- Самонесовместимость. Лекция 8

Содержание
- 2. ТИПЫ НЕСОВМЕСТИМОСТИ Три системы охарактеризовано на молекулярном уровне: SSI – спорофитная самонесовместимость (крестоцветные) S-гликопротеиновый (маковые) SРНКазный
- 3. ГСН/ССН Спорофитная СН: фенотип пыльцы определяется геномом родителя (спорофита) Гаметофитная СН: фенотип пыльцы определяется гаплоидным геномом
- 4. Гаметофитная самонесовместимость Рост пыльцы подавляется только тогда, когда она вошла в проводниковый тракт столбика. А на
- 5. Гаметофитная SI у мака Две S-детерминанты S-локуса мака уже достаточно хорошо изучены: PrpS кодирует трансмембранный белок
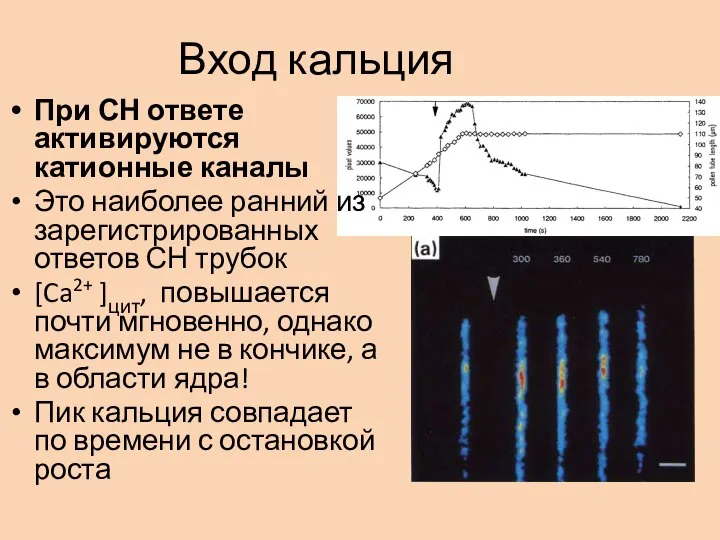
- 6. Вход кальция При СН ответе активируются катионные каналы Это наиболее ранний из зарегистрированных ответов СН трубок
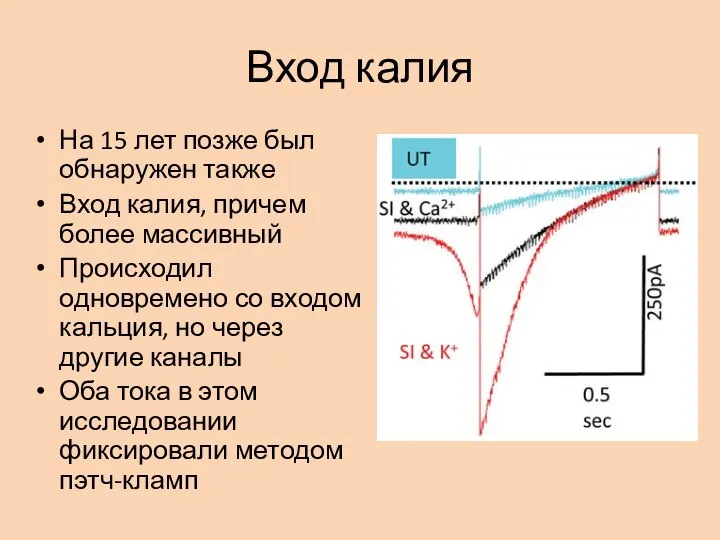
- 7. Вход калия На 15 лет позже был обнаружен также Вход калия, причем более массивный Происходил одновремено
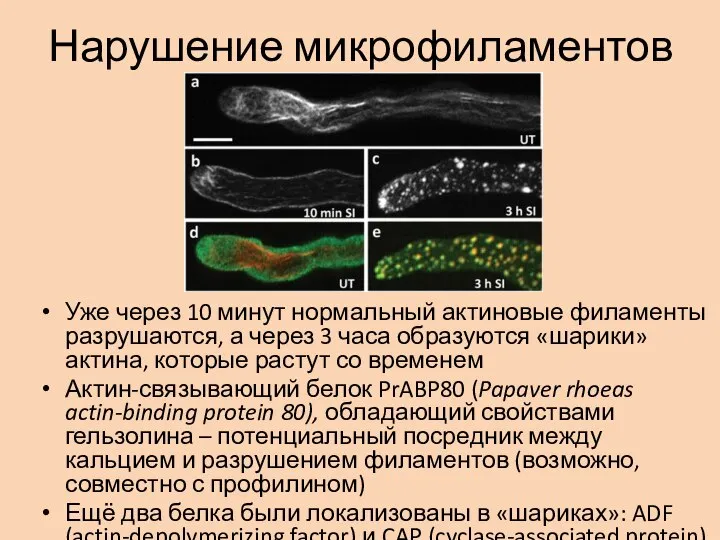
- 8. Нарушение микрофиламентов Уже через 10 минут нормальный актиновые филаменты разрушаются, а через 3 часа образуются «шарики»
- 9. Нарушение микротрубочек Микротрубочки в трубке полностью разрушаются уже через 1 минуту! В генеративной клетке МТ остаются
- 10. АФК – ангелы смерти Через 5 минут после индукции СН происходит быстрое транзиентное повышение цитозольных АФК
