- Радиационная генетика

Содержание
- 2. Историю изучения мутационного процесса можно разделить на 5 периодов. В течение первого периода (1900--1927) были развиты
- 3. Мутагенез Нобелевская премия - 1946
- 4. Третий период начался незадолго до второй мировой войны, когда открытие химического мутагенеза поставило новые проблемы, а
- 5. Это направление оказалось весьма плодотворным и до сих пор служит источником информации о действии мутагенов на
- 6. Первый период: с 1900 по 1927 г. Голландский ботаник Гуго Де Фриз, один из тех, кто
- 7. Он понял, что при рода мутаций нерасторжимо связана с природой гена. В своем выступлении, относящемся к
- 8. Фундаментальные проблемы, которые возникли в первый период: 1. Какое свойство генетического материала позволяет ему точно реплицироваться
- 9. Третий период: (с начала второй мировой войны до 1953 г.) Этот период в изучении мутагенеза был
- 10. К концу этого периода накопилось большое количество следующих проблем: 1. Загадка точной репликации генов лишь осложнилась
- 11. Четвертый период: (с 1953 до начала 1965 г.) Этот период менее четко определен во времени, чем
- 12. Пятый период: (с 1965 г. по сегодняшний день) Сегодняшние исследования целиком базируются на достижениях предыдущих периодов.
- 13. Типы зависимостей частоты образования мутаций от дозы облучения
- 14. Возрастание вероятности инактивации клеток, имеющих мутационное повреждение. Harm, Stein, 1956 Алиханян и соавт, 1960 Кривиский, 1960
- 15. ОСНОВНЫЕ ТИПЫ МУТАГЕННЫХ ПОВРЕЖДЕНИЙ ДНК Мутагенные факторы можно разделить на три группы. В первую входят агенты,
- 16. При действии ионизирующих излучений премутационными событиями служат в основном щелочестабильные повреждения ДНК. К ним следует прежде
- 17. Так как ОР в данном случае не могут перейти из донорской ДНК в хромосому клетки, то
- 18. МУТАГЕННОЕ ДЕЙСТВИЕ γ-ИЗЛУЧЕНИЯ НА КЛЕТКИ БАКТЕРИЙ Дозовая зависимость выхода мутаций к фагоустойчивости у бактерий, выявленная в
- 19. Квадратичная дозовая зависимость по образованию триптофановых реверсийу Е.соli - WР-2 была обнаружена в 1978 . В
- 20. Индукция lac- мутаций у бактерий E.coli при γ-облучении
- 21. ОСНОВНЫЕ ГИПОТЕЗЫ МЕХАНИЗМА МУТАГЕННОГО ДЕЙСТВИЯ ИОНИЗИРУЮЩИХ ИЗЛУЧЕНИЙ 1. Классическая модель Линейный тип зависимости Nm/N(D) часто наблюдается
- 22. 2. Репарационная модель. Для объяснения квадратичных зависимостей было предположено, что в формировании начального премутационного события участвуют
- 23. Nm/N(D) = (βD [1 - exp (- aD)], uде β= kMl/M - коэффициент пропорциональности. Из этого
- 24. 3. Биофизическая модель.
- 25. При действии ионизирующего излучения в ДНК клеток с некоторой частотой образуются прямые двунитевые разрывы (ПДР) ДНК,
- 26. Из неотрепарированных данным типом репарации КП и повреждений, которые не смогли быть отрепарированы polA-зависимой и recA-lexA-зависимой
- 27. Комплекс UmuDD’C играет ингибирующую роль в SOS-мутагенезе, секвестрируя UmuD’ активности. Участие ДНК-полимеразы III или некоторых её
- 28. Из неотрепарированных данным типом репарации КП и повреждений, которые не смогли быть отрепарированы polA-зависимой и recA-lexA-зависимой
- 29. RecA протеаза в ходе SOS ответа расщепляет UmuD белок, переводя его в активную UmuD’ форму. UmuD’
- 30. UmuC, UmuD’, RecA , SSB через сайты с повреждениями. Комплекс UmuDD’C играет ингибирующую роль в SOS-мутагенезе,
- 31. Таким образом, в биофизической модели закрепление премутационного повреждения в мутацию точкового типа при действии ионизирующих излучений
- 32. SOS –тест системы
- 33. RecA ген LexA ген >40 генов SOS регуляция
- 34. SOS-репарация ДНК γ Rec A protease rec A lex A >40 genes rec A lex A
- 35. β-галактозидаза лактоза β-галактозидаза lacZ галактоза глюкоза
- 36. Lac-оперон β-галактозидаза О
- 37. SOS-хромотест rec A lex A rec A lex A Rec A lacZ β-галактозидаза ОНФГ lacZ
- 38. SOS-хромотест
- 39. Конструкция pPLS-1 плазмиды FMNH2+ RCHO + O2 → FMN + RCOOH +H2O + hν luciferase
- 40. SOS – ответ при УФ-облучении Кинетика SOS lux индукции
- 41. Чувствительность SOS-lux теста к различным генотоксинам
- 42. Сравнительная оценка чувствительности различных тест-систем
- 43. фаг λ λ
- 44. Частица фага Хромосома фага длиной около 50000 пар оснований находится в головке фага, где она намотана
- 45. Пути развития фага
- 46. Репрессор Репрессор-белок из 236 аминокислот. Димер формируется главным образом за счет контактов между карбокси - концевыми
- 47. Белок Cro Cro -control of repressor and other genes
- 48. Два промотора,расположенные «спина к спине»,запускают движение полимеразы в противоположные стороны. Правый оператор состоит из трех частей
- 49. Действие репрессора и Cro
- 50. Репрессор,связанный с OR2 и OR1. Репрессор,связанный с OR1 и OR2,стимулируют промотор PRM и одновременно выключает PR
- 51. Как УФ-облучение изменяет поведение лизогенных клеток?
- 52. Расщепление репрессора белком RecA.
- 53. Расщепленные молекулы репрессора не могут образовывать димеры,поэтому в результате облучения они отсоединяются от оператора и не
- 54. Связывание белка Cro с OR. Димеры Cro независимо связываются с каждым из трех участков оператора.
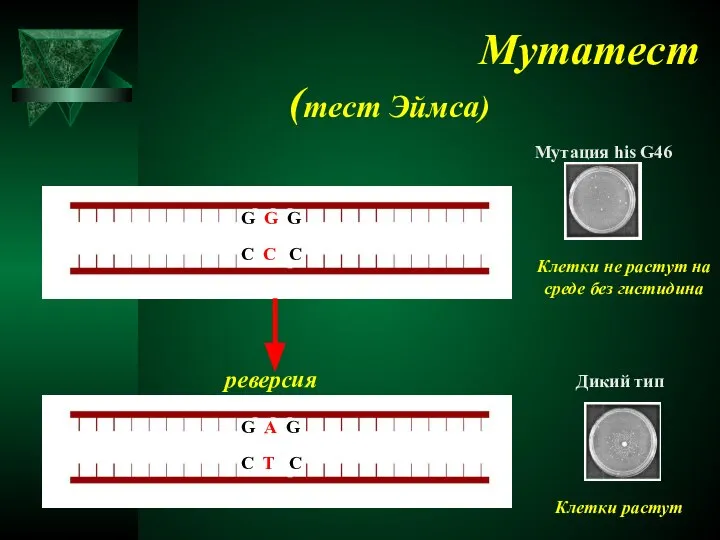
- 56. Мутатест (тест Эймса) Мутация his G46 Клетки не растут на среде без гистидина реверсия
- 58. Скачать презентацию

