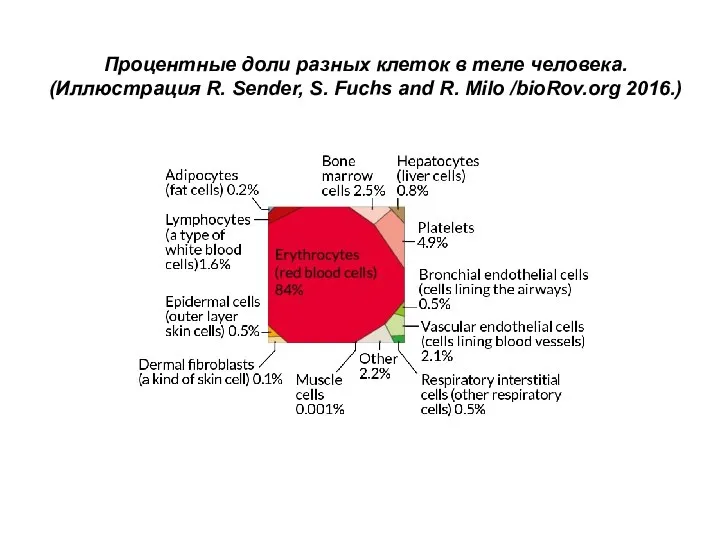
- 133. Процентные доли разных клеток в теле человека. (Иллюстрация R. Sender, S. Fuchs and R. Milo /bioRov.org
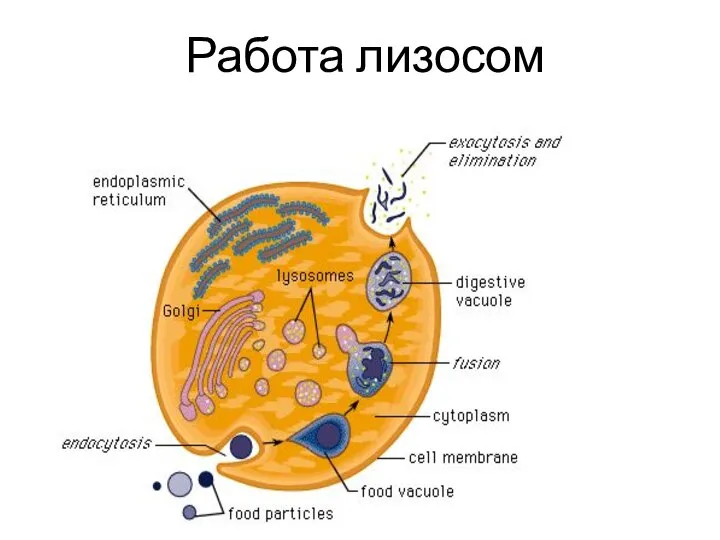
- 134. Работа лизосом
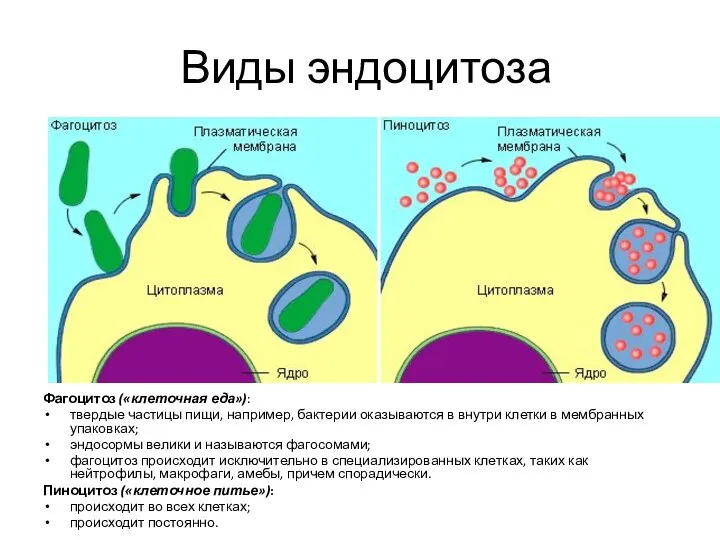
- 135. Виды эндоцитоза Фагоцитоз («клеточная еда»): твердые частицы пищи, например, бактерии оказываются в внутри клетки в мембранных
- 138. Скачать презентацию
Слайд 2Пищеварение в тонкой кишке
Ферменты пристеночного пищеварения более специализированы, это: альфа-глюкозидза, бэтта-галактозидаза, инвертаза,
Пищеварение в тонкой кишке
Ферменты пристеночного пищеварения более специализированы, это: альфа-глюкозидза, бэтта-галактозидаза, инвертаза,

Слайд 3Пищеварение в тонкой кишке
Идея мембранного пищеварения была высказана Уголевым А.М. в 1963
Пищеварение в тонкой кишке
Идея мембранного пищеварения была высказана Уголевым А.М. в 1963

Слайд 4Взаимоотношения внутриполостного и мембранного пищеварения
А — схема последовательной деполимеризации пищевых субстратов
Взаимоотношения внутриполостного и мембранного пищеварения
А — схема последовательной деполимеризации пищевых субстратов

Слайд 5ОСОБЕННОСТИ МЕМБРАННОГО ПИЩЕВАРЕНИЯ
Ферменты мембранного пищеварения концентрированы, структурированы, пространственно ориентированы и работают дольше,
ОСОБЕННОСТИ МЕМБРАННОГО ПИЩЕВАРЕНИЯ
Ферменты мембранного пищеварения концентрированы, структурированы, пространственно ориентированы и работают дольше,

Слайд 6Панкреатические ферменты в пристеночном пищеварении
Панкреатические ферменты в пристеночном пищеварении

Слайд 7Ферменты мембраны энтероцитов
Ферменты мембраны энтероцитов

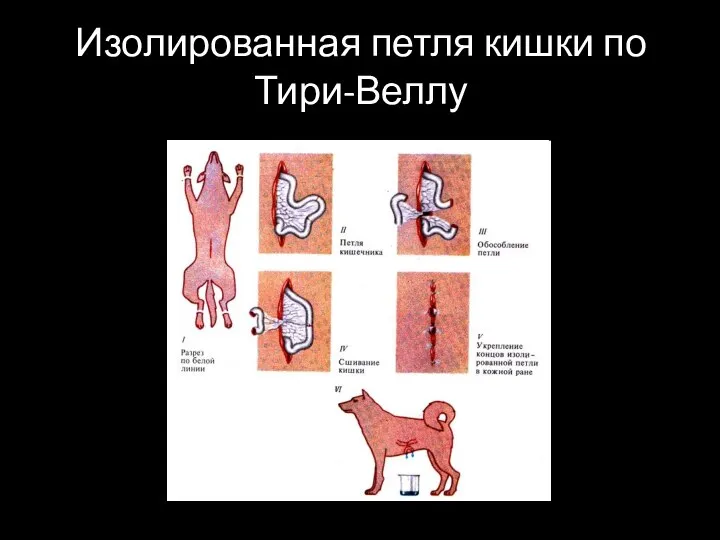
Слайд 8Изолированная петля кишки по Тири-Веллу
Изолированная петля кишки по Тири-Веллу
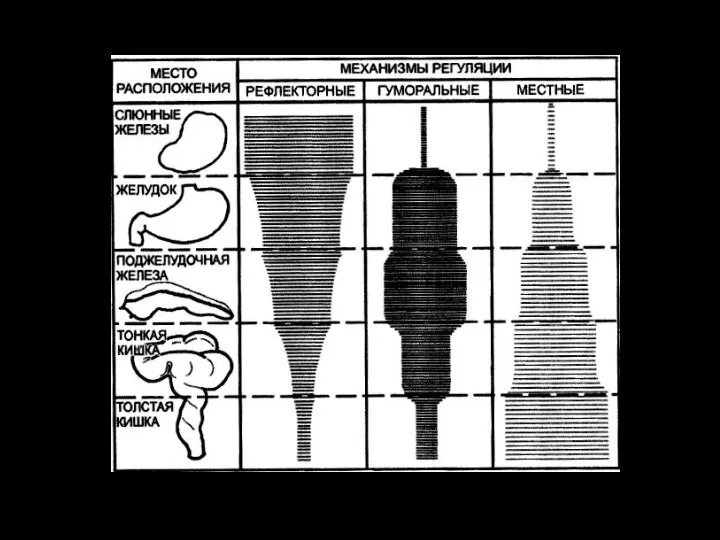
Слайд 9Регуляция секреции кишечного сока
Секреторные клетки желез слизистой оболочки тонкой кишки возбуждаются при
Регуляция секреции кишечного сока
Секреторные клетки желез слизистой оболочки тонкой кишки возбуждаются при

Слайд 10Регуляция секреции кишечного сока
Выработка пепетидаз не претерпевает существенных изменений даже при резком
Регуляция секреции кишечного сока
Выработка пепетидаз не претерпевает существенных изменений даже при резком

Слайд 11Стимулирующее действие на либеркюновы и бруннеровы железы оказывают, выделяющиеся из слизистой кишечника
Стимулирующее действие на либеркюновы и бруннеровы железы оказывают, выделяющиеся из слизистой кишечника

Слайд 13Физиология
всасывания
Физиология
всасывания
физиология всасывания
Физиология
всасывания
Физиология
всасывания
физиология всасывания
Слайд 14У взрослого человека число всасывающих кишечных клеток достигает 1010, а число соматических
У взрослого человека число всасывающих кишечных клеток достигает 1010, а число соматических

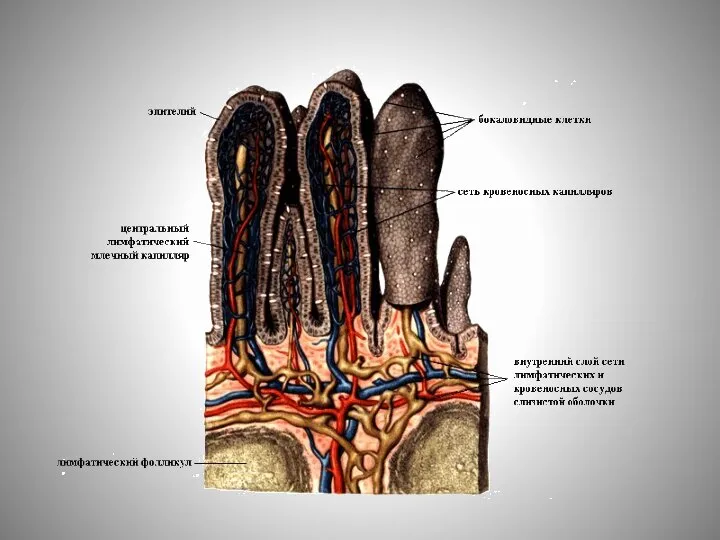
Слайд 15Характерной особенностью сосудистой системы кишечных ворсинок является высокая степень фенестрирования эндотелия капилляров
Характерной особенностью сосудистой системы кишечных ворсинок является высокая степень фенестрирования эндотелия капилляров

Слайд 17В слизистой тонкого кишечника постоянно осуществляются 2 процесса - секреция - переход
В слизистой тонкого кишечника постоянно осуществляются 2 процесса - секреция - переход

Слайд 18Всасывание – это физиологический процесс переноса веществ из просвета ЖКТ во внутреннею
Всасывание – это физиологический процесс переноса веществ из просвета ЖКТ во внутреннею

