- Катаболиз органических веществ на примере углеводов и жиров

Содержание
- 2. Этапы энергетического обмена
- 3. Роль печени и Цикл Кори
- 4. Классификация ферментов
- 5. Гликолиз или Брожение Схема гликолиза
- 6. Первая реакция Гексокиназа. Всего в организме существует 4 изоформы этого фермента. IV форма носит название Глюкокиназа
- 7. Вторая и Третья реакции Фосфоглюкозоизомераза катализирует обратимую изомеризацию глюкозо-6-фосфата (альдозы) во фруктозо-6-фосфат (кетозу) Механизм этой реакции
- 8. Четвертая реакция Существуют 2 класса альдолаз. Альдолазы класса I имеются у животных и растений, их работа
- 9. Пятая, Шестая и Седьмая реакции Количество NADH+ в клетке ( Механизм этой реакции схож с механизмом
- 10. Восьмая и Девятая реакции
- 11. Юбилейная Десятая реакция
- 12. Вовлечение других сахаров в гликолиз
- 13. Другие типы брожения
- 14. Брожение по типу клостридиевых, молочнокислых бактерий и дрожей.
- 15. Окислительное Декарбоксилирование Пирувата Цикл Кребса. Начало. Необратимые изменения
- 17. Цикл Трикарбоновых кислот. Цикл Кребса
- 18. Первая реакция
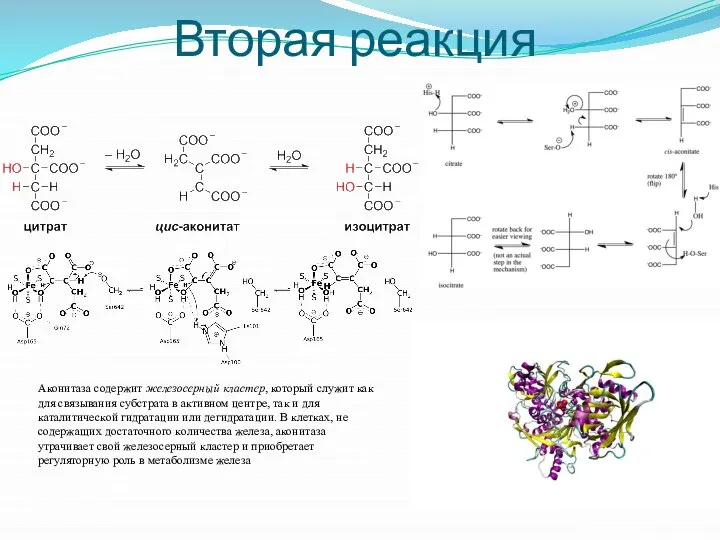
- 19. Вторая реакция Аконитаза содержит железосерный кластер, который служит как для связывания субстрата в активном центре, так
- 20. Третья и Четвертая реакция В клетках обнаружены две различных формы (изозима) изоцитратдегидрогеназы. Для функционирования одной из
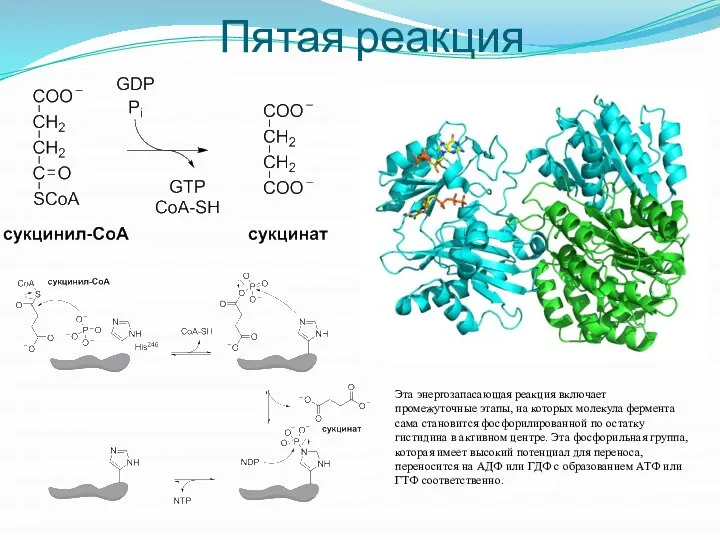
- 21. Пятая реакция Эта энергозапасающая реакция включает промежуточные этапы, на которых молекула фермента сама становится фосфорилированной по
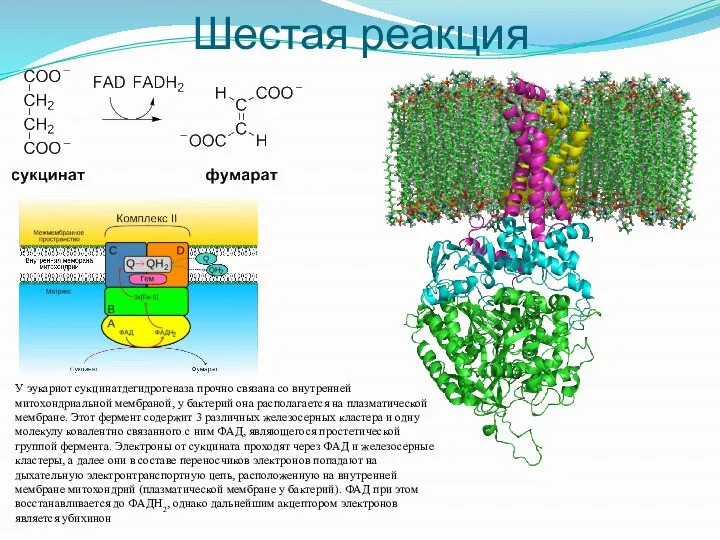
- 22. Шестая реакция У эукариот сукцинатдегидрогеназа прочно связана со внутренней митохондриальной мембраной, у бактерий она располагается на