Слайд 3Мутагенез
Нобелевская премия - 1946
Мутагенез
Нобелевская премия - 1946

Слайд 4Третий период начался незадолго до второй мировой войны, когда открытие химического мутагенеза
Третий период начался незадолго до второй мировой войны, когда открытие химического мутагенеза

Слайд 5Это направление оказалось весьма плодотворным и до сих пор служит источником информации
Это направление оказалось весьма плодотворным и до сих пор служит источником информации

Слайд 6Первый период: с 1900 по 1927 г.
Голландский ботаник Гуго Де Фриз, один
Первый период: с 1900 по 1927 г.
Голландский ботаник Гуго Де Фриз, один

Слайд 7Он понял, что при рода мутаций нерасторжимо связана с природой гена. В
Он понял, что при рода мутаций нерасторжимо связана с природой гена. В

Слайд 8Фундаментальные проблемы, которые возникли в первый период:
1. Какое свойство генетического материала позволяет
Фундаментальные проблемы, которые возникли в первый период:
1. Какое свойство генетического материала позволяет

Слайд 9Третий период:
(с начала второй мировой войны до 1953 г.)
Этот период в изучении
Третий период:
(с начала второй мировой войны до 1953 г.)
Этот период в изучении

Слайд 10К концу этого периода накопилось большое количество следующих проблем:
1. Загадка точной
К концу этого периода накопилось большое количество следующих проблем:
1. Загадка точной

Слайд 11Четвертый период:
(с 1953 до начала 1965 г.)
Этот период менее четко определен
Четвертый период:
(с 1953 до начала 1965 г.)
Этот период менее четко определен

Слайд 12Пятый период:
(с 1965 г. по сегодняшний день)
Сегодняшние исследования целиком базируются на
Пятый период:
(с 1965 г. по сегодняшний день)
Сегодняшние исследования целиком базируются на

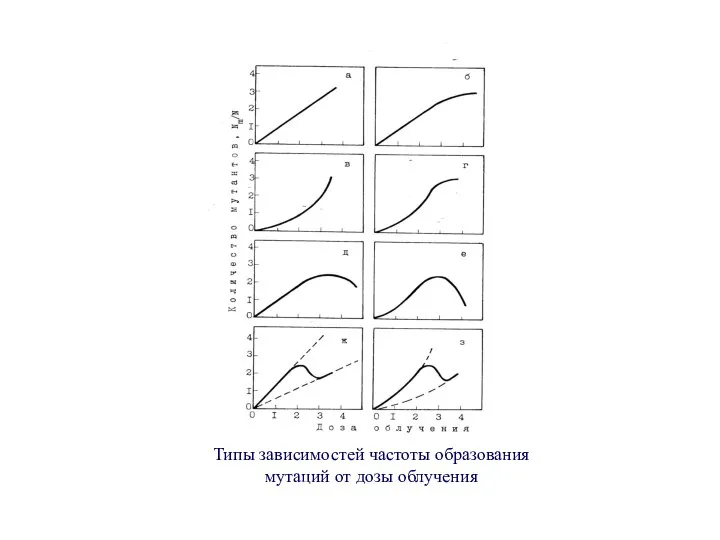
Слайд 13Типы зависимостей частоты образования мутаций от дозы облучения
Типы зависимостей частоты образования мутаций от дозы облучения
Слайд 14Возрастание вероятности инактивации клеток, имеющих мутационное повреждение.
Harm, Stein, 1956
Алиханян и соавт, 1960
Кривиский,
Возрастание вероятности инактивации клеток, имеющих мутационное повреждение.
Harm, Stein, 1956
Алиханян и соавт, 1960
Кривиский,

Слайд 15ОСНОВНЫЕ ТИПЫ МУТАГЕННЫХ ПОВРЕЖДЕНИЙ
ДНК
Мутагенные факторы можно разделить на три группы. В первую
ОСНОВНЫЕ ТИПЫ МУТАГЕННЫХ ПОВРЕЖДЕНИЙ
ДНК
Мутагенные факторы можно разделить на три группы. В первую

Слайд 16При действии ионизирующих излучений
премутационными событиями служат в основном щелочестабильные повреждения ДНК. К
При действии ионизирующих излучений
премутационными событиями служат в основном щелочестабильные повреждения ДНК. К

Слайд 17Так как ОР в данном случае не могут перейти из донорской ДНК
Так как ОР в данном случае не могут перейти из донорской ДНК

Слайд 18МУТАГЕННОЕ ДЕЙСТВИЕ γ-ИЗЛУЧЕНИЯ НА КЛЕТКИ БАКТЕРИЙ
Дозовая зависимость выхода мутаций к фагоустойчивости у
МУТАГЕННОЕ ДЕЙСТВИЕ γ-ИЗЛУЧЕНИЯ НА КЛЕТКИ БАКТЕРИЙ
Дозовая зависимость выхода мутаций к фагоустойчивости у

Слайд 19
Квадратичная дозовая зависимость по образованию триптофановых реверсийу Е.соli - WР-2 была
Квадратичная дозовая зависимость по образованию триптофановых реверсийу Е.соli - WР-2 была

