- Простые и сложные белки. Уровни структурной организации белка. Тема 4

Содержание
- 2. Белки́ (протеи́ны, полипепти́ды) (англ. Proteins) высокомолекулярные органические вещества, состоящие из альфа-аминокислот, соединённых в цепочку пептидной связью.
- 3. Образование полипептидов и биосинтез белка В живых организмах полипептидные (белковые) цепи синтезируются специальными молекулярными комплексами –
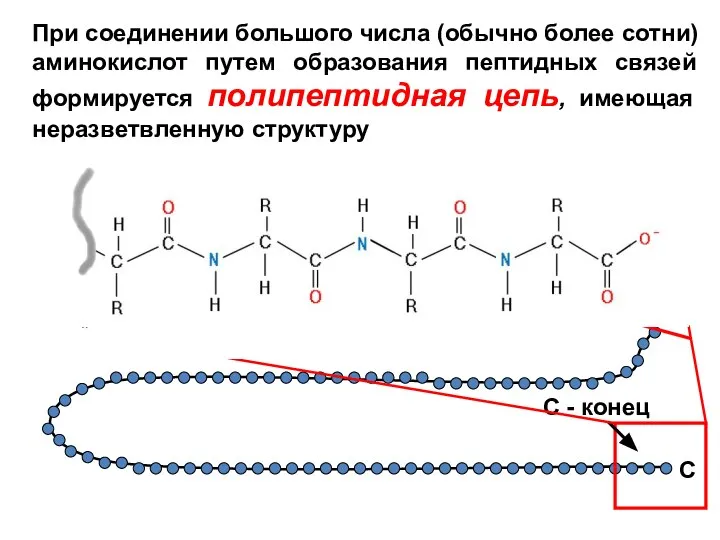
- 4. При соединении большого числа (обычно более сотни) аминокислот путем образования пептидных связей формируется полипептидная цепь, имеющая
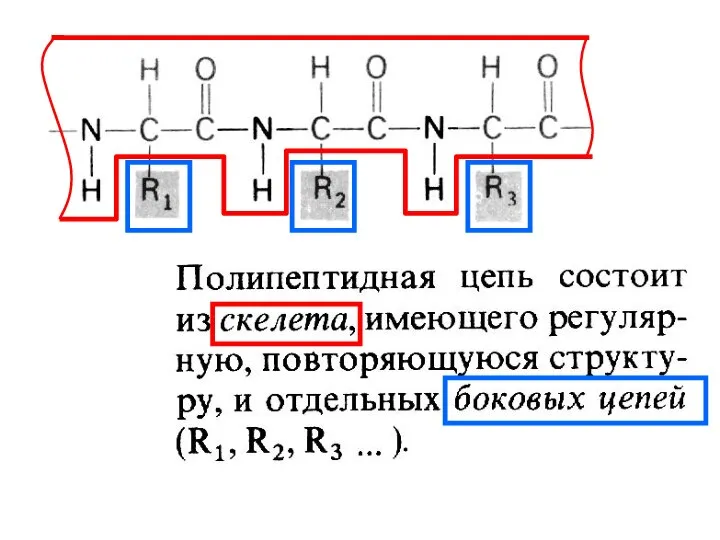
- 6. 1. Белки состоят из одной или нескольких полипептидных цепей 2. Последовательность аминокислот в полипептидных цепях белка
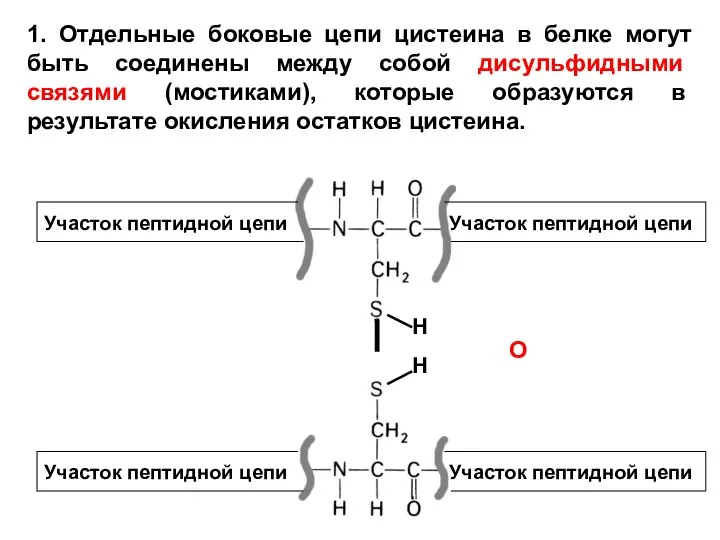
- 7. 1. Отдельные боковые цепи цистеина в белке могут быть соединены между собой дисульфидными связями (мостиками), которые
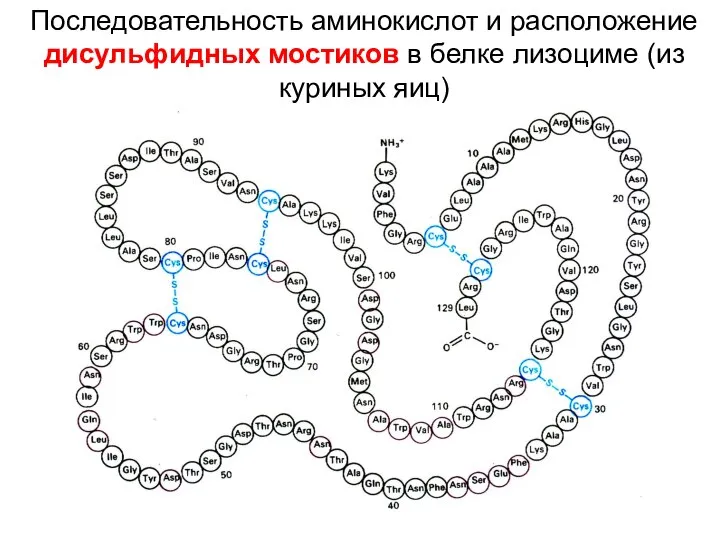
- 8. Последовательность аминокислот и расположение дисульфидных мостиков в белке лизоциме (из куриных яиц)
- 9. Ген – это участок ДНК, где зашифрована информация о последовательности аминокислот в каком-либо белке
- 10. 3. После синтеза белка некоторые аминокислотные остатки могут подвергаться специфической посттрансляционной модификации – к ним присоединяются
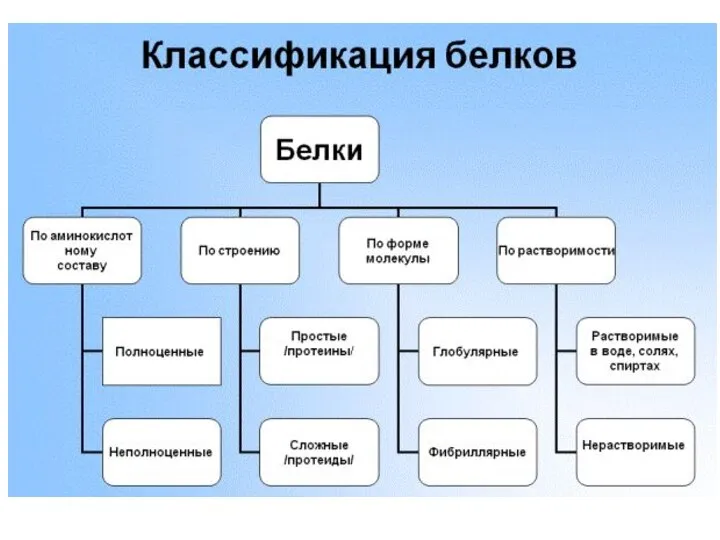
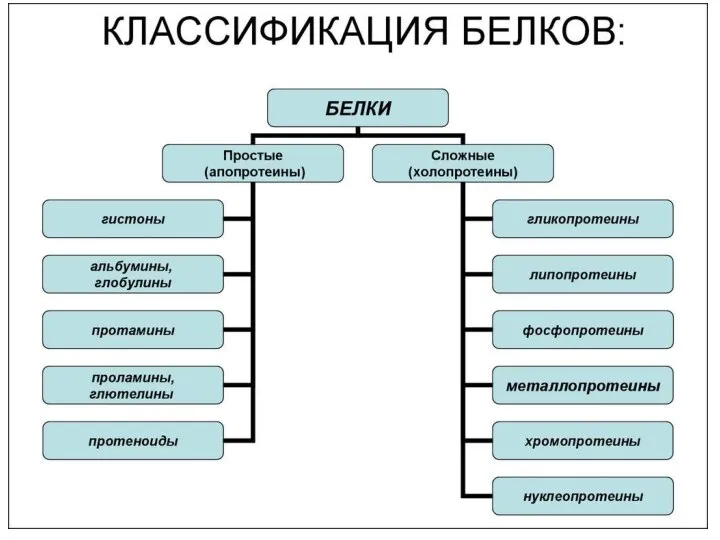
- 11. Простые и сложные белки Простые белки содержат только аминокислотные остатки Сложные белки содержат еще и неаминокислотные
- 12. Антитело IgG Гемоглобин Инсулин Аденилатциклаза Глютаминсинтетаза
- 13. Все белки обязательно имеют две характеристики: Молекулярная масса Изоэлектрическая точка А́томная едини́ца ма́ссы (русское обозначение: а.
- 15. Молекулярная масса и количество аминокислот в некоторых белках
- 16. Изоэлектрическая точка некоторых белков
- 18. Пищевая ценность белков определяется способностью перевариваться в ЖКТ и содержанием незаменимых аминокислот Например, белки яиц, молока,
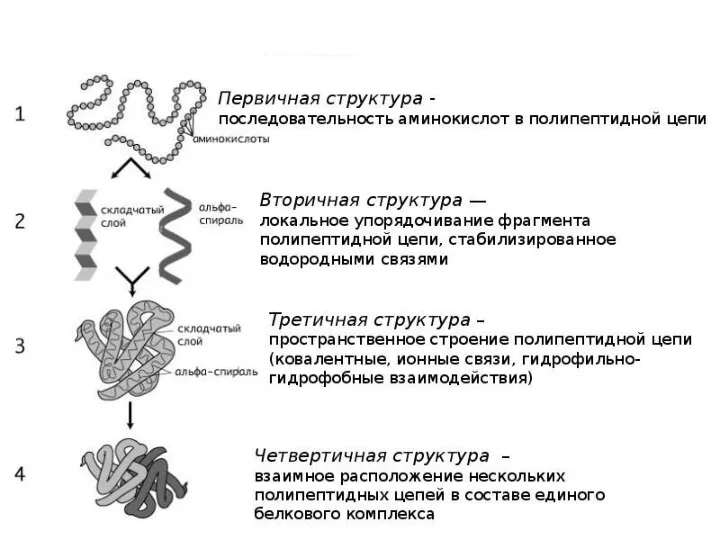
- 21. Уровни структуры белка Первичная структура – последовательность аминокислотных остатков в полипептидных цепях. Первичная структура полностью описывает
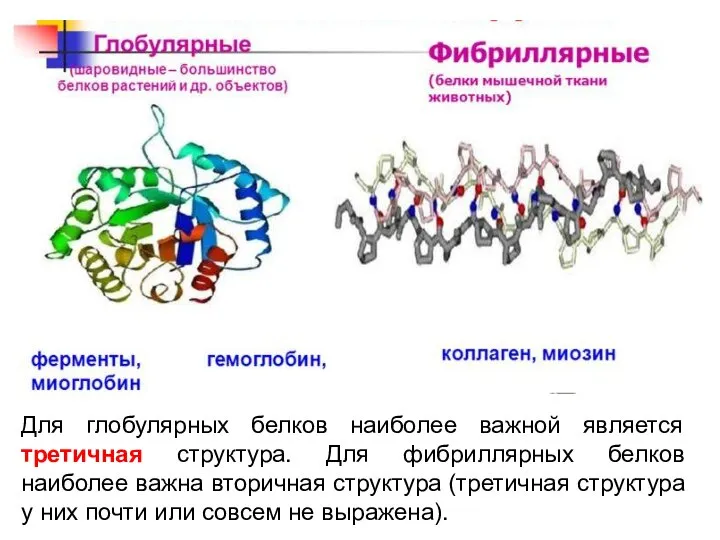
- 28. Для глобулярных белков наиболее важной является третичная структура. Для фибриллярных белков наиболее важна вторичная структура (третичная
- 29. Вторичная структура
- 30. Пептидная связь N H C R1 H C O N H R2 C H C O
- 31. Вращение невозможно Вращение возможно Вращение возможно
- 32. Типы регулярных вторичных структур α-спираль β-слой ! В белках обычно встречается правая альфа-спираль.
- 33. Свойства α-спирали Шаг правой α-спирали – 0,54 нанометра (3,6 аминокислотных остатка). Шаг левой α-спирали 0,96 нанометра
- 38. Типы β-слоев 1. Параллельный (соседние полипептидные цепи идут в одном направлении) 2. Антипараллельный (соседние цепи идут
- 39. Параллельный β-слой Анти-параллельный β-слой
- 41. В типичном белке около 60% аминокислотных остатков принимает участие в формировании регулярных вторичных структур.
- 42. 1. Третичная структура белка – общая укладка полипептидной цепи. 2. Третичная структура каждого белка уникальна. Третичная
- 43. Взаимодействия, формирующие третичную структуру белка Гидрофобные Гидрофобные взаимодействия связаны с тем, что гидрофобные (неполярные и незаряженные)
- 44. Неполярные группы Гидрофобные взаимодействия H2O H2O
- 45. Гидрофобные аминокислоты - неполярные и незаряженные Алифатические Ароматические Аланин Валин Лейцин Изолейцин Фенилаланин Триптофан Для тирозина:
- 46. На поверхности белка располагаются аминокислотные остатки с ионогенными боковыми цепями, которые могут нести заряд. Будут ли
- 47. Ионные мостики
- 48. Строение типичного глобулярного водорастворимого белка Вода Гидрофобное ядро Гидрофильная поверхность Гидрофобные островки
- 49. Процесс формирования трехмерной структуры белка – сворачивание (фолдинг) белка. Фолдинг белка происходит, в нормальных условиях, самопроизвольно
- 50. Схематичное представление укладки белка в третичную структуру
- 52. Отдельные аминокислотные остатки, удаленные в аминокислотной последовательности полипептида, сближены в третичной структуре белка. Так образуются активные
- 55. Домен белка — элемент третичной структуры белка, представляющий собой достаточно стабильную и независимую подструктуру белка, фолдинг
- 56. "Лейциновая застёжка-молния" между α-спиральными участками двух белков. Н2О Н2О Н2О
- 57. Локализация гидрофобных и гидрофильных радикалов аминокислотных остатков в молекуле белка. А - гидрофильный цитоплазматический белок; Б
- 62. !!! Для сложных белков (которые содержат не только аминокислоты, но и неаминокислотные компоненты) расположение неаминокислотных компонентов
- 63. Четвертичная структура Если белки состоят из двух и более полипептидных цепей, связанных между собой нековалентными связями,
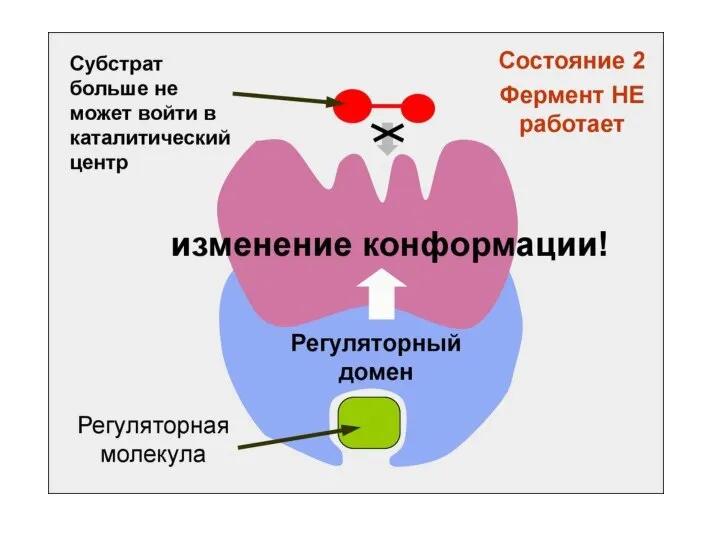
- 67. Многие ферменты имеют две каталитические субъединицы, которые согласованно меняют свою конформацию. В других случаях ферменты имеют
- 68. Сложная третичная и четвертичная структура белков необходима не только для выполнения ими их непосредственной функции, но
- 70. Денатурация белка Денатурация белка – нарушение вторичной и/или уникальной третичной структуры белка, сопровождающееся изменением его физико-химических
- 72. Денатурирующие агенты и воздействия: 1. Высокая температура 2. Сильные минеральные кислоты и основания. Разрушают ионные мостики
- 78. Денатурация белка детергентами Денатурация белка восстанавливающими и хаотропными агентами
- 80. Ренатурация белка Если белок при денатурации остался в растворе, он может быть ренатурирован при возвращении условий
- 82. Посттрансляционная модификация белков
- 84. 1. Гидроксилирование Гидроксилированием называют введение в молекулу органического соединения гидроксильной группы. 1) Гидроксилирование фенилаланина приводит к
- 85. 2. Гликозилирование
- 86. 3. Фосфорилирование Фосфотирозин
- 87. 4. Метилирование
- 88. 5. Сульфатирование
- 89. 6. Добавление гидрофильных групп
- 90. 7. Ацетилирование
- 91. Катаболизм белков
- 92. 1. В нейтральной среде полипептидная цепь является химически стабильной. В сильнокислой среде и при нагревании, полипептидная
- 93. Ферменты, осуществляющие деградацию белков, называются протеазами. По месту атаки молекулы субстрата протеолитические ферменты делятся на: 1.
- 98. Скачать презентацию
Слайд 2Белки́ (протеи́ны, полипепти́ды) (англ. Proteins) высокомолекулярные органические вещества, состоящие из альфа-аминокислот, соединённых в цепочку пептидной связью.
Белки
Белки́ (протеи́ны, полипепти́ды) (англ. Proteins) высокомолекулярные органические вещества, состоящие из альфа-аминокислот, соединённых в цепочку пептидной связью.
Белки