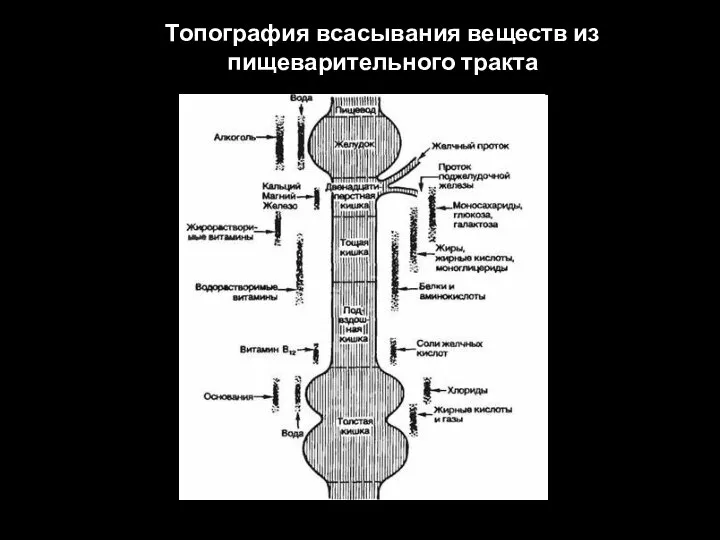
Слайд 19Топография всасывания веществ из пищеварительного тракта
Топография всасывания веществ из пищеварительного тракта
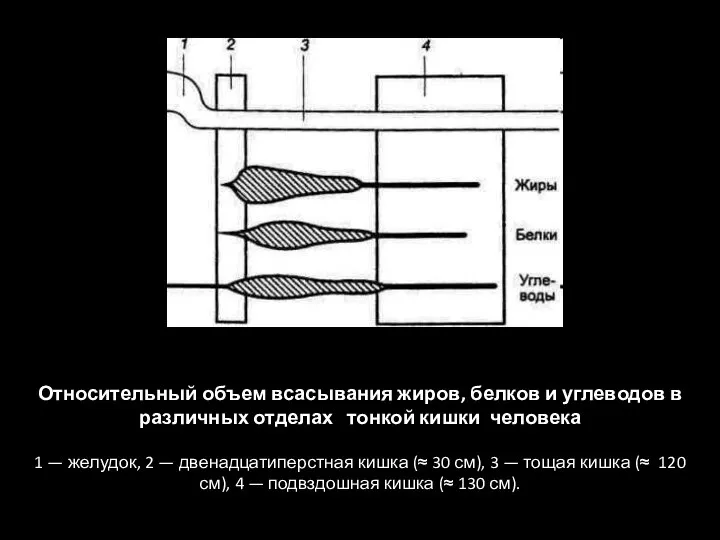
Слайд 20Относительный объем всасывания жиров, белков и углеводов в различных отделах тонкой кишки
Относительный объем всасывания жиров, белков и углеводов в различных отделах тонкой кишки
Слайд 21Всасывание осуществляется путём активного и пассивного транспорта.
Пассивный транспорт:
- диффузия
- осмос
- фильтрация
- персобция
Активный
Всасывание осуществляется путём активного и пассивного транспорта.
Пассивный транспорт:
- диффузия
- осмос
- фильтрация
- персобция
Активный

Слайд 22ОБЩИЕ МЕХАНИЗМЫ ВСАСЫВАНИЯ
ПАССИВНЫЙ ТРАНСПОРТ - ПЕРЕНОС БЕЗ ЗАТРАТ ЭНЕРГИИ -
ОБЩИЕ МЕХАНИЗМЫ ВСАСЫВАНИЯ
ПАССИВНЫЙ ТРАНСПОРТ - ПЕРЕНОС БЕЗ ЗАТРАТ ЭНЕРГИИ -

Слайд 23Соседние клетки образуют вместе с плотным контактом и межклеточным пространством функциональную единицу.
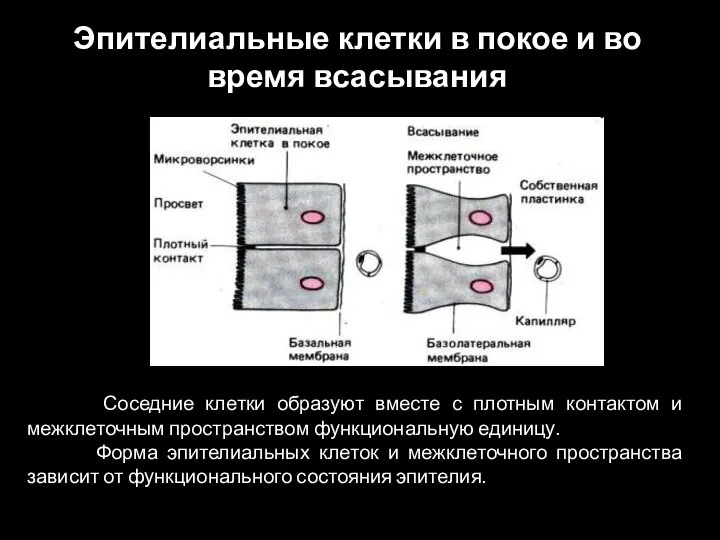
Слайд 24Пассивный перенос через эпителий
Зависит от размера пор плотных контактов, который уменьшается в
Пассивный перенос через эпителий
Зависит от размера пор плотных контактов, который уменьшается в

Слайд 25Переваривание белков
происходит за счет панкреатических протеаз (гидролиз на короткие пептидные фрагменты и
Переваривание белков
происходит за счет панкреатических протеаз (гидролиз на короткие пептидные фрагменты и
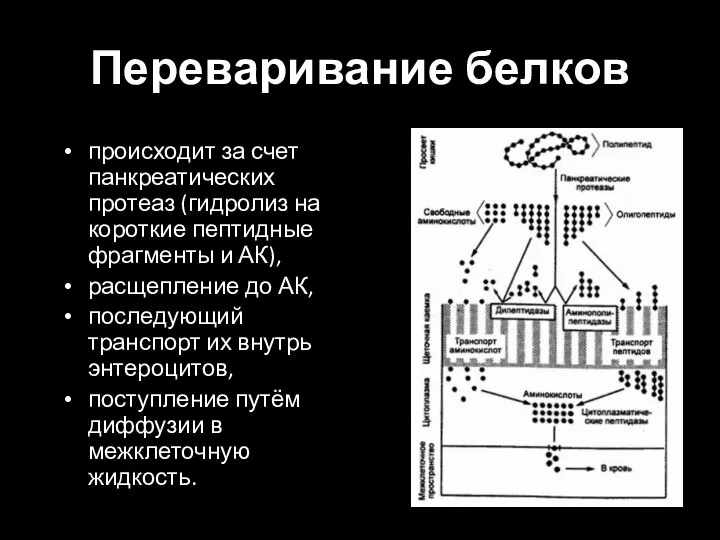
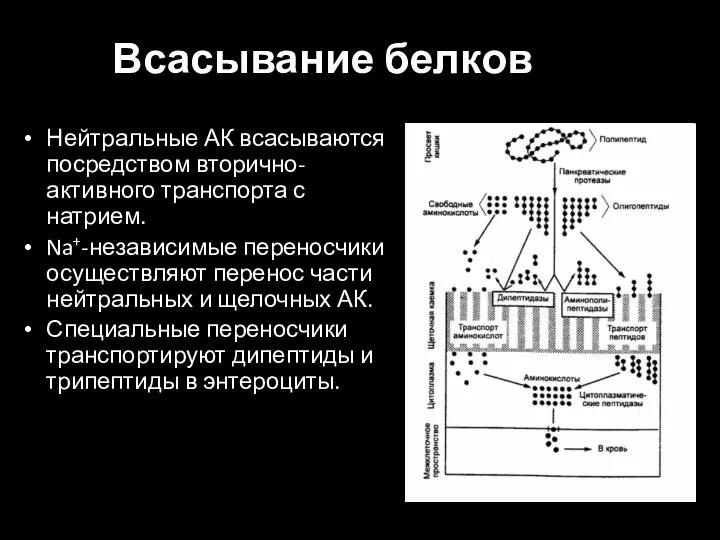
Слайд 26Всасывание белков
Нейтральные АК всасываются посредством вторично-активного транспорта с натрием.
Na+-независимые переносчики осуществляют перенос
Всасывание белков
Нейтральные АК всасываются посредством вторично-активного транспорта с натрием.
Na+-независимые переносчики осуществляют перенос
Слайд 27Предполагают наличие различных транспортных систем для всасывания аминокислот:
- нейтральных
- основных
Предполагают наличие различных транспортных систем для всасывания аминокислот:
- нейтральных
- основных

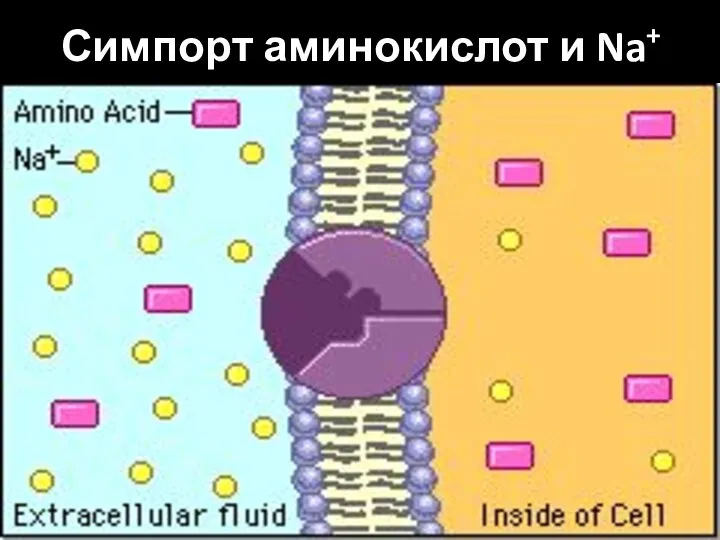
Слайд 28Симпорт аминокислот и Na+
Симпорт аминокислот и Na+
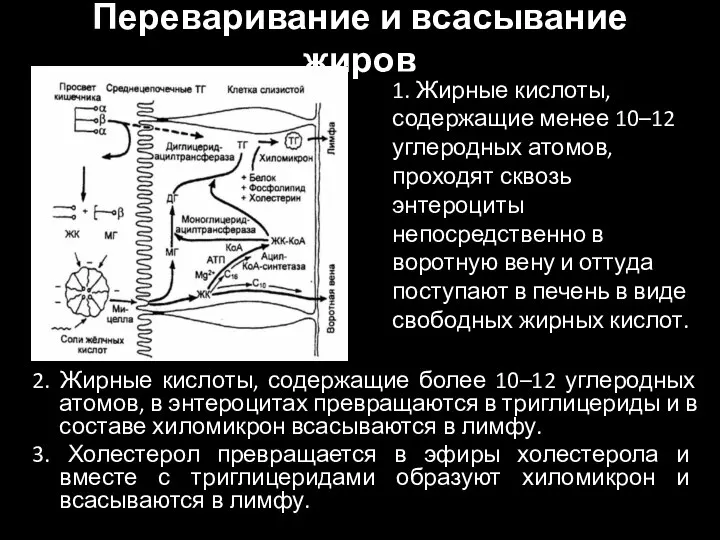
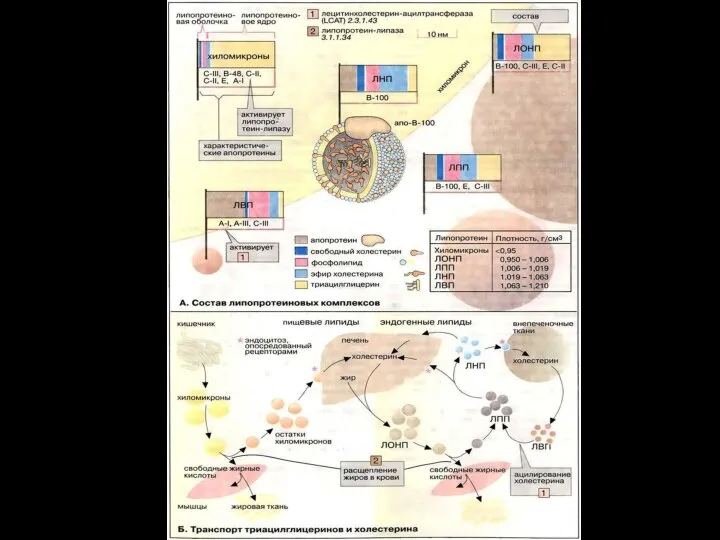
Слайд 29Переваривание и всасывание жиров
2. Жирные кислоты, содержащие более 10–12 углеродных атомов, в
Переваривание и всасывание жиров
2. Жирные кислоты, содержащие более 10–12 углеродных атомов, в
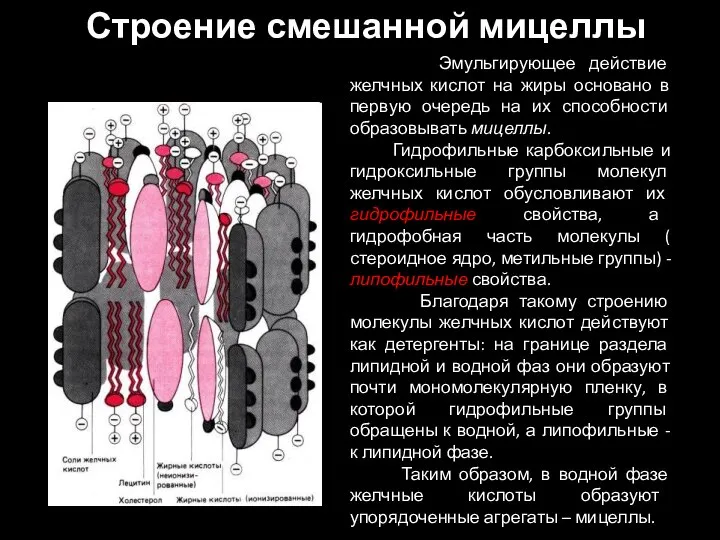
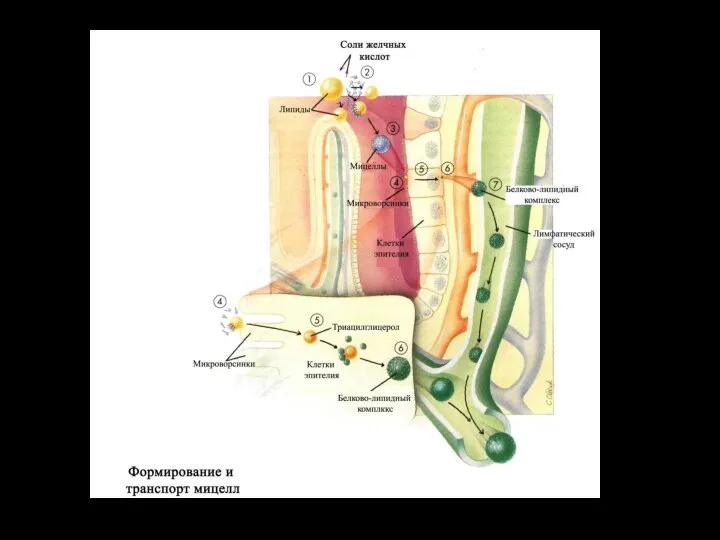
Слайд 30Строение смешанной мицеллы
Эмульгирующее действие желчных кислот на жиры основано в первую
Строение смешанной мицеллы
Эмульгирующее действие желчных кислот на жиры основано в первую