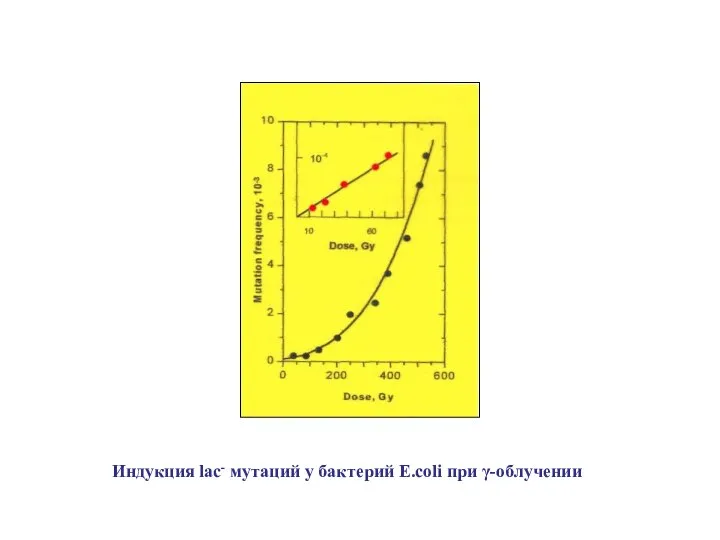
Слайд 20Индукция lac- мутаций у бактерий E.coli при γ-облучении
Индукция lac- мутаций у бактерий E.coli при γ-облучении
Слайд 21ОСНОВНЫЕ ГИПОТЕЗЫ МЕХАНИЗМА МУТАГЕННОГО ДЕЙСТВИЯ ИОНИЗИРУЮЩИХ ИЗЛУЧЕНИЙ
1. Классическая модель
Линейный тип зависимости Nm/N(D)
ОСНОВНЫЕ ГИПОТЕЗЫ МЕХАНИЗМА МУТАГЕННОГО ДЕЙСТВИЯ ИОНИЗИРУЮЩИХ ИЗЛУЧЕНИЙ
1. Классическая модель
Линейный тип зависимости Nm/N(D)

Слайд 222. Репарационная модель.
Для объяснения квадратичных зависимостей было предположено, что в формировании
Для объяснения квадратичных зависимостей было предположено, что в формировании

Слайд 23Nm/N(D) = (βD [1 - exp (- aD)],
uде β= kMl/M - коэффициент
Nm/N(D) = (βD [1 - exp (- aD)],
uде β= kMl/M - коэффициент
![Nm/N(D) = (βD [1 - exp (- aD)], uде β= kMl/M -](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/946729/slide-22.jpg)
Слайд 243. Биофизическая модель.
3. Биофизическая модель.
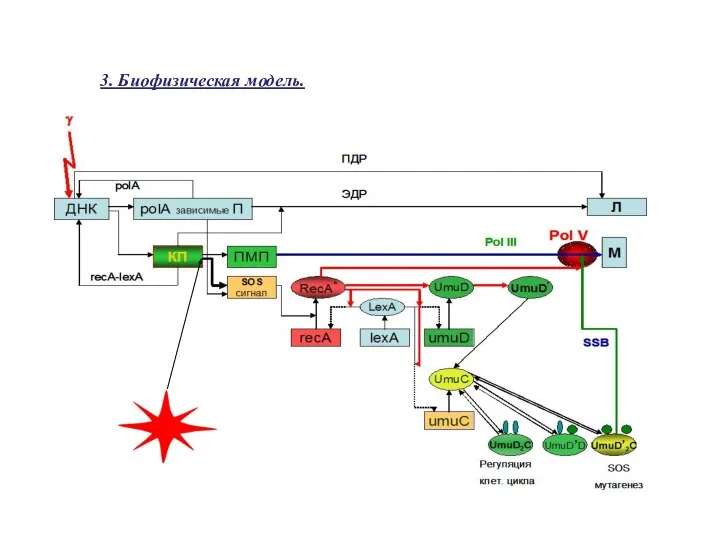
Слайд 25При действии ионизирующего излучения в ДНК клеток с некоторой частотой образуются прямые
При действии ионизирующего излучения в ДНК клеток с некоторой частотой образуются прямые

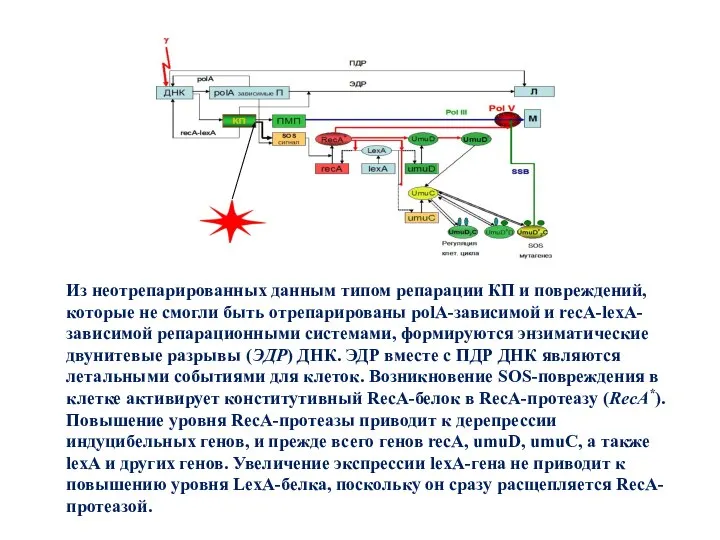
Слайд 26Из неотрепарированных данным типом репарации КП и повреждений, которые не смогли быть
Из неотрепарированных данным типом репарации КП и повреждений, которые не смогли быть

Слайд 27Комплекс UmuDD’C играет ингибирующую роль в SOS-мутагенезе, секвестрируя UmuD’ активности. Участие ДНК-полимеразы
Комплекс UmuDD’C играет ингибирующую роль в SOS-мутагенезе, секвестрируя UmuD’ активности. Участие ДНК-полимеразы

