- Пластический обмен

Содержание
- 2. Пластический обмен Репликация – удвоение ДНК. Транскрипция – процесс синтеза РНК по матрице ДНК; Трансляция –
- 3. Пластический обмен По другому называется анаболизмом. Всегда проходит с затратой энергии. В процессе фотосинтеза используется физическая
- 4. Аденинтрифосфорная кислота В латинской литерации также обозначается как аденозинтрифосфорная кислота (АТФ). Потеряв одну фосфорную группу называется
- 5. Как работает АТФ АТФ отдает свою фосфатную группу субстрату или ферменту. При этом происходит упругое изменение
- 6. Репликация ДНК - удвоение Происходит в ядре. Расплетение и удвоение ДНК происходит за счет ДНК-полимеразы. Репликация
- 7. Репликация ДНК (копирование) Механизм копирования называется полуконсервативным, так как матрицей новой молекуле ДНК служит одна нить
- 8. Описание репликации Этапы репликации: Синтез комплементарной цепи (ДНК-полимераза); Разрезание водородных связей двух нитей материнской цепочки ДНК
- 9. Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова Три первоначальные гипотезы репликации ДНК Полуконсервативный метод Консервативный метод
- 10. Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова Три первоначальные гипотезы репликации ДНК Консервативный; Полуконсервативный; Дисперсный. Изначальная
- 11. Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
- 12. Репликационная вилка В процессе репликации ферментом хеликазой разрушающей водородные связи между нуклеотидами и разделяющей ДНК на
- 13. Репликационная вилка Особенность ДНК полимераза движется по цепочки ДНК исключительно в одну сторону: от 3’-конца к
- 14. Репликон Реплико́н — молекула или участок ДНК или РНК, реплицирующийся из одной точки начала репликации. Гипотеза
- 15. Монорепликонная репликация У прокариот репликация начинается в одной точки ori* и идет в обе стороны по
- 16. Репликация у доядерных и ядерных форм клеток Прокариоты: циклическая ДНК, репликация начинается с одного конца. ДНК
- 17. Репликационная вилка Схематическое изображение процесса репликации, цифрами отмечены: (1) запаздывающая нить, (2) лидирующая нить, (3) ДНК-полимераза
- 18. Описание репликации Принципиально, чтобы к моменту деления клетки ДНК была реплицирована полностью и при этом только
- 19. Синтез белка включает следующие стадии Транскрипция (по матрице ДНК синтезируется иРНК); Трансляция (по матрице иРНК синтезируется
- 20. Теория гена Информация о строении белка содержится в гене. Ген – это структурная единица ДНК. Ген
- 21. Отклонения Первый пример отклонения от стандартного генетического кода был открыт в 1979 году при исследовании генов
- 22. Транскрипция Транскрипция выполняется с помощью РНК-полимеразы; РНК-полимераза двигается с 3’ к 5’ концу по кодирующей цепочки
- 23. ТРАНСКРИЦИЯ Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
- 24. Посттранскрипционные механизмы После завершения трансляции РНК может претерпевать изменения. РНК скомпонованное из транскриптов с нескольких экзонов
- 25. Механизм транскрипции у бактерий
- 26. СТРОЕНИЕ ГЕНОВ У ПРОКАРИОТ Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
- 27. Механизм транскрипции у бактерий Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
- 28. https://en.ppt-online.org/347103 Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
- 29. Механизм транскрипции у эукариот Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова Poly-A или Полиаденилирование — это
- 30. Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
- 31. В чем сходства и отличия транскрипции бактерий и эукариот? Сходства: 1) Синтез начинается с АУГ-кодона, кодирующего
- 32. РНК в цитоплазме Чтобы экзонуклеазы не разрушили РНК, она закрывается от них с 3’-конца cap-белком (место
- 33. Трансляция Трансляция идет с 5’ к 3’концу. В цитоплазме иРНК (информационная) соединяется с рибосомой (рибосомальная РНК
- 34. Стадии трансляции Инициация Связывание иРНК с малой субъединицей рибосомы. Присоединение к м. суб. первой тРНК Связывание
- 36. Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова 1 – малая субъединица рибосомы 2 – иРНК 3
- 38. Модель генетического кода Открыт Маршалом Ниренбенргом и С Очао в 1961 Получили Нобелевскую премию 1968 г.
- 39. Свойства генетического кода Триплетен - каждая аминокислота кодируется тремя нуклеотидами. Эта последовательность в молекуле иРНК называется
- 40. Процессы играет важную роль в Репликации – матричном синтезе новой цепочки ДНК по старой; Транскрипции –
- 41. Таблица генетического кода Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
- 43. Синтез аминоацил-тРНК Конечный продукт активации аминокислот: молекула тРНК, ковалентно связанная с аминокислотой через ацильную связь между
- 44. Рамка считывания последовательность нуклеотидов в составе ДНК или РНК, потенциально способная кодировать белок. Нарушение рамки считывания
- 45. Синонимы – омонимы генетического кода Определение соответствия кодону и аминокислоте наглядно продемонстрировано в таблице генетического кода
- 46. Модель Маршала Ниренбенрга и С Очао 1961
- 47. Мутации Генные - вставки, выпадения, замена нуклеотидов, триплетов, части гена; Ограничены размером гена; Хромосомные – делеция
- 48. Генные мутации. Причина - следствие Замена одного нуклеотида связана с изменением аминокислотного состава белка. Не изменится:
- 49. Хемосинтез Хемосинтез — способ автотрофного питания, при котором источником энергии для синтеза органических веществ из CO2
- 50. Хемосинтез Железобактерии (Geobacter, Gallionella) окисляют двухвалентное железо до трёхвалентного. Серобактерии (Desulfuromonas, Desulfobacter, Beggiatoa) окисляют сероводород до
- 51. Регуляция экспрессии генов https://biology.su/molecular/gene-activity https://studopedia.ru/5_169574_regulyatsiya-ekspressii-genov-u-prokariot.html Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
- 52. ДНК - аддукт https://yandex.ru/search/?clid=2332287&win=421&from=chromesearch&text=%D0%B0%D0%B4%D0%B4%D1%83%D0%BA%D1%82%D1%8B%20%D0%B4%D0%BD%D0%BA&lr=213 Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
- 54. Скачать презентацию
Слайд 2Пластический обмен
Репликация – удвоение ДНК.
Транскрипция – процесс синтеза РНК по матрице ДНК;
Трансляция
Пластический обмен
Репликация – удвоение ДНК.
Транскрипция – процесс синтеза РНК по матрице ДНК;
Трансляция

