- Синапс

Содержание
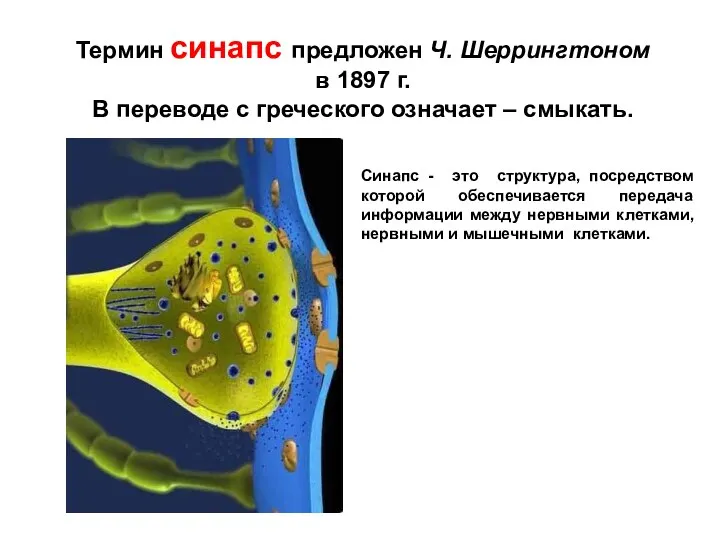
- 2. Синапс - это структура, посредством которой обеспечивается передача информации между нервными клетками, нервными и мышечными клетками.
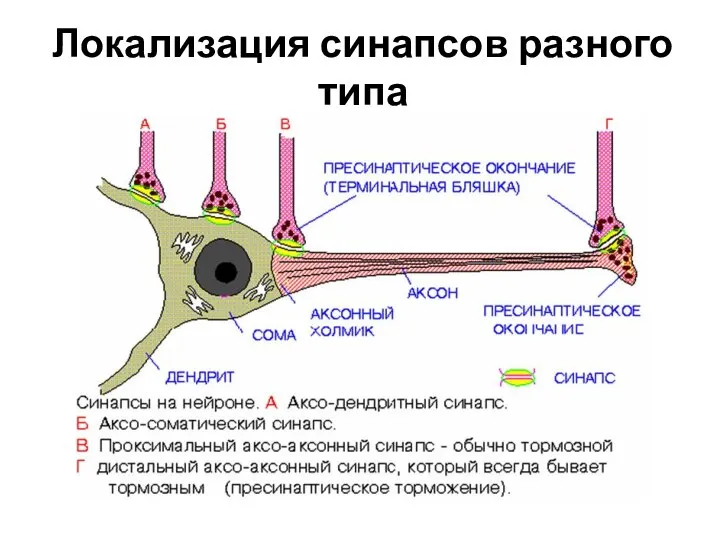
- 3. КЛАССИФИКАЦИЯ СИНАПСОВ 1. По местоположению: а.) центральные (головной и спинной мозг) - аксосоматические, аксоаксональные, аксодендретические; -
- 4. 5.) По природе медиатора химические делятся на следующие группы: I. Группа биогенных аминов: а.) катехоламины –
- 5. Локализация синапсов разного типа
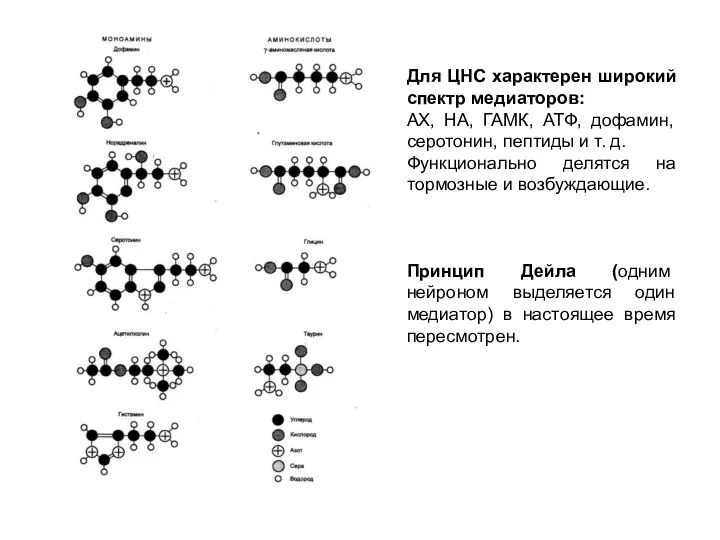
- 6. Для ЦНС характерен широкий спектр медиаторов: АХ, НА, ГАМК, АТФ, дофамин, серотонин, пептиды и т. д.
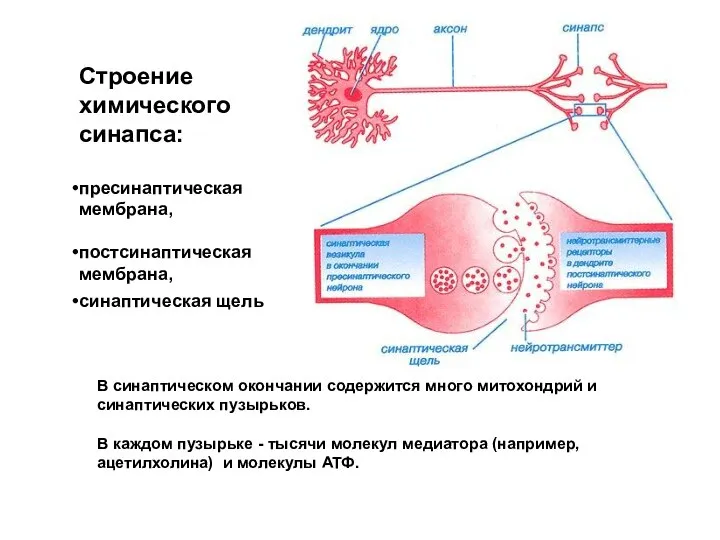
- 8. Строение химического синапса: пресинаптическая мембрана, постсинаптическая мембрана, синаптическая щель В синаптическом окончании содержится много митохондрий и
- 9. Строение химического синапса Пресинаптическая мембрана образована аксональным окончанием, которое теряет в этом месте миелиновую оболочку. Здесь
- 10. Постсинаптическая мембрана (или концевая пластинка) имеет многочисленные складки, увеличивающие площадь ее взаимодействия с медитором. На мембране
- 11. Элементы нервно-мышечного синапса
- 12. Активные зоны в двигательных нервных окончаниях лягушки и мыши лягушка мышь Синаптический бутон
- 13. 1954 Дель Кастилло и Катц детально описывают ПКП и МПКП в нервно-мышечном синапсе. Они предположили, что
- 14. А Б В Г
- 16. Потенциал концевой пластинки Возбуждающий потенциал концевой пластинки (ВПСП) существует только локально на постсинаптической мембране. Его величина
- 17. Потенциалы и токи концевой пластинки на разных расстояниях от нее
- 18. Если синапс возбуждающий, то увеличивается проницаемость постсинаптической мембраны для натрия и калия. Возникает ВПСП. Он существует
- 19. Постсинаптические потенциалы
- 20. МЕХАНИЗМ ТРАНСФОРМАЦИИ ВПКП В ПД КЛЕТКИ После возникновения ВПКП, между деполяризованной мембраной концевой пластинки и находящемся
- 21. Расположение рецепор-управляемых и потенциалозависимых каналов на мембране мышечной клетки. ПП= -80 мВ ПП= -80мВ -80 мВ
- 22. ТРАНСФОРМАЦИЯ ВПКП В ПД -80 мВ -30 мВ -80 мВ Nа+ Nа+ Екр=-60 мВ Екр=-60мВ -30
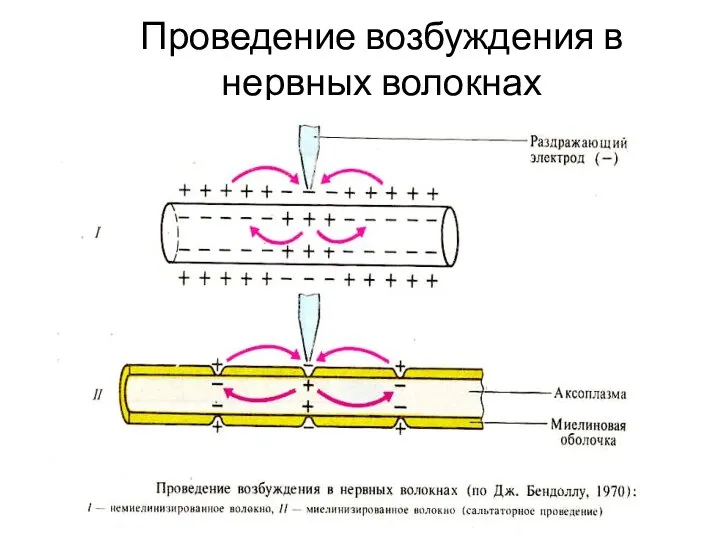
- 23. Проведение возбуждения в нервных волокнах