Слайд 28Из неотрепарированных данным типом репарации КП и повреждений, которые не смогли быть
Из неотрепарированных данным типом репарации КП и повреждений, которые не смогли быть
Слайд 29 RecA протеаза в ходе SOS ответа расщепляет UmuD белок, переводя его
RecA протеаза в ходе SOS ответа расщепляет UmuD белок, переводя его
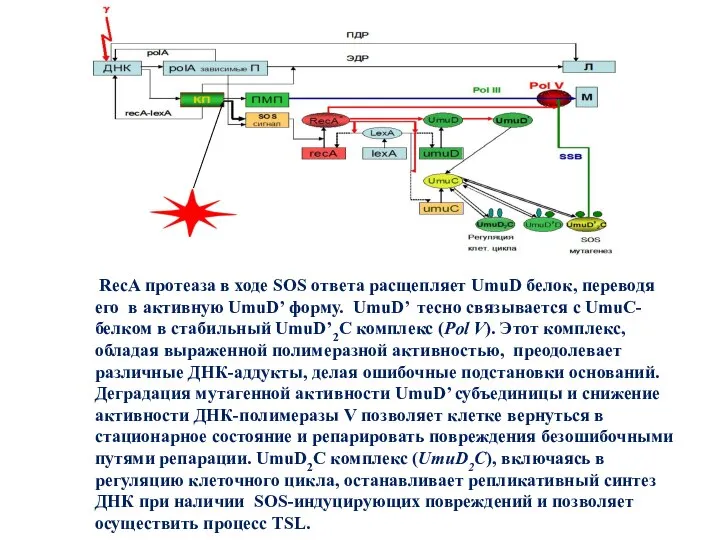
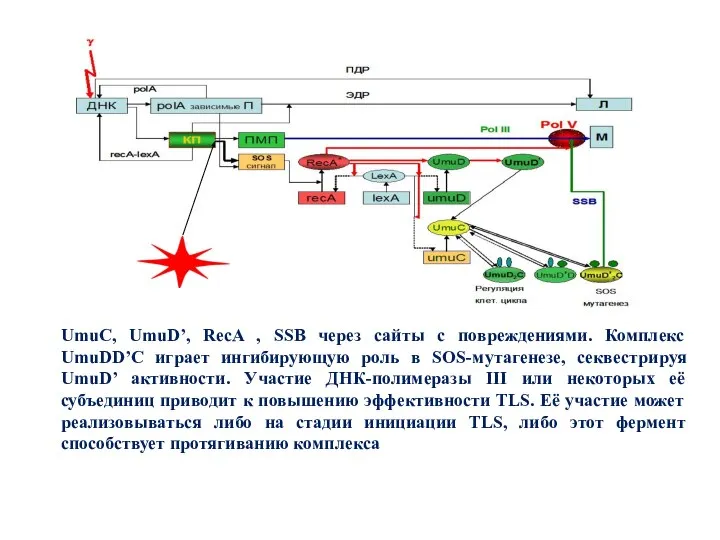
Слайд 30UmuC, UmuD’, RecA , SSB через сайты с повреждениями. Комплекс UmuDD’C играет
UmuC, UmuD’, RecA , SSB через сайты с повреждениями. Комплекс UmuDD’C играет
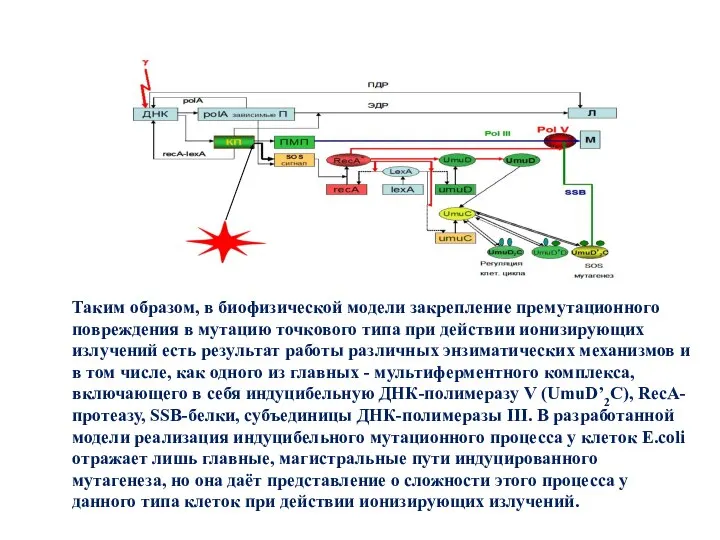
Слайд 31Таким образом, в биофизической модели закрепление премутационного повреждения в мутацию точкового типа
Таким образом, в биофизической модели закрепление премутационного повреждения в мутацию точкового типа
Слайд 32SOS –тест системы
SOS –тест системы