Слайд 3Пластический обмен
По другому называется анаболизмом. Всегда проходит с затратой энергии.
В процессе
Пластический обмен
По другому называется анаболизмом. Всегда проходит с затратой энергии.
В процессе

Слайд 4Аденинтрифосфорная кислота
В латинской литерации также обозначается как аденозинтрифосфорная кислота (АТФ).
Потеряв одну фосфорную
Аденинтрифосфорная кислота
В латинской литерации также обозначается как аденозинтрифосфорная кислота (АТФ).
Потеряв одну фосфорную

Слайд 5Как работает АТФ
АТФ отдает свою фосфатную группу субстрату или ферменту. При этом
Как работает АТФ
АТФ отдает свою фосфатную группу субстрату или ферменту. При этом

Слайд 6Репликация ДНК - удвоение
Происходит в ядре.
Расплетение и удвоение ДНК происходит за счет
Репликация ДНК - удвоение
Происходит в ядре.
Расплетение и удвоение ДНК происходит за счет

Слайд 7Репликация ДНК (копирование)
Механизм копирования называется полуконсервативным, так как матрицей новой молекуле ДНК
Репликация ДНК (копирование)
Механизм копирования называется полуконсервативным, так как матрицей новой молекуле ДНК

Слайд 8Описание репликации
Этапы репликации:
Синтез комплементарной цепи (ДНК-полимераза);
Разрезание водородных связей двух нитей
Описание репликации
Этапы репликации:
Синтез комплементарной цепи (ДНК-полимераза);
Разрезание водородных связей двух нитей

Слайд 9Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
Три первоначальные гипотезы репликации ДНК
Полуконсервативный
Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
Три первоначальные гипотезы репликации ДНК
Полуконсервативный

Слайд 10Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
Три первоначальные гипотезы репликации ДНК
Консервативный;
Полуконсервативный;
Дисперсный.
Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
Три первоначальные гипотезы репликации ДНК
Консервативный;
Полуконсервативный;
Дисперсный.