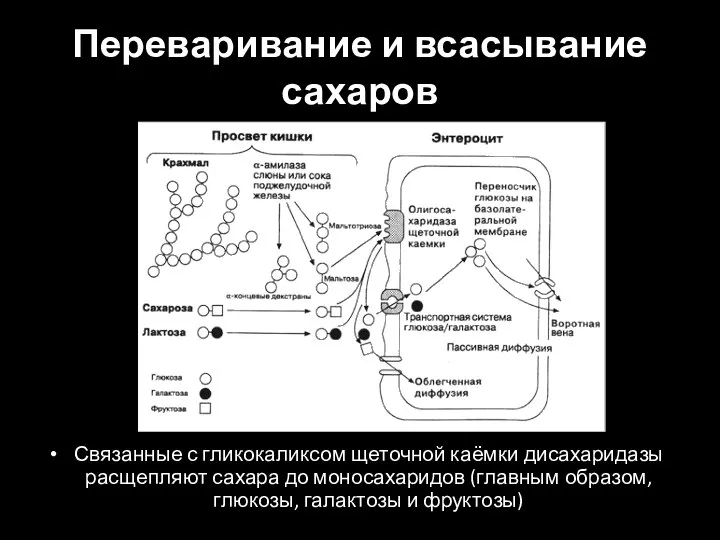
Слайд 34Переваривание и всасывание сахаров
Связанные с гликокаликсом щеточной каёмки дисахаридазы расщепляют сахара до
Переваривание и всасывание сахаров
Связанные с гликокаликсом щеточной каёмки дисахаридазы расщепляют сахара до
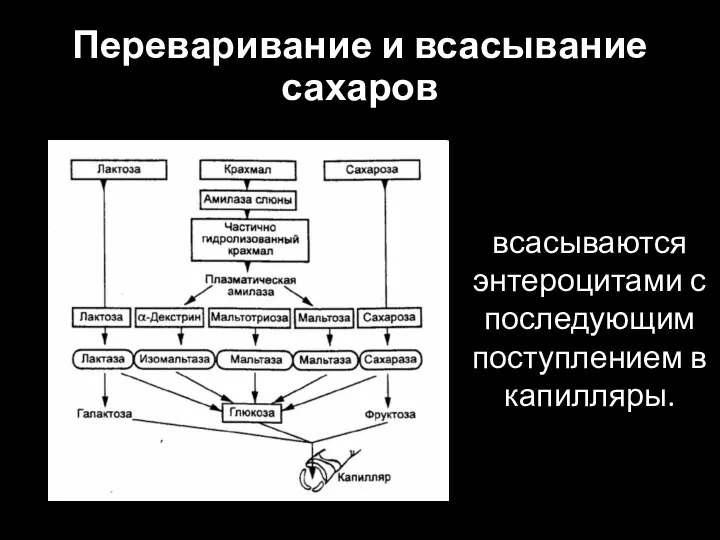
Слайд 35Переваривание и всасывание сахаров
всасываются энтероцитами с последующим поступлением в капилляры.
Переваривание и всасывание сахаров
всасываются энтероцитами с последующим поступлением в капилляры.
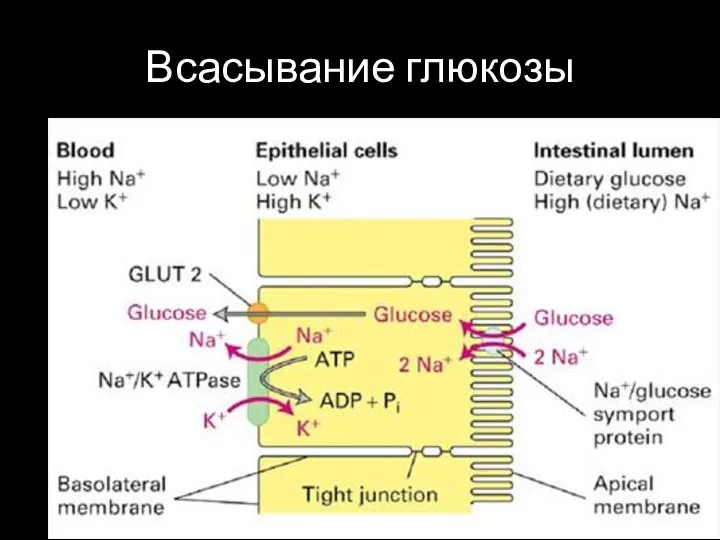
Слайд 36Всасывание глюкозы
Всасывание глюкозы
Слайд 37Схема работы переносчика глюкозы
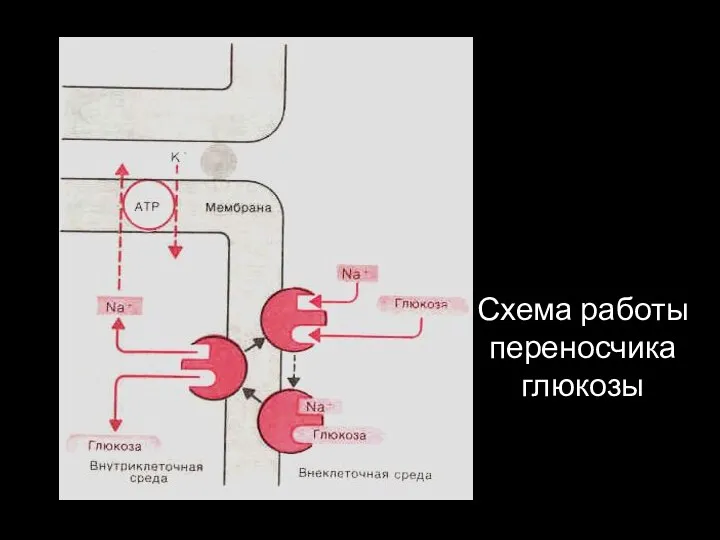
Схема работы переносчика глюкозы
Слайд 38Симпорт натрия и глюкозы
Глюкоза всасывается в энтероцит путем симпорта совместно с натрием.
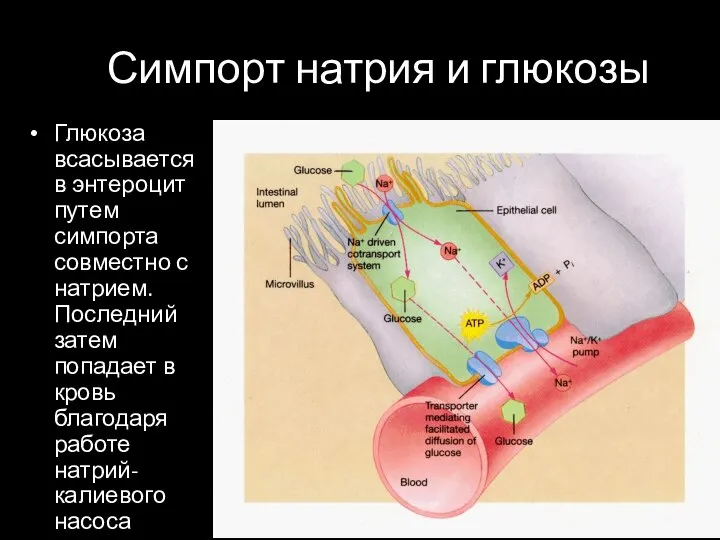
Симпорт натрия и глюкозы
Глюкоза всасывается в энтероцит путем симпорта совместно с натрием.
Слайд 39Симпорт сахарозы и протонов
Сахароза всасывается с помощью симпорта с протонами, выводящимися затем
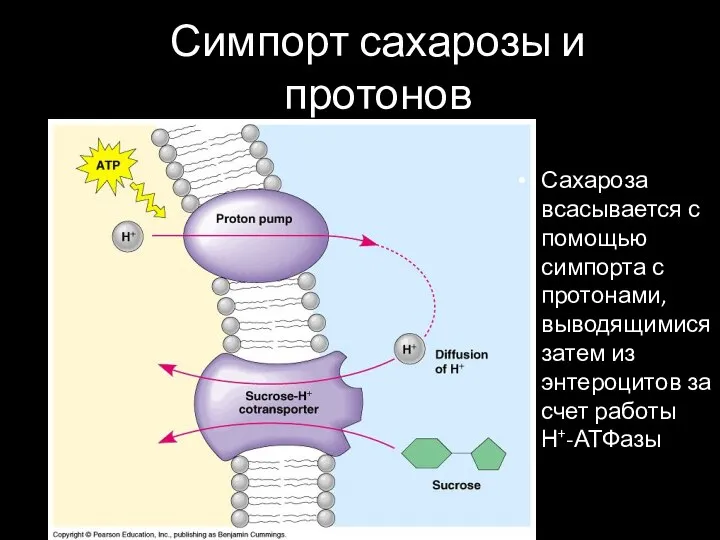
Симпорт сахарозы и протонов
Сахароза всасывается с помощью симпорта с протонами, выводящимися затем
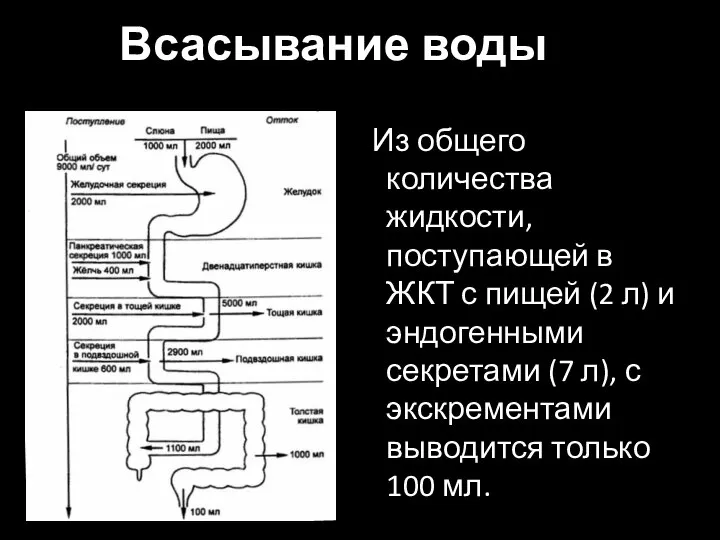
Слайд 40Всасывание воды
Из общего количества жидкости, поступающей в ЖКТ с пищей (2 л)
Всасывание воды
Из общего количества жидкости, поступающей в ЖКТ с пищей (2 л)
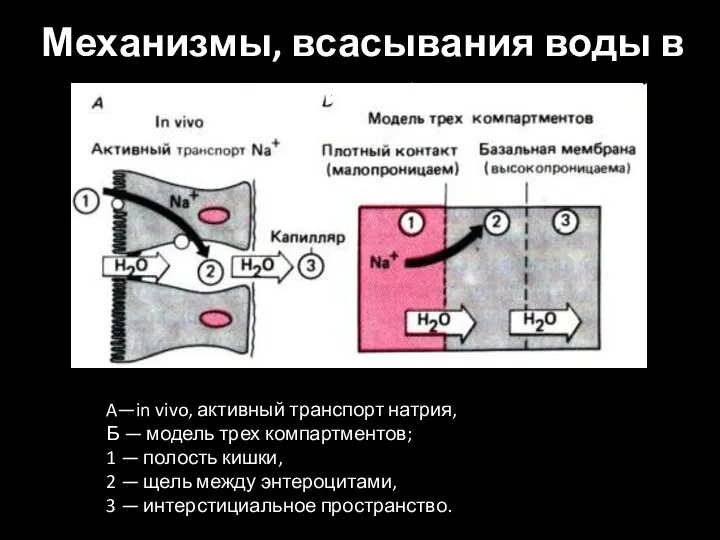
Слайд 41A—in vivo, активный транспорт натрия,
Б — модель трех компартментов;
1 —
Б — модель трех компартментов;
1 —
Слайд 42Механизм
экскреции воды
Гипертоничность химуса вызывает движение воды из плазмы в химус посредством
Механизм
экскреции воды
Гипертоничность химуса вызывает движение воды из плазмы в химус посредством