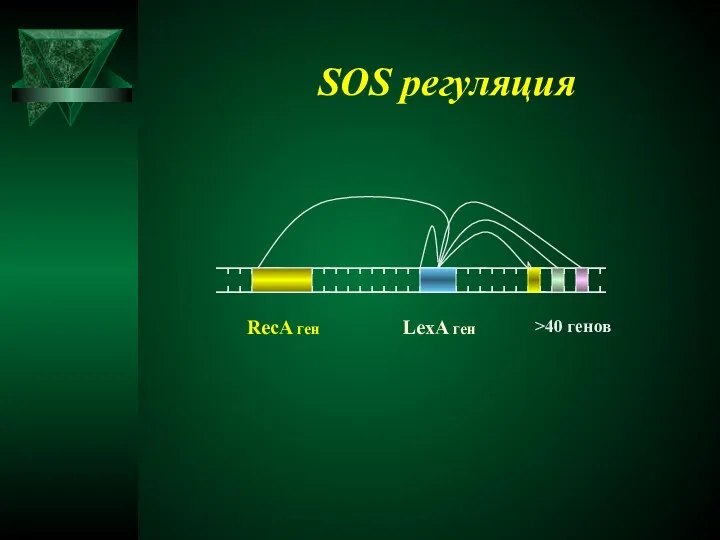
Слайд 33RecA ген
LexA ген
>40 генов
SOS регуляция
RecA ген
LexA ген
>40 генов
SOS регуляция
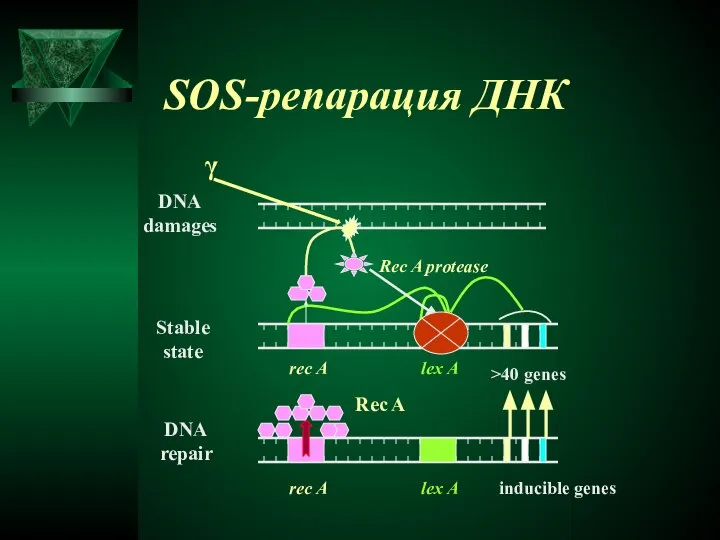
Слайд 34SOS-репарация ДНК
γ
Rec A protease
rec A
lex A
>40 genes
rec A
lex A
inducible genes
DNA damages
Stable
SOS-репарация ДНК
γ
Rec A protease
rec A
lex A
>40 genes
rec A
lex A
inducible genes
DNA damages
Stable
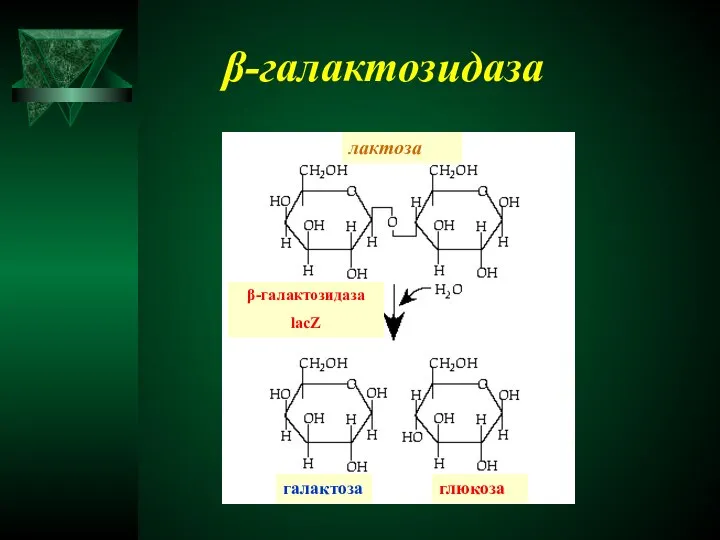
Слайд 35β-галактозидаза
лактоза
β-галактозидаза
lacZ
галактоза
глюкоза
β-галактозидаза
лактоза
β-галактозидаза
lacZ
галактоза
глюкоза
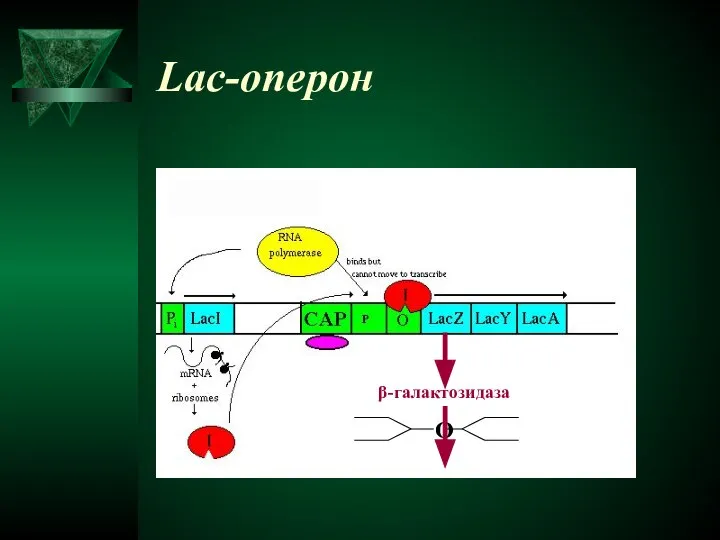
Слайд 36Lac-оперон
β-галактозидаза
О
Lac-оперон
β-галактозидаза
О
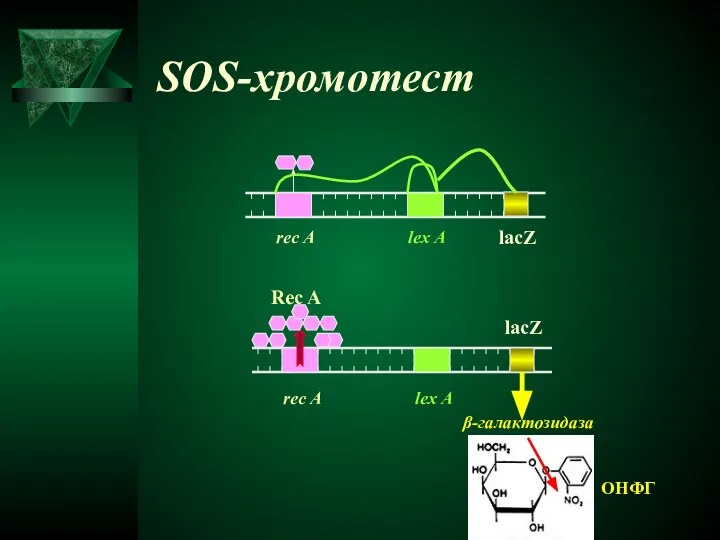
Слайд 37SOS-хромотест
rec A
lex A
rec A
lex A
Rec A
lacZ
β-галактозидаза
ОНФГ
lacZ
SOS-хромотест
rec A
lex A