- 25. Химические синапсы по эффекту , оказываемому медиатором на постсинаптическую мембрану делят на: Ионотропные Метаботропные
- 26. Ионотропный синапс
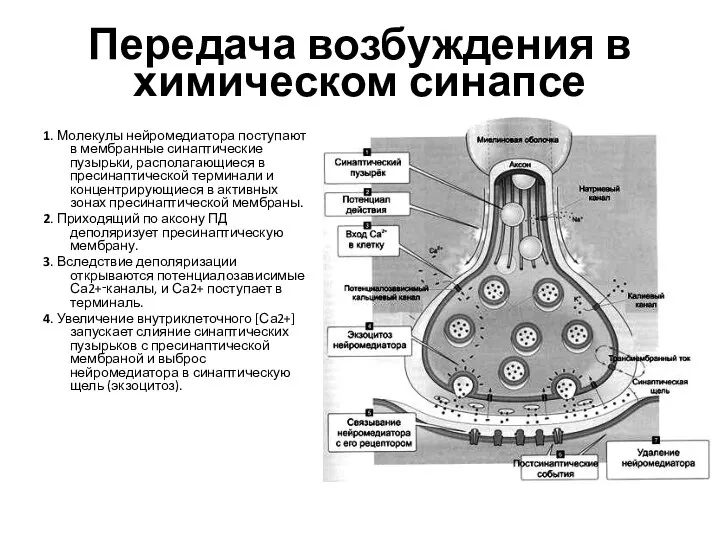
- 27. Передача возбуждения в химическом синапсе 1. Молекулы нейромедиатора поступают в мембранные синаптические пузырьки, располагающиеся в пресинаптической
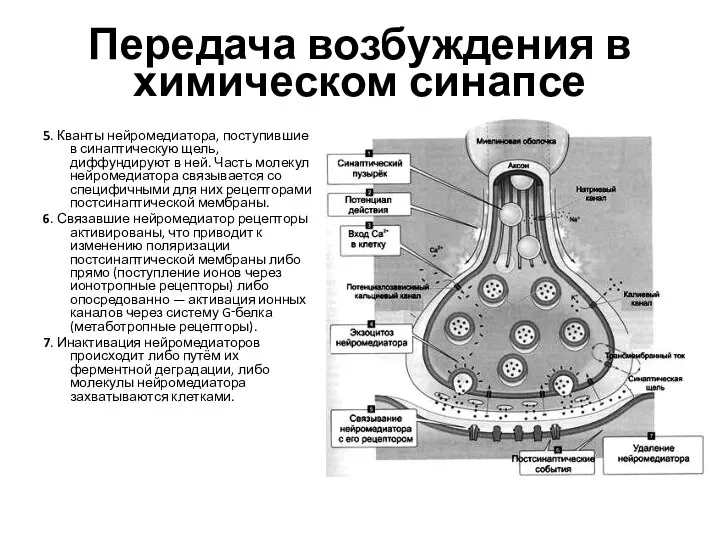
- 28. Передача возбуждения в химическом синапсе 5. Кванты нейромедиатора, поступившие в синаптическую щель, диффундируют в ней. Часть
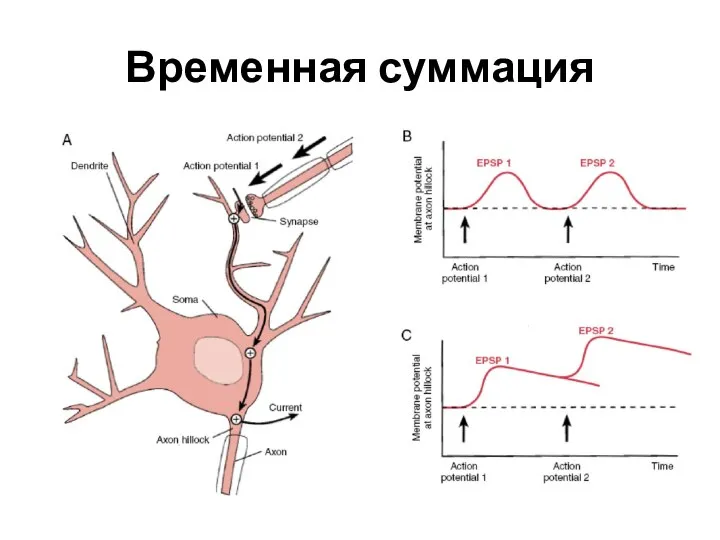
- 29. Временная суммация
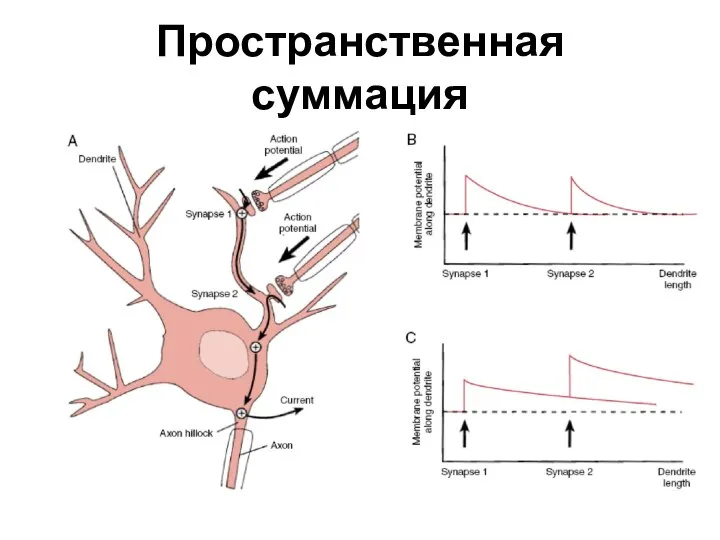
- 30. Пространственная суммация
- 31. Метаботропный синапс
- 32. Постсинаптические рецепторы Ионотропные Быстрые Единый комплекс с ионным каналом Работают на открытие каналов Никотиновые холинорецепторы, рецепторы
- 33. Никотиновый и мускариновый холинорецепторы
- 34. Метаболизм медиаторов: АХ
- 35. Метаболизм медиаторов: НА
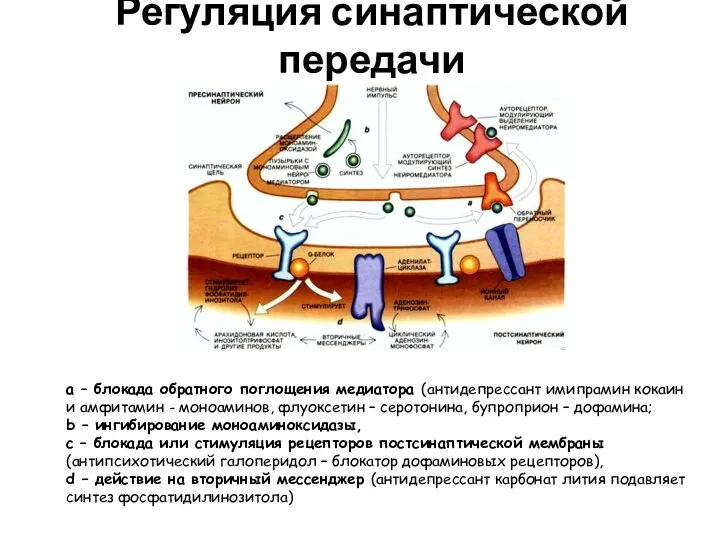
- 36. Регуляция синаптической передачи а – блокада обратного поглощения медиатора (антидепрессант имипрамин кокаин и амфитамин - моноаминов,
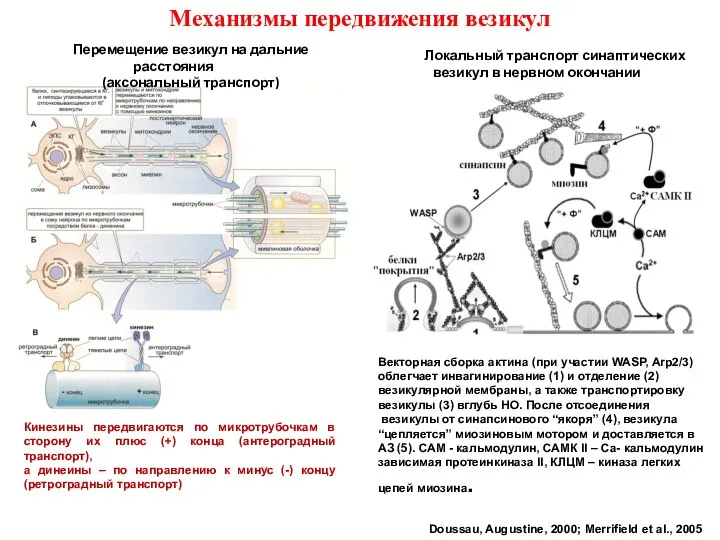
- 37. Механизмы передвижения везикул Doussau, Augustine, 2000; Merrifield et al., 2005 Перемещение везикул на дальние расстояния (аксональный
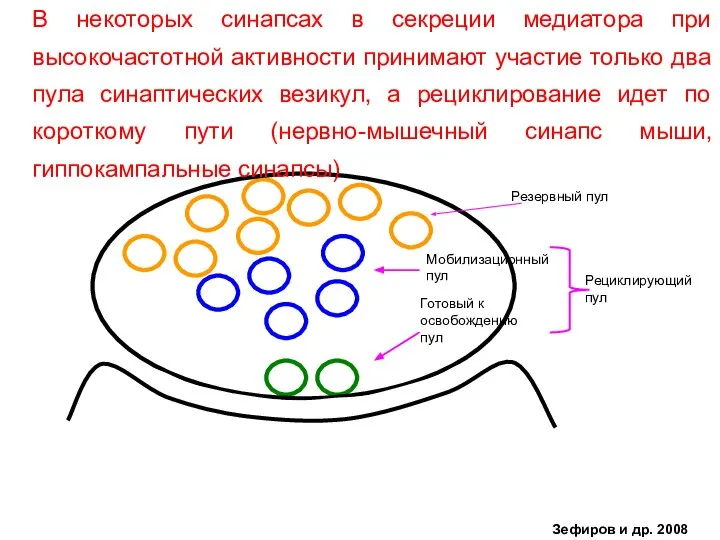
- 38. Готовый к освобождению пул Мобилизационный пул Рециклирующий пул Резервный пул В некоторых синапсах в секреции медиатора
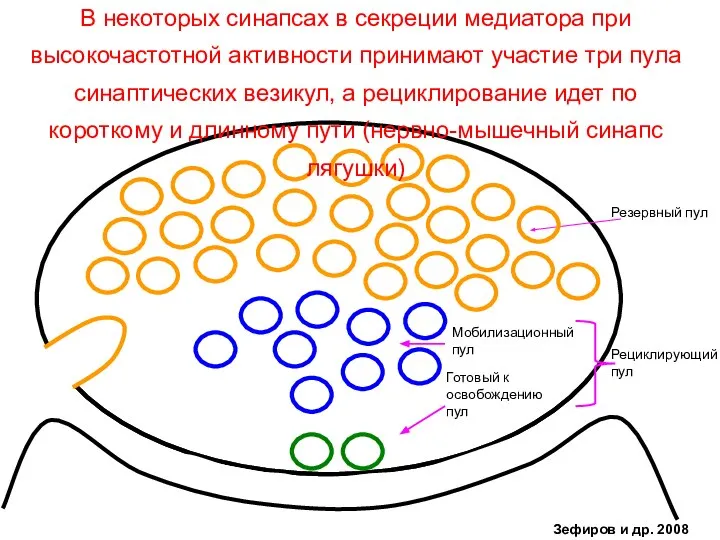
- 39. Готовый к освобождению пул Мобилизационный пул Рециклирующий пул Резервный пул В некоторых синапсах в секреции медиатора
- 40. Пулы синаптических везикул и пути рециклирования Зефиров, 2007 Короткий путь в рециклирующий пул (50 секунд) Длинный
- 41. Современные представления о синаптической везикуле (Takamori et al. 2006) Протонная помпа Везикулярный транспортер ацетилхолина (ВТАХ) Заполнение
- 43. Отпочковывание пузырьков от эндосомы Подход к мембране Подготовка к слиянию Слияние и опорожнение пузырька (экзоцитоз) Заполнение
- 44. Синаптические везикулы фиксируются большей частью на цитоскелете посредством протеина синапсина (sinapsin). Oн покрывает везикулы снаружи и
- 47. Экзоцитоз синаптических везикул: - Вызванный (синхронный и асинхронный) - Спонтанный Tang et al., 2006