- 11. ROS NO Ca2+
- 12. Резюме по маку PrsS and PrpS узнают друг друга → Вход Ca2+ и сопутствующее повышение [Ca2+]цит
- 13. Модификация цитоскелета: деполимеризация актиновых филаментов и образование «шариков» Активация MAPK (mitogen-activated protein kinase) p56 Закисление цитозоля
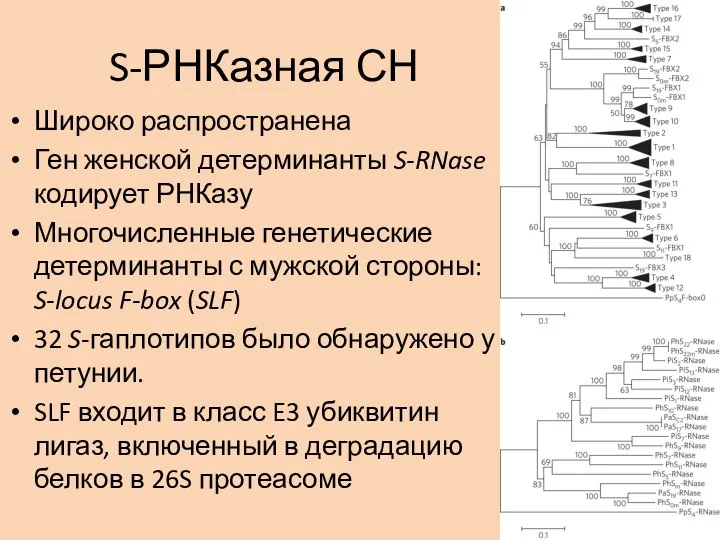
- 14. S-РНКазная СН Широко распространена Ген женской детерминанты S-RNase кодирует РНКазу Многочисленные генетические детерминанты с мужской стороны:
- 15. Партнер или паразит?! Ученые заметили, что коэволюция женских и мужских детерминант идёт разными путями (у пасленовых).
- 16. Каков же паттерн? Sx-РНКаза является мишенью для SLFn если Sx-аллель SLFn изменён или удалён. Этот упрощённый
- 17. Так думали раньше… Считали, что транспорт S-РНКаз в трубку специфичный, и «несвои» просто не заходят Но
- 18. S-РНКаза S-РНКаза синтезируется в проводниковом тракте , пока пыльцевые трубки растут через столбик. Трубки поглощают все
- 19. S-РНКаза (продолжение) Этот фермент деградирует рРНК в самонесовместимой пыльце При потере РНКазной активности СН не работает
- 20. Более ранние модели Предполагали, что узнается своя РНКаза и не деградирует, а начинает своё чёрное дело
- 21. Чужие SРНКазы «не узнаются» и отправляются на деградацию в протеасому Своя SРНКаза узнаётся рецептором SLF (S-locus