Слайд 11Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
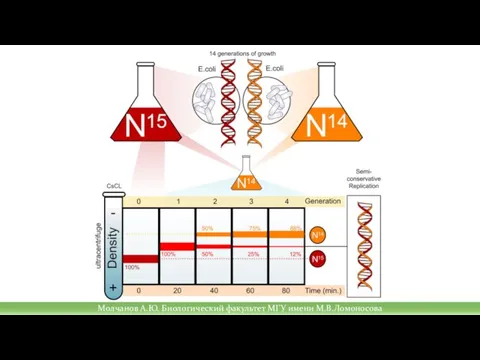
Слайд 12Репликационная вилка
В процессе репликации ферментом хеликазой разрушающей водородные связи между нуклеотидами и
Репликационная вилка
В процессе репликации ферментом хеликазой разрушающей водородные связи между нуклеотидами и

Слайд 13Репликационная вилка
Особенность ДНК полимераза движется по цепочки ДНК исключительно в одну сторону:
Репликационная вилка
Особенность ДНК полимераза движется по цепочки ДНК исключительно в одну сторону:

Слайд 14Репликон
Реплико́н — молекула или участок ДНК или РНК, реплицирующийся из одной точки начала
Репликон
Реплико́н — молекула или участок ДНК или РНК, реплицирующийся из одной точки начала

Слайд 15Монорепликонная репликация
У прокариот репликация начинается в одной точки ori* и идет в
Монорепликонная репликация
У прокариот репликация начинается в одной точки ori* и идет в

Слайд 16Репликация у доядерных и ядерных форм клеток
Прокариоты: циклическая ДНК, репликация начинается с
Репликация у доядерных и ядерных форм клеток
Прокариоты: циклическая ДНК, репликация начинается с

Слайд 17Репликационная вилка
Схематическое изображение процесса репликации, цифрами отмечены: (1) запаздывающая нить, (2) лидирующая
Репликационная вилка
Схематическое изображение процесса репликации, цифрами отмечены: (1) запаздывающая нить, (2) лидирующая

Слайд 18Описание репликации
Принципиально, чтобы к моменту деления клетки ДНК была реплицирована полностью
Описание репликации
Принципиально, чтобы к моменту деления клетки ДНК была реплицирована полностью

Слайд 19Синтез белка включает следующие стадии
Транскрипция (по матрице ДНК синтезируется иРНК);
Трансляция (по матрице
Синтез белка включает следующие стадии
Транскрипция (по матрице ДНК синтезируется иРНК);
Трансляция (по матрице

Слайд 20Теория гена
Информация о строении белка содержится в гене. Ген – это структурная
Теория гена
Информация о строении белка содержится в гене. Ген – это структурная

Слайд 21Отклонения
Первый пример отклонения от стандартного генетического кода был открыт в 1979 году при исследовании
Отклонения
Первый пример отклонения от стандартного генетического кода был открыт в 1979 году при исследовании

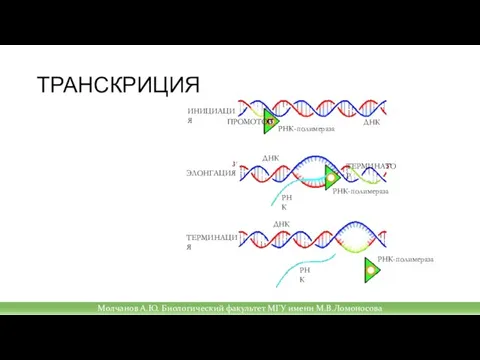
Слайд 22Транскрипция
Транскрипция выполняется с помощью РНК-полимеразы;
РНК-полимераза двигается с 3’ к 5’ концу по
Транскрипция
Транскрипция выполняется с помощью РНК-полимеразы;
РНК-полимераза двигается с 3’ к 5’ концу по