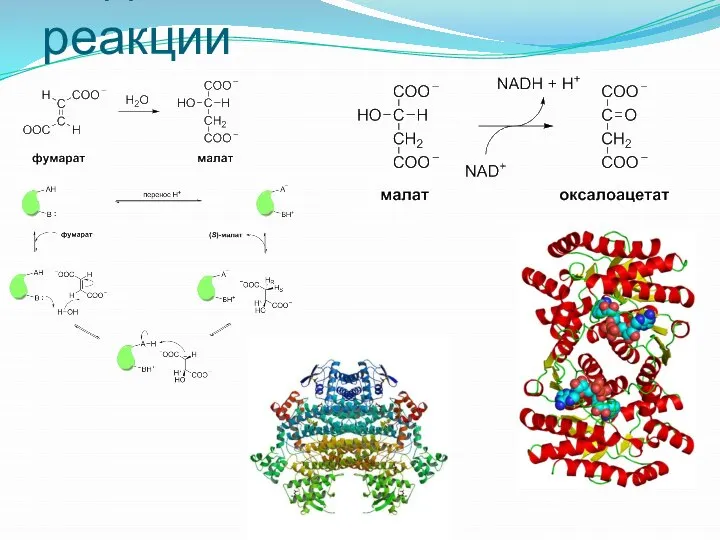
- 23. Седьмая и Восьмая реакции
- 24. Катаболиз Жиров и активация ЖК
- 25. β-окисление жирных кислот
- 26. β-окисление жирных кислот Окисление Жирных кислот с нечётным числом атомов углерода
- 27. Окисление ЖК с нечётным числом атомов углерода
- 28. α-Окисление жирных кислот в пероксисомах Рассмотрим α-окисление жирных кислот в животных клетках на примере фитановой кислоты.
- 29. Глиоксилатный цикл у Растений Механизм реакции катализируемой изоцитратлиазой и структура этого фермента
- 31. Скачать презентацию
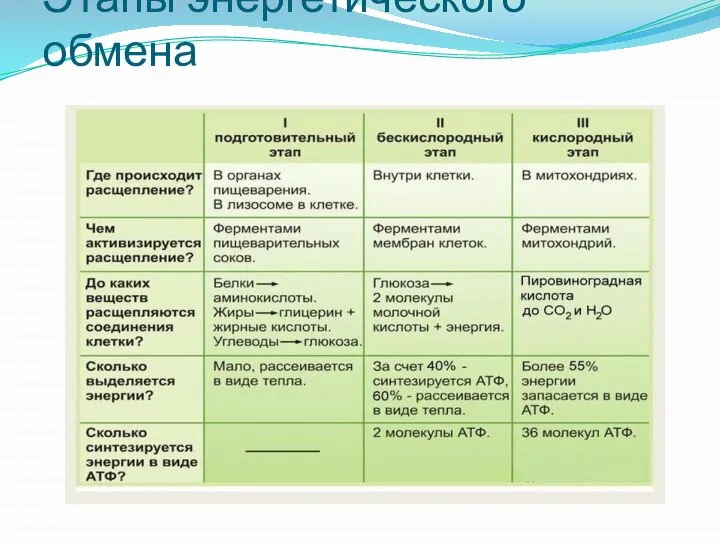
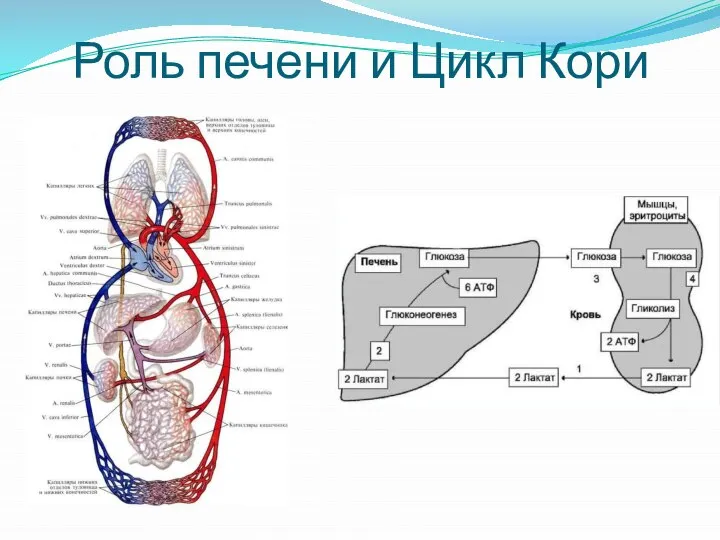
Слайд 3Роль печени и Цикл Кори
Роль печени и Цикл Кори
Слайд 4Классификация ферментов
Классификация ферментов

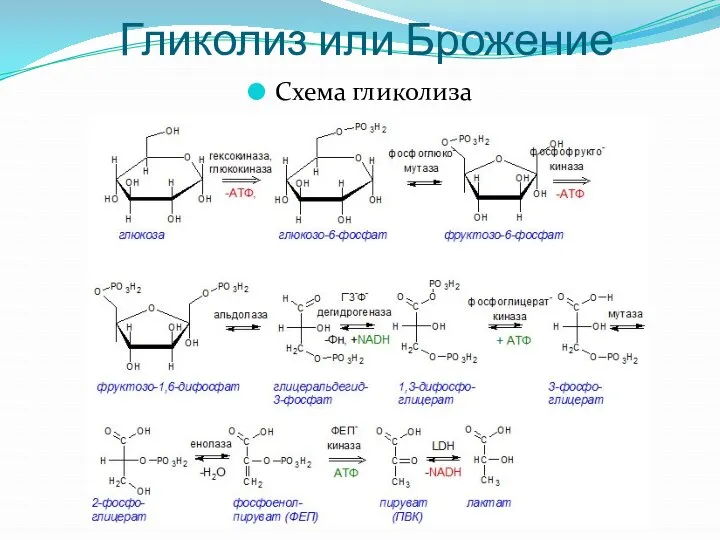