- 22. Новая модель: коллективное узнавание Многочисленные SLF белки, которые производит пыльца данного S-гаплотипа, вместе узнают и дезактивируют
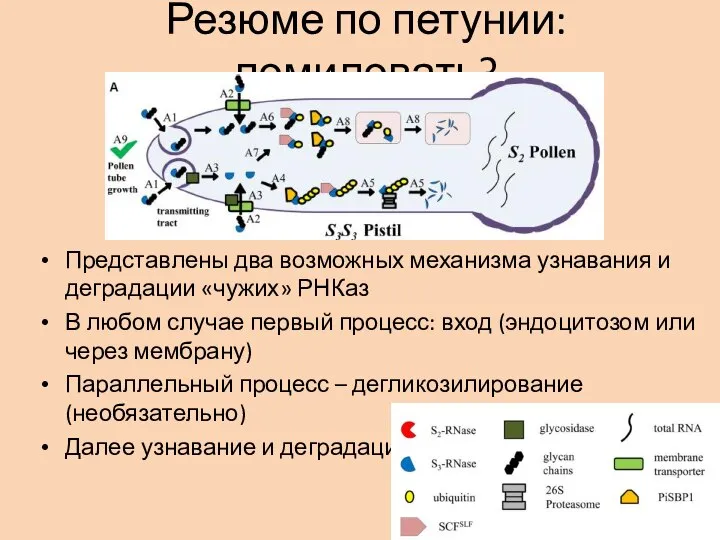
- 23. Резюме по петунии: помиловать? Представлены два возможных механизма узнавания и деградации «чужих» РНКаз В любом случае
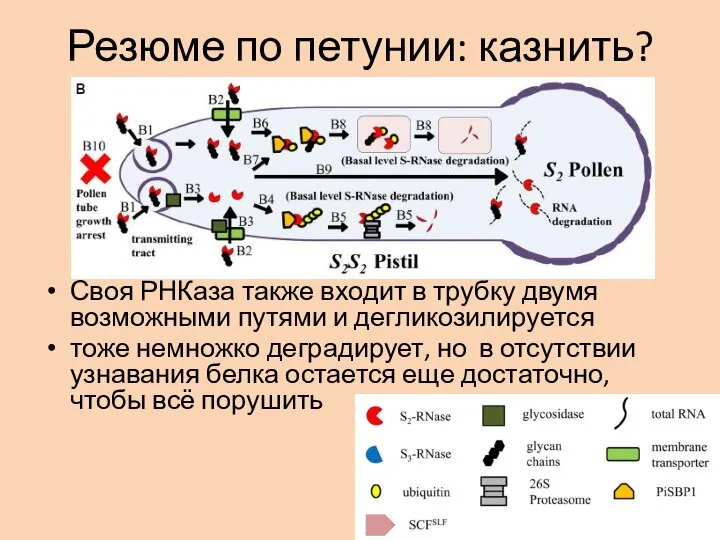
- 24. Резюме по петунии: казнить? Своя РНКаза также входит в трубку двумя возможными путями и дегликозилируется тоже
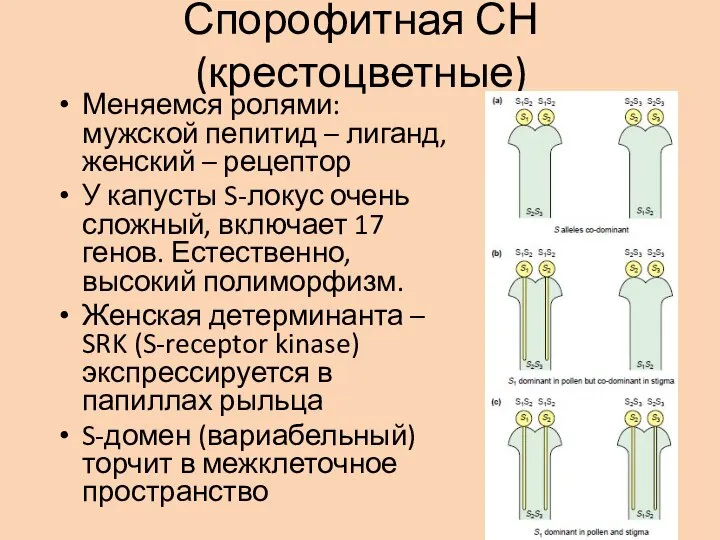
- 25. Спорофитная СН (крестоцветные) Меняемся ролями: мужской пепитид – лиганд, женский – рецептор У капусты S-локус очень
- 26. Мужская детерминанта – SCR/SPP (S-locus cystein-rich protein, S-locus pollen protein) Полиморфизм мужской детерминанты выше Она экспрессируется
- 27. Cубклеточная локализация SRK SRK обнаружены в плазмалемме и эндосомах у капусты Присутствует на ПМ В цитоплазме
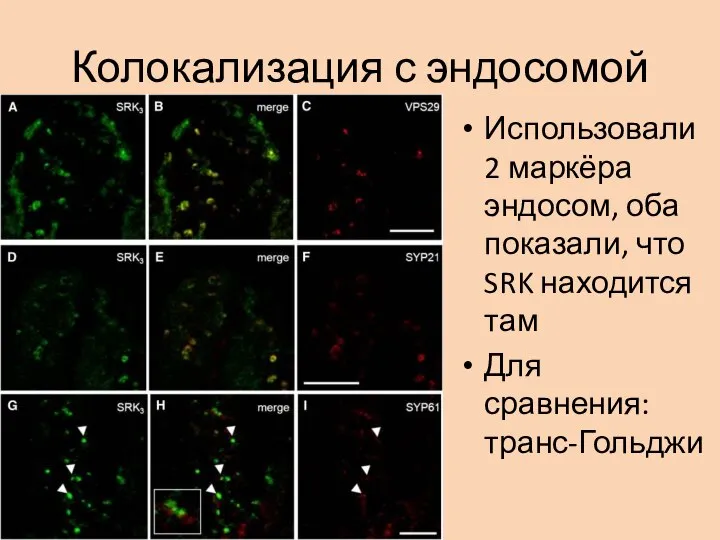
- 28. Колокализация с эндосомой Использовали 2 маркёра эндосом, оба показали, что SRK находится там Для сравнения: транс-Гольджи
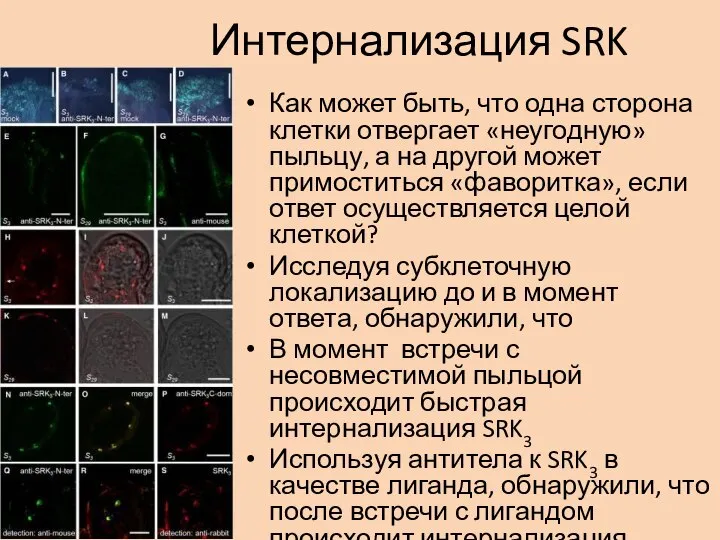
- 29. Интернализация SRK Как может быть, что одна сторона клетки отвергает «неугодную» пыльцу, а на другой может