Слайд 3Образование полипептидов и биосинтез белка
В живых организмах полипептидные (белковые) цепи синтезируются специальными
Образование полипептидов и биосинтез белка
В живых организмах полипептидные (белковые) цепи синтезируются специальными

Слайд 4При соединении большого числа (обычно более сотни) аминокислот путем образования пептидных связей
При соединении большого числа (обычно более сотни) аминокислот путем образования пептидных связей
Слайд 61. Белки состоят из одной или нескольких полипептидных цепей
2. Последовательность аминокислот
1. Белки состоят из одной или нескольких полипептидных цепей
2. Последовательность аминокислот

Слайд 71. Отдельные боковые цепи цистеина в белке могут быть соединены между собой
1. Отдельные боковые цепи цистеина в белке могут быть соединены между собой
Слайд 8Последовательность аминокислот и расположение дисульфидных мостиков в белке лизоциме (из куриных яиц)
Последовательность аминокислот и расположение дисульфидных мостиков в белке лизоциме (из куриных яиц)
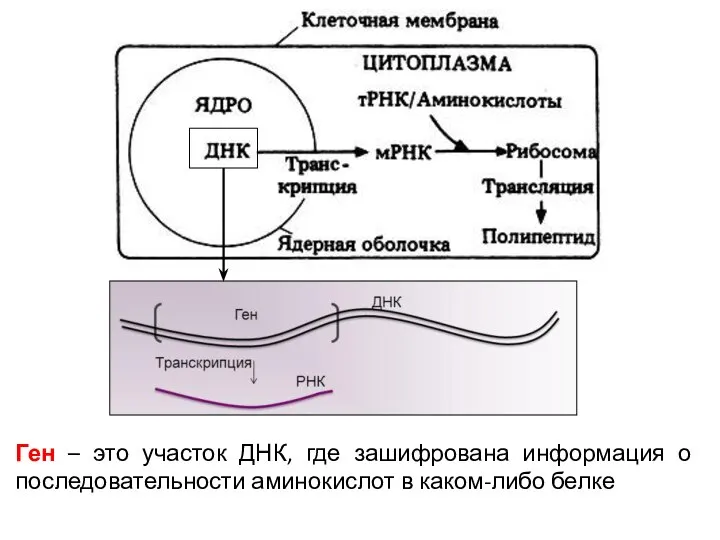
Слайд 9Ген – это участок ДНК, где зашифрована информация о последовательности аминокислот в
Ген – это участок ДНК, где зашифрована информация о последовательности аминокислот в
Слайд 103. После синтеза белка некоторые аминокислотные остатки могут подвергаться специфической посттрансляционной модификации
3. После синтеза белка некоторые аминокислотные остатки могут подвергаться специфической посттрансляционной модификации

Слайд 11Простые и сложные белки
Простые белки содержат только аминокислотные остатки
Сложные белки содержат еще
Простые и сложные белки
Простые белки содержат только аминокислотные остатки
Сложные белки содержат еще

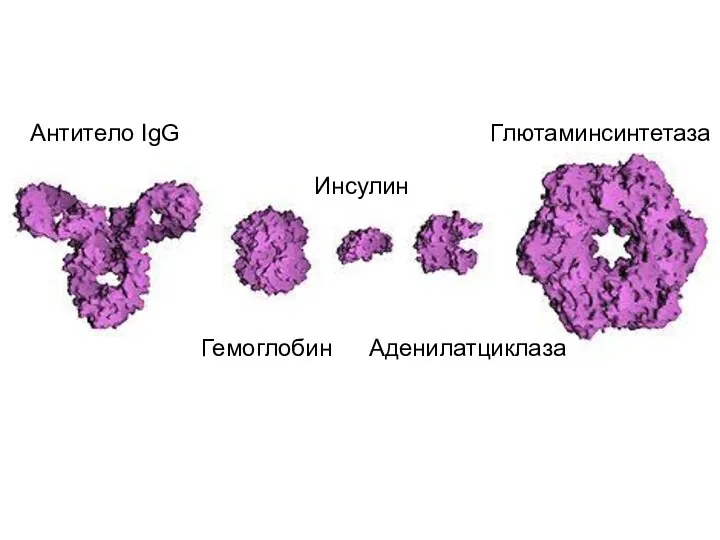
Слайд 12Антитело IgG
Гемоглобин
Инсулин
Аденилатциклаза
Глютаминсинтетаза
Антитело IgG
Гемоглобин
Инсулин
Аденилатциклаза
Глютаминсинтетаза
Слайд 13Все белки обязательно имеют две характеристики:
Молекулярная масса
Изоэлектрическая точка
А́томная едини́ца
Все белки обязательно имеют две характеристики:
Молекулярная масса
Изоэлектрическая точка
А́томная едини́ца