Слайд 43Механизм
всасывания воды
Клетки ворсинок «накачивают» Na+ в межклеточное пространство и таким образом
Механизм
всасывания воды
Клетки ворсинок «накачивают» Na+ в межклеточное пространство и таким образом

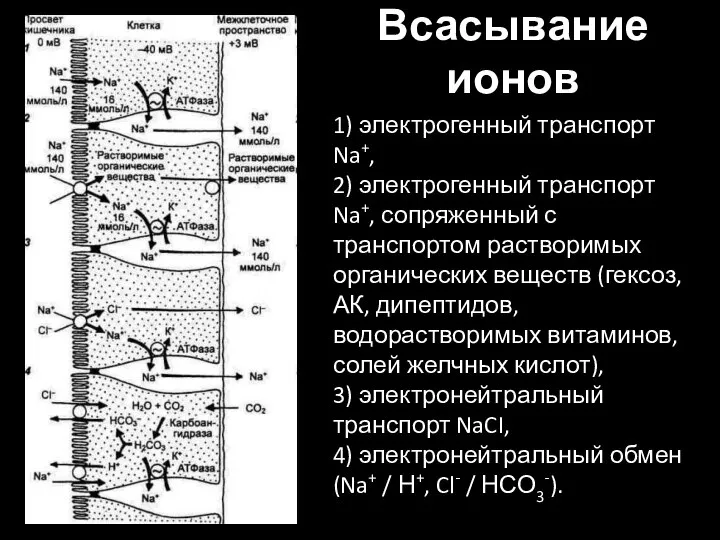
Слайд 44Всасывание ионов
1) электрогенный транспорт Na+,
2) электрогенный транспорт Na+, сопряженный с транспортом
Всасывание ионов
1) электрогенный транспорт Na+, 2) электрогенный транспорт Na+, сопряженный с транспортом
Слайд 45Всасывание витаминов
Водорастворимые витамины всасываются очень быстро.
Всасывание жирорастворимых витаминов A, D, E и
Всасывание витаминов
Водорастворимые витамины всасываются очень быстро.
Всасывание жирорастворимых витаминов A, D, E и

Слайд 46Регуляция всасывания
Осуществляется за счет изменения процессов кровотока через слизистую кишечника и желудка,
Регуляция всасывания
Осуществляется за счет изменения процессов кровотока через слизистую кишечника и желудка,

Слайд 47Регуляция всасывания
При усилении функциональной активности ЖКТ он может возрастать в 8-10 раз.
Регуляция всасывания
При усилении функциональной активности ЖКТ он может возрастать в 8-10 раз.

Слайд 48Регуляция всасывания
Интенсивность кровотока и особенно лимфотока может также регулироваться за счет сократительной
Регуляция всасывания
Интенсивность кровотока и особенно лимфотока может также регулироваться за счет сократительной

Слайд 50Регуляция всасывания
На активность всасывания оказывает непосредственное влияние гладкая мускулатура кишечника: способствуя перемешиванию
Регуляция всасывания
На активность всасывания оказывает непосредственное влияние гладкая мускулатура кишечника: способствуя перемешиванию

Слайд 51Регуляция всасывания
Следует отметить, что гормоны, меняющие процесс реабсорбции данного вещества в кишечнике,
Регуляция всасывания
Следует отметить, что гормоны, меняющие процесс реабсорбции данного вещества в кишечнике,

Слайд 52Пищеварение
в толстом кишечнике
Пищеварение
в толстом кишечнике
Слайд 53Функции толстого кишечника
1. Поддержание водного и электролитного баланса
2. Участие в углеводном обмене
3.
Функции толстого кишечника
1. Поддержание водного и электролитного баланса
2. Участие в углеводном обмене
3.

Слайд 54Сок толстого кишечника
Состоит из плотной и жидкой частей и имеет щелочную реакцию
Сок толстого кишечника
Состоит из плотной и жидкой частей и имеет щелочную реакцию

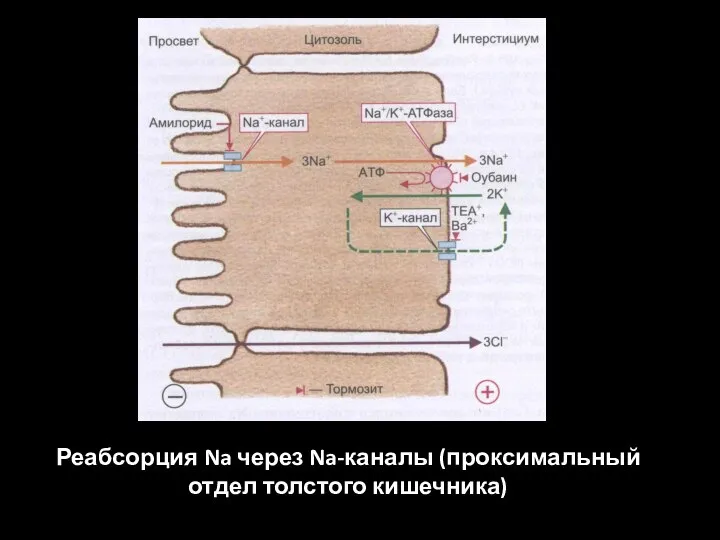
Слайд 55Реабсорция Na через Na-каналы (проксимальный отдел толстого кишечника)
Реабсорция Na через Na-каналы (проксимальный отдел толстого кишечника)
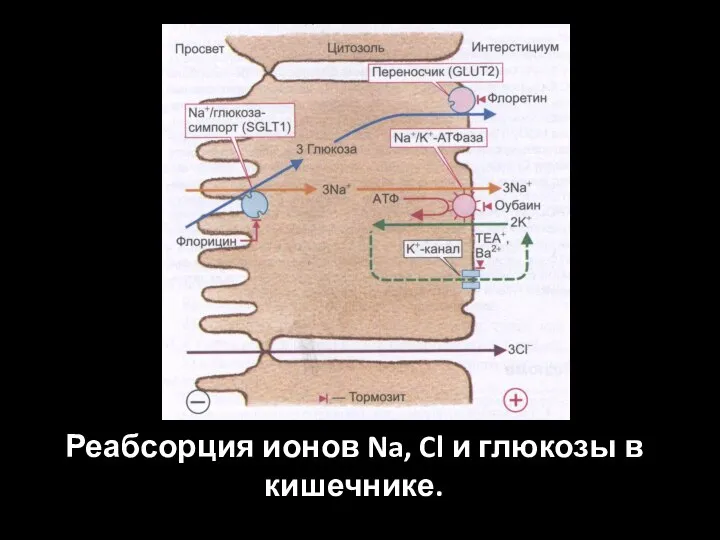
Слайд 56Реабсорция ионов Na, Cl и глюкозы в кишечнике.
Реабсорция ионов Na, Cl и глюкозы в кишечнике.
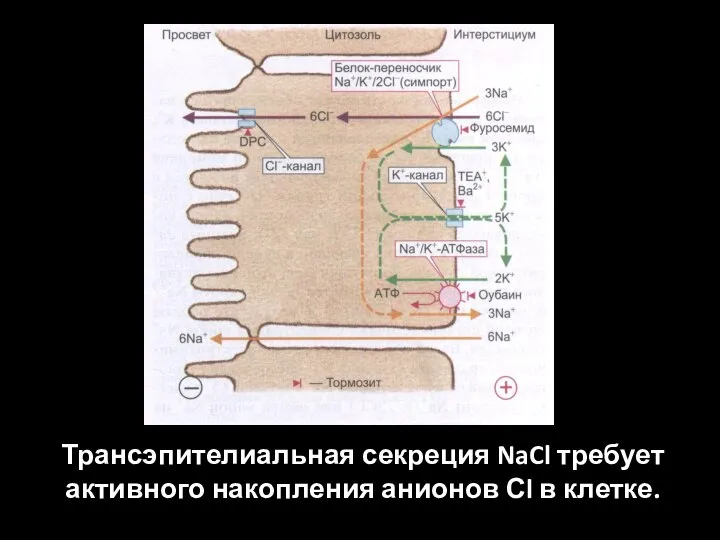
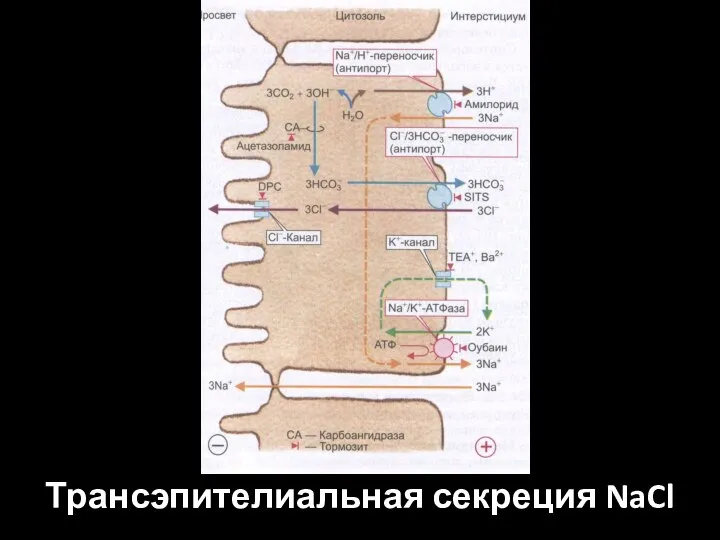
Слайд 57Трансэпителиальная секреция NaCl требует активного накопления анионов Сl в клетке.
Трансэпителиальная секреция NaCl требует активного накопления анионов Сl в клетке.
Слайд 58Трансэпителиальная секреция NaCl
Трансэпителиальная секреция NaCl
Слайд 59Вверху: активное поглощение Na+ (стимулируется альдостероном) и пассивная диффузия К+ в противоположном
Вверху: активное поглощение Na+ (стимулируется альдостероном) и пассивная диффузия К+ в противоположном