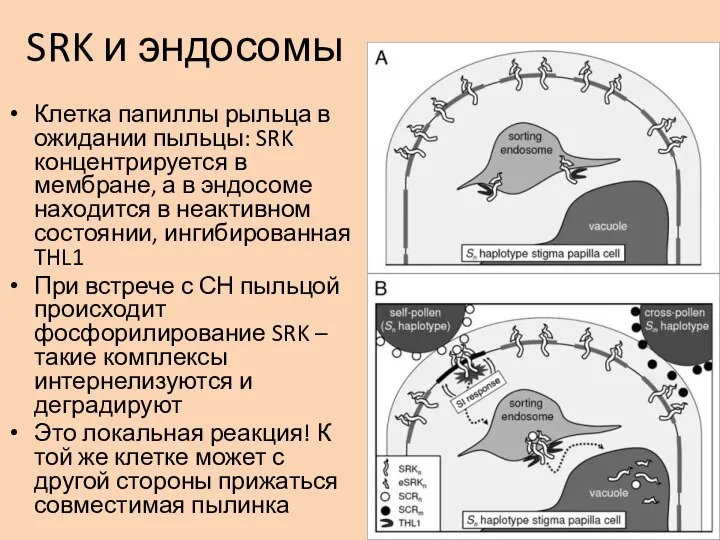
- 30. SRK и эндосомы Клетка папиллы рыльца в ожидании пыльцы: SRK концентрируется в мембране, а в эндосоме
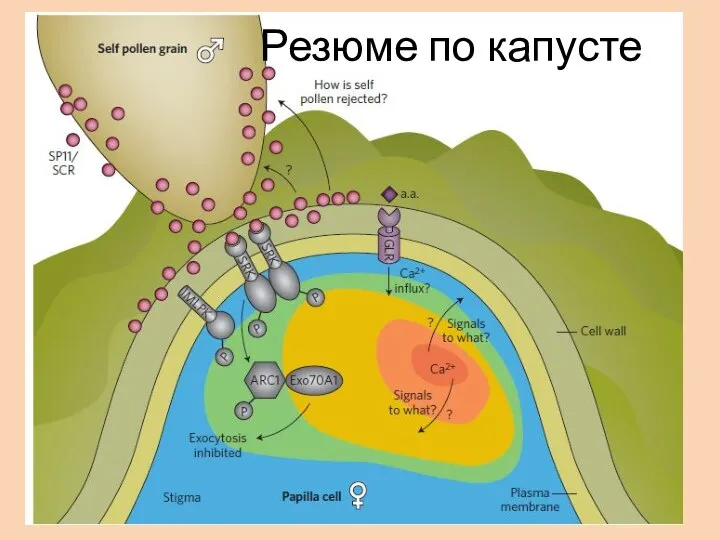
- 31. Резюме по капусте
- 33. Скачать презентацию
Слайд 2ТИПЫ НЕСОВМЕСТИМОСТИ
Три системы охарактеризовано на молекулярном уровне:
SSI – спорофитная самонесовместимость (крестоцветные)
S-гликопротеиновый (маковые)
SРНКазный
ТИПЫ НЕСОВМЕСТИМОСТИ
Три системы охарактеризовано на молекулярном уровне:
SSI – спорофитная самонесовместимость (крестоцветные)
S-гликопротеиновый (маковые)
SРНКазный
Слайд 3ГСН/ССН
Спорофитная СН: фенотип пыльцы определяется геномом родителя (спорофита)
Гаметофитная СН: фенотип пыльцы определяется
ГСН/ССН
Спорофитная СН: фенотип пыльцы определяется геномом родителя (спорофита)
Гаметофитная СН: фенотип пыльцы определяется

Слайд 4Гаметофитная самонесовместимость
Рост пыльцы подавляется только тогда, когда она вошла в проводниковый тракт
Гаметофитная самонесовместимость
Рост пыльцы подавляется только тогда, когда она вошла в проводниковый тракт

Слайд 5Гаметофитная SI у мака
Две S-детерминанты S-локуса мака уже достаточно хорошо изучены:
PrpS кодирует
Гаметофитная SI у мака
Две S-детерминанты S-локуса мака уже достаточно хорошо изучены:
PrpS кодирует

Слайд 6Вход кальция
При СН ответе активируются катионные каналы