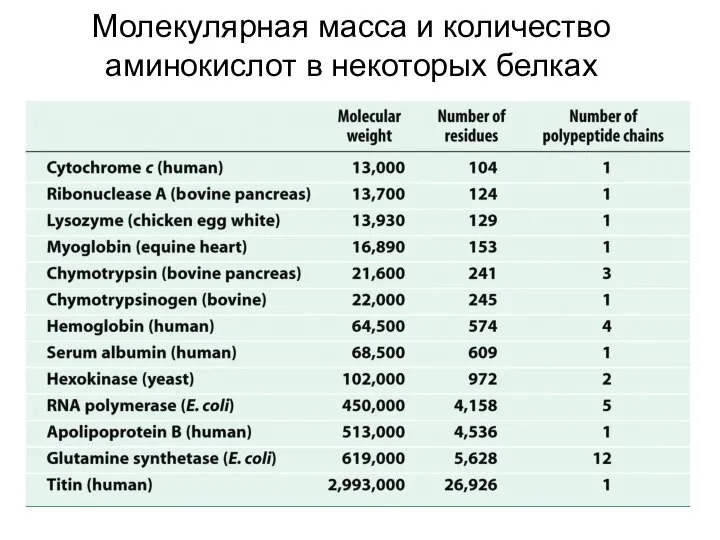
Слайд 15Молекулярная масса и количество аминокислот в некоторых белках
Молекулярная масса и количество аминокислот в некоторых белках
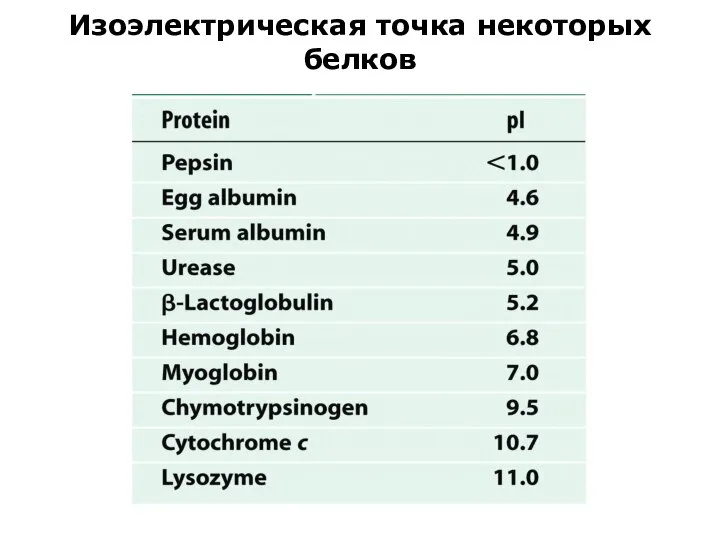
Слайд 16Изоэлектрическая точка некоторых белков
Изоэлектрическая точка некоторых белков
Слайд 18Пищевая ценность белков определяется способностью перевариваться в ЖКТ и содержанием незаменимых аминокислот
Например,
Пищевая ценность белков определяется способностью перевариваться в ЖКТ и содержанием незаменимых аминокислот
Например,


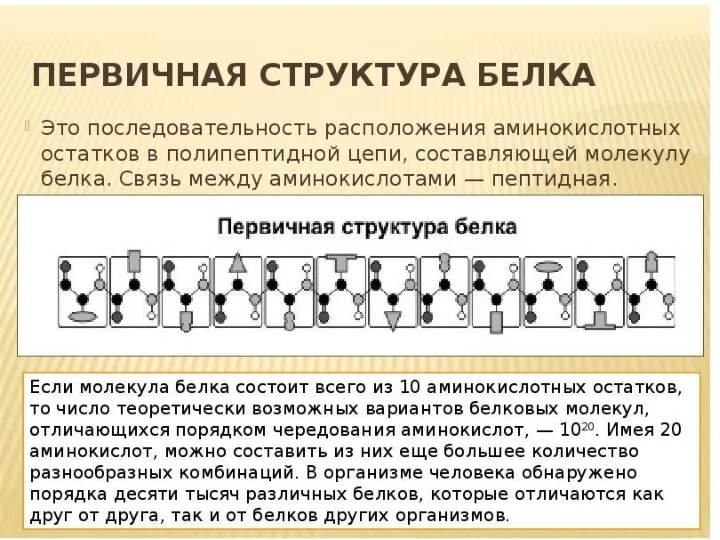
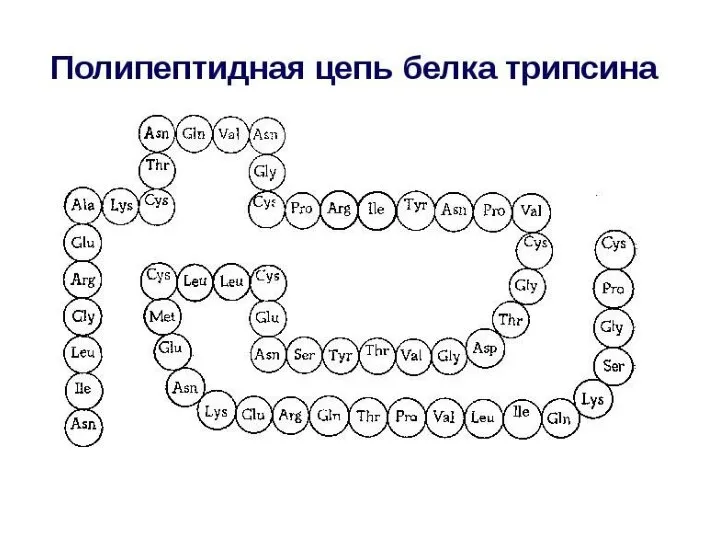
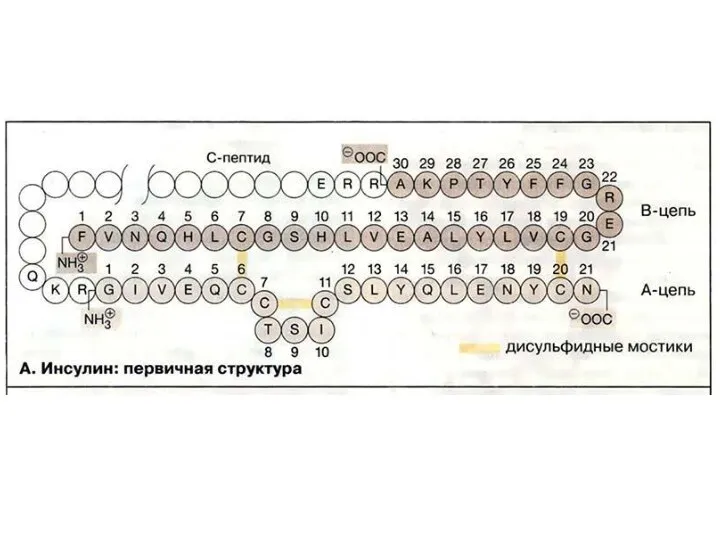
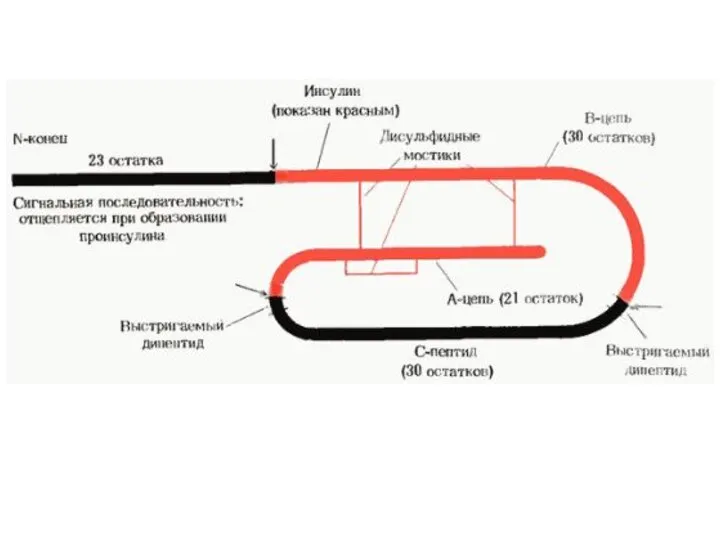
Слайд 21Уровни структуры белка
Первичная структура – последовательность аминокислотных остатков в полипептидных цепях. Первичная
Уровни структуры белка
Первичная структура – последовательность аминокислотных остатков в полипептидных цепях. Первичная


Слайд 28Для глобулярных белков наиболее важной является третичная структура. Для фибриллярных белков наиболее
Для глобулярных белков наиболее важной является третичная структура. Для фибриллярных белков наиболее
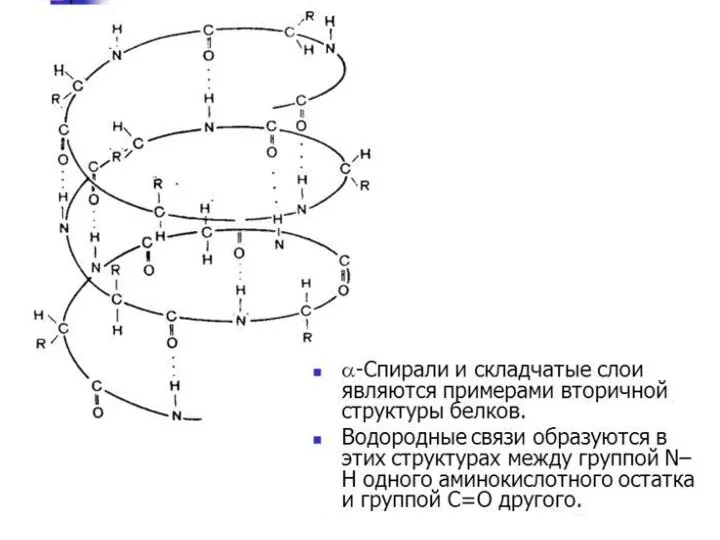
Слайд 29Вторичная структура
Вторичная структура

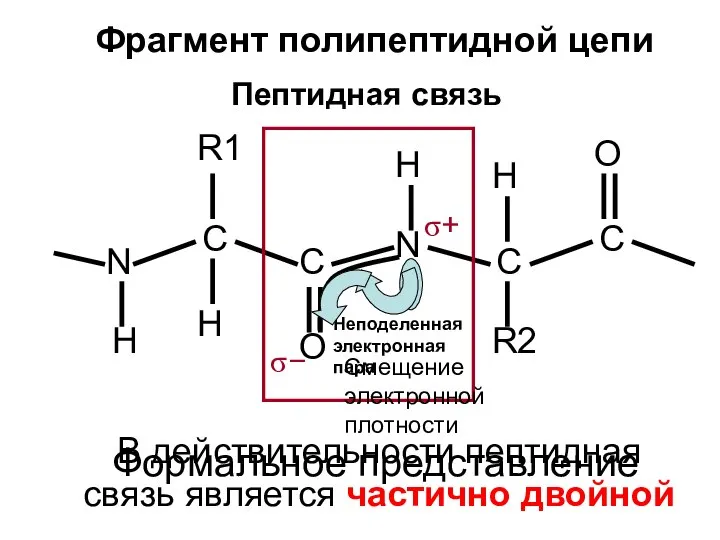
Слайд 30Пептидная связь
N
H
C
R1
H
C
O
N
H
R2
C
H
C
O
Формальное представление
• •
В действительности пептидная
связь является частично двойной
Неподеленная
электронная пара
Пептидная связь
N
H
C
R1
H
C
O
N
H
R2
C
H
C
O
Формальное представление
• •
В действительности пептидная
связь является частично двойной
Неподеленная
электронная пара
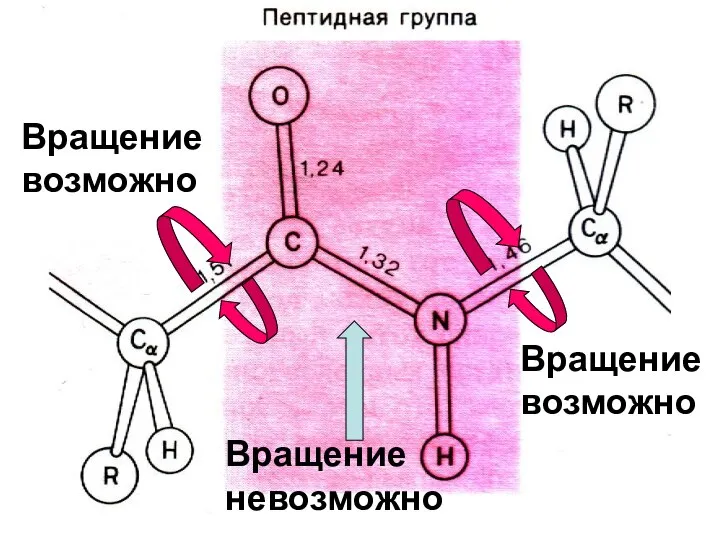
Слайд 31Вращение
невозможно
Вращение
возможно
Вращение
возможно
Вращение
невозможно
Вращение
возможно
Вращение
возможно
Слайд 32Типы регулярных вторичных структур
α-спираль
β-слой
! В белках обычно встречается правая альфа-спираль.
Типы регулярных вторичных структур
α-спираль
β-слой
! В белках обычно встречается правая альфа-спираль.