Слайд 23ТРАНСКРИЦИЯ
Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
ТРАНСКРИЦИЯ
Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
Слайд 24Посттранскрипционные механизмы
После завершения трансляции РНК может претерпевать изменения.
РНК скомпонованное из транскриптов
Посттранскрипционные механизмы
После завершения трансляции РНК может претерпевать изменения.
РНК скомпонованное из транскриптов

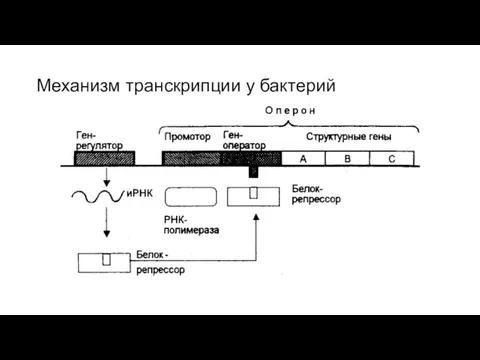
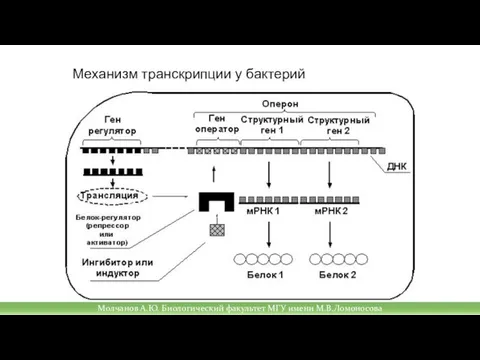
Слайд 25Механизм транскрипции у бактерий
Механизм транскрипции у бактерий
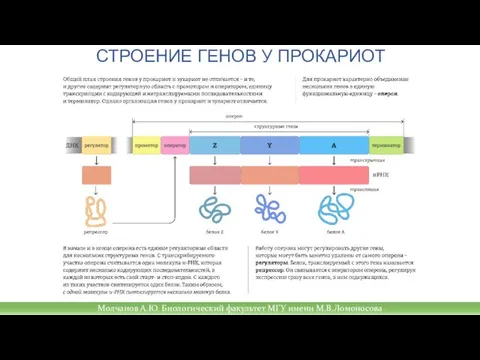
Слайд 26СТРОЕНИЕ ГЕНОВ У ПРОКАРИОТ
Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
СТРОЕНИЕ ГЕНОВ У ПРОКАРИОТ
Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
Слайд 27Механизм транскрипции у бактерий
Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
Механизм транскрипции у бактерий
Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
Слайд 28https://en.ppt-online.org/347103
Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
https://en.ppt-online.org/347103
Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова

Слайд 29Механизм транскрипции у эукариот
Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
Poly-A или
Механизм транскрипции у эукариот
Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
Poly-A или

Слайд 30Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова

Слайд 31В чем сходства и отличия транскрипции бактерий и эукариот?
Сходства:
1) Синтез начинается с
В чем сходства и отличия транскрипции бактерий и эукариот?
Сходства:
1) Синтез начинается с

Слайд 32РНК в цитоплазме
Чтобы экзонуклеазы не разрушили РНК, она закрывается от них с
РНК в цитоплазме
Чтобы экзонуклеазы не разрушили РНК, она закрывается от них с