Слайд 5Гликолиз или Брожение
Схема гликолиза
Гликолиз или Брожение
Схема гликолиза
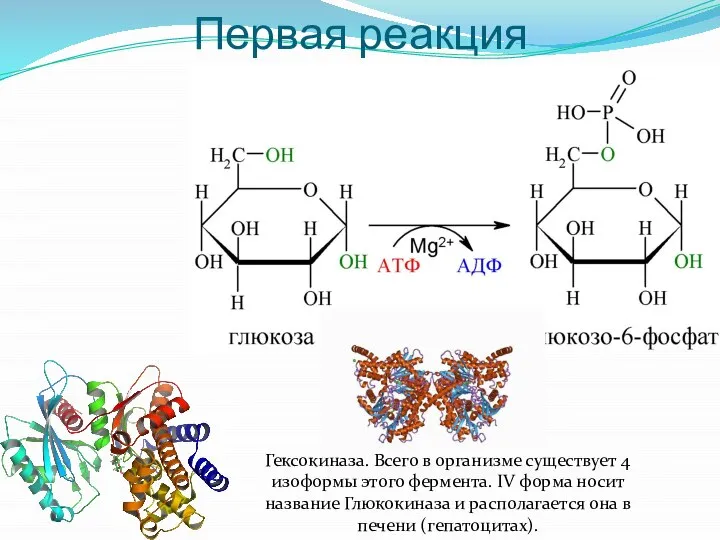
Слайд 6Первая реакция
Гексокиназа. Всего в организме существует 4 изоформы этого фермента. IV форма
Первая реакция
Гексокиназа. Всего в организме существует 4 изоформы этого фермента. IV форма
Слайд 7Вторая и Третья реакции
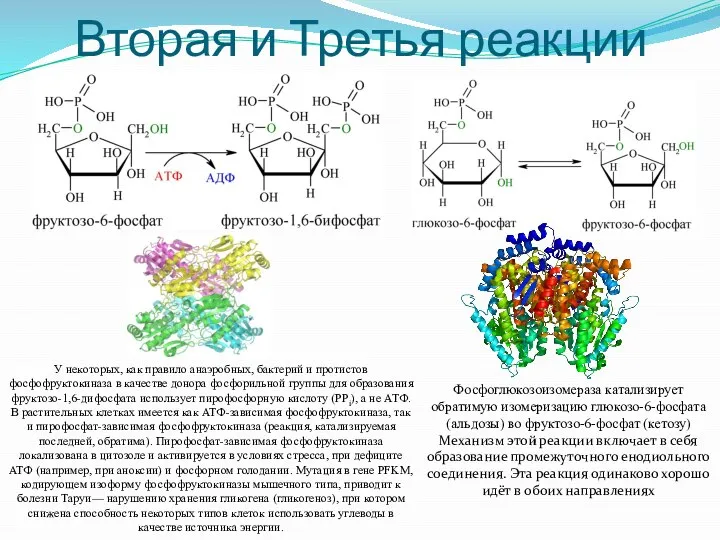
Фосфоглюкозоизомераза катализирует обратимую изомеризацию глюкозо-6-фосфата (альдозы) во фруктозо-6-фосфат (кетозу)
Механизм
Вторая и Третья реакции
Фосфоглюкозоизомераза катализирует обратимую изомеризацию глюкозо-6-фосфата (альдозы) во фруктозо-6-фосфат (кетозу)
Механизм
Слайд 8Четвертая реакция
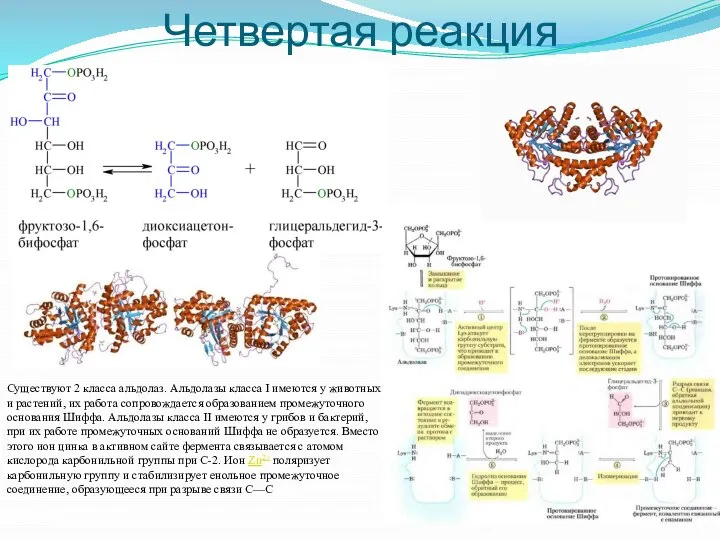
Существуют 2 класса альдолаз. Альдолазы класса I имеются у животных и
Четвертая реакция