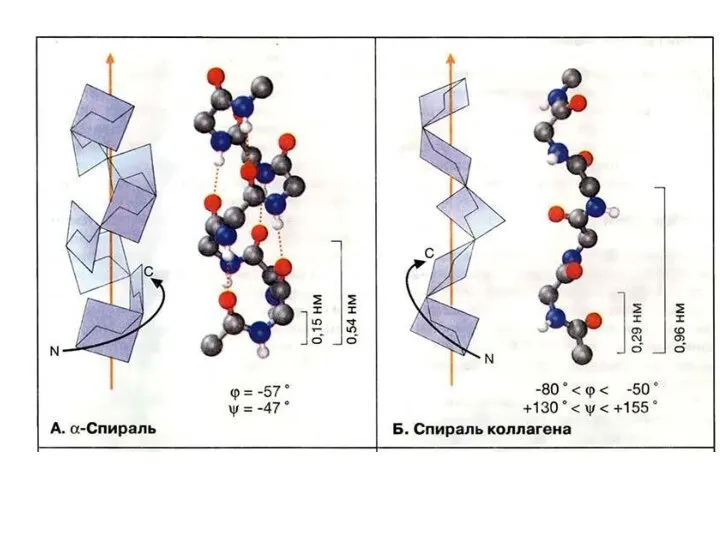
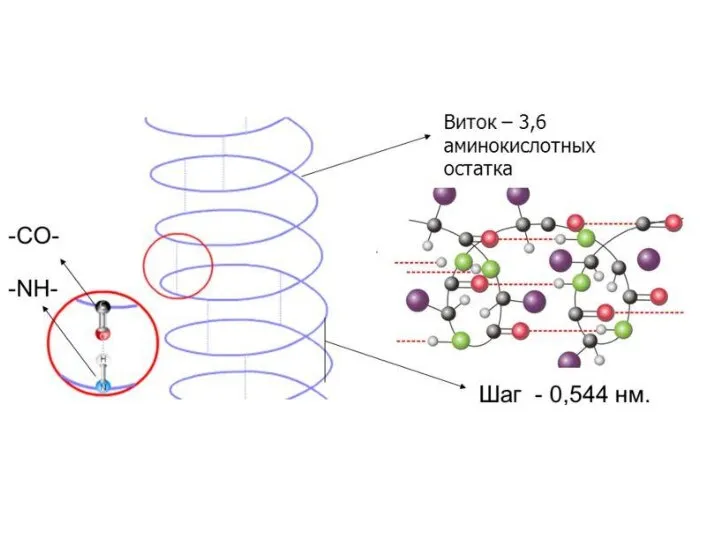
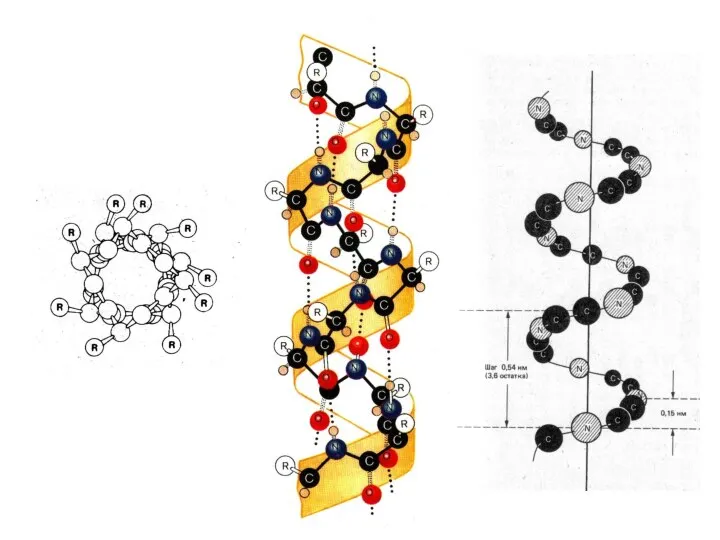
Слайд 33Свойства α-спирали
Шаг правой α-спирали – 0,54 нанометра (3,6 аминокислотных остатка).
Шаг
Свойства α-спирали
Шаг правой α-спирали – 0,54 нанометра (3,6 аминокислотных остатка).
Шаг

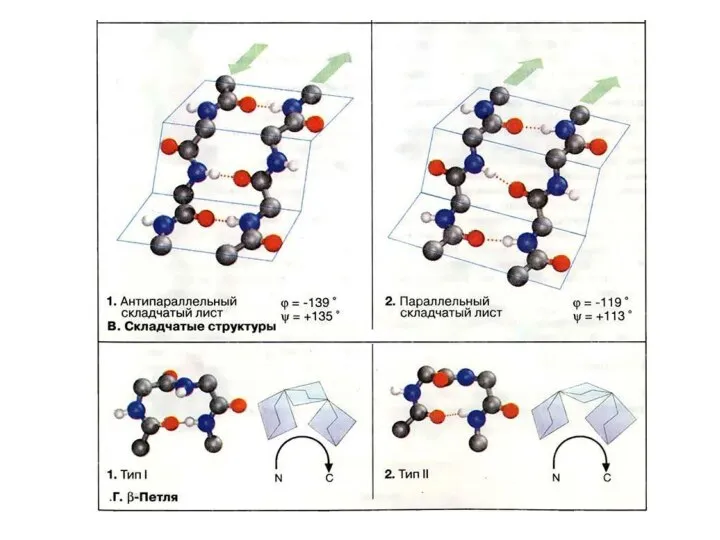
Слайд 38Типы β-слоев
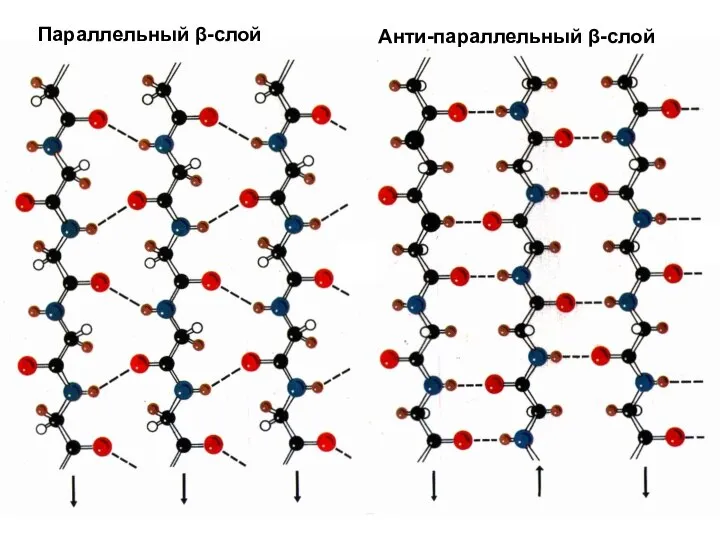
1. Параллельный (соседние полипептидные цепи идут в одном направлении)
2. Антипараллельный
Типы β-слоев
1. Параллельный (соседние полипептидные цепи идут в одном направлении)
2. Антипараллельный

Слайд 39Параллельный β-слой
Анти-параллельный β-слой
Параллельный β-слой
Анти-параллельный β-слой
Слайд 41В типичном белке около 60% аминокислотных остатков принимает участие в формировании регулярных
В типичном белке около 60% аминокислотных остатков принимает участие в формировании регулярных

Слайд 421. Третичная структура белка – общая укладка полипептидной цепи.
2. Третичная структура
1. Третичная структура белка – общая укладка полипептидной цепи.
2. Третичная структура

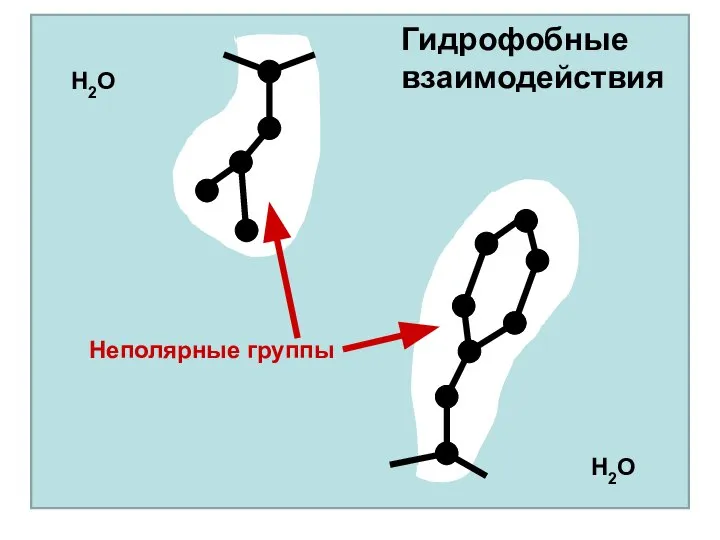
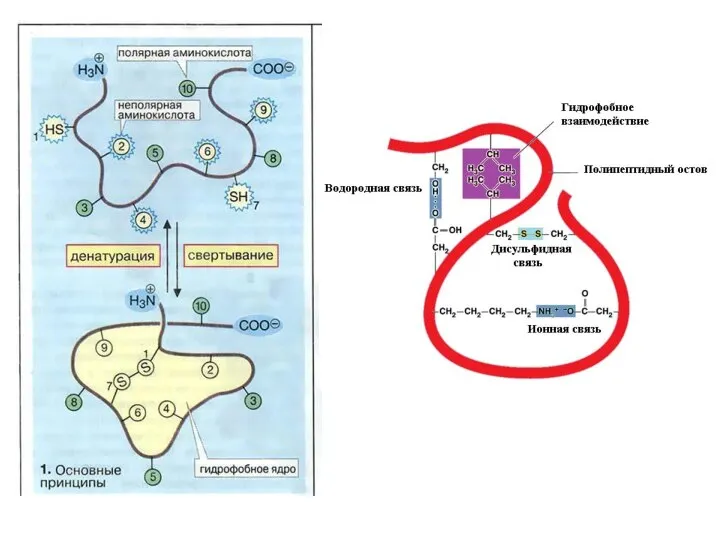
Слайд 43Взаимодействия, формирующие третичную структуру белка
Гидрофобные
Гидрофобные взаимодействия связаны с тем, что гидрофобные (неполярные
Взаимодействия, формирующие третичную структуру белка
Гидрофобные
Гидрофобные взаимодействия связаны с тем, что гидрофобные (неполярные