- 49. Докирование Праймирование Экзоцитоз АДФ+Ф АТФ Ca++ Полное слияние Разборка SNARE- комплекса Ca++ “Kiss-and-run” Приближение VAMP/синаптобревин -
- 50. Скорость везикулярного цикла и надежность синаптической передачи Нарушение везикулярного цикла (блокирование эндоцитоза) и синаптическая эффективность при
- 51. Физиологические особенности химических синапсов: - односторонняя проводимость - синаптическая задержка - квантовый характер выделения медиаторов -истощение
- 52. Классификация блокаторов нервно-мышечной передачи 1.) Местные анестетики, блокируют проведение возбуждение к пресинаптической мембране (новокаин, лидокаин и
- 53. Электрический синапс. Характерны для ЦНС, но встречаются и на периферии (сердце, гладкомышечная ткань ). Представляют собой
- 54. Ультраструктура нексуса (щелевого контакта)
- 55. 2 1 3 3 Электрический синапс 1 – пресинаптическая мембрана 2 – постсинаптическая мембрана 3 -
- 57. Электрический синапс А. Распределение токов. При возбуждении клетки 1 в нее входит натриевый ток (INa). Она
- 58. Проведение возбуждения в нервных волокнах
- 59. Отличия электрического синапса от химического: - отсутствие синаптической задержки двустороннее проведением возбуждения относится к возбуждающим синапсам
- 60. Структура смешанного синапса А - участок химической передачи. Б - участок электрической передачи. 1. Пресинаптическая мембрана.
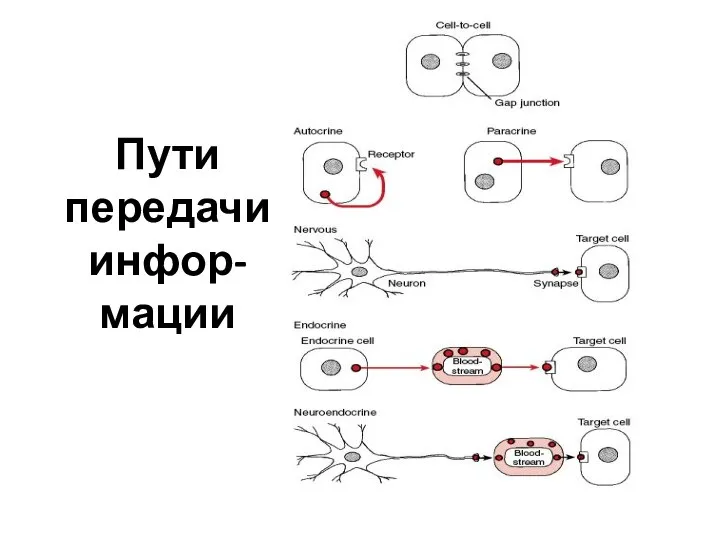
- 61. Пути передачи инфор-мации
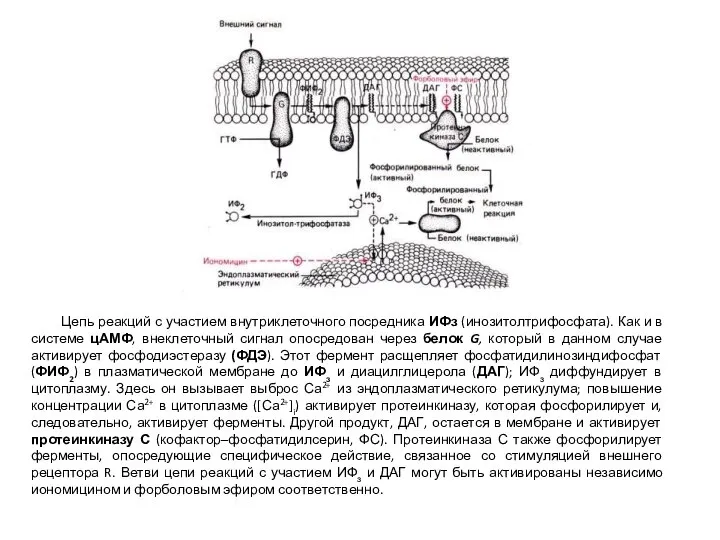
- 62. Цепь реакций с участием внутриклеточного посредника ИФз (инозитолтрифосфата). Как и в системе цАМФ, внеклеточный сигнал опосредован
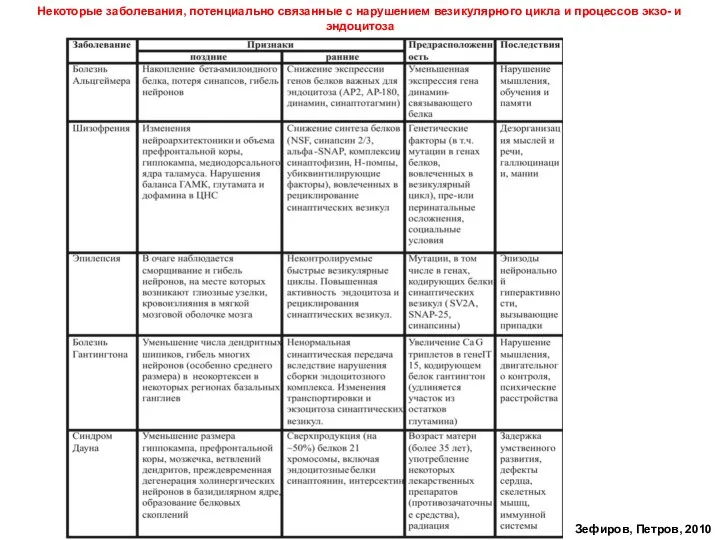
- 63. Некоторые заболевания, потенциально связанные с нарушением везикулярного цикла и процессов экзо- и эндоцитоза Зефиров, Петров, 2010
- 65. Скачать презентацию
Слайд 2Синапс - это структура, посредством которой обеспечивается передача информации между нервными клетками,
Синапс - это структура, посредством которой обеспечивается передача информации между нервными клетками,
Слайд 3КЛАССИФИКАЦИЯ СИНАПСОВ
1. По местоположению:
а.) центральные (головной и спинной мозг)
- аксосоматические, аксоаксональные,
КЛАССИФИКАЦИЯ СИНАПСОВ
1. По местоположению:
а.) центральные (головной и спинной мозг)
- аксосоматические, аксоаксональные,