Это наиболее ранний из зарегистрированных ответов
Вход кальция
При СН ответе активируются катионные каналы
Это наиболее ранний из зарегистрированных ответов
Слайд 7Вход калия
На 15 лет позже был обнаружен также
Вход калия, причем более массивный
Происходил
Вход калия
На 15 лет позже был обнаружен также
Вход калия, причем более массивный
Происходил
Слайд 8Нарушение микрофиламентов
Уже через 10 минут нормальный актиновые филаменты разрушаются, а через 3
Нарушение микрофиламентов
Уже через 10 минут нормальный актиновые филаменты разрушаются, а через 3
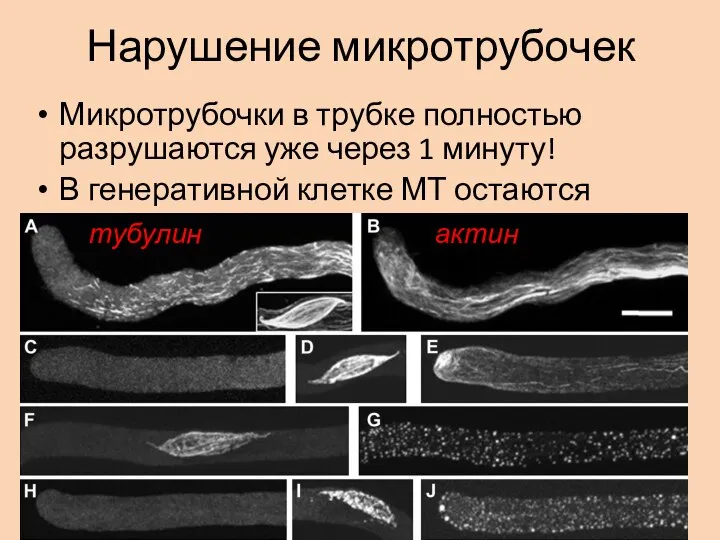
Слайд 9Нарушение микротрубочек
Микротрубочки в трубке полностью разрушаются уже через 1 минуту!
В генеративной клетке
Нарушение микротрубочек
Микротрубочки в трубке полностью разрушаются уже через 1 минуту!
В генеративной клетке
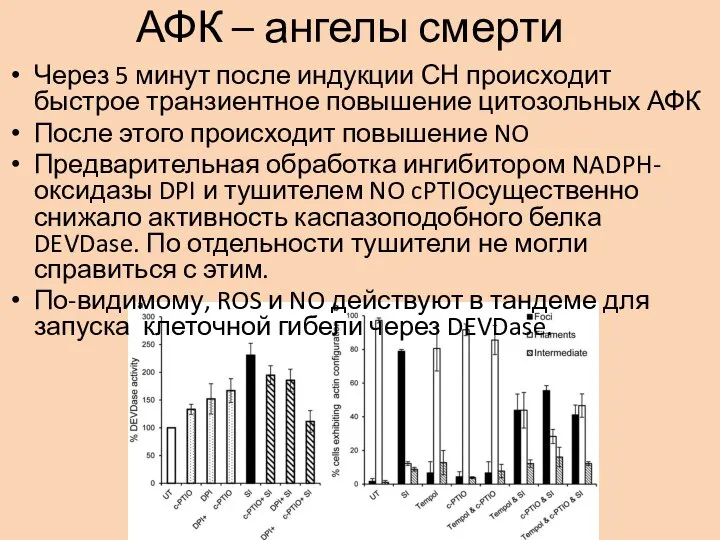
Слайд 10АФК – ангелы смерти
Через 5 минут после индукции СН происходит быстрое транзиентное
АФК – ангелы смерти
Через 5 минут после индукции СН происходит быстрое транзиентное
Слайд 11ROS
NO
Ca2+
ROS
NO
Ca2+

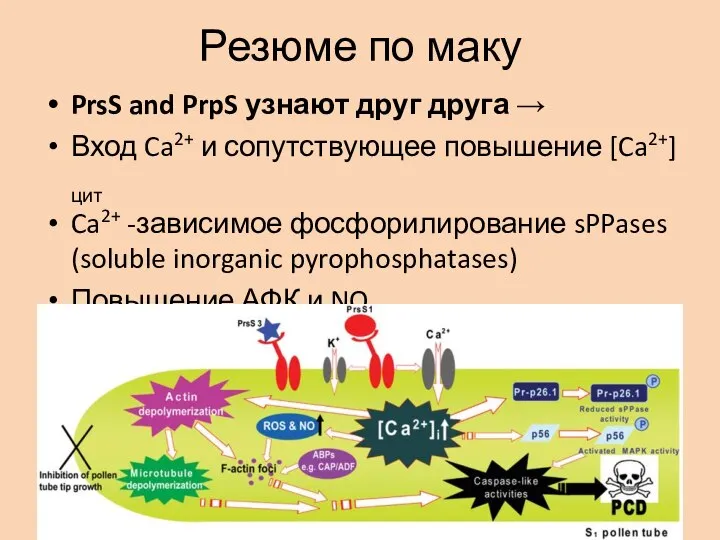
Слайд 12Резюме по маку
PrsS and PrpS узнают друг друга →
Вход Ca2+ и сопутствующее
Резюме по маку
PrsS and PrpS узнают друг друга →
Вход Ca2+ и сопутствующее