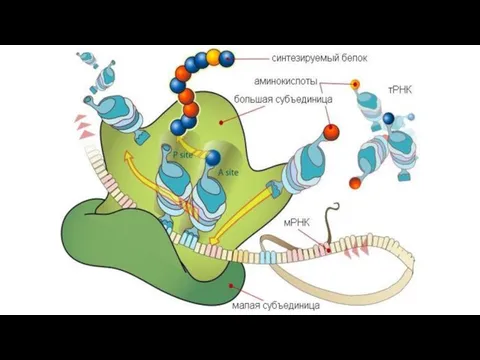
Слайд 33Трансляция
Трансляция идет с 5’ к 3’концу.
В цитоплазме иРНК (информационная) соединяется с рибосомой
Трансляция
Трансляция идет с 5’ к 3’концу.
В цитоплазме иРНК (информационная) соединяется с рибосомой

Слайд 34Стадии трансляции
Инициация
Связывание иРНК с малой субъединицей рибосомы.
Присоединение к м. суб. первой
Стадии трансляции
Инициация
Связывание иРНК с малой субъединицей рибосомы.
Присоединение к м. суб. первой


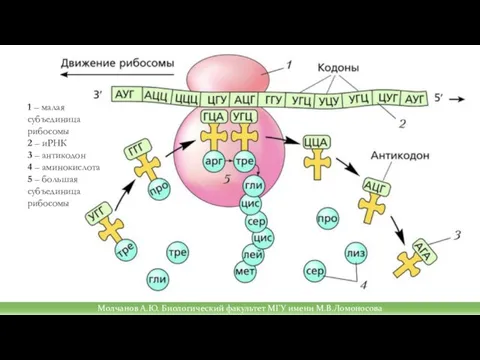
Слайд 36Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
1 – малая субъединица рибосомы
2
Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
1 – малая субъединица рибосомы
2
Слайд 38Модель генетического кода
Открыт Маршалом Ниренбенргом и С Очао в 1961
Получили Нобелевскую премию
Модель генетического кода
Открыт Маршалом Ниренбенргом и С Очао в 1961
Получили Нобелевскую премию

Слайд 39Свойства генетического кода
Триплетен - каждая аминокислота кодируется тремя нуклеотидами. Эта последовательность в
Свойства генетического кода
Триплетен - каждая аминокислота кодируется тремя нуклеотидами. Эта последовательность в

Слайд 40Процессы играет важную роль в
Репликации – матричном синтезе новой цепочки ДНК
Процессы играет важную роль в
Репликации – матричном синтезе новой цепочки ДНК

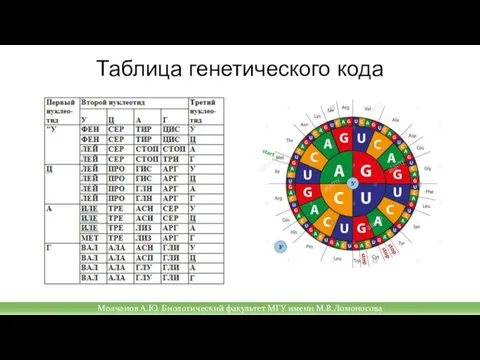
Слайд 41Таблица генетического кода
Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
Таблица генетического кода
Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова

Слайд 43Синтез аминоацил-тРНК
Конечный продукт активации аминокислот: молекула тРНК, ковалентно связанная с аминокислотой через
Синтез аминоацил-тРНК
Конечный продукт активации аминокислот: молекула тРНК, ковалентно связанная с аминокислотой через

Слайд 44Рамка считывания
последовательность нуклеотидов в составе ДНК или РНК, потенциально способная кодировать белок.
Рамка считывания
последовательность нуклеотидов в составе ДНК или РНК, потенциально способная кодировать белок.

Слайд 45Синонимы – омонимы генетического кода
Определение соответствия кодону и аминокислоте наглядно продемонстрировано
Синонимы – омонимы генетического кода
Определение соответствия кодону и аминокислоте наглядно продемонстрировано

Слайд 46Модель Маршала Ниренбенрга и С Очао 1961
Модель Маршала Ниренбенрга и С Очао 1961

Слайд 47Мутации
Генные - вставки, выпадения, замена нуклеотидов, триплетов, части гена; Ограничены размером гена;
Хромосомные
Мутации
Генные - вставки, выпадения, замена нуклеотидов, триплетов, части гена; Ограничены размером гена;
Хромосомные