Слайд 4
5.) По природе медиатора химические делятся на следующие группы:
I. Группа биогенных аминов:
а.)
5.) По природе медиатора химические делятся на следующие группы: I. Группа биогенных аминов: а.)

Слайд 5Локализация синапсов разного типа
Локализация синапсов разного типа
Слайд 6Для ЦНС характерен широкий спектр медиаторов:
АХ, НА, ГАМК, АТФ, дофамин, серотонин,
Для ЦНС характерен широкий спектр медиаторов:
АХ, НА, ГАМК, АТФ, дофамин, серотонин,

Слайд 8Строение химического синапса:
пресинаптическая мембрана,
постсинаптическая мембрана,
синаптическая щель
В синаптическом окончании содержится много
Строение химического синапса:
пресинаптическая мембрана,
постсинаптическая мембрана,
синаптическая щель
В синаптическом окончании содержится много
Слайд 9Строение химического синапса
Пресинаптическая мембрана образована аксональным окончанием, которое теряет в этом месте
Строение химического синапса
Пресинаптическая мембрана образована аксональным окончанием, которое теряет в этом месте

Слайд 10Постсинаптическая мембрана (или концевая пластинка) имеет многочисленные складки, увеличивающие площадь ее взаимодействия
Постсинаптическая мембрана (или концевая пластинка) имеет многочисленные складки, увеличивающие площадь ее взаимодействия

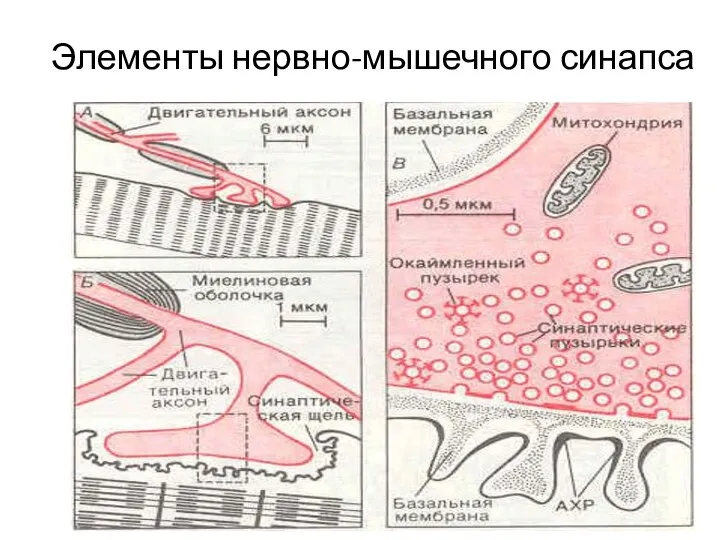
Слайд 11Элементы нервно-мышечного синапса
Элементы нервно-мышечного синапса
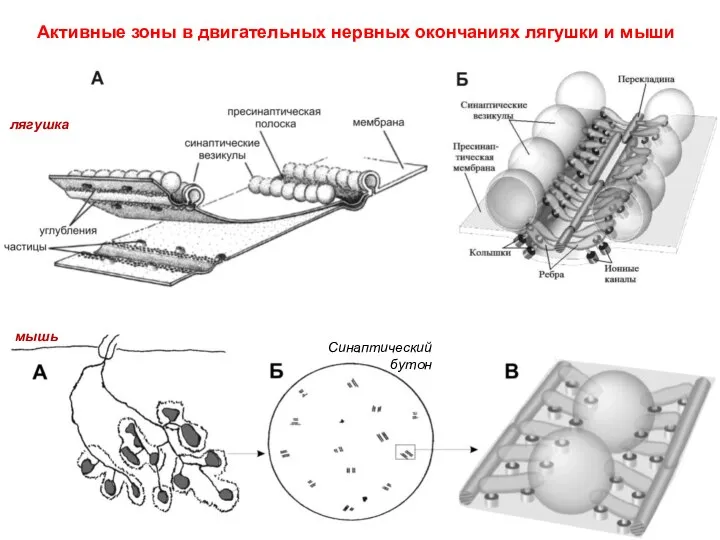
Слайд 12Активные зоны в двигательных нервных окончаниях лягушки и мыши
лягушка
мышь
Синаптический бутон
Активные зоны в двигательных нервных окончаниях лягушки и мыши
лягушка
мышь
Синаптический бутон
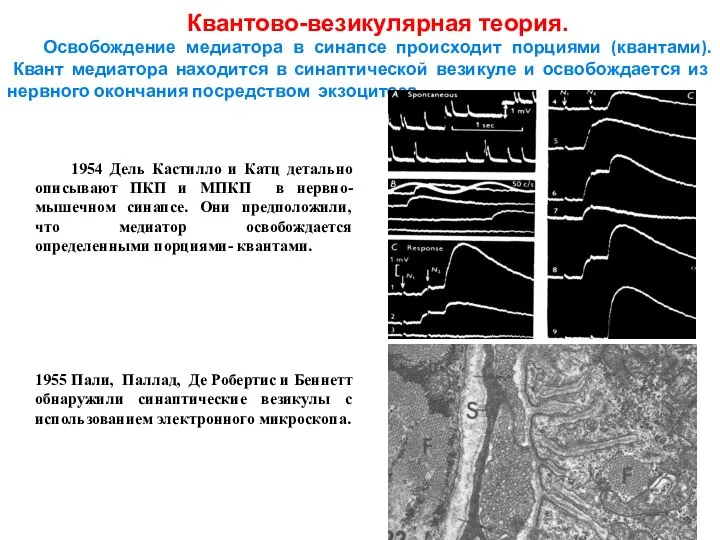
Слайд 13 1954 Дель Кастилло и Катц детально описывают ПКП и МПКП в нервно-мышечном
1954 Дель Кастилло и Катц детально описывают ПКП и МПКП в нервно-мышечном
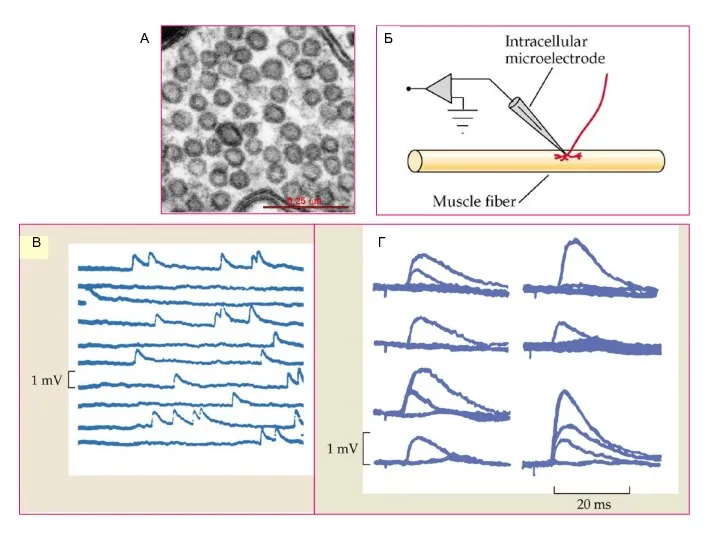
Слайд 14А
Б
В
Г
А
Б
В
Г
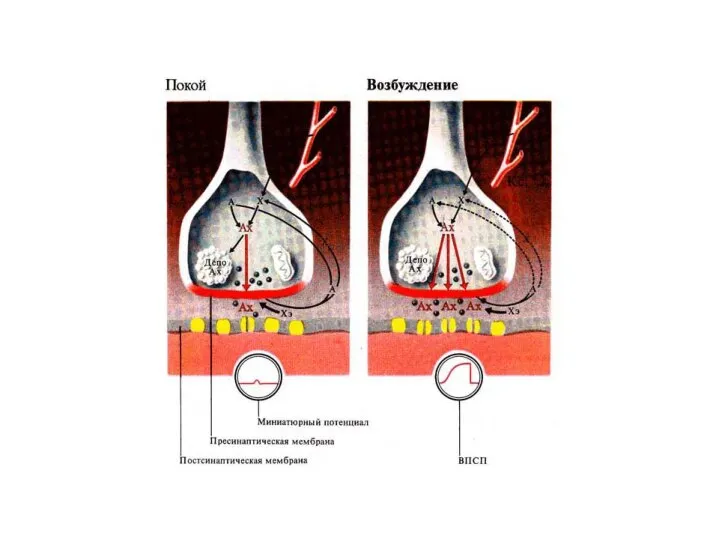
Слайд 16Потенциал концевой пластинки
Возбуждающий потенциал концевой пластинки (ВПСП) существует только локально на
Потенциал концевой пластинки
Возбуждающий потенциал концевой пластинки (ВПСП) существует только локально на

Слайд 17Потенциалы и токи концевой пластинки на разных расстояниях от нее
Потенциалы и токи концевой пластинки на разных расстояниях от нее
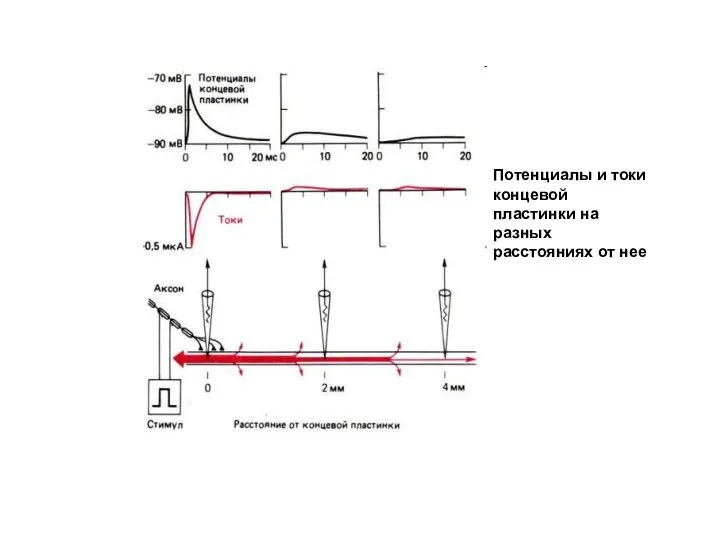
Слайд 18Если синапс возбуждающий, то увеличивается проницаемость постсинаптической мембраны для натрия и калия.
Если синапс возбуждающий, то увеличивается проницаемость постсинаптической мембраны для натрия и калия.