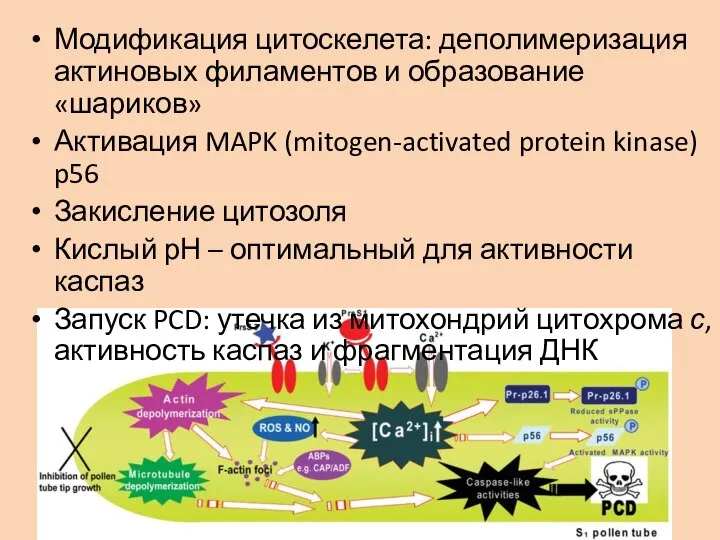
Слайд 13Модификация цитоскелета: деполимеризация актиновых филаментов и образование «шариков»
Активация MAPK (mitogen-activated protein kinase)
Модификация цитоскелета: деполимеризация актиновых филаментов и образование «шариков»
Активация MAPK (mitogen-activated protein kinase)
Слайд 14S-РНКазная СН
Широко распространена
Ген женской детерминанты S-RNase кодирует РНКазу
Многочисленные генетические детерминанты с мужской
S-РНКазная СН
Широко распространена
Ген женской детерминанты S-RNase кодирует РНКазу
Многочисленные генетические детерминанты с мужской
Слайд 15Партнер или паразит?!
Ученые заметили, что коэволюция женских и мужских детерминант идёт разными
Партнер или паразит?!
Ученые заметили, что коэволюция женских и мужских детерминант идёт разными

Слайд 16Каков же паттерн?
Sx-РНКаза является мишенью для SLFn если Sx-аллель SLFn изменён или
Каков же паттерн?
Sx-РНКаза является мишенью для SLFn если Sx-аллель SLFn изменён или

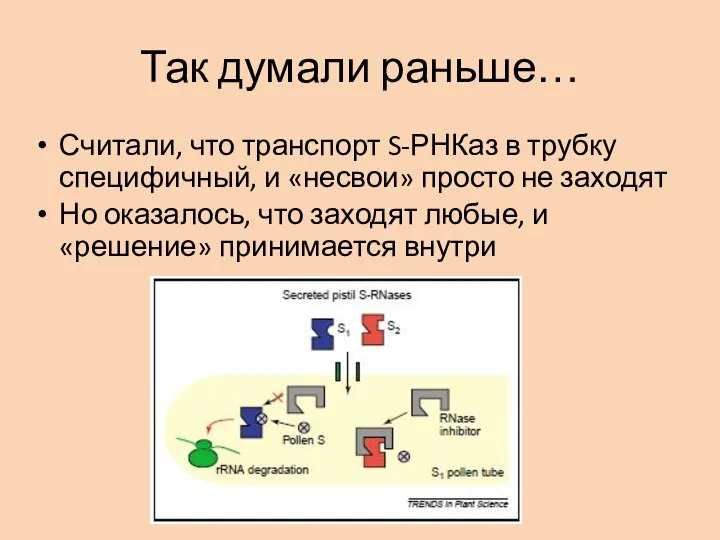
Слайд 17Так думали раньше…
Считали, что транспорт S-РНКаз в трубку специфичный, и «несвои» просто
Так думали раньше…
Считали, что транспорт S-РНКаз в трубку специфичный, и «несвои» просто
Слайд 18S-РНКаза
S-РНКаза синтезируется в проводниковом тракте , пока пыльцевые трубки растут через столбик.