Существуют 2 класса альдолаз. Альдолазы класса I имеются у животных и
Слайд 9Пятая, Шестая и Седьмая реакции
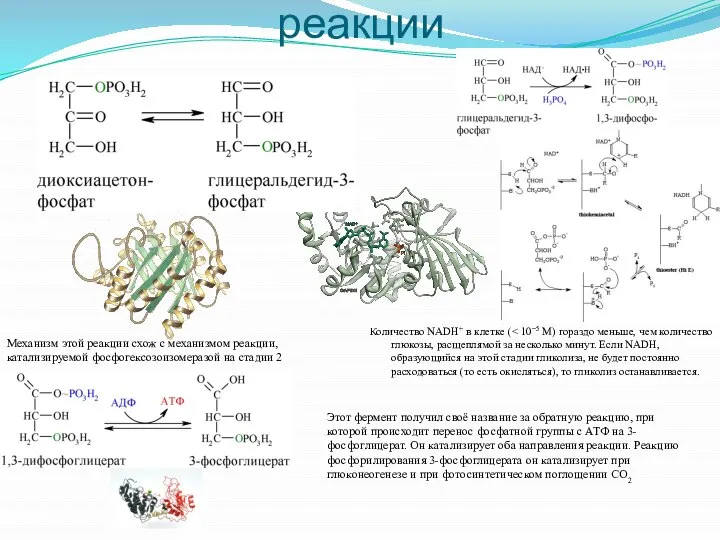
Количество NADH+ в клетке (< 10−5 М) гораздо
Пятая, Шестая и Седьмая реакции
Количество NADH+ в клетке (< 10−5 М) гораздо
Слайд 10Восьмая и Девятая реакции
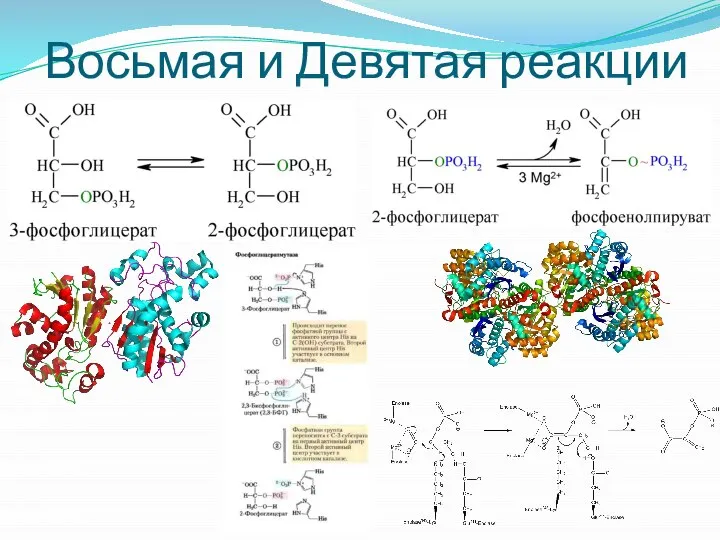
Восьмая и Девятая реакции
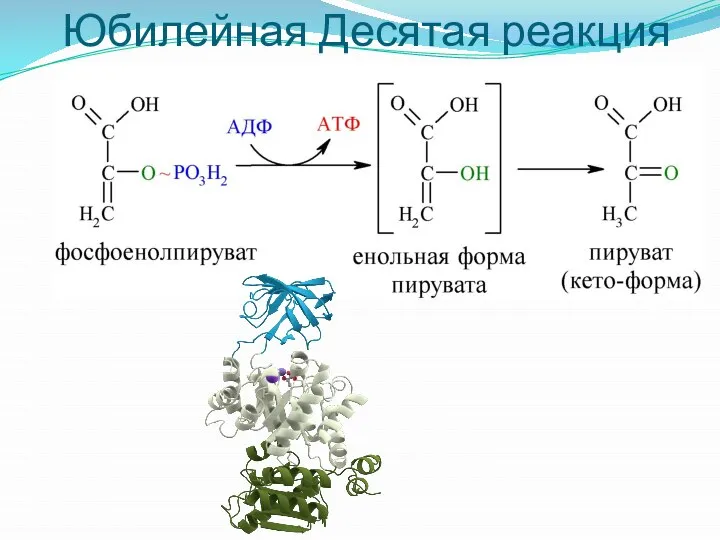
Слайд 11Юбилейная Десятая реакция
Юбилейная Десятая реакция
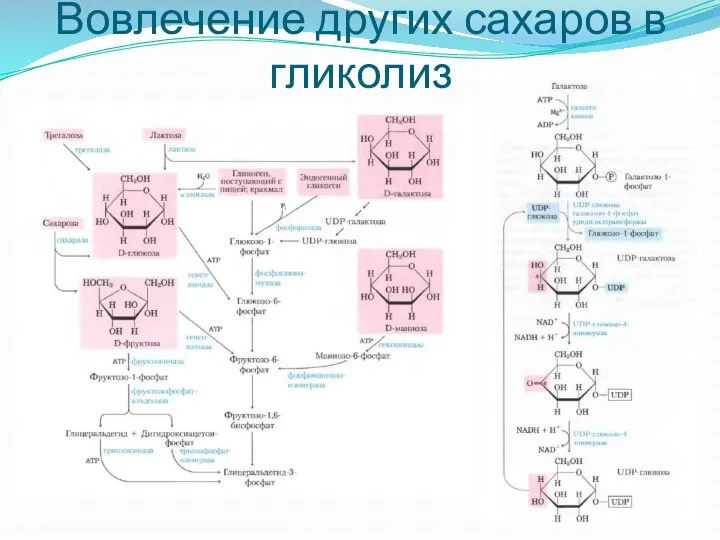
Слайд 12Вовлечение других сахаров в гликолиз
Вовлечение других сахаров в гликолиз
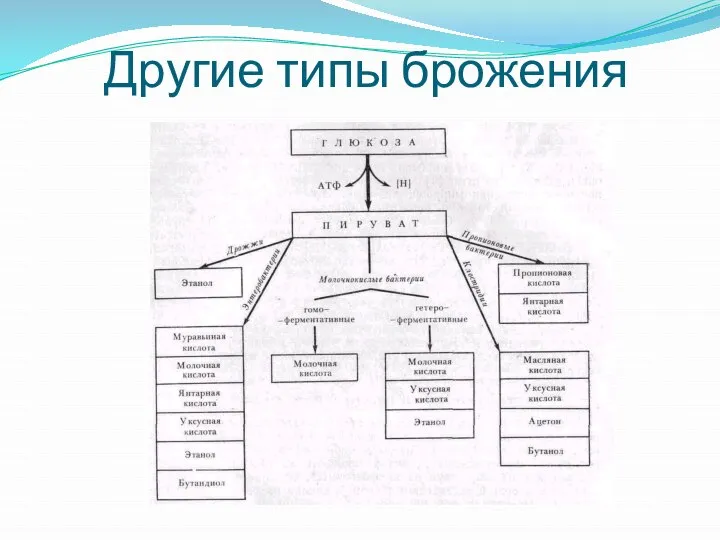
Слайд 13Другие типы брожения
Другие типы брожения
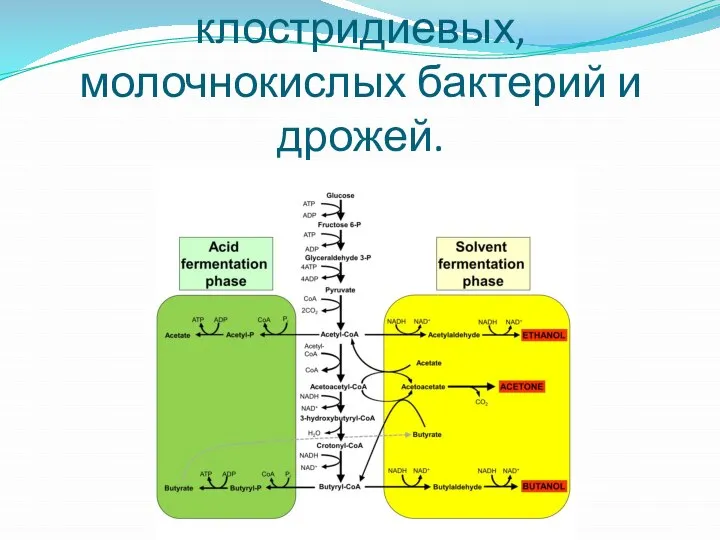
Слайд 14Брожение по типу клостридиевых, молочнокислых бактерий и дрожей.
Брожение по типу клостридиевых, молочнокислых бактерий и дрожей.