rec A
lex A
Rec A
lacZ
β-галактозидаза
ОНФГ
lacZ
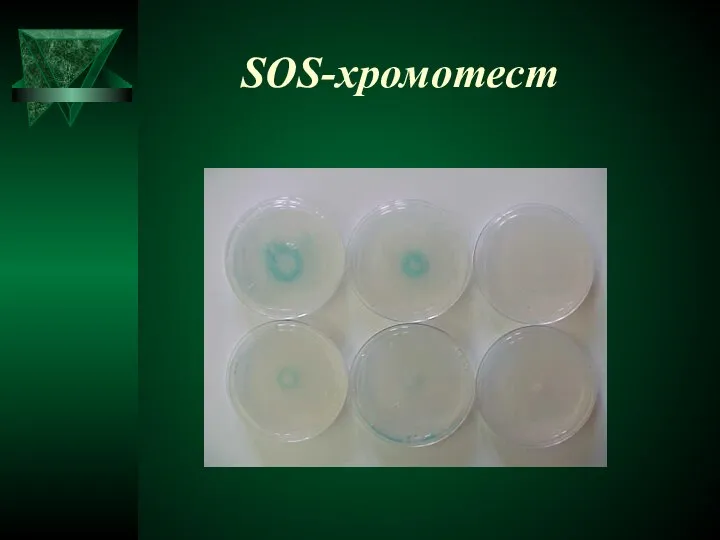
Слайд 38SOS-хромотест
SOS-хромотест
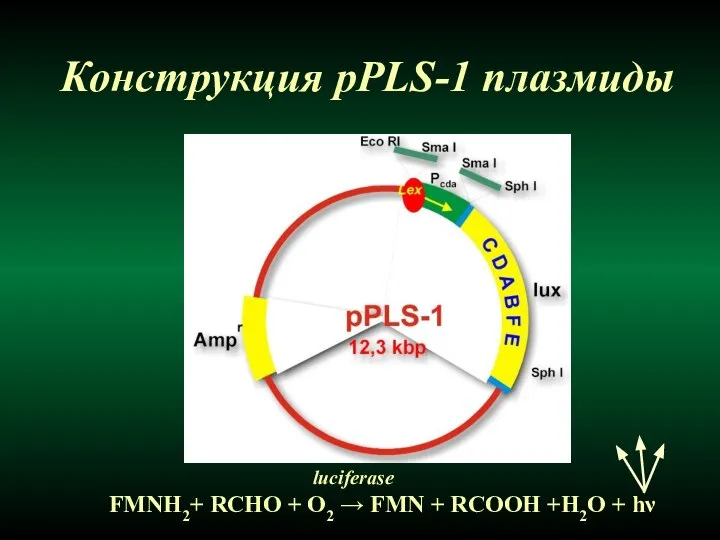
Слайд 39Конструкция pPLS-1 плазмиды
FMNH2+ RCHO + O2 → FMN + RCOOH +H2O +
Конструкция pPLS-1 плазмиды
FMNH2+ RCHO + O2 → FMN + RCOOH +H2O +
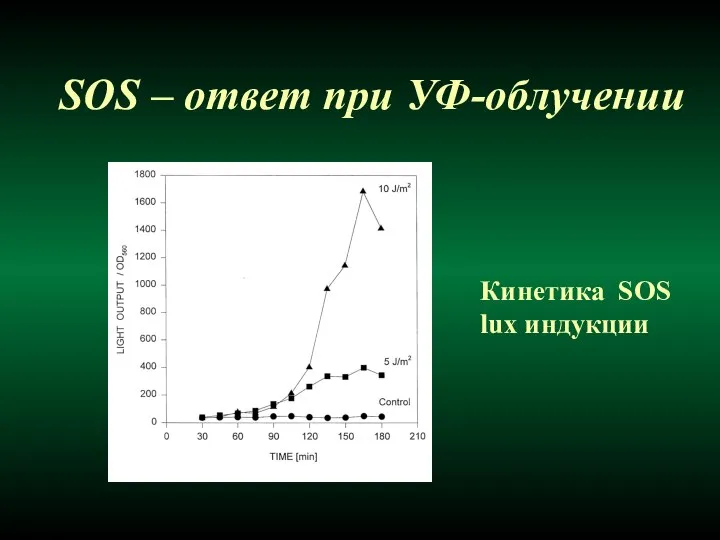
Слайд 40SOS – ответ при УФ-облучении
Кинетика SOS lux индукции
SOS – ответ при УФ-облучении
Кинетика SOS lux индукции
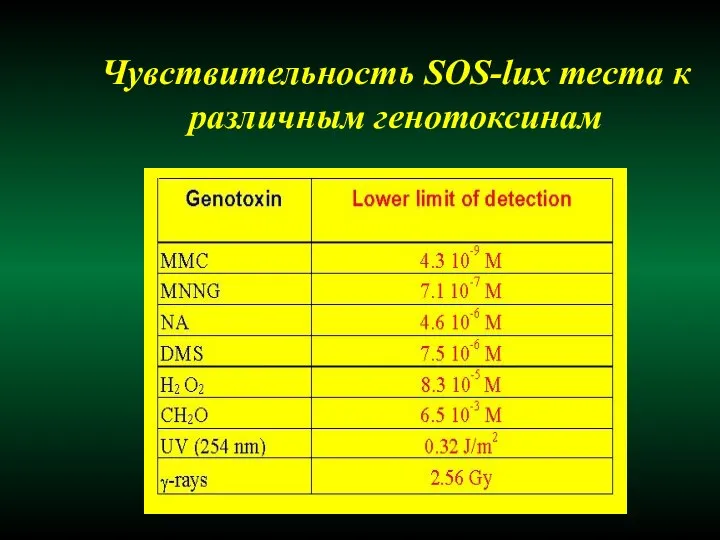
Слайд 41Чувствительность SOS-lux теста к различным генотоксинам
Чувствительность SOS-lux теста к различным генотоксинам
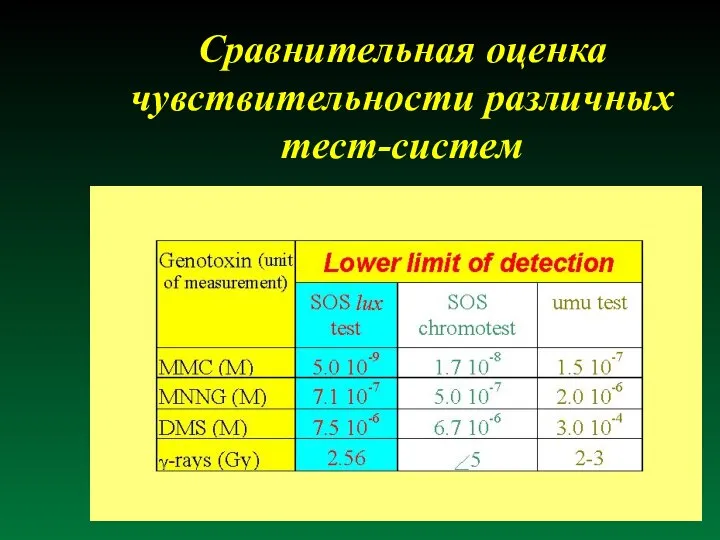
Слайд 42Сравнительная оценка чувствительности различных тест-систем
Сравнительная оценка чувствительности различных тест-систем
Слайд 43 фаг
λ
λ
фаг
λ
λ