Слайд 44Неполярные группы
Гидрофобные
взаимодействия
H2O
H2O
Неполярные группы
Гидрофобные
взаимодействия
H2O
H2O
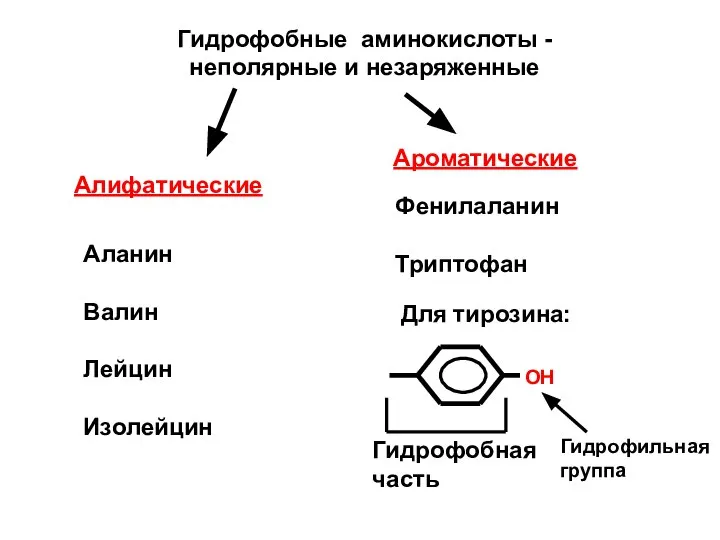
Слайд 45Гидрофобные аминокислоты -
неполярные и незаряженные
Алифатические
Ароматические
Аланин
Валин
Лейцин
Изолейцин
Фенилаланин
Триптофан
Для тирозина:
ОН
Гидрофобная
часть
Гидрофильная
группа
Гидрофобные аминокислоты -
неполярные и незаряженные
Алифатические
Ароматические
Аланин
Валин
Лейцин
Изолейцин
Фенилаланин
Триптофан
Для тирозина:
ОН
Гидрофобная
часть
Гидрофильная
группа
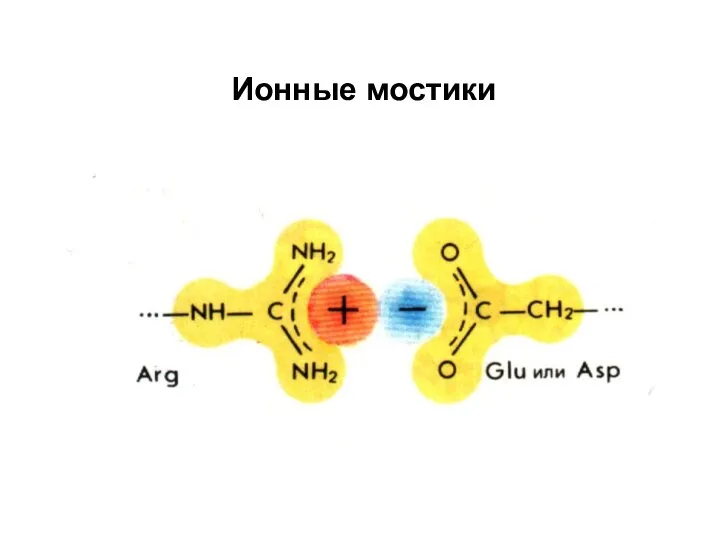
Слайд 46На поверхности белка располагаются аминокислотные остатки с ионогенными боковыми цепями, которые могут
На поверхности белка располагаются аминокислотные остатки с ионогенными боковыми цепями, которые могут

Слайд 47Ионные мостики
Ионные мостики
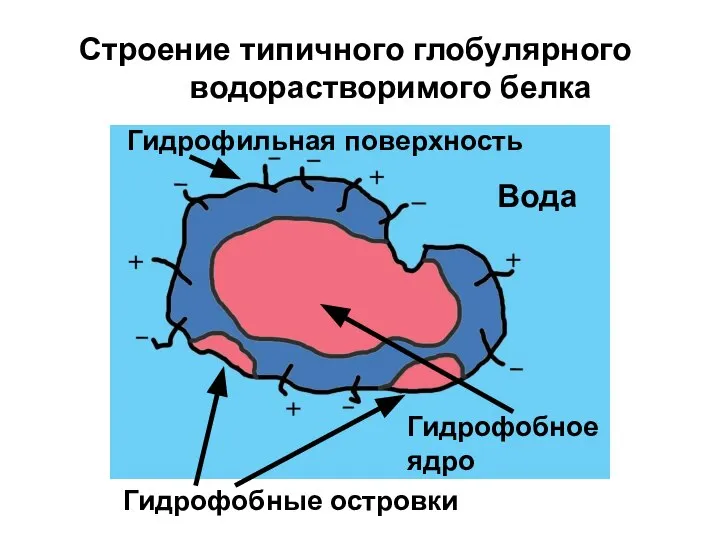
Слайд 48Строение типичного глобулярного
водорастворимого белка
Вода
Гидрофобное
ядро
Гидрофильная поверхность
Гидрофобные островки
Строение типичного глобулярного
водорастворимого белка
Вода
Гидрофобное
ядро
Гидрофильная поверхность
Гидрофобные островки
Слайд 49Процесс формирования трехмерной структуры белка – сворачивание (фолдинг) белка.
Фолдинг белка происходит,
Процесс формирования трехмерной структуры белка – сворачивание (фолдинг) белка.
Фолдинг белка происходит,

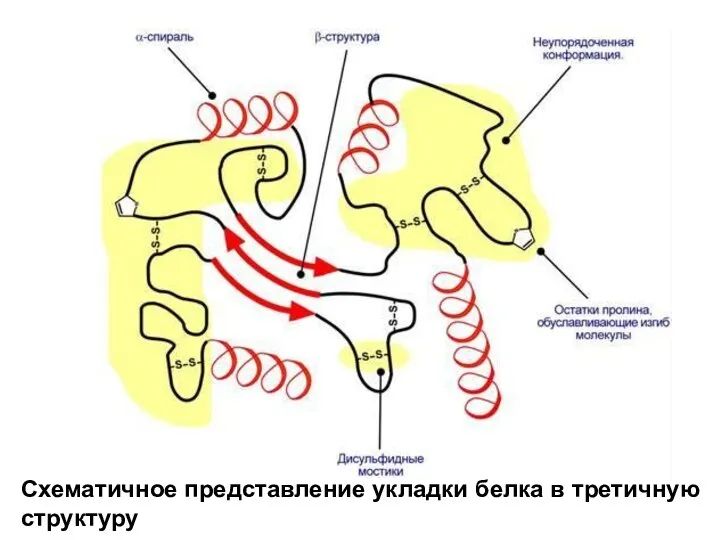
Слайд 50Схематичное представление укладки белка в третичную структуру
Схематичное представление укладки белка в третичную структуру
Слайд 52Отдельные аминокислотные остатки, удаленные в аминокислотной последовательности полипептида, сближены в третичной структуре
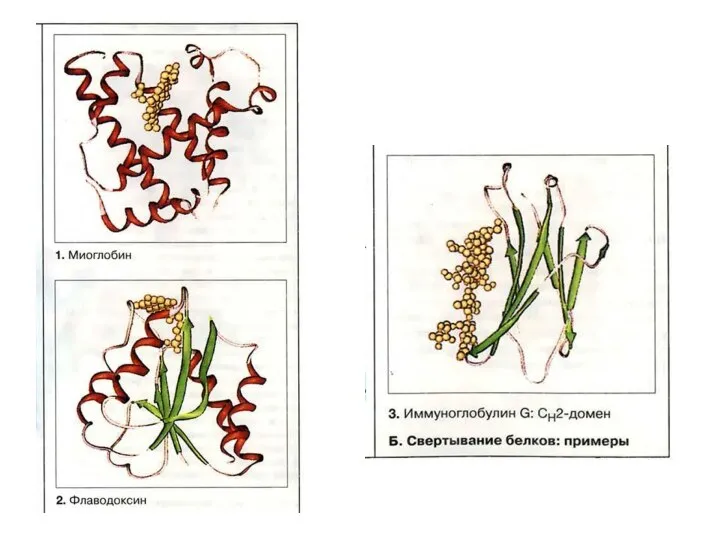
Отдельные аминокислотные остатки, удаленные в аминокислотной последовательности полипептида, сближены в третичной структуре

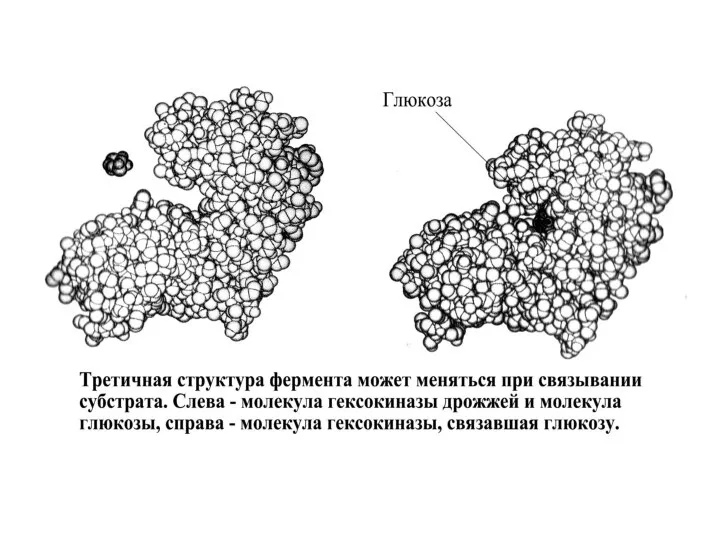
Слайд 55Домен белка — элемент третичной структуры белка, представляющий собой достаточно стабильную и независимую подструктуру белка, фолдинг которой
Домен белка — элемент третичной структуры белка, представляющий собой достаточно стабильную и независимую подструктуру белка, фолдинг которой