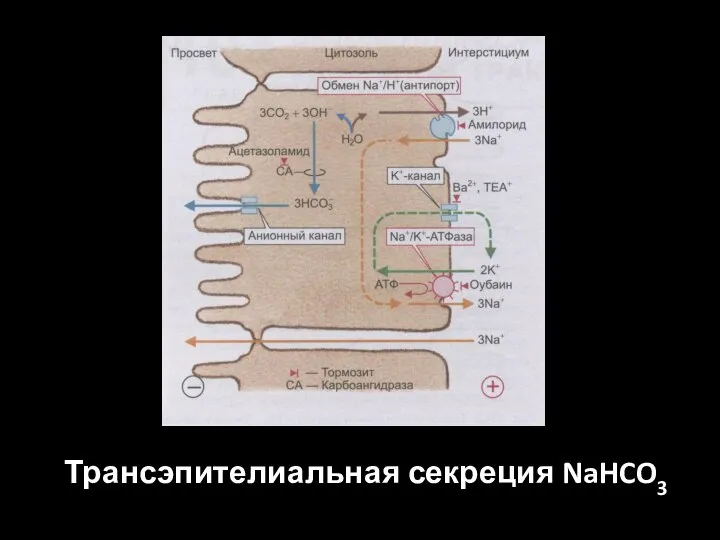
Слайд 60Трансэпителиальная секреция NaHCO3
Трансэпителиальная секреция NaHCO3
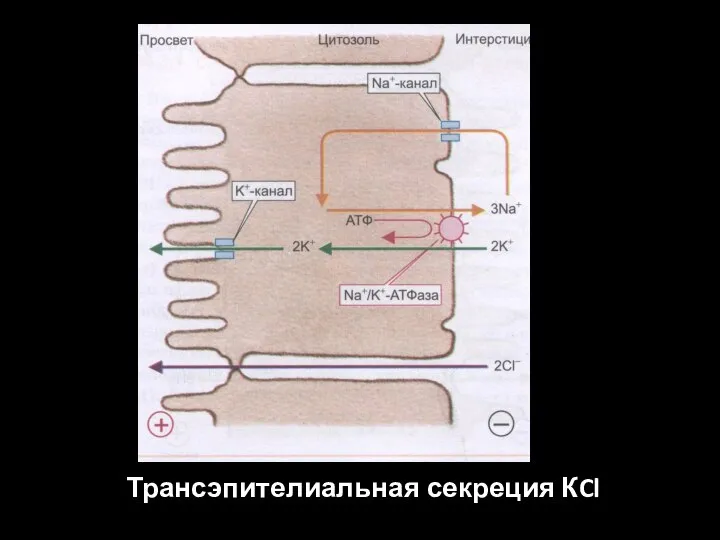
Слайд 61Трансэпителиальная секреция КCl
Трансэпителиальная секреция КCl
Слайд 62Стимуляторы секреции
1. Эндогенные стимуляторы
2. Бактериальные энтеротоксины
3. Гормоны (VIP, ADG и др.)
4. Местные
Стимуляторы секреции
1. Эндогенные стимуляторы
2. Бактериальные энтеротоксины
3. Гормоны (VIP, ADG и др.)
4. Местные

Слайд 63Микрофлора толстого кишечника
Дистальный отдел пищеварительного канала является местом самого обильного
Микрофлора толстого кишечника
Дистальный отдел пищеварительного канала является местом самого обильного

Слайд 64По подсчётам исследователей из Института Вейцманна : «среднее» человеческое тело весом 70
По подсчётам исследователей из Института Вейцманна : «среднее» человеческое тело весом 70

Слайд 65Нормальная микрофлора толстого кишечника (эубиоз) делится на три группы:
1-я – (главная) включает
Нормальная микрофлора толстого кишечника (эубиоз) делится на три группы:
1-я – (главная) включает

Слайд 66Микроорганизмы, связанные со слизистой кишечника, относятся к мукозной микрофлоре – М-микрофлоре, а
Микроорганизмы, связанные со слизистой кишечника, относятся к мукозной микрофлоре – М-микрофлоре, а

Слайд 67защитная функция
инактивация тонкокишечных ферментов
расщепление компонентов пищеварительных секретов
синтез витаминов и др. биологических
защитная функция
инактивация тонкокишечных ферментов
расщепление компонентов пищеварительных секретов
синтез витаминов и др. биологических

Слайд 68Защитная функция
Состоит в том, что кишечная микрофлора в организме хозяина действует
Защитная функция
Состоит в том, что кишечная микрофлора в организме хозяина действует

Слайд 69Так называемая, нормальная микрофлора заселяет не только ЖКТ, но и органы дыхания,
Так называемая, нормальная микрофлора заселяет не только ЖКТ, но и органы дыхания,

Слайд 70Рост и развитие бактериальной флоры находится под контролем иммунной системы (иммуноглобулины, лейкоциты
Рост и развитие бактериальной флоры находится под контролем иммунной системы (иммуноглобулины, лейкоциты

Слайд 71Роль кишечной флоры в регуляции моторной функции толстого кишечника
Метаболическая активность кишечной флоры
Роль кишечной флоры в регуляции моторной функции толстого кишечника
Метаболическая активность кишечной флоры

Слайд 72Роль кишечной флоры в регуляции моторной функции толстого кишечника
Микрофлора кишечника синтезирует оксид
Роль кишечной флоры в регуляции моторной функции толстого кишечника
Микрофлора кишечника синтезирует оксид

Слайд 73Детские особенности. Становление кишечной микрофлоры
Внутриутробно ребенок развивается со стерильным желудочно-кишечным трактом.
Первая
Детские особенности. Становление кишечной микрофлоры
Внутриутробно ребенок развивается со стерильным желудочно-кишечным трактом.
Первая

Слайд 74Детские особенности. Становление кишечной микрофлоры
Во вторую фазу (длительностью 2-4 дня) происходит активное
Детские особенности. Становление кишечной микрофлоры
Во вторую фазу (длительностью 2-4 дня) происходит активное

Слайд 75Детские особенности. Становление кишечной микрофлоры
Заселение кишечника новорожденных бифидофлорой тесно связано с грудным
Детские особенности. Становление кишечной микрофлоры
Заселение кишечника новорожденных бифидофлорой тесно связано с грудным

Слайд 76Детские особенности. Становление кишечной микрофлоры
Третья фаза становления характеризуется стабилизацией микрофлоры, при этом
Детские особенности. Становление кишечной микрофлоры
Третья фаза становления характеризуется стабилизацией микрофлоры, при этом

Слайд 77Для сохранения бифидофлоры имеет значение количество получаемого ребенком женского молока: если оно
Для сохранения бифидофлоры имеет значение количество получаемого ребенком женского молока: если оно

Слайд 78Детские особенности. Становление кишечной микрофлоры
Переход на дефинитивное питание существенно изменяет микрофлору кишечника,
Детские особенности. Становление кишечной микрофлоры
Переход на дефинитивное питание существенно изменяет микрофлору кишечника,

Слайд 79МОТОРНАЯ ФУНКЦИЯ ЖКТ
МОТОРНАЯ ФУНКЦИЯ ЖКТ
МОТОРНАЯ ФУНКЦИЯ ЖКТ
МОТОРНАЯ ФУНКЦИЯ ЖКТ
МОТОРНАЯ ФУНКЦИЯ ЖКТ
МОТОРНАЯ ФУНКЦИЯ ЖКТ
Слайд 80Оказывает влияние на все этапы процесса пищеварения. Она обеспечивает:
механическую обработку пищи,
Оказывает влияние на все этапы процесса пищеварения. Она обеспечивает:
механическую обработку пищи,

Слайд 81Моторная функция желудка
Обеспечивается тремя слоями гладкомышечной ткани (продольным, циркулярным и косым). Обеспечивает
Моторная функция желудка
Обеспечивается тремя слоями гладкомышечной ткани (продольным, циркулярным и косым). Обеспечивает