S-РНКаза
S-РНКаза синтезируется в проводниковом тракте , пока пыльцевые трубки растут через столбик.

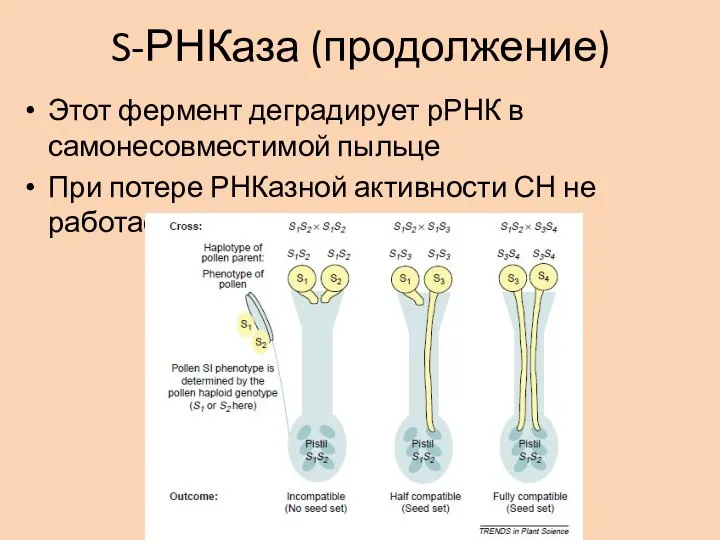
Слайд 19S-РНКаза (продолжение)
Этот фермент деградирует рРНК в самонесовместимой пыльце
При потере РНКазной активности СН
S-РНКаза (продолжение)
Этот фермент деградирует рРНК в самонесовместимой пыльце
При потере РНКазной активности СН
Слайд 20Более ранние модели
Предполагали, что узнается своя РНКаза и не деградирует, а начинает
Более ранние модели
Предполагали, что узнается своя РНКаза и не деградирует, а начинает

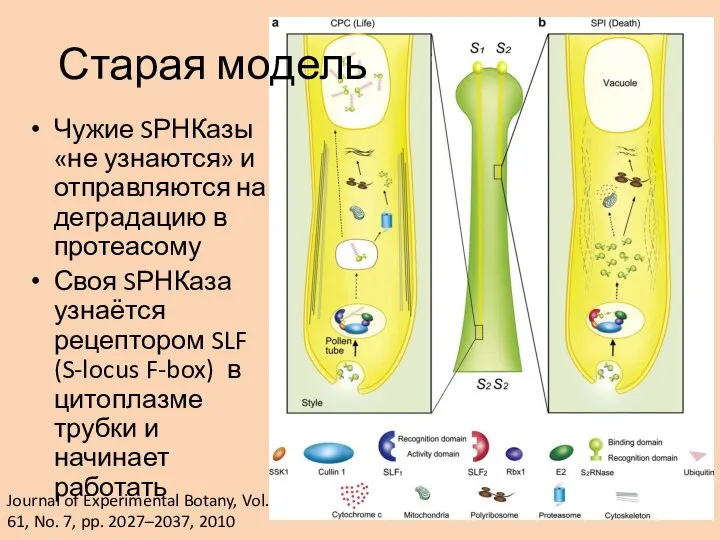
Слайд 21Чужие SРНКазы «не узнаются» и отправляются на деградацию в протеасому
Своя SРНКаза узнаётся
Чужие SРНКазы «не узнаются» и отправляются на деградацию в протеасому
Своя SРНКаза узнаётся
Слайд 22Новая модель: коллективное узнавание
Многочисленные SLF белки, которые производит пыльца данного S-гаплотипа, вместе
Новая модель: коллективное узнавание
Многочисленные SLF белки, которые производит пыльца данного S-гаплотипа, вместе

Слайд 23Резюме по петунии: помиловать?
Представлены два возможных механизма узнавания и деградации «чужих» РНКаз
В
Резюме по петунии: помиловать?
Представлены два возможных механизма узнавания и деградации «чужих» РНКаз
В
Слайд 24Резюме по петунии: казнить?
Своя РНКаза также входит в трубку двумя возможными путями
Резюме по петунии: казнить?