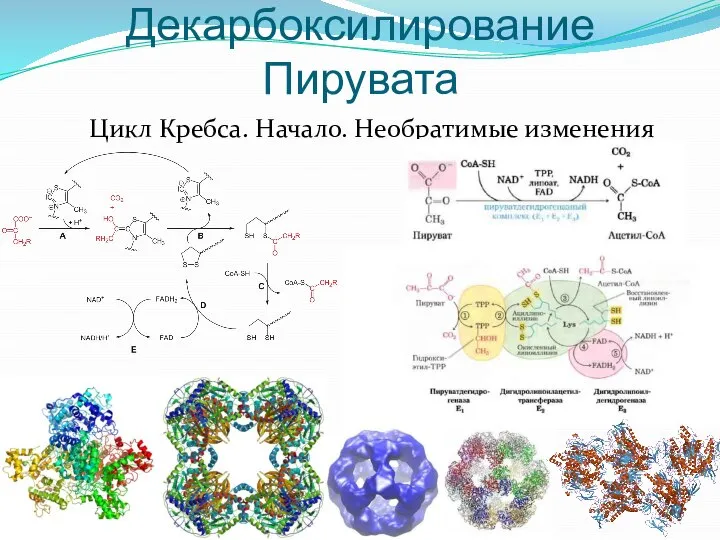
Слайд 15Окислительное Декарбоксилирование Пирувата
Цикл Кребса. Начало. Необратимые изменения
Окислительное Декарбоксилирование Пирувата
Цикл Кребса. Начало. Необратимые изменения
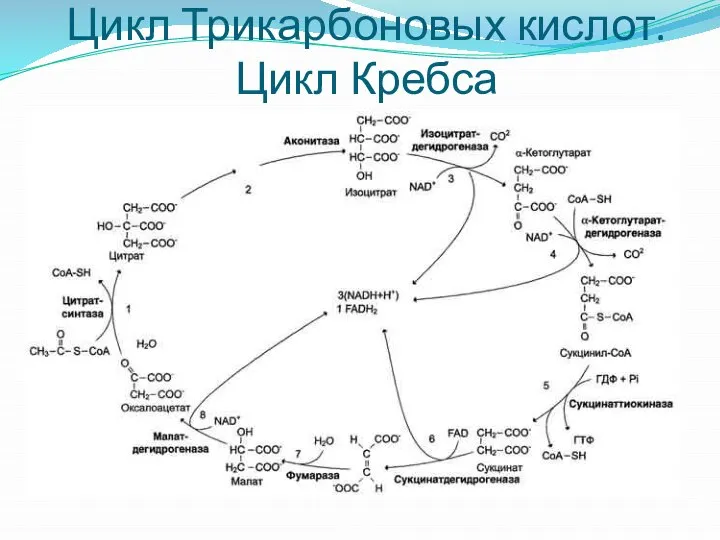
Слайд 17Цикл Трикарбоновых кислот. Цикл Кребса
Цикл Трикарбоновых кислот. Цикл Кребса
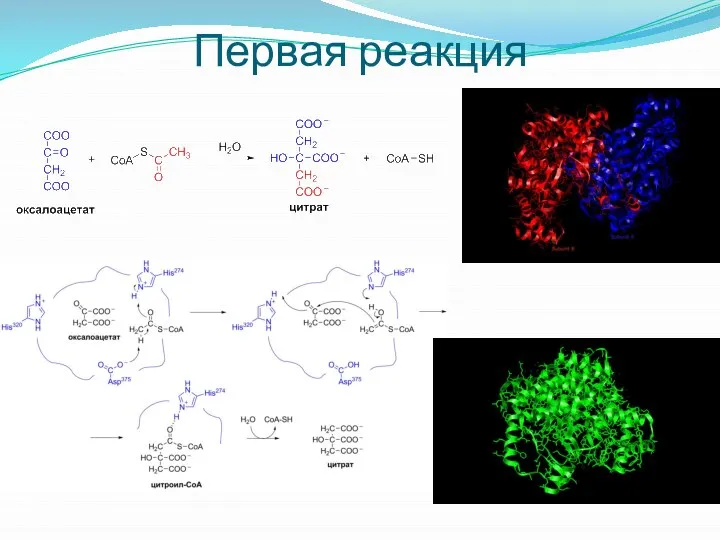
Слайд 18Первая реакция
Первая реакция
Слайд 19Вторая реакция
Аконитаза содержит железосерный кластер, который служит как для связывания субстрата в
Вторая реакция
Аконитаза содержит железосерный кластер, который служит как для связывания субстрата в
Слайд 20Третья и Четвертая реакция
В клетках обнаружены две различных формы (изозима) изоцитратдегидрогеназы. Для
Третья и Четвертая реакция
В клетках обнаружены две различных формы (изозима) изоцитратдегидрогеназы. Для

Слайд 21Пятая реакция
Эта энергозапасающая реакция включает промежуточные этапы, на которых молекула фермента сама
Пятая реакция
Эта энергозапасающая реакция включает промежуточные этапы, на которых молекула фермента сама
Слайд 22Шестая реакция
У эукариот сукцинатдегидрогеназа прочно связана со внутренней митохондриальной мембраной, у бактерий
Шестая реакция
У эукариот сукцинатдегидрогеназа прочно связана со внутренней митохондриальной мембраной, у бактерий
Слайд 23Седьмая и Восьмая реакции
Седьмая и Восьмая реакции
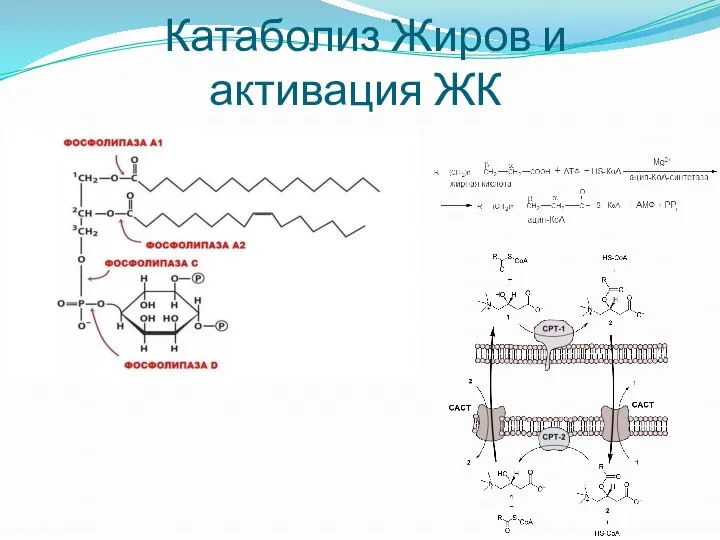