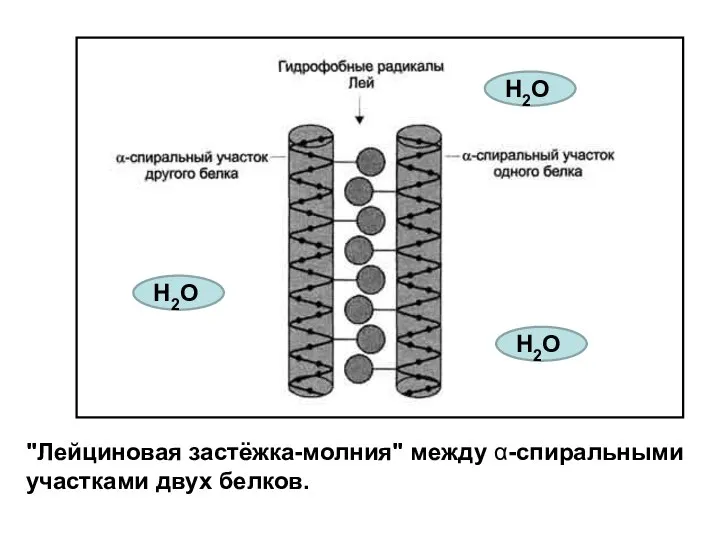
Слайд 56"Лейциновая застёжка-молния" между α-спиральными участками двух белков.
Н2О
Н2О
Н2О
"Лейциновая застёжка-молния" между α-спиральными участками двух белков.
Н2О
Н2О
Н2О
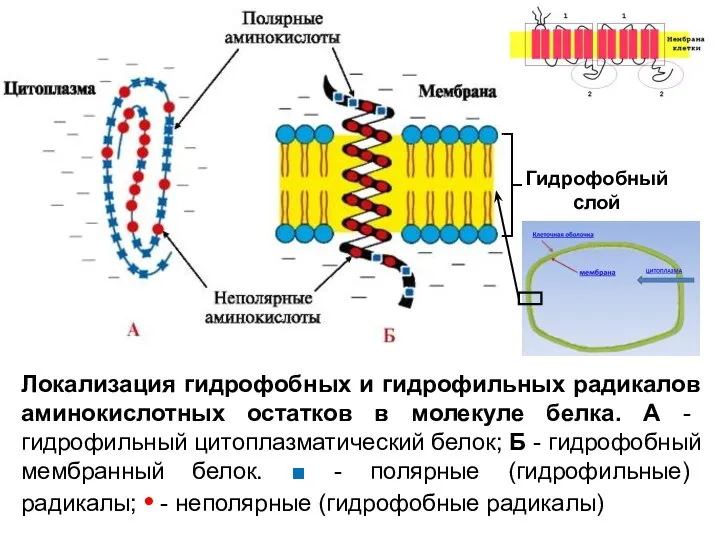
Слайд 57Локализация гидрофобных и гидрофильных радикалов аминокислотных остатков в молекуле белка. А -
Локализация гидрофобных и гидрофильных радикалов аминокислотных остатков в молекуле белка. А -
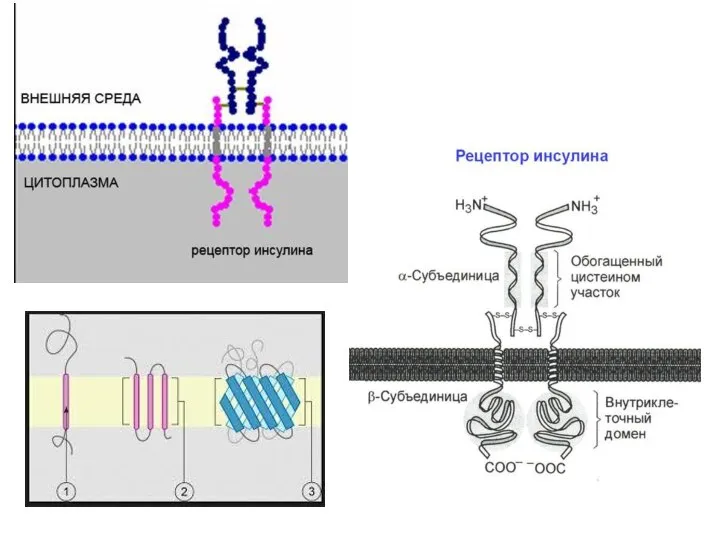
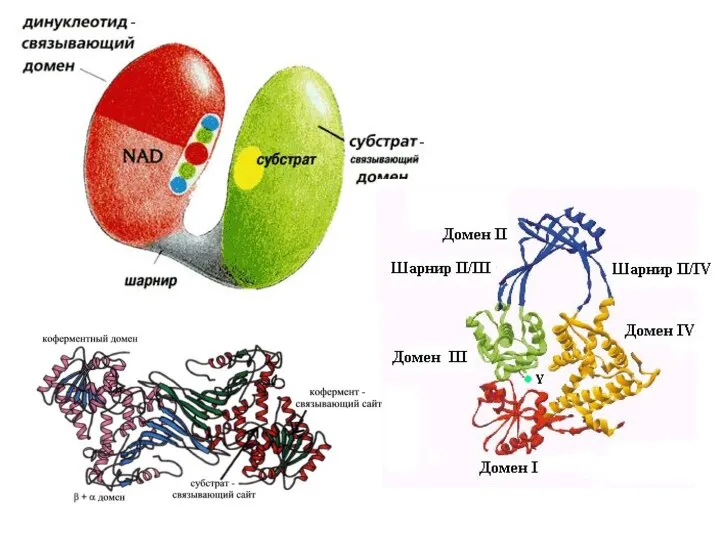
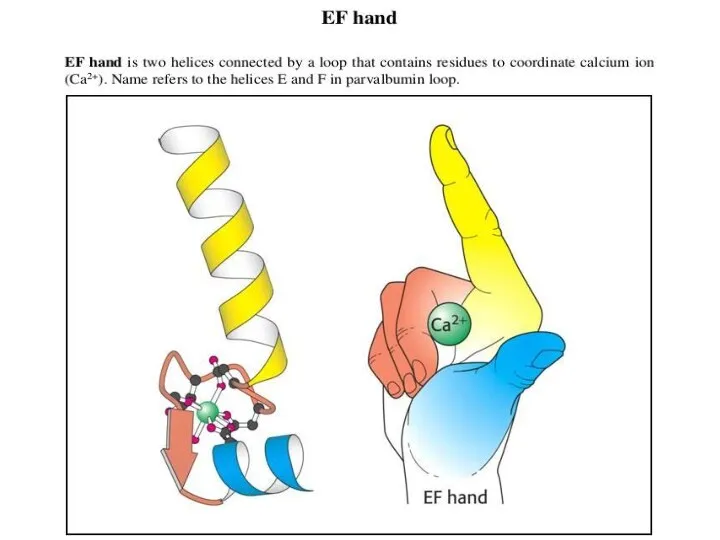
Слайд 62!!! Для сложных белков (которые содержат не только аминокислоты, но и неаминокислотные
!!! Для сложных белков (которые содержат не только аминокислоты, но и неаминокислотные

Слайд 63Четвертичная структура
Если белки состоят из двух и более полипептидных цепей, связанных между
Четвертичная структура
Если белки состоят из двух и более полипептидных цепей, связанных между

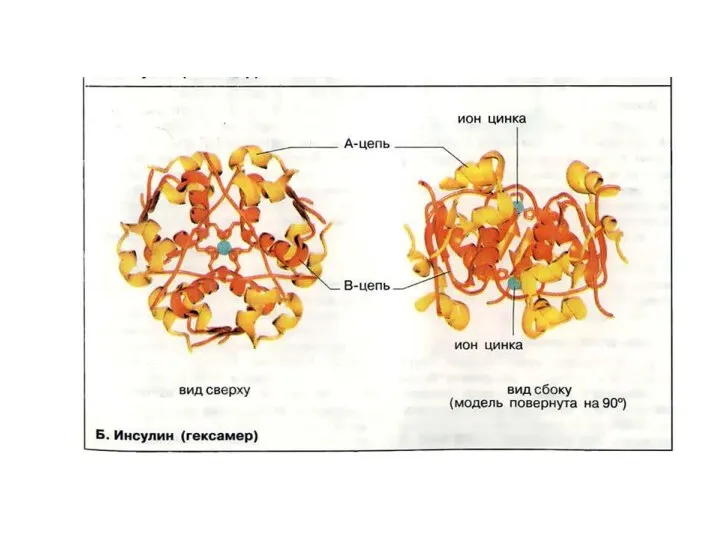
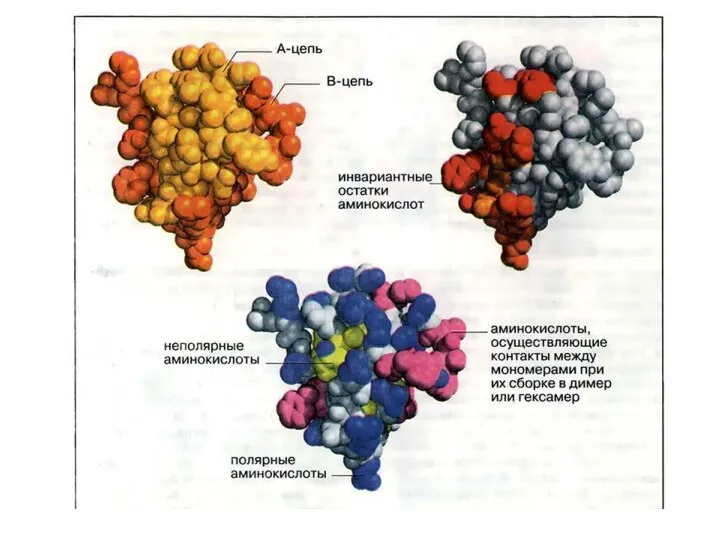
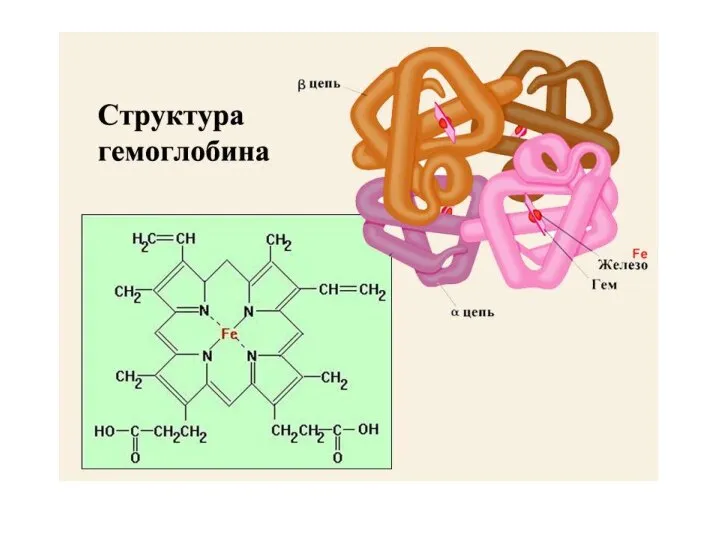
Слайд 67Многие ферменты имеют две каталитические субъединицы, которые согласованно меняют свою конформацию. В
Многие ферменты имеют две каталитические субъединицы, которые согласованно меняют свою конформацию. В

Слайд 68Сложная третичная и четвертичная структура белков необходима не только для выполнения ими
Сложная третичная и четвертичная структура белков необходима не только для выполнения ими

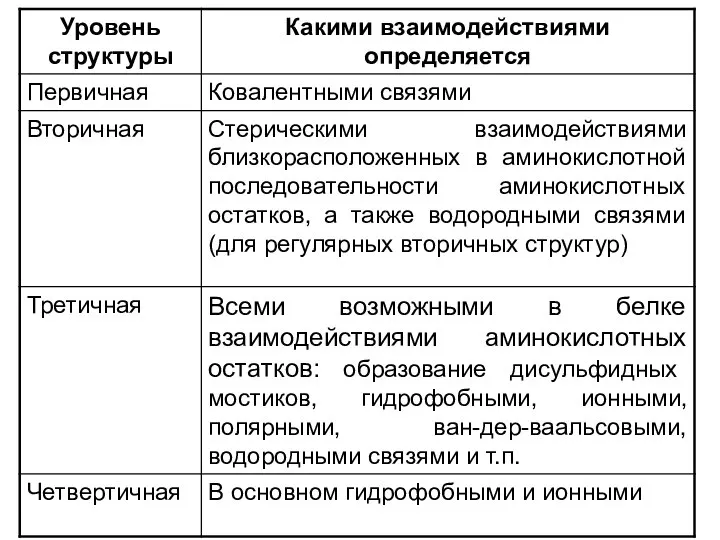
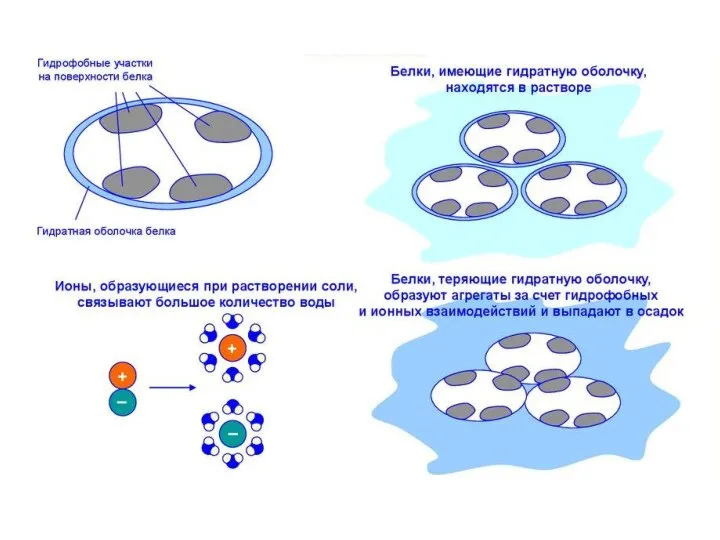
Слайд 70Денатурация белка
Денатурация белка – нарушение вторичной и/или уникальной третичной структуры белка, сопровождающееся
Денатурация белка
Денатурация белка – нарушение вторичной и/или уникальной третичной структуры белка, сопровождающееся