Слайд 82МОТОРИКА ЖЕЛУДКА
МОТОРИКА ЖЕЛУДКА
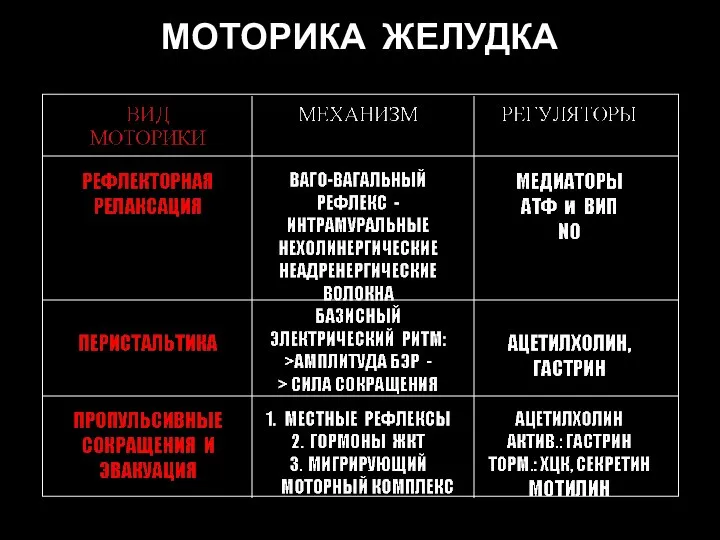
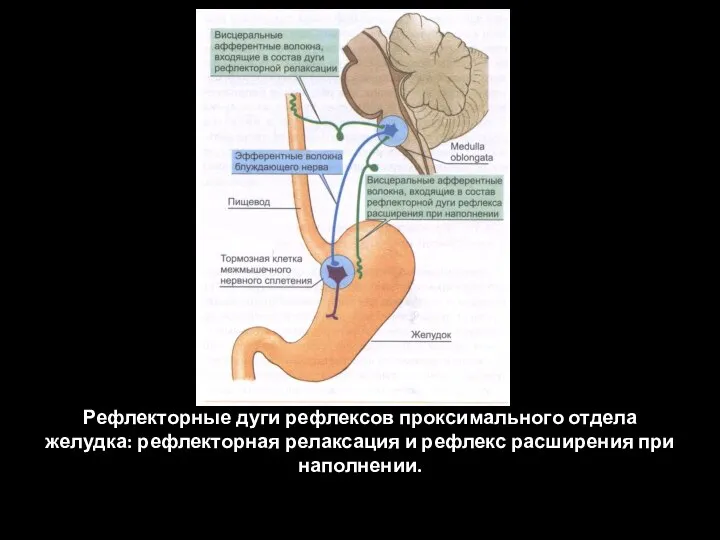
Слайд 83Рефлекторные дуги рефлексов проксимального отдела желудка: рефлекторная релаксация и рефлекс расширения при
Рефлекторные дуги рефлексов проксимального отдела желудка: рефлекторная релаксация и рефлекс расширения при
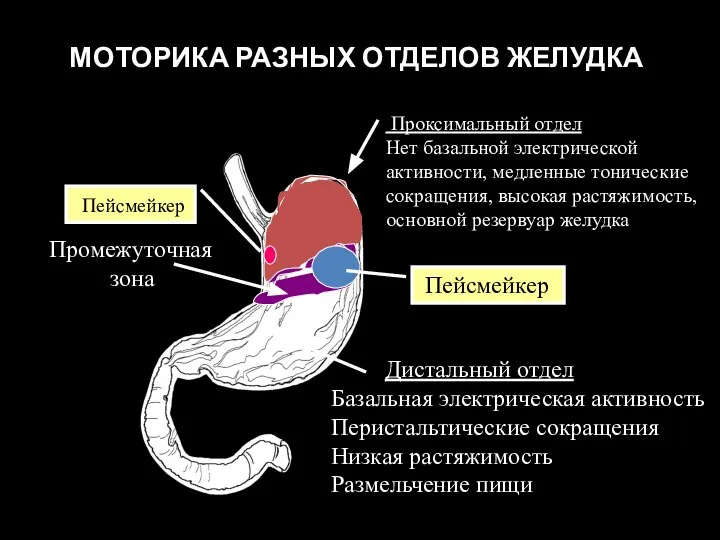
Слайд 84 МОТОРИКА РАЗНЫХ ОТДЕЛОВ ЖЕЛУДКА
Проксимальный отдел
Нет базальной электрической активности, медленные тонические
МОТОРИКА РАЗНЫХ ОТДЕЛОВ ЖЕЛУДКА
Проксимальный отдел
Нет базальной электрической активности, медленные тонические
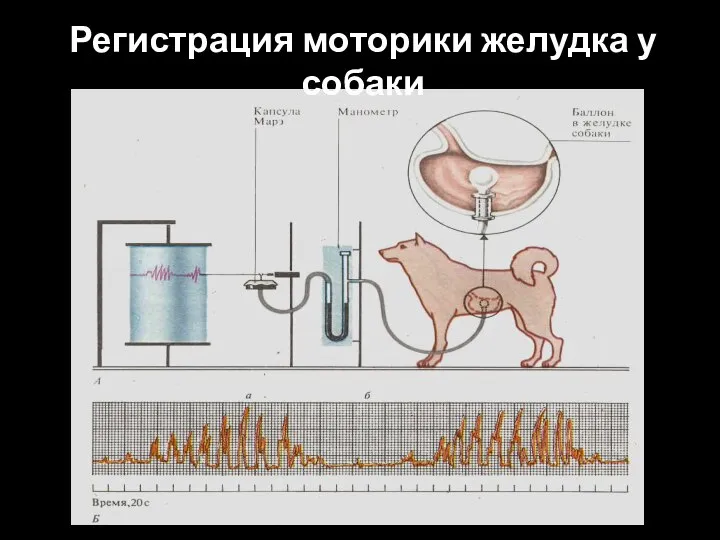
Слайд 85Регистрация моторики желудка у собаки
Регистрация моторики желудка у собаки
Слайд 86Три типа волн сокращений желудка здорового человека (I—III).
а — сокращения желудка;
Три типа волн сокращений желудка здорового человека (I—III).
а — сокращения желудка;
Слайд 87Типы сокращений фундальной и пилорической частей желудка:
1 – перистальтические; 2 –
Типы сокращений фундальной и пилорической частей желудка: 1 – перистальтические; 2 –
Слайд 88Влияние глотания (указано стрелкой) на моторику пищевода (1), кардиального сфинктера (2), фундального
Влияние глотания (указано стрелкой) на моторику пищевода (1), кардиального сфинктера (2), фундального
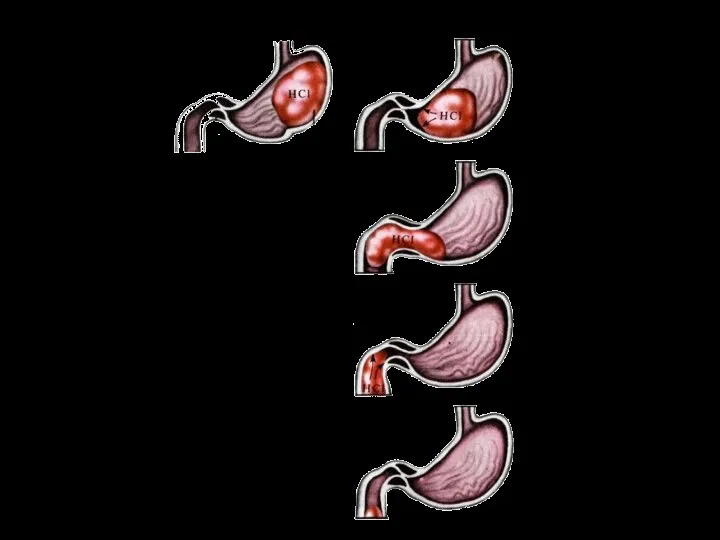
Слайд 91Факторы, определяющие переход пищи из желудка в двенадцатиперстную кишку:
моторика желудка (увеличивает давление
Факторы, определяющие переход пищи из желудка в двенадцатиперстную кишку:
моторика желудка (увеличивает давление

Слайд 92Виды моторики тонкого кишечника
1. Ритмическая сегментация (8-10 в мин)
2. Перистальтика (1-20 см/сек)
3.
Виды моторики тонкого кишечника
1. Ритмическая сегментация (8-10 в мин)
2. Перистальтика (1-20 см/сек)
3.

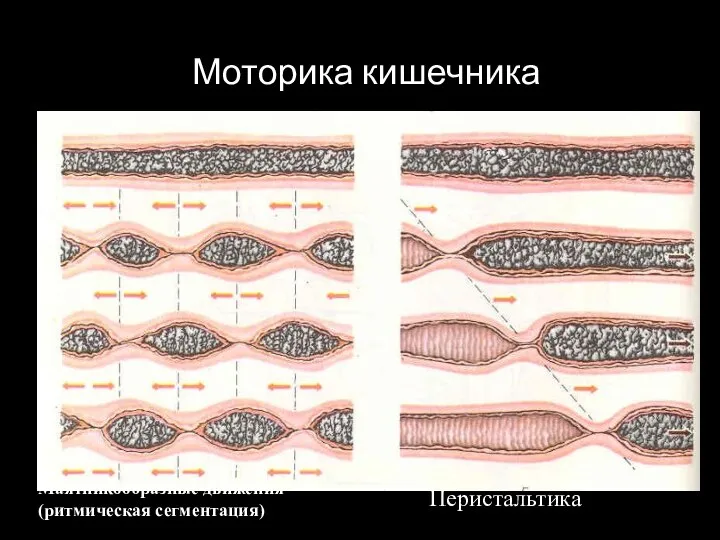
Слайд 93Моторика кишечника
Маятникообразные движения (ритмическая сегментация)
Перистальтика
Моторика кишечника
Маятникообразные движения (ритмическая сегментация)
Перистальтика
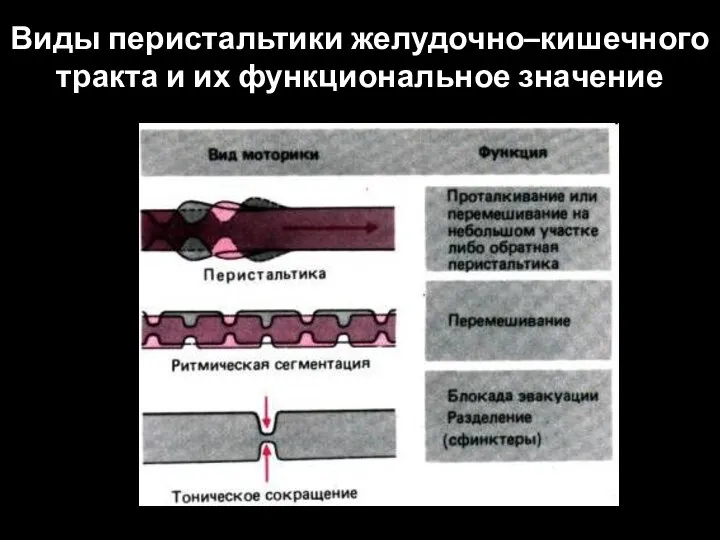
Слайд 94Виды перистальтики желудочно–кишечного тракта и их функциональное значение
Виды перистальтики желудочно–кишечного тракта и их функциональное значение
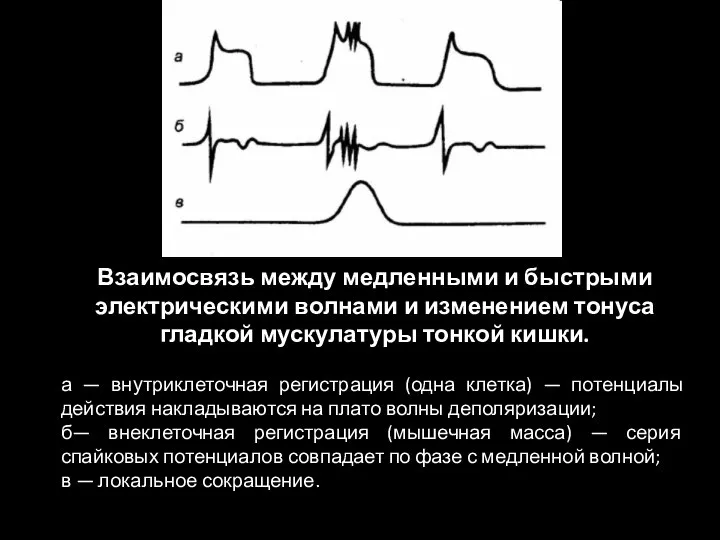
Слайд 95Взаимосвязь между медленными и быстрыми электрическими волнами и изменением тонуса гладкой мускулатуры
Взаимосвязь между медленными и быстрыми электрическими волнами и изменением тонуса гладкой мускулатуры
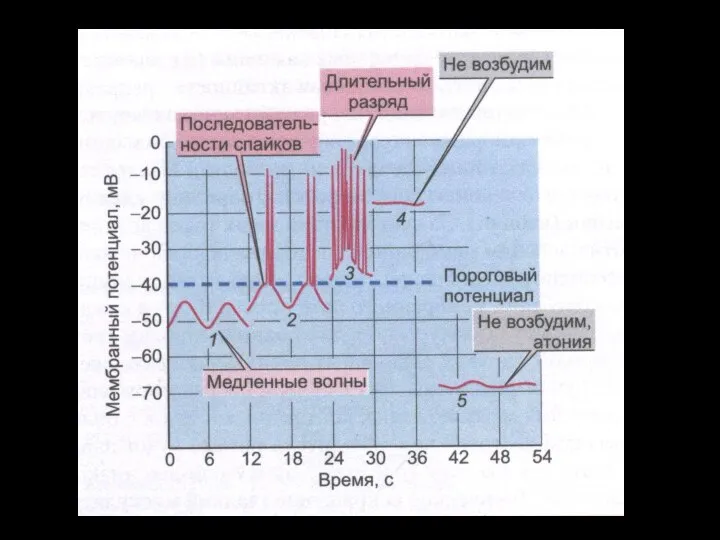
Слайд 96Мембранный ПП и ПД ГМК ЖКТ.
Мембранный ПП и ПД ГМК ЖКТ.
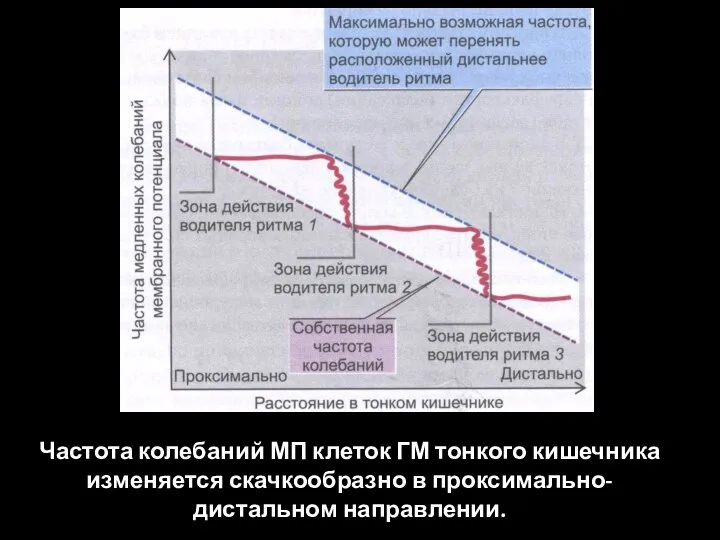
Слайд 97Частота колебаний МП клеток ГМ тонкого кишечника изменяется скачкообразно в проксимально-дистальном направлении.
Частота колебаний МП клеток ГМ тонкого кишечника изменяется скачкообразно в проксимально-дистальном направлении.
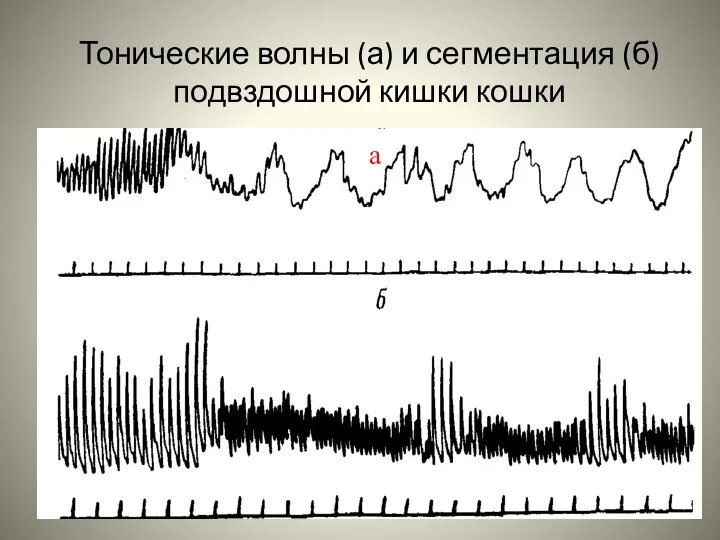
Слайд 98Тонические волны (а) и сегментация (б) подвздошной кишки кошки
а
Тонические волны (а) и сегментация (б) подвздошной кишки кошки
а
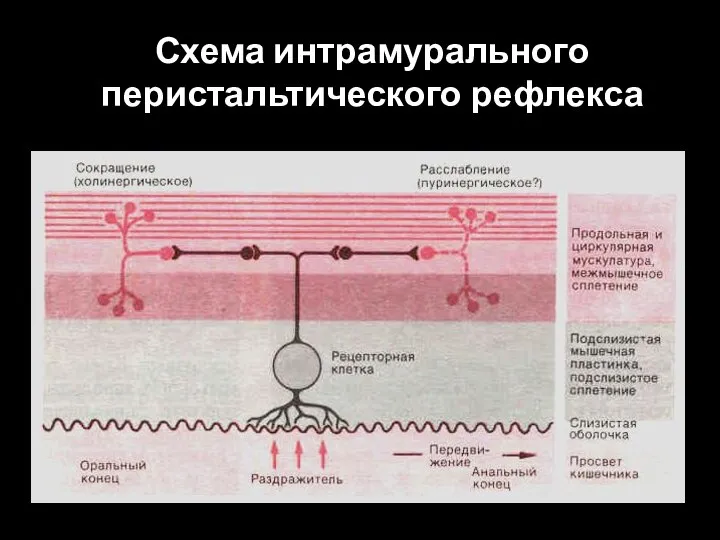
Слайд 99Схема интрамурального перистальтического рефлекса
Схема интрамурального перистальтического рефлекса
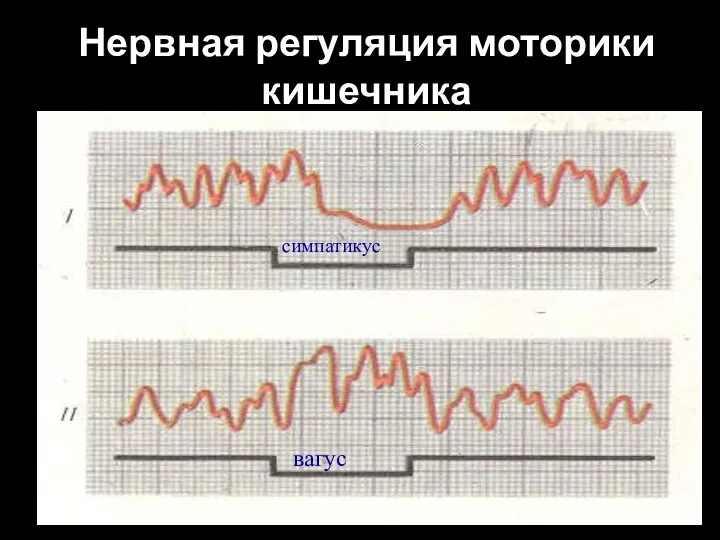
Слайд 100Нервная регуляция моторики кишечника
симпатикус
вагус
Нервная регуляция моторики кишечника
симпатикус
вагус
Слайд 101ГУМОРАЛЬНАЯ РЕГУЛЯЦИЯ МОТОРИКИ ТОНКОЙ КИШКИ
АМПЛИТУДУ СОКРАЩЕНИЙ
УМЕНЬШАЮТ УВЕЛИЧИВАЮТ
СЕКРЕТИН ГАСТРИН
ГЛЮКАГОН ХОЛЕЦИСТОКИНИН
МОТИЛИН
ГУМОРАЛЬНАЯ РЕГУЛЯЦИЯ МОТОРИКИ ТОНКОЙ КИШКИ
АМПЛИТУДУ СОКРАЩЕНИЙ
УМЕНЬШАЮТ УВЕЛИЧИВАЮТ
СЕКРЕТИН ГАСТРИН
ГЛЮКАГОН ХОЛЕЦИСТОКИНИН
МОТИЛИН