Слайд 24 Катаболиз Жиров и активация ЖК
Катаболиз Жиров и активация ЖК
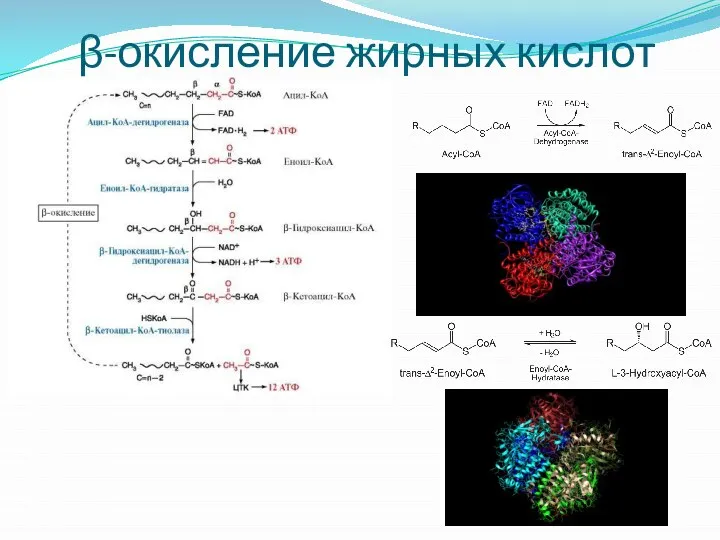
Слайд 25β-окисление жирных кислот
β-окисление жирных кислот
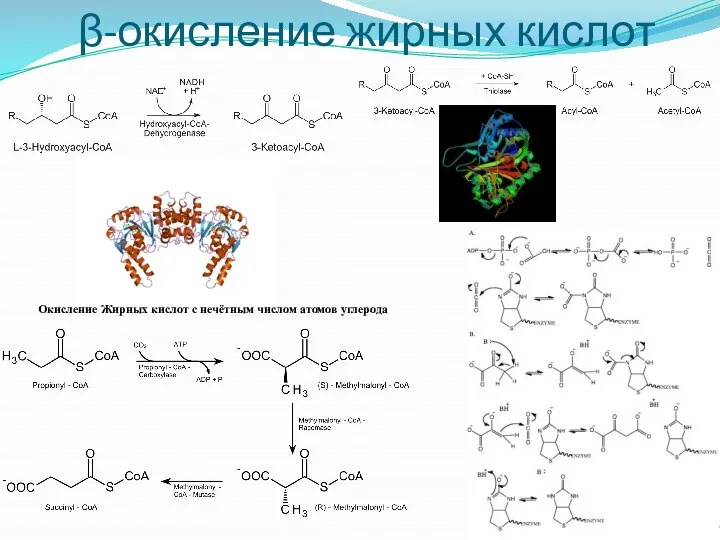
Слайд 26β-окисление жирных кислот
Окисление Жирных кислот с нечётным числом атомов углерода
β-окисление жирных кислот
Окисление Жирных кислот с нечётным числом атомов углерода
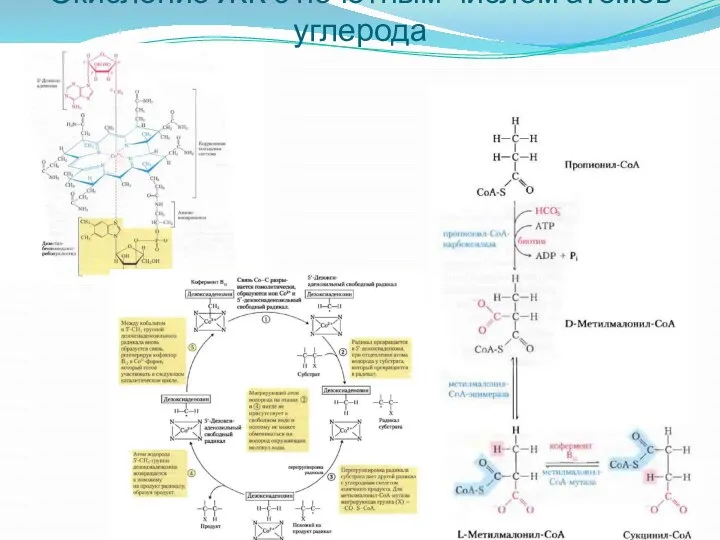
Слайд 27Окисление ЖК с нечётным числом атомов углерода
Окисление ЖК с нечётным числом атомов углерода
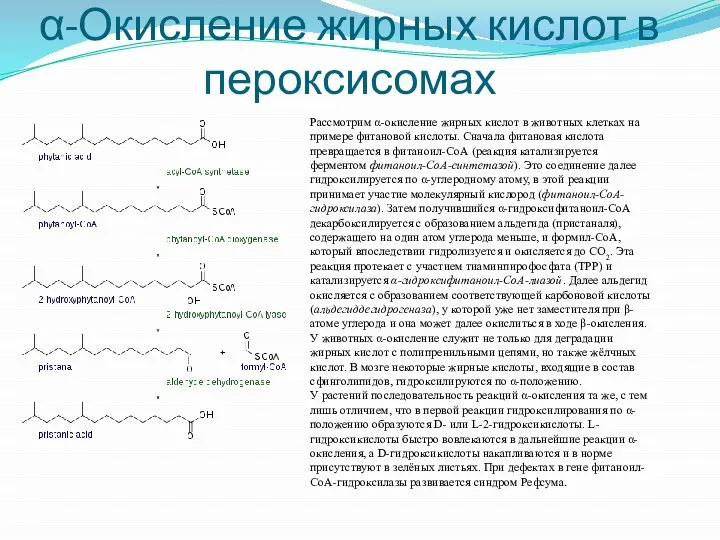
Слайд 28α-Окисление жирных кислот в пероксисомах
Рассмотрим α-окисление жирных кислот в животных клетках на
α-Окисление жирных кислот в пероксисомах
Рассмотрим α-окисление жирных кислот в животных клетках на
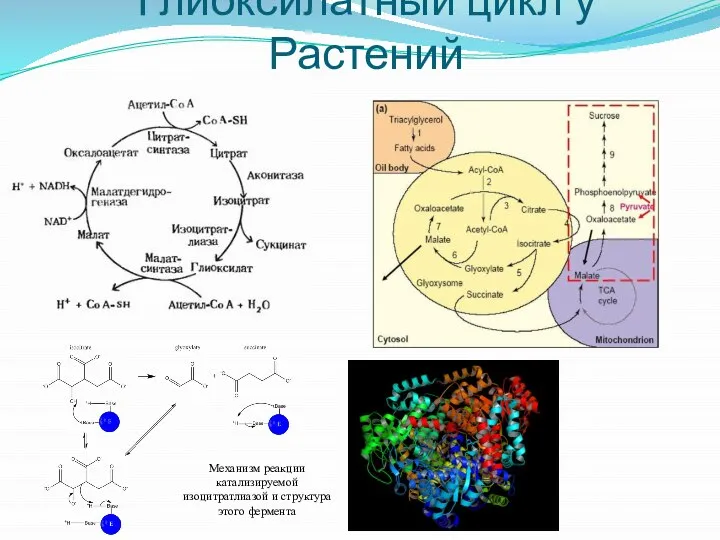
Слайд 29Глиоксилатный цикл у Растений
Механизм реакции катализируемой изоцитратлиазой и структура этого фермента
Глиоксилатный цикл у Растений
Механизм реакции катализируемой изоцитратлиазой и структура этого фермента
 Зимующие птицы
Зимующие птицы Опорно-двигательная система. Строение кости
Опорно-двигательная система. Строение кости Методика работы с микроскопом. Биологический рисунок
Методика работы с микроскопом. Биологический рисунок Весёлая биология. Викторина
Весёлая биология. Викторина Репродукция. (Тема 6)
Репродукция. (Тема 6) Кокетльові господарочки
Кокетльові господарочки Отряд хоботные