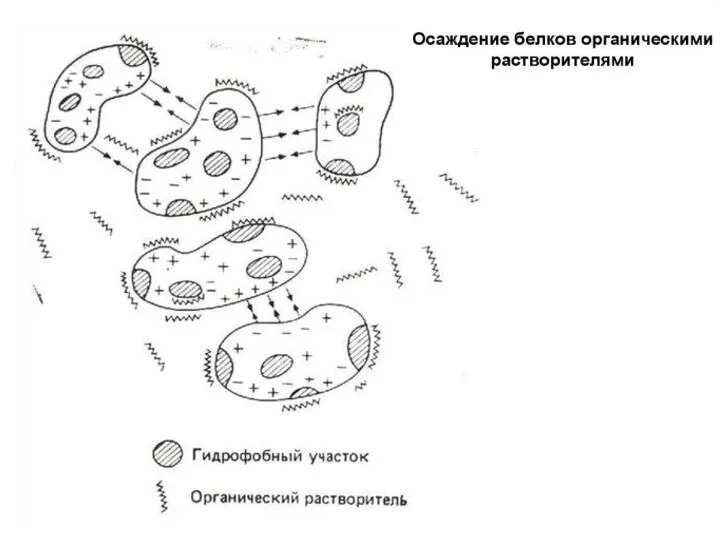
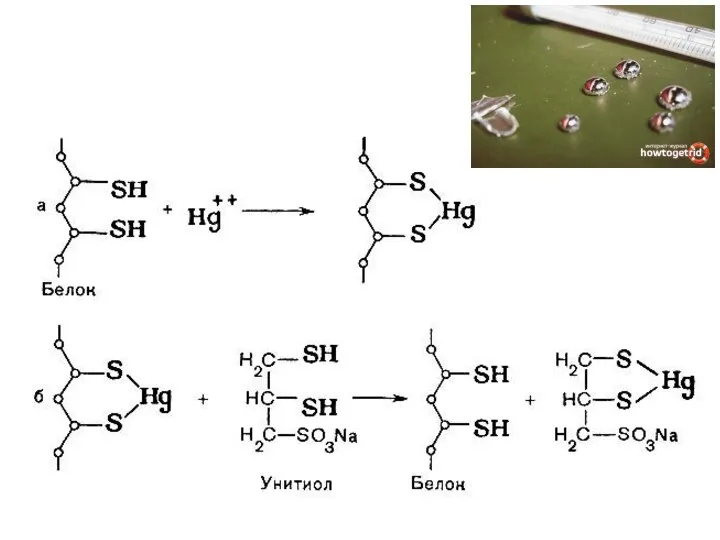
Слайд 72Денатурирующие агенты и воздействия:
1. Высокая температура
2. Сильные минеральные кислоты и основания.
Денатурирующие агенты и воздействия:
1. Высокая температура
2. Сильные минеральные кислоты и основания.


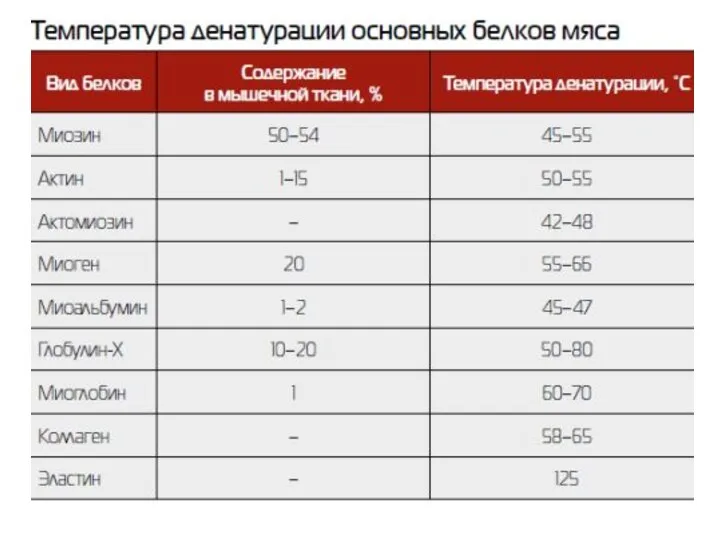

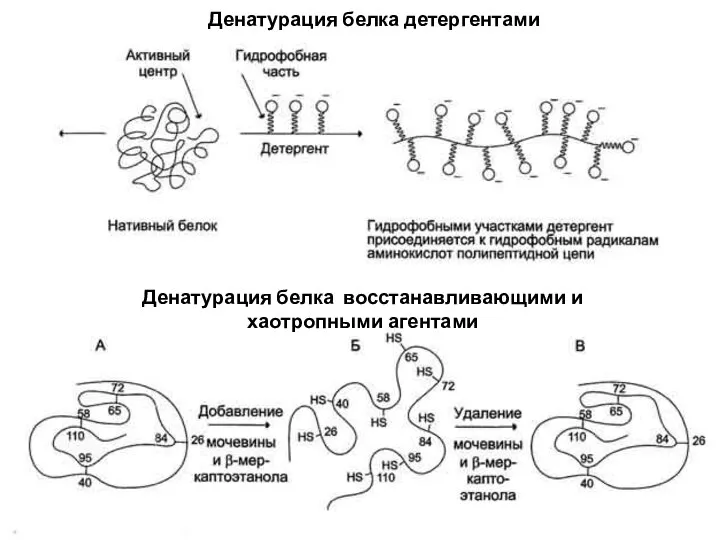
Слайд 78Денатурация белка детергентами
Денатурация белка восстанавливающими и хаотропными агентами
Денатурация белка детергентами
Денатурация белка восстанавливающими и хаотропными агентами
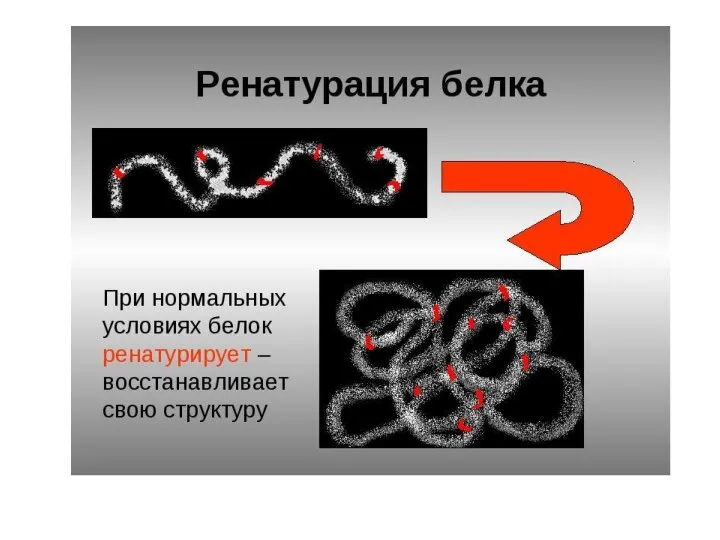
Слайд 80Ренатурация белка
Если белок при денатурации остался в растворе, он может быть ренатурирован
Ренатурация белка
Если белок при денатурации остался в растворе, он может быть ренатурирован

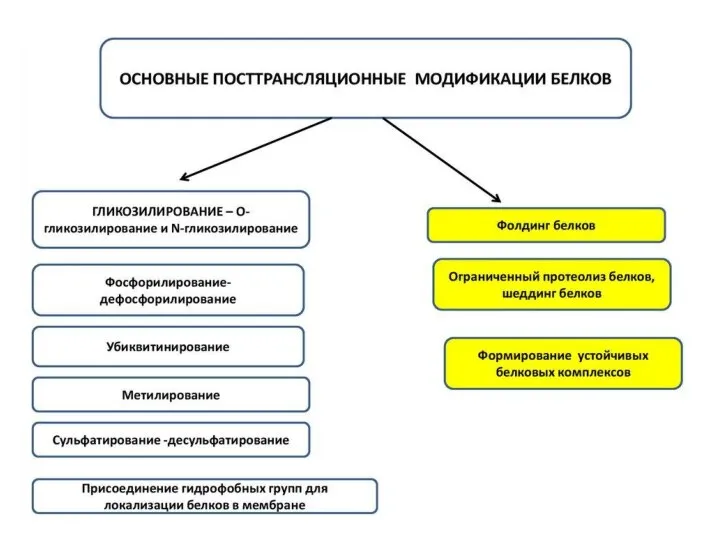
Слайд 82Посттрансляционная модификация белков
Посттрансляционная модификация белков