Слайд 48Генные мутации. Причина - следствие
Замена одного нуклеотида связана с изменением аминокислотного состава
Генные мутации. Причина - следствие
Замена одного нуклеотида связана с изменением аминокислотного состава

Слайд 49Хемосинтез
Хемосинтез — способ автотрофного питания, при котором источником энергии для синтеза органических
Хемосинтез
Хемосинтез — способ автотрофного питания, при котором источником энергии для синтеза органических

Слайд 50Хемосинтез
Железобактерии (Geobacter, Gallionella) окисляют двухвалентное железо до трёхвалентного.
Серобактерии (Desulfuromonas, Desulfobacter, Beggiatoa) окисляют
Хемосинтез
Железобактерии (Geobacter, Gallionella) окисляют двухвалентное железо до трёхвалентного.
Серобактерии (Desulfuromonas, Desulfobacter, Beggiatoa) окисляют

Слайд 51Регуляция экспрессии генов
https://biology.su/molecular/gene-activity
https://studopedia.ru/5_169574_regulyatsiya-ekspressii-genov-u-prokariot.html
Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
Регуляция экспрессии генов
https://biology.su/molecular/gene-activity
https://studopedia.ru/5_169574_regulyatsiya-ekspressii-genov-u-prokariot.html
Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова

Слайд 52ДНК - аддукт
https://yandex.ru/search/?clid=2332287&win=421&from=chromesearch&text=%D0%B0%D0%B4%D0%B4%D1%83%D0%BA%D1%82%D1%8B%20%D0%B4%D0%BD%D0%BA&lr=213
Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова
ДНК - аддукт
https://yandex.ru/search/?clid=2332287&win=421&from=chromesearch&text=%D0%B0%D0%B4%D0%B4%D1%83%D0%BA%D1%82%D1%8B%20%D0%B4%D0%BD%D0%BA&lr=213
Молчанов А.Ю. Биологический факультет МГУ имени М.В.Ломоносова

 Презентация на тему СОСНОВЫЙ ЛЕС
Презентация на тему СОСНОВЫЙ ЛЕС  Формы размножения организмов и их цитологические основы
Формы размножения организмов и их цитологические основы Презентация на тему ПИЩЕВАЯ БИОТЕХНОЛОГИЯ
Презентация на тему ПИЩЕВАЯ БИОТЕХНОЛОГИЯ  Образование шейного сплетения
Образование шейного сплетения Факторы эволюции человека
Факторы эволюции человека Животные жарких стран и Севера
Животные жарких стран и Севера Тыңлагыз. Кошлар- безнең
Тыңлагыз. Кошлар- безнең Листья деревьев и кустарников
Листья деревьев и кустарников Пищеварительная система
Пищеварительная система Царство: Растения. Отдел: Покрытосеменные. Класс: Однодольные
Царство: Растения. Отдел: Покрытосеменные. Класс: Однодольные Рыбы. Внутренний костный скелет
Рыбы. Внутренний костный скелет Lektsia_14-15_golovnoy_Mozg
Lektsia_14-15_golovnoy_Mozg Патологическая физиология типовых нарушений обмена веществ
Патологическая физиология типовых нарушений обмена веществ Презентация на тему Многообразие насекомых, роль в природе и на практике
Презентация на тему Многообразие насекомых, роль в природе и на практике  Подсолнухи
Подсолнухи Функции и строение нервной системы
Функции и строение нервной системы Who wants to be a millionaire
Who wants to be a millionaire Весна: березень, квітень, травень
Весна: березень, квітень, травень Витамин В6
Витамин В6 Многообразие и значение птиц
Многообразие и значение птиц Арзамасский дендрарий
Арзамасский дендрарий В царстве растений
В царстве растений Комнатные растения
Комнатные растения Основные группы микроорганизмов
Основные группы микроорганизмов Морские уточки
Морские уточки Плоские, круглые, кольчатые черви
Плоские, круглые, кольчатые черви What makes our blood different from others
What makes our blood different from others Вегетативные органы растений
Вегетативные органы растений