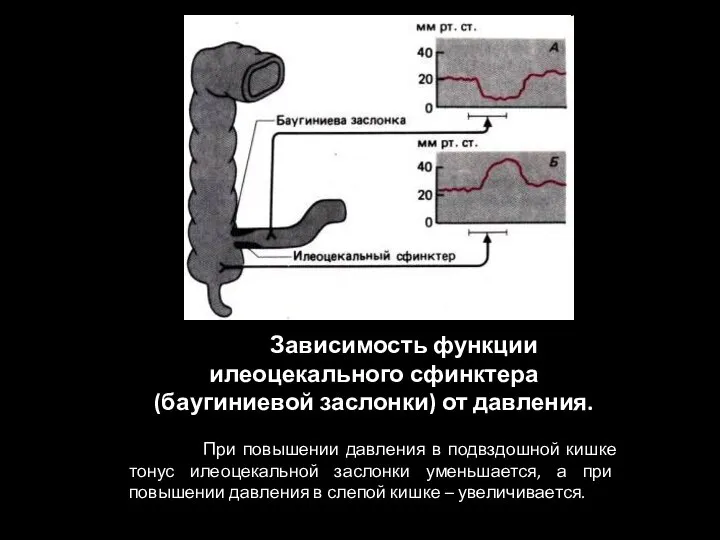
Слайд 102 Зависимость функции илеоцекального сфинктера (баугиниевой заслонки) от давления.
При повышении
Зависимость функции илеоцекального сфинктера (баугиниевой заслонки) от давления.
При повышении
Слайд 103Моторика толстого кишечника
Время пребывания химуса в толстой кишке исчисляется днями, в целом
Моторика толстого кишечника
Время пребывания химуса в толстой кишке исчисляется днями, в целом

Слайд 104Моторика толстого кишечника
Только для толстой кишки свойственны масс - сокращения, которые захватывают
Моторика толстого кишечника
Только для толстой кишки свойственны масс - сокращения, которые захватывают

Слайд 105Моторика толстого кишечника
По сравнению с восходящей ободочной кишкой частота сокращений поперечной и
Моторика толстого кишечника
По сравнению с восходящей ободочной кишкой частота сокращений поперечной и

Слайд 106РЕГУЛЯЦИЯ МОТОРИКИ ТОЛСТОГО КИШЕЧНИКА
Базальный тонус кишечника задается авторитмической активностью гладкомышечных клеток (циркулярный
РЕГУЛЯЦИЯ МОТОРИКИ ТОЛСТОГО КИШЕЧНИКА
Базальный тонус кишечника задается авторитмической активностью гладкомышечных клеток (циркулярный

Слайд 107РЕГУЛЯЦИЯ МОТОРИКИ ТОЛСТОГО КИШЕЧНИКА
Экстрамуральная иннервация осуществляется симпатическим и парасимпатическим отделами нервной системы:
симпатический
РЕГУЛЯЦИЯ МОТОРИКИ ТОЛСТОГО КИШЕЧНИКА
Экстрамуральная иннервация осуществляется симпатическим и парасимпатическим отделами нервной системы:
симпатический

Слайд 108РЕГУЛЯЦИЯ МОТОРИКИ ТОЛСТОГО КИШЕЧНИКА
Существенную роль в регуляции моторики играют рефлекторные связи толстой
РЕГУЛЯЦИЯ МОТОРИКИ ТОЛСТОГО КИШЕЧНИКА
Существенную роль в регуляции моторики играют рефлекторные связи толстой

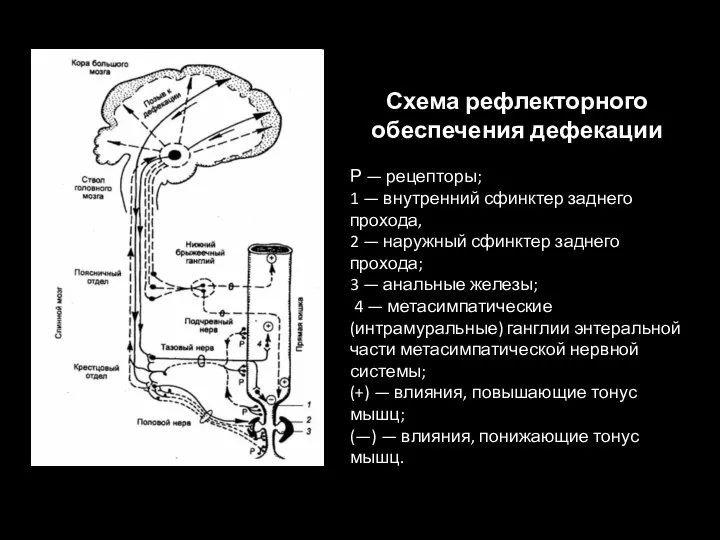
Слайд 109Схема рефлекторного обеспечения дефекации
Р — рецепторы;
1 — внутренний сфинктер заднего прохода,
Схема рефлекторного обеспечения дефекации
Р — рецепторы;
1 — внутренний сфинктер заднего прохода,
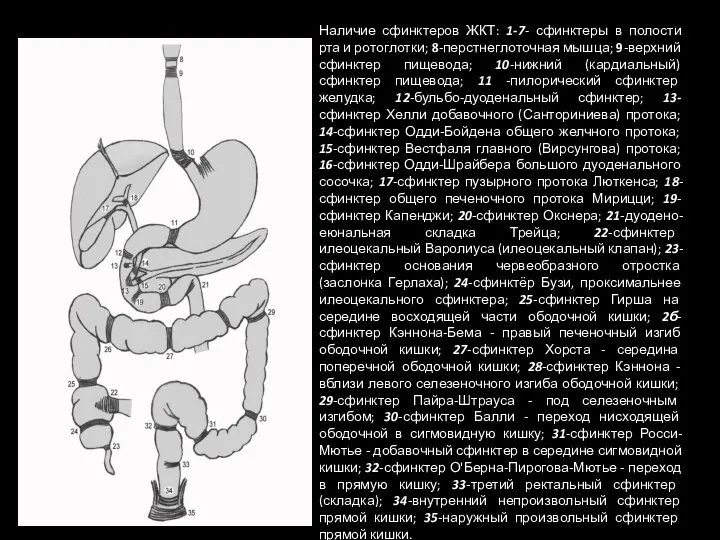
Слайд 110Наличие сфинктеров ЖКТ: 1-7- сфинктеры в полости рта и ротоглотки; 8-перстнеглоточная мышца;
Наличие сфинктеров ЖКТ: 1-7- сфинктеры в полости рта и ротоглотки; 8-перстнеглоточная мышца;
Слайд 111РОЛЬ ИНТЕРСТИЦИАЛЬНЫХ КЛЕТОК КАХАЛЯ В МЕХНАНИЗМЕ ЭЛЕКТРОМЕХАНИЧЕСКОГО РИТМОГЕНЕЗА ЖЕЛУДОЧНО-КИШЕЧНОГО ТРАКТА.
РОЛЬ ИНТЕРСТИЦИАЛЬНЫХ КЛЕТОК КАХАЛЯ В МЕХНАНИЗМЕ ЭЛЕКТРОМЕХАНИЧЕСКОГО РИТМОГЕНЕЗА ЖЕЛУДОЧНО-КИШЕЧНОГО ТРАКТА.