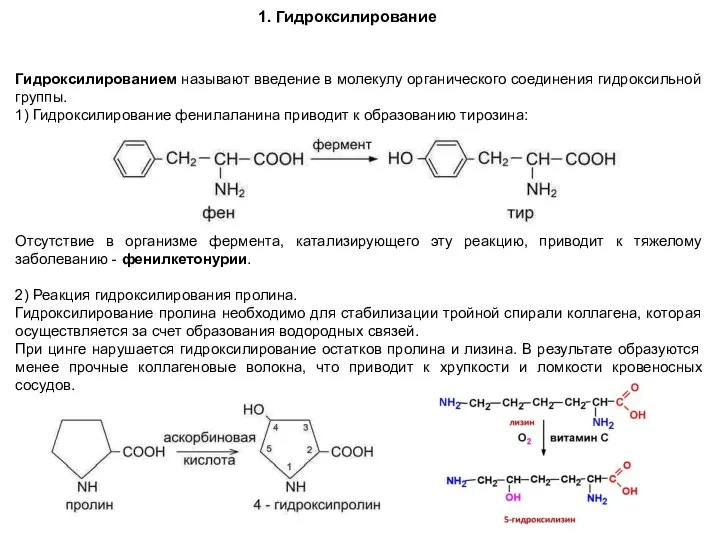
Слайд 841. Гидроксилирование
Гидроксилированием называют введение в молекулу органического соединения гидроксильной группы.
1) Гидроксилирование фенилаланина
1. Гидроксилирование
Гидроксилированием называют введение в молекулу органического соединения гидроксильной группы.
1) Гидроксилирование фенилаланина
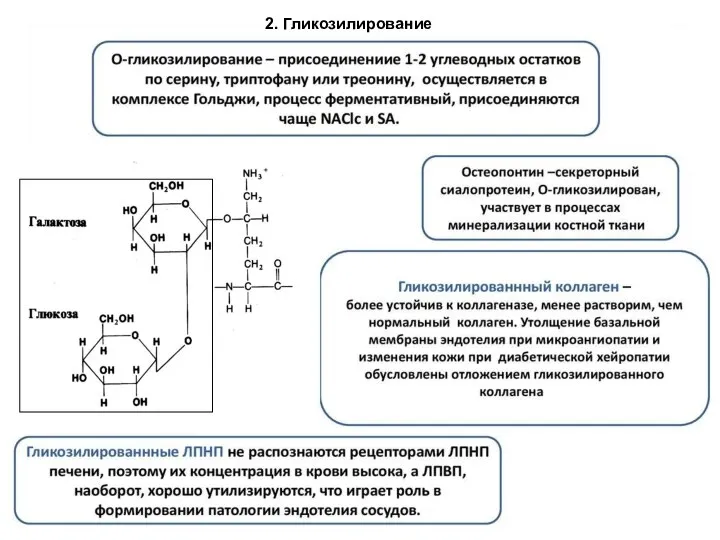
Слайд 852. Гликозилирование
2. Гликозилирование
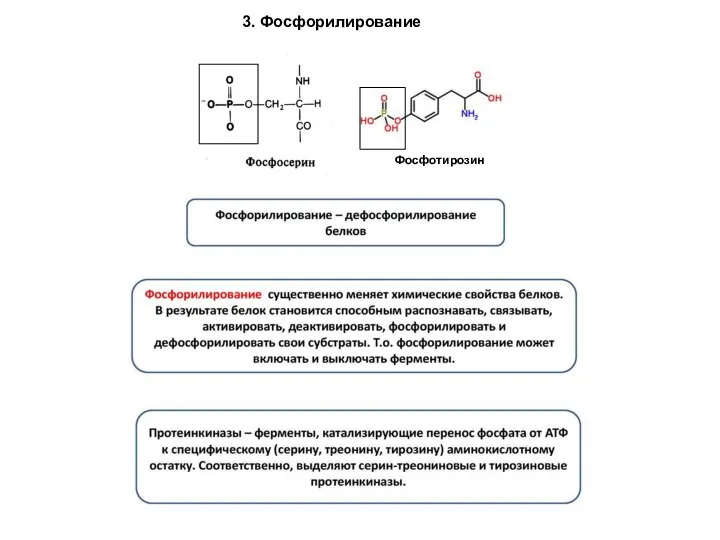
Слайд 863. Фосфорилирование
Фосфотирозин
3. Фосфорилирование
Фосфотирозин
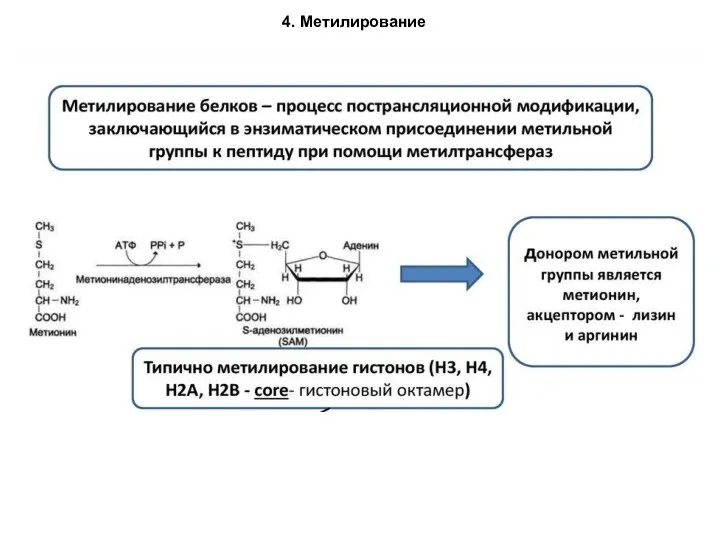
Слайд 874. Метилирование
4. Метилирование
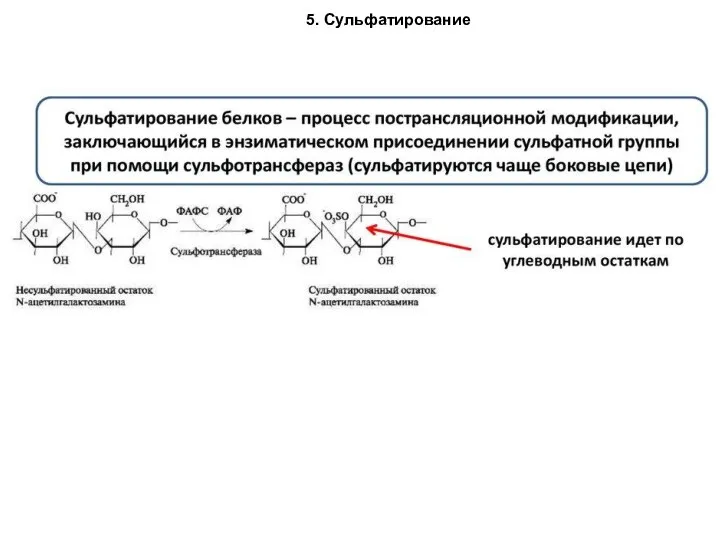
Слайд 885. Сульфатирование
5. Сульфатирование
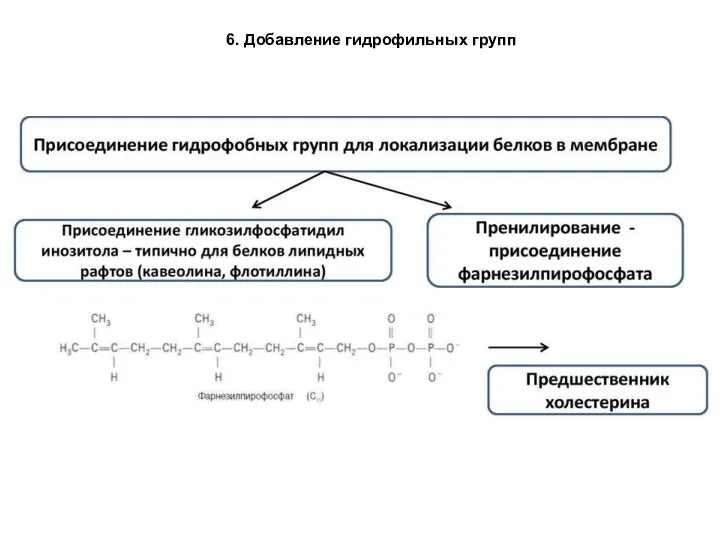
Слайд 896. Добавление гидрофильных групп
6. Добавление гидрофильных групп
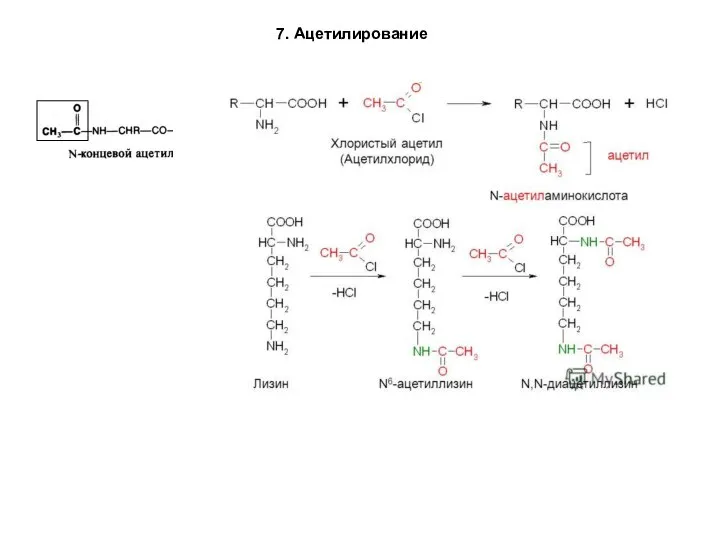
Слайд 907. Ацетилирование
7. Ацетилирование
Слайд 91Катаболизм белков
Катаболизм белков

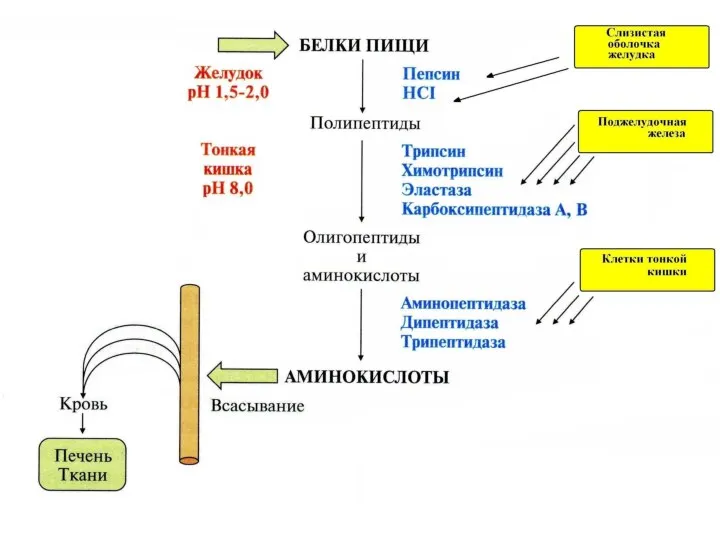
Слайд 921. В нейтральной среде полипептидная цепь является химически стабильной. В сильнокислой среде
1. В нейтральной среде полипептидная цепь является химически стабильной. В сильнокислой среде

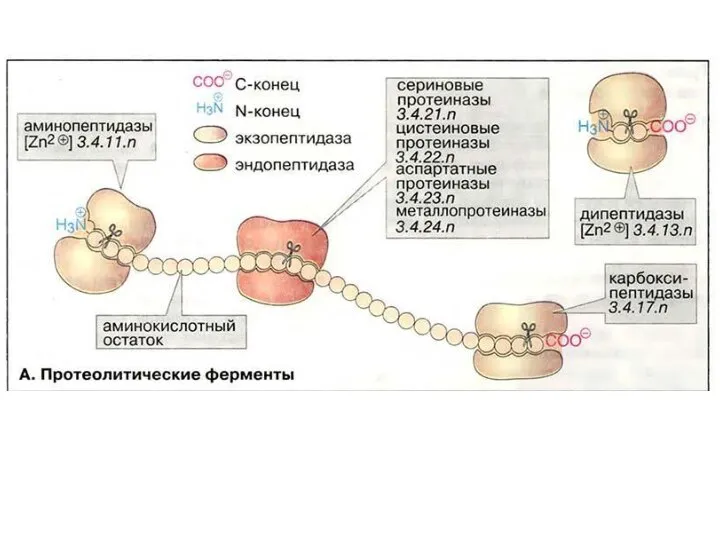
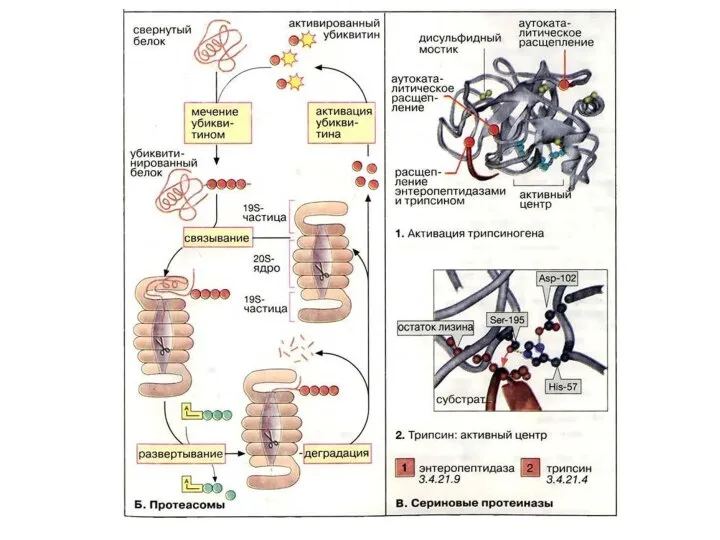
Слайд 93Ферменты, осуществляющие деградацию белков, называются протеазами. По месту атаки молекулы субстрата протеолитические
Ферменты, осуществляющие деградацию белков, называются протеазами. По месту атаки молекулы субстрата протеолитические

 Бионика, как наука
Бионика, как наука Пути и направления эволюции
Пути и направления эволюции Пол и гендер
Пол и гендер ПОЛЕЗНАЯ АЗБУКА ПИТАНИЯ Проблема: не всегда умеем правильно выбирать продукты для здорового питания (не знаем о пользе ов
ПОЛЕЗНАЯ АЗБУКА ПИТАНИЯ Проблема: не всегда умеем правильно выбирать продукты для здорового питания (не знаем о пользе ов Гигиена кормов и кормления животных
Гигиена кормов и кормления животных Чудесные цветники весной
Чудесные цветники весной Холодолюбивая фауна в антропогенном периоде
Холодолюбивая фауна в антропогенном периоде Строение и функции клетки. Цитоплазма и её органоиды
Строение и функции клетки. Цитоплазма и её органоиды Презентация на тему Первоцветы Особенности раннецветущих растений
Презентация на тему Первоцветы Особенности раннецветущих растений  Рассматривание клеток и тканей в оптический микроскоп
Рассматривание клеток и тканей в оптический микроскоп Отдел голосеменные
Отдел голосеменные Аноклазе: кудонь жуватат
Аноклазе: кудонь жуватат Общая характеристика обмена веществ и преобразование энергии
Общая характеристика обмена веществ и преобразование энергии Обмен углеводов
Обмен углеводов Філогенія Excavata
Філогенія Excavata Дыхательная система. Полость носа, гортань, трахея
Дыхательная система. Полость носа, гортань, трахея Многообразие кишечнополостных (7 класс)
Многообразие кишечнополостных (7 класс) Слайды по теме Царство Грибы
Слайды по теме Царство Грибы Соматический и автономный отделы нервной системы
Соматический и автономный отделы нервной системы Филин
Филин Вирусология и открытие вирусов
Вирусология и открытие вирусов Свертывание крови. Группы крови. Переливание. 8 класс
Свертывание крови. Группы крови. Переливание. 8 класс Витаминдер және олардың маңызы
Витаминдер және олардың маңызы Медицинская арахноэнтомология
Медицинская арахноэнтомология Вирусы грибов
Вирусы грибов Белки. Функции белков
Белки. Функции белков Развитие жизни в криптозое
Развитие жизни в криптозое Les kaktas
Les kaktas