Слайд 19Постсинаптические потенциалы
Постсинаптические потенциалы
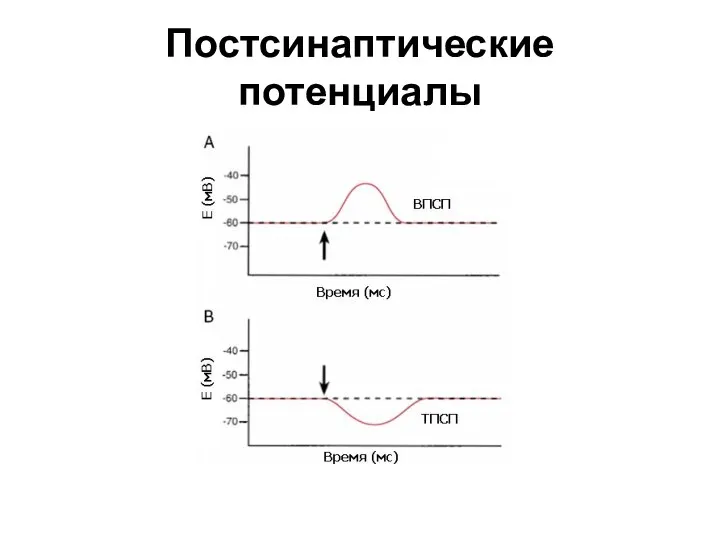
Слайд 20МЕХАНИЗМ ТРАНСФОРМАЦИИ ВПКП В ПД КЛЕТКИ
После возникновения ВПКП, между деполяризованной мембраной концевой
МЕХАНИЗМ ТРАНСФОРМАЦИИ ВПКП В ПД КЛЕТКИ
После возникновения ВПКП, между деполяризованной мембраной концевой

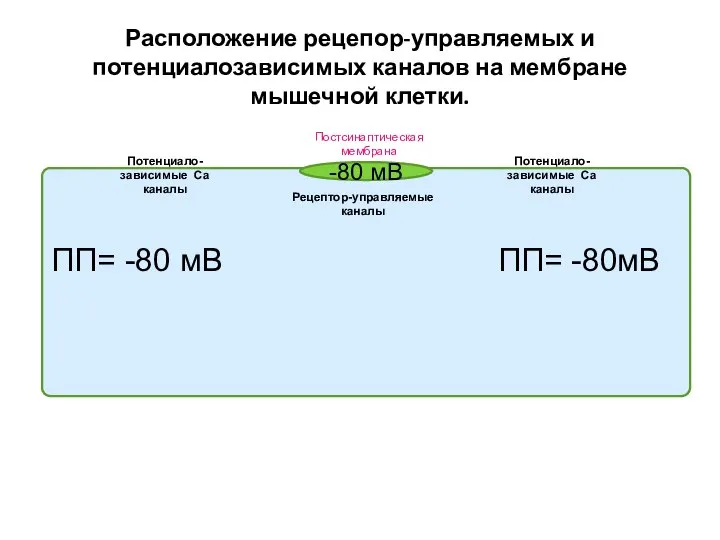
Слайд 21Расположение рецепор-управляемых и потенциалозависимых каналов на мембране мышечной клетки.
ПП= -80 мВ ПП=
Расположение рецепор-управляемых и потенциалозависимых каналов на мембране мышечной клетки.
ПП= -80 мВ ПП=
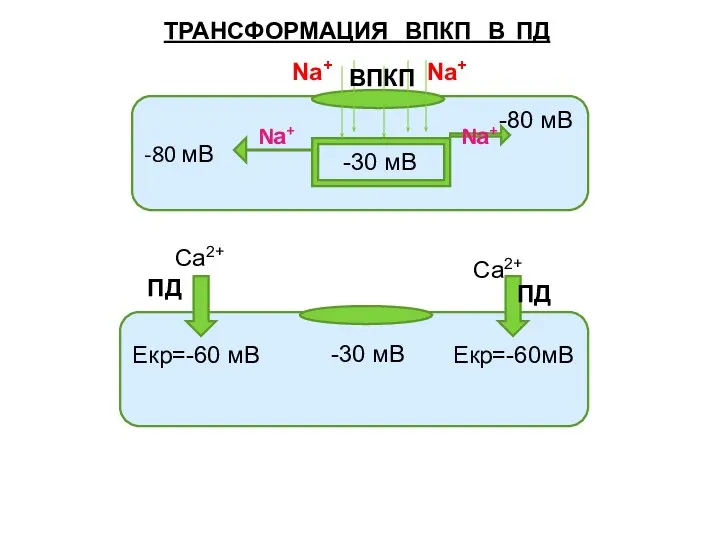
Слайд 22ТРАНСФОРМАЦИЯ ВПКП В ПД
-80 мВ
-30 мВ
-80 мВ
Nа+
Nа+
Екр=-60 мВ Екр=-60мВ
-30
ТРАНСФОРМАЦИЯ ВПКП В ПД
-80 мВ
-30 мВ
-80 мВ
Nа+
Nа+
Екр=-60 мВ Екр=-60мВ
-30
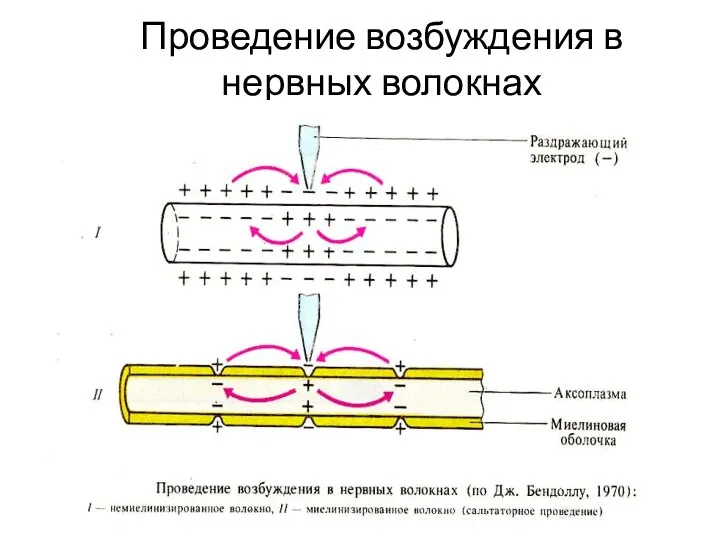
Слайд 23Проведение возбуждения в нервных волокнах
Проведение возбуждения в нервных волокнах
Слайд 25Химические синапсы по эффекту , оказываемому медиатором на постсинаптическую мембрану делят на:
Химические синапсы по эффекту , оказываемому медиатором на постсинаптическую мембрану делят на:

Слайд 26Ионотропный синапс
Ионотропный синапс
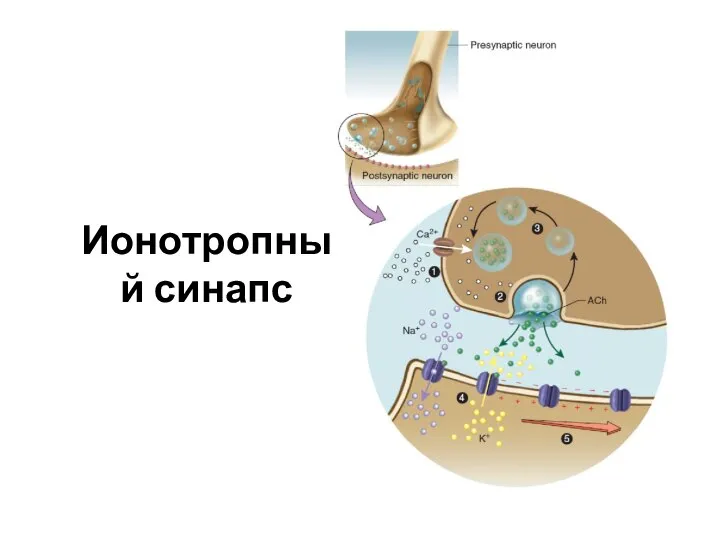
Слайд 27Передача возбуждения в химическом синапсе
1. Молекулы нейромедиатора поступают в мембранные синаптические пузырьки,
Передача возбуждения в химическом синапсе
1. Молекулы нейромедиатора поступают в мембранные синаптические пузырьки,
Слайд 28Передача возбуждения в химическом синапсе
5. Кванты нейромедиатора, поступившие в синаптическую щель, диффундируют
Передача возбуждения в химическом синапсе
5. Кванты нейромедиатора, поступившие в синаптическую щель, диффундируют
Слайд 29Временная суммация
Временная суммация
Слайд 30Пространственная суммация
Пространственная суммация
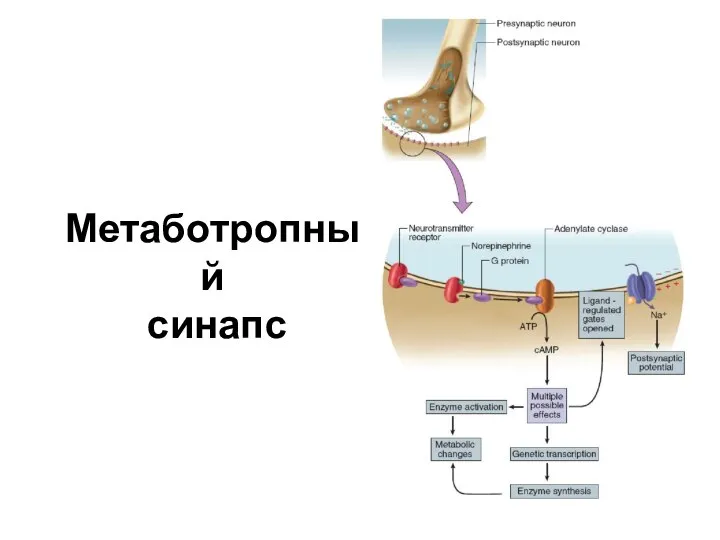
Слайд 31Метаботропный
синапс
Метаботропный
синапс
Слайд 32Постсинаптические рецепторы
Ионотропные
Быстрые
Единый комплекс с ионным каналом
Работают на открытие каналов
Никотиновые холинорецепторы, рецепторы ГАМК,
Постсинаптические рецепторы
Ионотропные
Быстрые
Единый комплекс с ионным каналом
Работают на открытие каналов
Никотиновые холинорецепторы, рецепторы ГАМК,