Своя РНКаза также входит в трубку двумя возможными путями
Слайд 25Спорофитная СН (крестоцветные)
Меняемся ролями: мужской пепитид – лиганд, женский – рецептор
У капусты
Спорофитная СН (крестоцветные)
Меняемся ролями: мужской пепитид – лиганд, женский – рецептор
У капусты
Слайд 26Мужская детерминанта – SCR/SPP (S-locus cystein-rich protein, S-locus pollen protein)
Полиморфизм мужской детерминанты
Мужская детерминанта – SCR/SPP (S-locus cystein-rich protein, S-locus pollen protein)
Полиморфизм мужской детерминанты

Слайд 27Cубклеточная локализация SRK
SRK обнаружены в плазмалемме и эндосомах у капусты
Присутствует на ПМ
В
Cубклеточная локализация SRK
SRK обнаружены в плазмалемме и эндосомах у капусты
Присутствует на ПМ
В
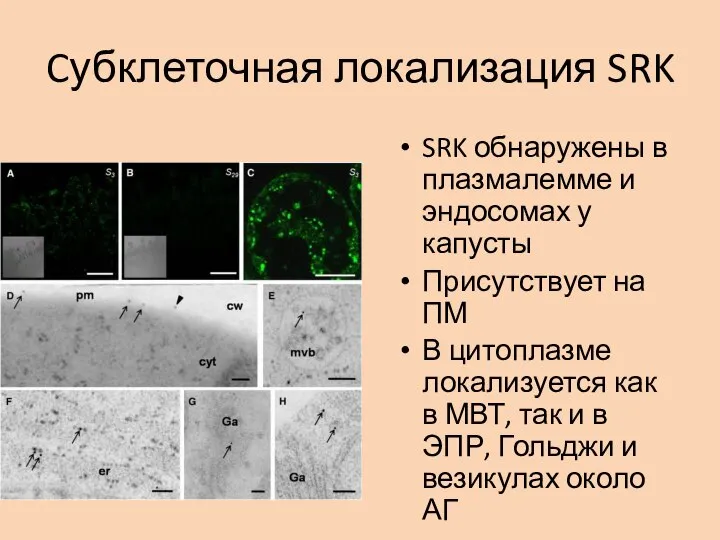
Слайд 28Колокализация с эндосомой
Использовали 2 маркёра эндосом, оба показали, что SRK находится там
Для
Колокализация с эндосомой
Использовали 2 маркёра эндосом, оба показали, что SRK находится там
Для
Слайд 29Интернализация SRK
Как может быть, что одна сторона клетки отвергает «неугодную» пыльцу, а
Интернализация SRK
Как может быть, что одна сторона клетки отвергает «неугодную» пыльцу, а
Слайд 30SRK и эндосомы
Клетка папиллы рыльца в ожидании пыльцы: SRK концентрируется в мембране,
SRK и эндосомы
Клетка папиллы рыльца в ожидании пыльцы: SRK концентрируется в мембране,
Слайд 31Резюме по капусте
Резюме по капусте
 Презентация на тему ТЕЛА И ВЕЩЕСТВА
Презентация на тему ТЕЛА И ВЕЩЕСТВА  Основные группы микроорганизмов
Основные группы микроорганизмов Отгадай грызуна зоопарка
Отгадай грызуна зоопарка Грибы. Часть 2
Грибы. Часть 2 Эти забавные домашние питомцы
Эти забавные домашние питомцы Грибная пора. Интерактивная игра
Грибная пора. Интерактивная игра Питание ягодных культур
Питание ягодных культур Көбелек сабағы
Көбелек сабағы Группы биотических связей между организмами
Группы биотических связей между организмами Тип членистоногие
Тип членистоногие Близнецы в генетике
Близнецы в генетике Что растёт на клумбе
Что растёт на клумбе Особенности сенсорной функции у детей и подростков
Особенности сенсорной функции у детей и подростков Педагогическая физиология детей и подростков
Педагогическая физиология детей и подростков Деление клетки (митоз)
Деление клетки (митоз) Механизмы регуляции ферментативной активности
Механизмы регуляции ферментативной активности Органы цветкового растения
Органы цветкового растения Изучение автоматизированной системы управления воздушным режимом
Изучение автоматизированной системы управления воздушным режимом Ученика 9б класса СОШ № 288 КОШЕЛЕВА НИКИТЫ
Ученика 9б класса СОШ № 288 КОШЕЛЕВА НИКИТЫ Защита растений в ландшафте
Защита растений в ландшафте ГМО. Аргументы за
ГМО. Аргументы за Паразитизм
Паразитизм Презентация на тему Первые вестники весны - Первоцветы
Презентация на тему Первые вестники весны - Первоцветы  Картофель - пищевое паслёновое растение
Картофель - пищевое паслёновое растение Презентация на тему Человеческий мозг
Презентация на тему Человеческий мозг  Отдел покрытосеменные: капустные, маковые
Отдел покрытосеменные: капустные, маковые Розы (фотографии)
Розы (фотографии) Растения
Растения