Слайд 44Частица фага
Хромосома фага длиной около 50000 пар оснований находится в головке
Частица фага
Хромосома фага длиной около 50000 пар оснований находится в головке

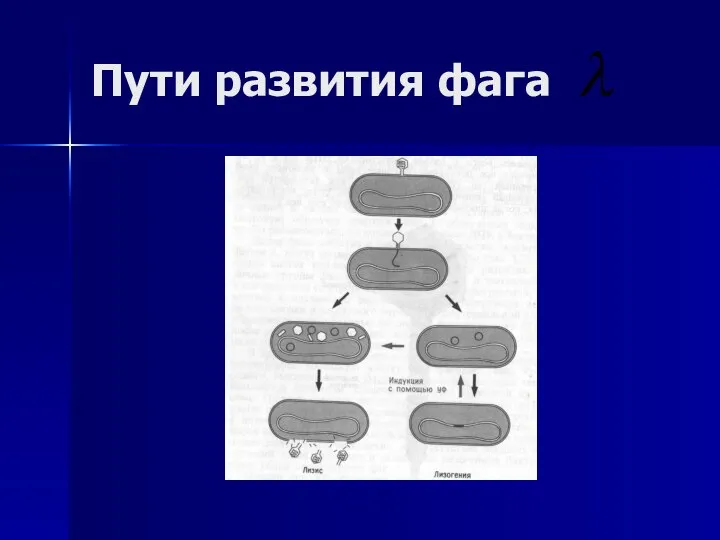
Слайд 45Пути развития фага
Пути развития фага
Слайд 46Репрессор
Репрессор-белок из 236 аминокислот.
Димер формируется главным образом за счет контактов между
Репрессор
Репрессор-белок из 236 аминокислот.
Димер формируется главным образом за счет контактов между

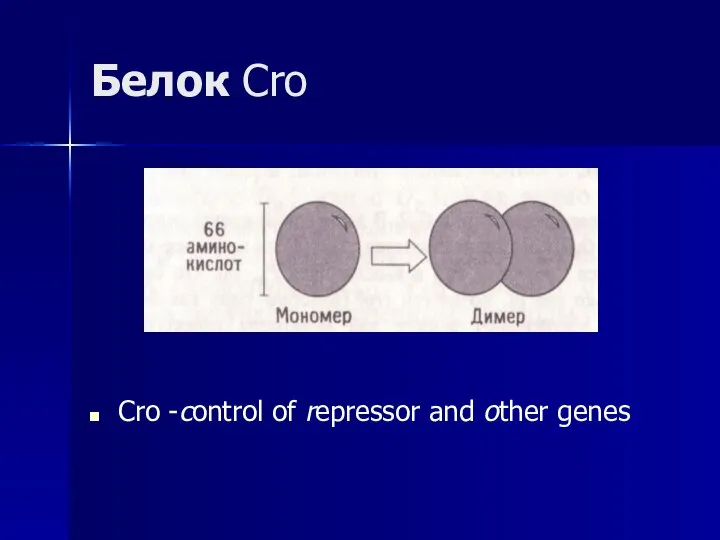
Слайд 47Белок Cro
Cro -control of repressor and other genes
Белок Cro
Cro -control of repressor and other genes
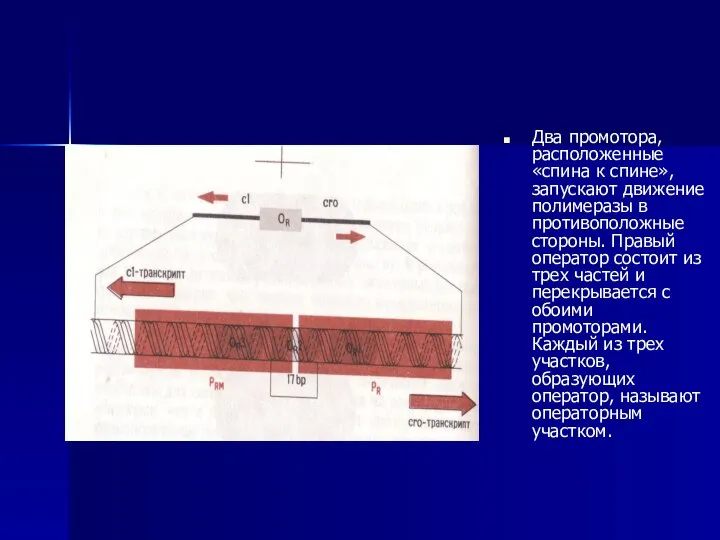
Слайд 48Два промотора,расположенные «спина к спине»,запускают движение полимеразы в противоположные стороны. Правый оператор
Два промотора,расположенные «спина к спине»,запускают движение полимеразы в противоположные стороны. Правый оператор
Слайд 49Действие репрессора и Cro
Действие репрессора и Cro

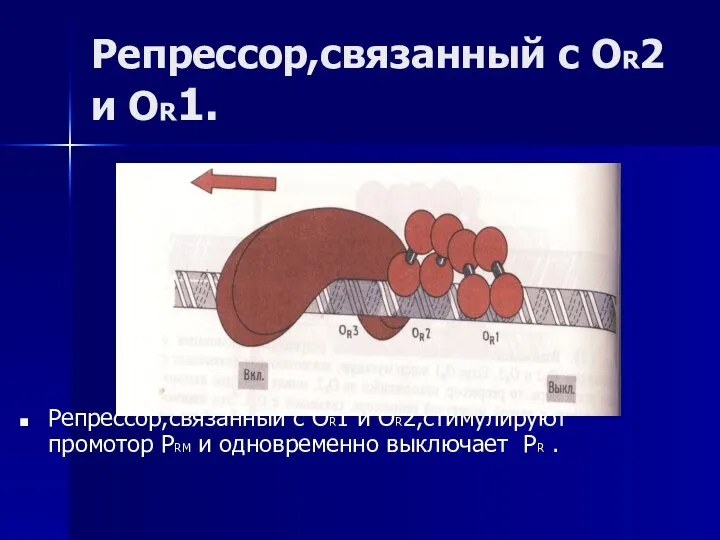
Слайд 50Репрессор,связанный с OR2 и OR1.
Репрессор,связанный с OR1 и OR2,стимулируют промотор PRM и
Репрессор,связанный с OR2 и OR1.
Репрессор,связанный с OR1 и OR2,стимулируют промотор PRM и
Слайд 51Как УФ-облучение изменяет поведение лизогенных клеток?
Как УФ-облучение изменяет поведение лизогенных клеток?