Отряд хоботные Развитие жизни в архее и протерозое
Развитие жизни в архее и протерозое Кроманьонцы. Этимология
Кроманьонцы. Этимология Отряды динозавров
Отряды динозавров חיות בעולם
חיות בעולם Класс Паукообразные. 7 класс
Класс Паукообразные. 7 класс Козлов Н.А. Общая гистология. Ткани домашних млекопитающих животных. 2004
Козлов Н.А. Общая гистология. Ткани домашних млекопитающих животных. 2004 Capybara
Capybara Вирусы. История открытия вирусов
Вирусы. История открытия вирусов стафилокок (1)
стафилокок (1) Внутреннее строение земноводных. Размножение. Развитие
Внутреннее строение земноводных. Размножение. Развитие Презентация на тему Оплодотворение у цветковых растений
Презентация на тему Оплодотворение у цветковых растений  Вегетативное размножение (стеблевое)
Вегетативное размножение (стеблевое)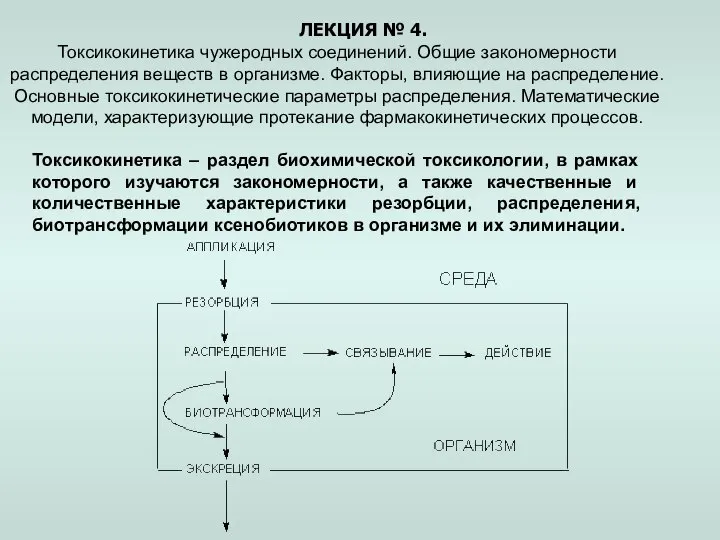 Токсикокинетика чужеродных соединений. Закономерности распределения веществ в организме. (Лекция 4)
Токсикокинетика чужеродных соединений. Закономерности распределения веществ в организме. (Лекция 4) Магнолия. Наша Миссия
Магнолия. Наша Миссия Деление клетки 5 класс
Деление клетки 5 класс Голо- и Покрытосеменные
Голо- и Покрытосеменные Пектиновые вещества
Пектиновые вещества Органы растений
Органы растений Презентация на тему Первый и второй законы Менделя
Презентация на тему Первый и второй законы Менделя  Классификация животных
Классификация животных Класс Пресмыкающиеся
Класс Пресмыкающиеся