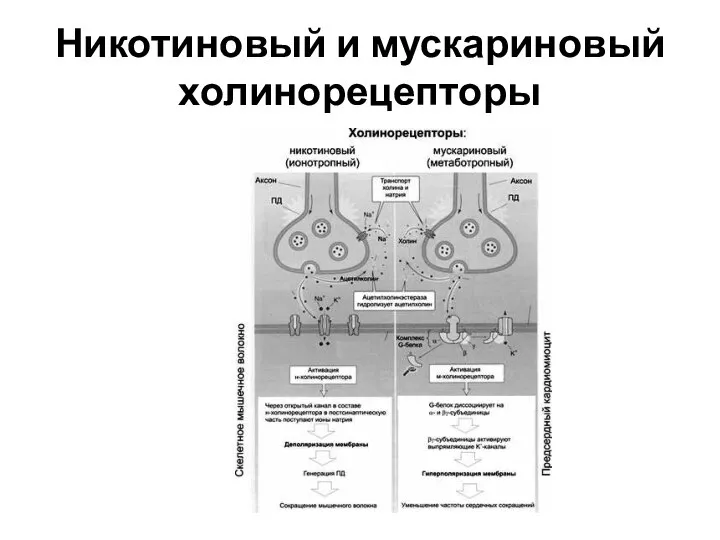
Слайд 33Никотиновый и мускариновый холинорецепторы
Никотиновый и мускариновый холинорецепторы
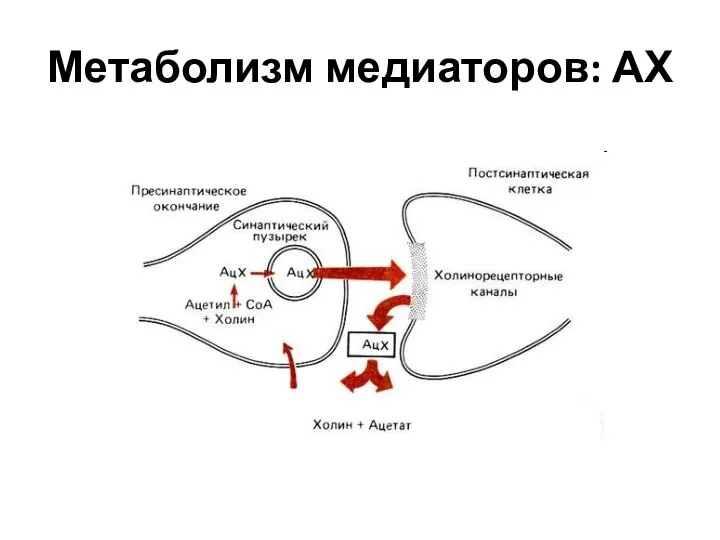
Слайд 34Метаболизм медиаторов: АХ
Метаболизм медиаторов: АХ
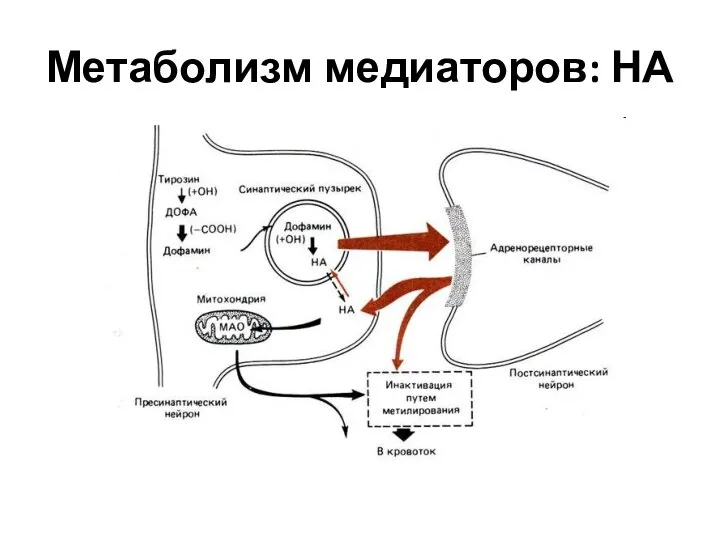
Слайд 35Метаболизм медиаторов: НА
Метаболизм медиаторов: НА
Слайд 36Регуляция синаптической передачи
а – блокада обратного поглощения медиатора (антидепрессант имипрамин кокаин и
Регуляция синаптической передачи
а – блокада обратного поглощения медиатора (антидепрессант имипрамин кокаин и
Слайд 37Механизмы передвижения везикул
Doussau, Augustine, 2000; Merrifield et al., 2005
Перемещение везикул на
Механизмы передвижения везикул
Doussau, Augustine, 2000; Merrifield et al., 2005
Перемещение везикул на
Слайд 38Готовый к
освобождению
пул
Мобилизационный
пул
Рециклирующий
пул
Резервный пул
В некоторых синапсах в секреции медиатора при высокочастотной активности
Готовый к
освобождению
пул
Мобилизационный
пул
Рециклирующий
пул
Резервный пул
В некоторых синапсах в секреции медиатора при высокочастотной активности
Слайд 39Готовый к
освобождению
пул
Мобилизационный
пул
Рециклирующий
пул
Резервный пул
В некоторых синапсах в секреции медиатора при высокочастотной активности
Готовый к
освобождению
пул
Мобилизационный
пул
Рециклирующий
пул
Резервный пул
В некоторых синапсах в секреции медиатора при высокочастотной активности
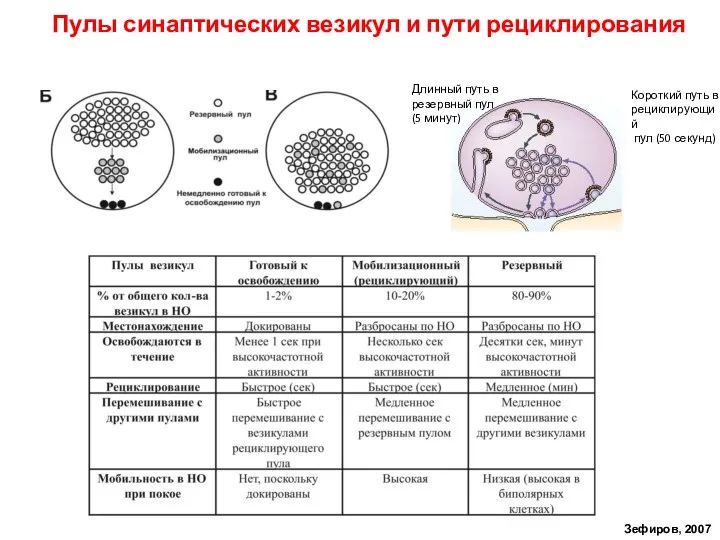
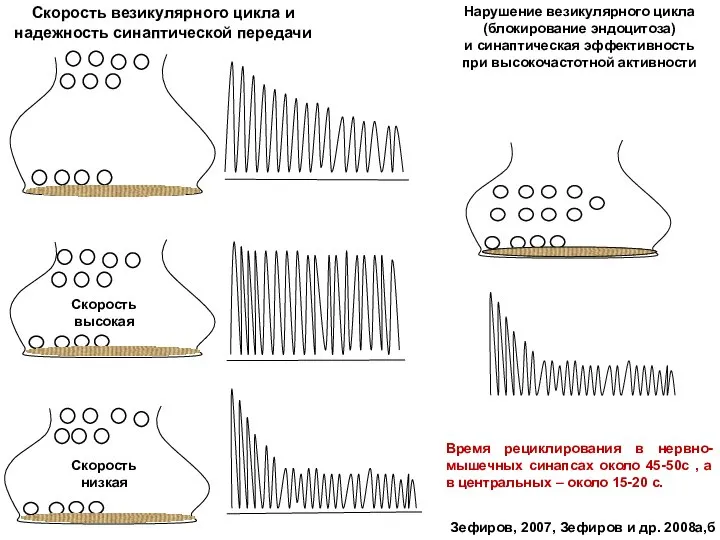
Слайд 40Пулы синаптических везикул и пути рециклирования
Зефиров, 2007
Короткий путь в
рециклирующий
пул (50
Пулы синаптических везикул и пути рециклирования
Зефиров, 2007
Короткий путь в
рециклирующий
пул (50
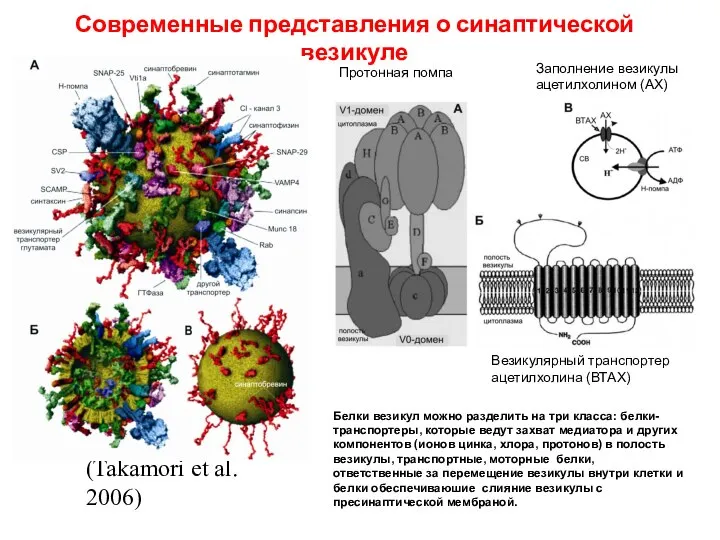
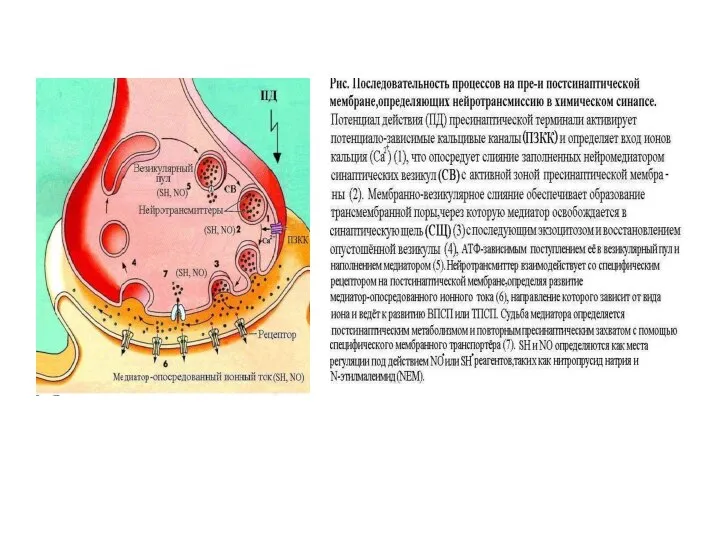
Слайд 41Современные представления о синаптической везикуле
(Takamori et al. 2006)
Протонная помпа
Везикулярный транспортер ацетилхолина
Современные представления о синаптической везикуле
(Takamori et al. 2006)
Протонная помпа
Везикулярный транспортер ацетилхолина
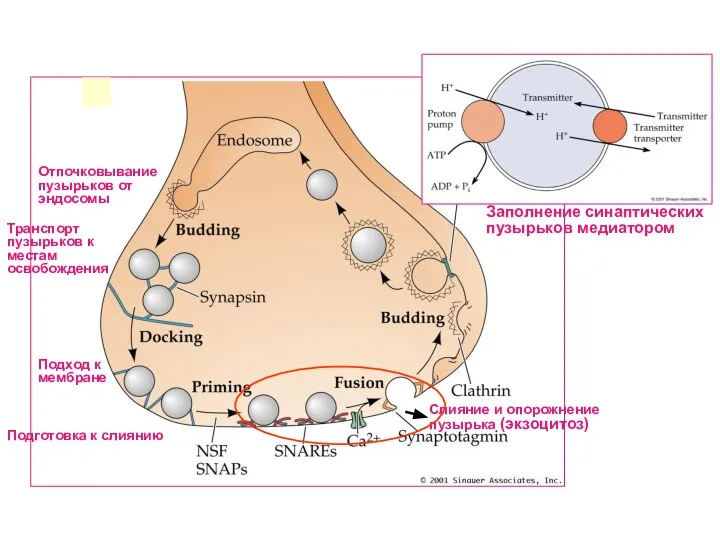
Слайд 43Отпочковывание пузырьков от эндосомы
Подход к мембране
Подготовка к слиянию
Слияние и опорожнение пузырька (экзоцитоз)
Заполнение
Отпочковывание пузырьков от эндосомы
Подход к мембране
Подготовка к слиянию
Слияние и опорожнение пузырька (экзоцитоз)
Заполнение
Слайд 44Синаптические везикулы фиксируются большей частью на цитоскелете посредством протеина синапсина (sinapsin).
Oн
Синаптические везикулы фиксируются большей частью на цитоскелете посредством протеина синапсина (sinapsin).
Oн