Слайд 112Сантья́го Рамо́н-и-Каха́ль (исп. Santiago Ramón y Cajal) -испанский врач и гистолог,
лауреат
Сантья́го Рамо́н-и-Каха́ль (исп. Santiago Ramón y Cajal) -испанский врач и гистолог, лауреат
Слайд 113Keith A. An account of six specimens of the great bowel removed
Keith A. An account of six specimens of the great bowel removed

Слайд 114Имеется четыре основных популяции ICC в тонком кишечнике мышей.
Ультраструктурные особенности ICC, как
Имеется четыре основных популяции ICC в тонком кишечнике мышей.
Ультраструктурные особенности ICC, как

Слайд 115 Возможности изучения роли ICC были значительно расширены, когда было показано, что в
Возможности изучения роли ICC были значительно расширены, когда было показано, что в

Слайд 116Кроме этого, в настоящее время для исследования роли ICC в регуляции моторики
Кроме этого, в настоящее время для исследования роли ICC в регуляции моторики

Слайд 117СХЕМАТИЧЕСКОЕ ИЗОБРАЖЕНИЕ ТИПОВ ICC, ЛОКАЛИЗОВАННЫХ В РАЗЛИЧНЫХ СЛОЯХ ТКАНИ СТЕНКИ ЖКТ
ICC –
СХЕМАТИЧЕСКОЕ ИЗОБРАЖЕНИЕ ТИПОВ ICC, ЛОКАЛИЗОВАННЫХ В РАЗЛИЧНЫХ СЛОЯХ ТКАНИ СТЕНКИ ЖКТ
ICC –
Слайд 118Классификация ICC.
ICC миэнтерического плексуса (ICC – MP – myenteric plexus). ICC –MP
Классификация ICC.
ICC миэнтерического плексуса (ICC – MP – myenteric plexus). ICC –MP

Слайд 119Сопряжение ICC и ГМК.
Ультрамикроскопические исследования показывают, что отдельные типы ICC имеют плотные
Сопряжение ICC и ГМК.
Ультрамикроскопические исследования показывают, что отдельные типы ICC имеют плотные

Слайд 120ICC – CM в желудке крысы характеризуются электронно-плотной цитоплазмой, каналами (стрелка), щелевым
ICC – CM в желудке крысы характеризуются электронно-плотной цитоплазмой, каналами (стрелка), щелевым
Слайд 121КОМПОНЕНТЫ РИТМОНЕЙРОМЫШЕЧНОГО КОМПЛЕКСА ЖКТ
КОМПОНЕНТЫ РИТМОНЕЙРОМЫШЕЧНОГО КОМПЛЕКСА ЖКТ
Слайд 122Схематическое изображение пейсмекерных потенциалов в ICC – MP медленных волн в ГМК.
Фазы:
Схематическое изображение пейсмекерных потенциалов в ICC – MP медленных волн в ГМК.
Фазы:
Слайд 123Иммуногистохимическое окрашивание сети ICC - зеленая и нервных волокон – красная в
Иммуногистохимическое окрашивание сети ICC - зеленая и нервных волокон – красная в
Слайд 124А и В: Электрические активности, зарегистрированные от циркулярных ГМК тонкого кишечника мышей
А и В: Электрические активности, зарегистрированные от циркулярных ГМК тонкого кишечника мышей
Слайд 125Модель СПР-митохондриального механизма пейсмекерной активности ICC.
Модель СПР-митохондриального механизма пейсмекерной активности ICC.
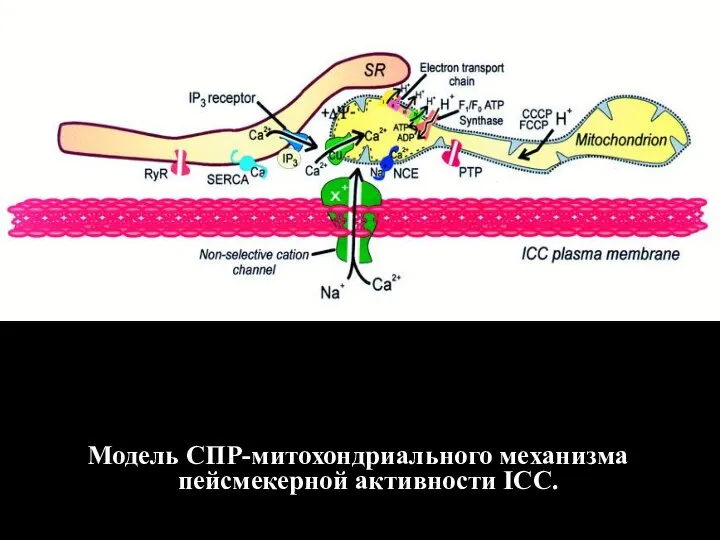
Слайд 126Цитозольный Са2+ генератор, отвечающий за пейсмекерную активность в интерстициальных клетках Кахаля, освобождает
Цитозольный Са2+ генератор, отвечающий за пейсмекерную активность в интерстициальных клетках Кахаля, освобождает
Слайд 127СПАСИБО ЗА ВНИМАНИЕ
СПАСИБО ЗА ВНИМАНИЕ
Слайд 128Всасывание веществ в кишечнике
Всасывание веществ в кишечнике
Слайд 130Механизмы транспорта. Всасывание веществ осуществляется с участием различных видов транспорта.
Пассивный транспорт жидкостей
Механизмы транспорта. Всасывание веществ осуществляется с участием различных видов транспорта.
Пассивный транспорт жидкостей

Слайд 131Активный транспорт веществ является однонаправленным и связан с затратами энергии, источником которой
Активный транспорт веществ является однонаправленным и связан с затратами энергии, источником которой

Слайд 132Облегченная диффузия, как и простая диффузи, осуществляется без затрат энергии, по градиенту
Облегченная диффузия, как и простая диффузи, осуществляется без затрат энергии, по градиенту

Слайд 133Процентные доли разных клеток в теле человека.
(Иллюстрация R. Sender, S. Fuchs
Процентные доли разных клеток в теле человека.
(Иллюстрация R. Sender, S. Fuchs
Слайд 134Работа лизосом
Работа лизосом
Слайд 135Виды эндоцитоза
Фагоцитоз («клеточная еда»):
твердые частицы пищи, например, бактерии оказываются в внутри клетки
Виды эндоцитоза
Фагоцитоз («клеточная еда»):
твердые частицы пищи, например, бактерии оказываются в внутри клетки
 Земноводные Тюменской области
Земноводные Тюменской области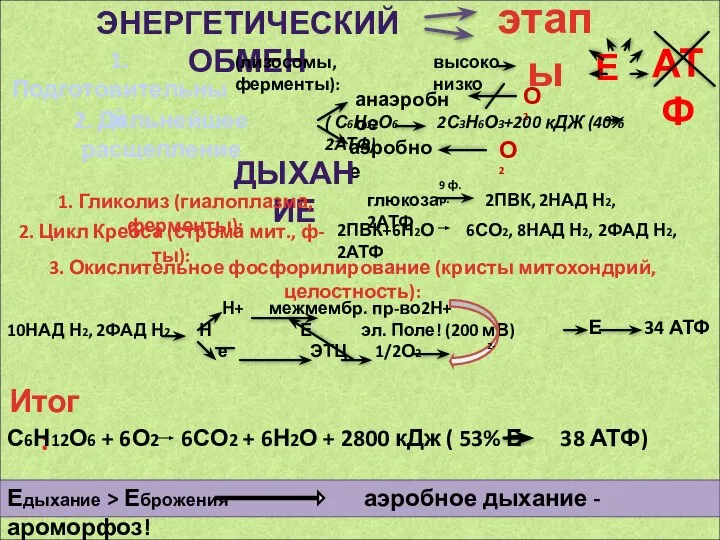 Энергетический обмен
Энергетический обмен Движение крови по сосудам
Движение крови по сосудам Экологические факторы и их влияние на организм
Экологические факторы и их влияние на организм Сравнительная характеристика типов червей
Сравнительная характеристика типов червей Založení neziskové organizace
Založení neziskové organizace Тип Членистоногие
Тип Членистоногие Применения информационных технологий в изучении влияния фитогормонов на рост орхидей
Применения информационных технологий в изучении влияния фитогормонов на рост орхидей Клык нижней челюсти
Клык нижней челюсти Химический состав клетки
Химический состав клетки Жизнь на Земле
Жизнь на Земле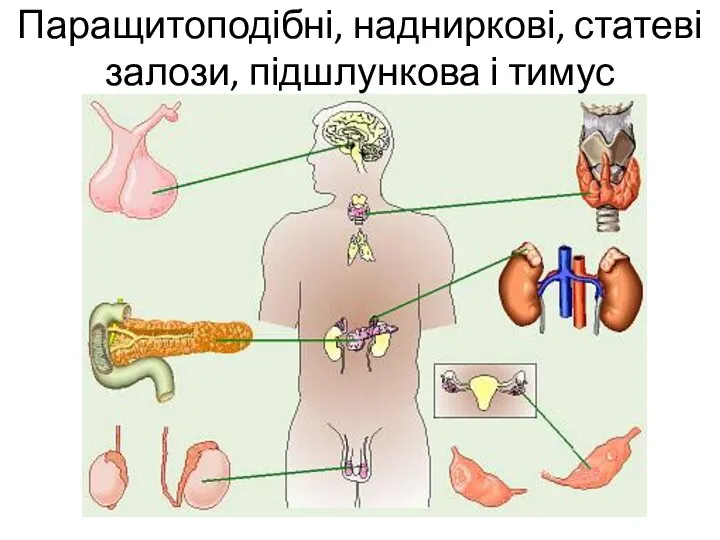 Паращитоподібні, надниркові, статеві залози, підшлункова і тимус
Паращитоподібні, надниркові, статеві залози, підшлункова і тимус 5dc9ea4996b8de47
5dc9ea4996b8de47 Биосинтез белка: Транскрипция. Процессинг мРНК. Трансляция. Фолдинг
Биосинтез белка: Транскрипция. Процессинг мРНК. Трансляция. Фолдинг Ткани человека. 3
Ткани человека. 3 Отличие одного вида растений от другого на основании морфологического критерия
Отличие одного вида растений от другого на основании морфологического критерия Птицы. Внешнее строение
Птицы. Внешнее строение Жүйкелік және гуморальдік реттелудің механизмі
Жүйкелік және гуморальдік реттелудің механизмі Пищеварение полостное, внутриклеточное
Пищеварение полостное, внутриклеточное Изменение живых организмов в зависимости от среды обитания
Изменение живых организмов в зависимости от среды обитания Оценка перспективных селекционных линий ярового ячменя в условиях Западной Сибири
Оценка перспективных селекционных линий ярового ячменя в условиях Западной Сибири Лекарственные растения
Лекарственные растения Фізіологія серця, судин, гемодинаміка
Фізіологія серця, судин, гемодинаміка Занимательная биология
Занимательная биология Формы естественного отбора. Задание
Формы естественного отбора. Задание Назовите признаки жизни. 9 класс
Назовите признаки жизни. 9 класс Презентация на тему Пути достижения биологического прогресса
Презентация на тему Пути достижения биологического прогресса  Закономерности происхождения и развития жизни на Земле. 9 класс
Закономерности происхождения и развития жизни на Земле. 9 класс