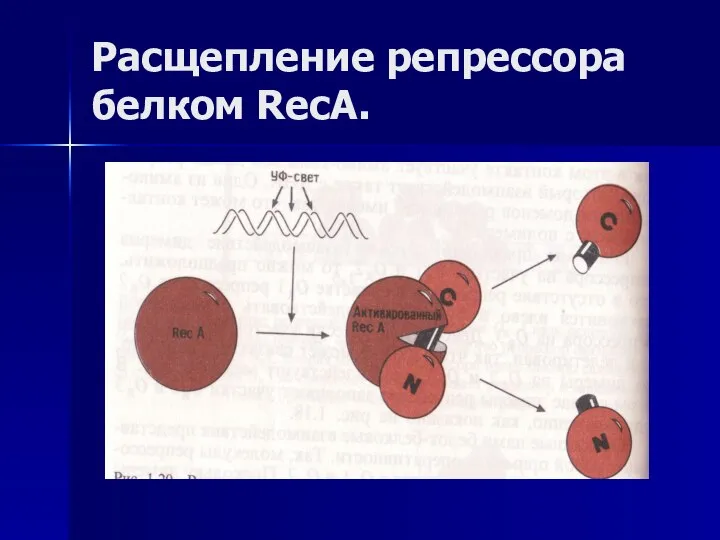
Слайд 52Расщепление репрессора белком RecA.
Расщепление репрессора белком RecA.
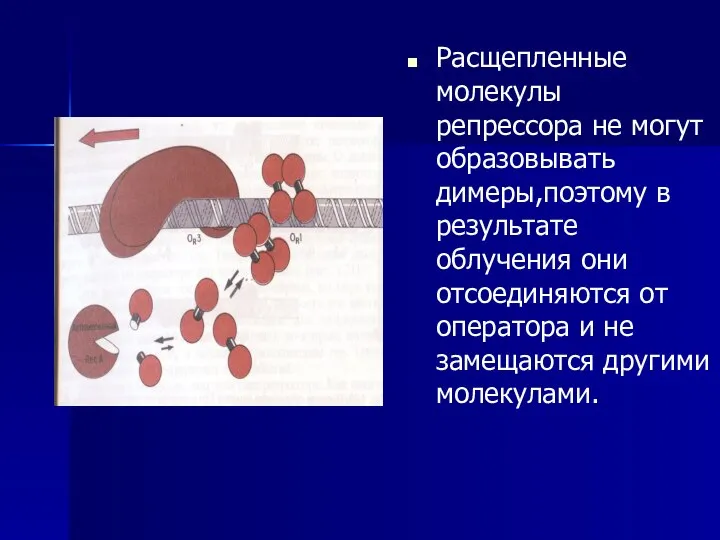
Слайд 53Расщепленные молекулы репрессора не могут образовывать димеры,поэтому в результате облучения они отсоединяются
Расщепленные молекулы репрессора не могут образовывать димеры,поэтому в результате облучения они отсоединяются
Слайд 54Связывание белка Cro с OR.
Димеры Cro независимо связываются с каждым из трех
Связывание белка Cro с OR.
Димеры Cro независимо связываются с каждым из трех

Слайд 56 Мутатест
(тест Эймса)
Мутация his G46
Клетки не растут на среде без гистидина
реверсия
Мутатест
(тест Эймса)
Мутация his G46
Клетки не растут на среде без гистидина
реверсия
 Позвоночный столб
Позвоночный столб Брюхоногие моллюски Выполнил : Севостьянов Дмитрий
Брюхоногие моллюски Выполнил : Севостьянов Дмитрий Разбор заданий ЕГЭ по биологии
Разбор заданий ЕГЭ по биологии Астра однолетняя
Астра однолетняя Удивительные растения
Удивительные растения Белок кинезин
Белок кинезин Нарцисс
Нарцисс Высотная поясность
Высотная поясность Дефекты зрения
Дефекты зрения Экологические системы
Экологические системы Иннервация тазового дна, синдром Алкока, синдром запирательного нерва
Иннервация тазового дна, синдром Алкока, синдром запирательного нерва Анатомическое строение листа
Анатомическое строение листа Высшие, или плацентарные звери: насекомоядные и рукокрылые, грызуны и зайцеобразные, хищные
Высшие, или плацентарные звери: насекомоядные и рукокрылые, грызуны и зайцеобразные, хищные Кошечки. Породы кошек
Кошечки. Породы кошек Покрытосеменные растения. Урок обобщения, повторения и контроля
Покрытосеменные растения. Урок обобщения, повторения и контроля Популяции. Изменчивость популяции
Популяции. Изменчивость популяции Біологічні основи запліднення та розвитку організму
Біологічні основи запліднення та розвитку організму Опасные растения и насекомые Крыма
Опасные растения и насекомые Крыма Сцепленное наследование и типы взаимодействия генов
Сцепленное наследование и типы взаимодействия генов Класс земноводные. Amphibia
Класс земноводные. Amphibia Джозеф Редьярд Кіплінг (1865-1936)
Джозеф Редьярд Кіплінг (1865-1936) Презентация на тему Глаз как орган зрения и оптическая система
Презентация на тему Глаз как орган зрения и оптическая система  Фотосинтез. Общая схема фотосинтеза
Фотосинтез. Общая схема фотосинтеза История открытия пенициллина
История открытия пенициллина Многообразие животного мира
Многообразие животного мира Презентация на тему Змеи
Презентация на тему Змеи  Общие закономерности физиологии и ее основные понятия
Общие закономерности физиологии и ее основные понятия Органы чувств. Орган слуха и равновесия
Органы чувств. Орган слуха и равновесия