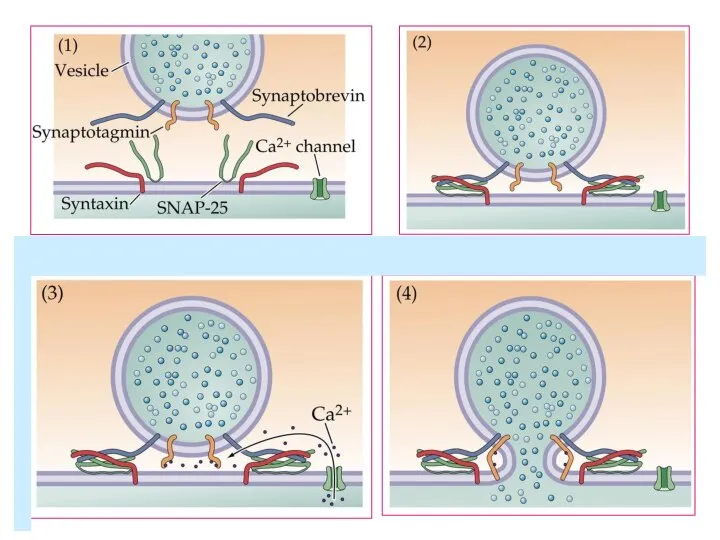
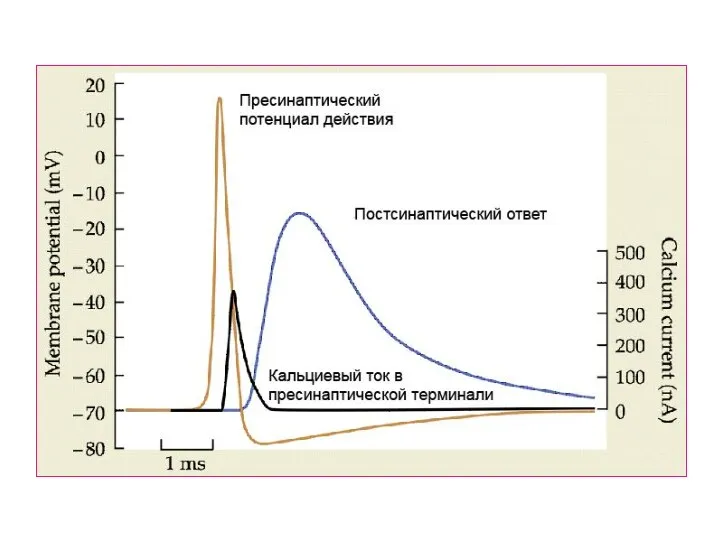
Слайд 47Экзоцитоз синаптических везикул:
- Вызванный
(синхронный и асинхронный)
- Спонтанный
Tang et al., 2006
Экзоцитоз синаптических везикул:
- Вызванный
(синхронный и асинхронный)
- Спонтанный
Tang et al., 2006
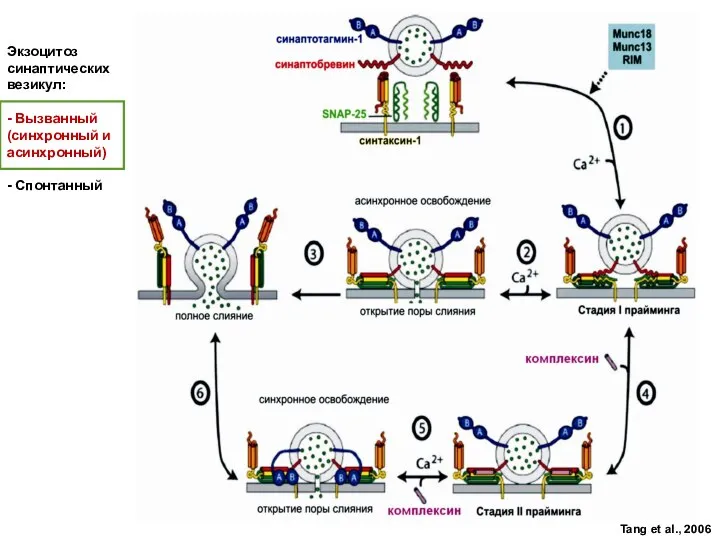
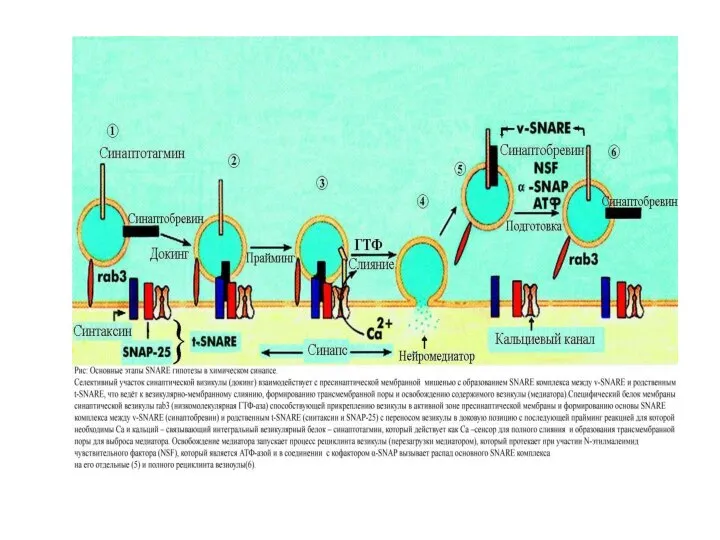
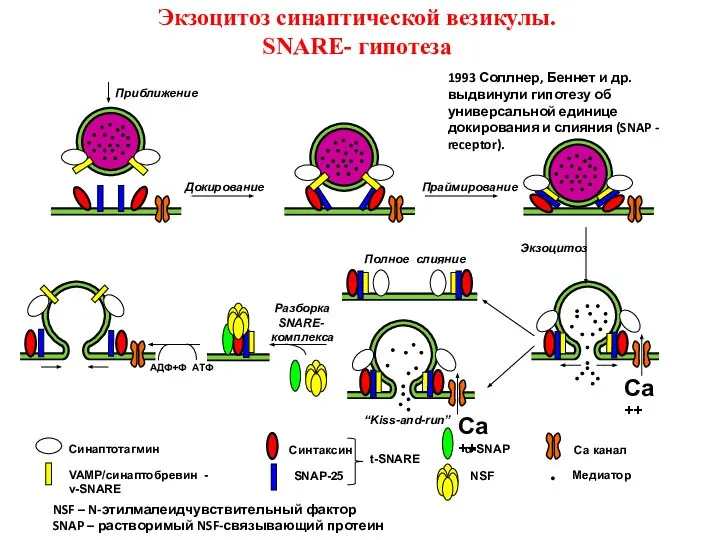
Слайд 49Докирование
Праймирование
Экзоцитоз
АДФ+Ф АТФ
Ca++
Полное слияние
Разборка
SNARE-
комплекса
Ca++
“Kiss-and-run”
Приближение
VAMP/синаптобревин - v-SNARE
SNAP-25
Синтаксин
Синаптотагмин
α-SNAP
NSF
Са канал
Медиатор
t-SNARE
Экзоцитоз синаптической везикулы.
SNARE-
Докирование
Праймирование
Экзоцитоз
АДФ+Ф АТФ
Ca++
Полное слияние
Разборка
SNARE-
комплекса
Ca++
“Kiss-and-run”
Приближение
VAMP/синаптобревин - v-SNARE
SNAP-25
Синтаксин
Синаптотагмин
α-SNAP
NSF
Са канал
Медиатор
t-SNARE
Экзоцитоз синаптической везикулы.
SNARE-
Слайд 50Скорость везикулярного цикла и надежность синаптической передачи
Нарушение везикулярного цикла (блокирование эндоцитоза)
и
Скорость везикулярного цикла и надежность синаптической передачи
Нарушение везикулярного цикла (блокирование эндоцитоза)
и
Слайд 51Физиологические особенности
химических синапсов:
- односторонняя проводимость
- синаптическая задержка
- квантовый характер выделения
Физиологические особенности
химических синапсов:
- односторонняя проводимость
- синаптическая задержка
- квантовый характер выделения

Слайд 52Классификация блокаторов нервно-мышечной передачи
1.) Местные анестетики, блокируют проведение возбуждение к пресинаптической мембране
Классификация блокаторов нервно-мышечной передачи
1.) Местные анестетики, блокируют проведение возбуждение к пресинаптической мембране

Слайд 53Электрический синапс.
Характерны для ЦНС, но встречаются и на периферии (сердце, гладкомышечная ткань
Электрический синапс.
Характерны для ЦНС, но встречаются и на периферии (сердце, гладкомышечная ткань

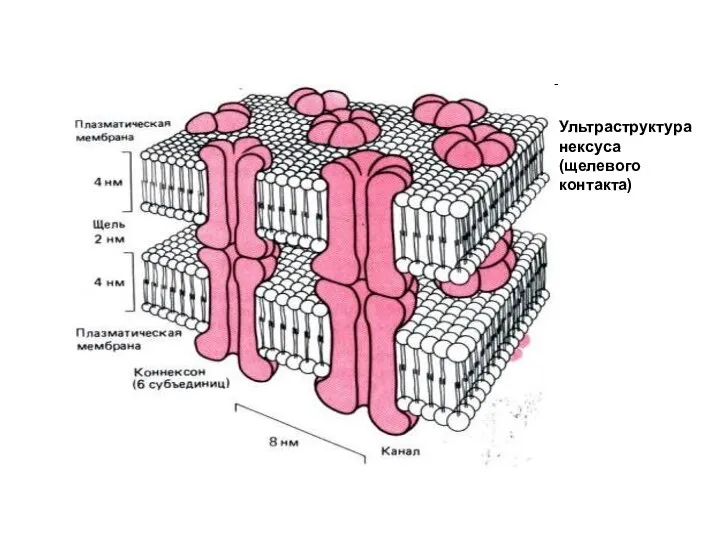
Слайд 54Ультраструктура нексуса (щелевого контакта)
Ультраструктура нексуса (щелевого контакта)
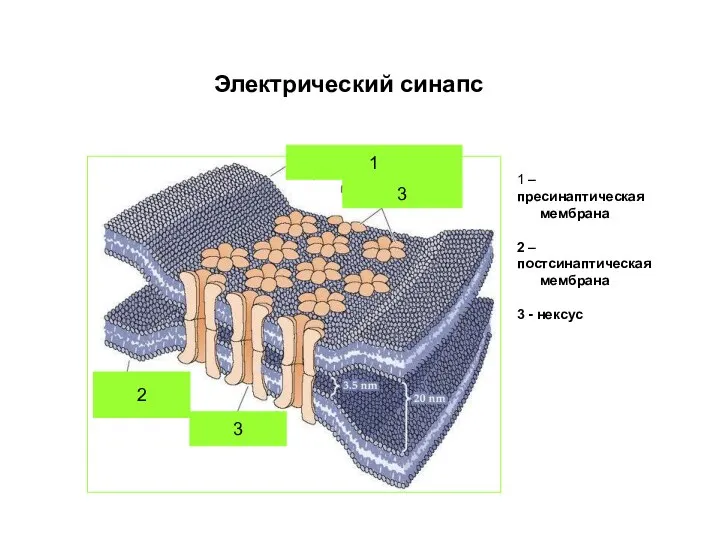
Слайд 552
1
3
3
Электрический синапс
1 – пресинаптическая
мембрана
2 – постсинаптическая
мембрана
3 - нексус
2
1
3
3
Электрический синапс
1 – пресинаптическая
мембрана
2 – постсинаптическая
мембрана
3 - нексус
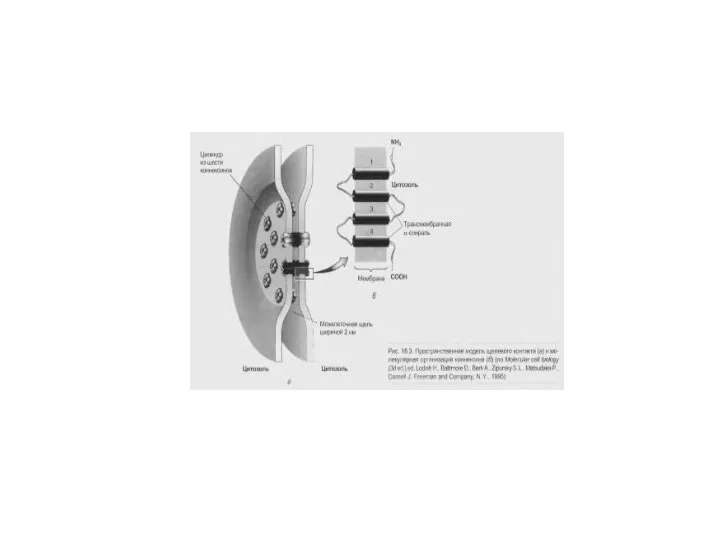
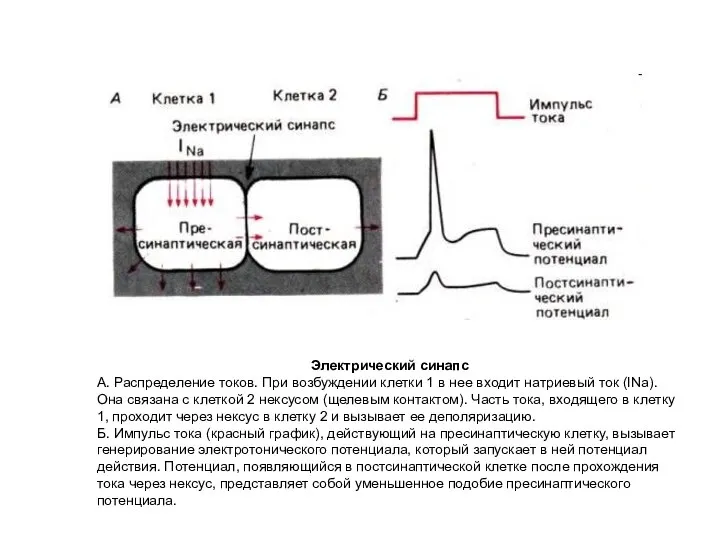
Слайд 57Электрический синапс
А. Распределение токов. При возбуждении клетки 1 в нее входит натриевый
Электрический синапс
А. Распределение токов. При возбуждении клетки 1 в нее входит натриевый
Слайд 58Проведение возбуждения в нервных волокнах
Проведение возбуждения в нервных волокнах
Слайд 59
Отличия электрического синапса от химического:
- отсутствие синаптической задержки
двустороннее проведением возбуждения
относится к возбуждающим
Отличия электрического синапса от химического:
- отсутствие синаптической задержки
двустороннее проведением возбуждения
относится к возбуждающим

Слайд 60Структура смешанного синапса
А - участок химической
передачи.
Б - участок электрической
передачи.
1. Пресинаптическая
Структура смешанного синапса
А - участок химической
передачи.
Б - участок электрической
передачи.
1. Пресинаптическая
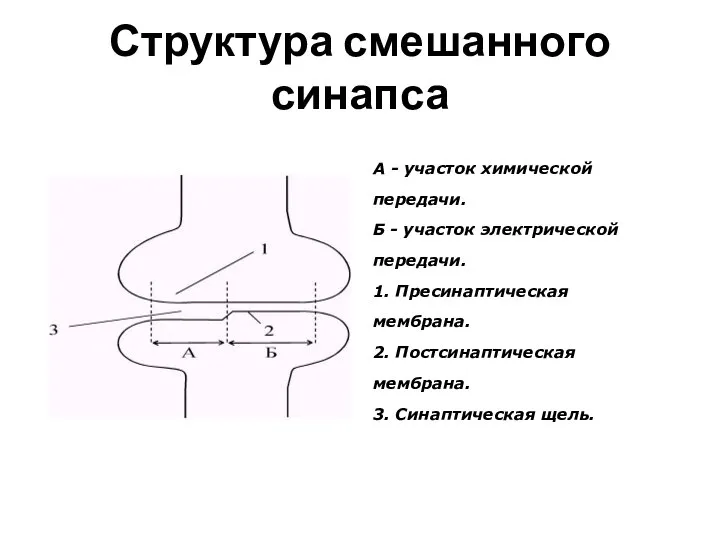
Слайд 61Пути передачи инфор-мации
Пути передачи инфор-мации
Слайд 62 Цепь реакций с участием внутриклеточного посредника ИФз (инозитолтрифосфата). Как и в
Цепь реакций с участием внутриклеточного посредника ИФз (инозитолтрифосфата). Как и в
Слайд 63Некоторые заболевания, потенциально связанные с нарушением везикулярного цикла и процессов экзо- и
Некоторые заболевания, потенциально связанные с нарушением везикулярного цикла и процессов экзо- и
 Развитие половых клеток
Развитие половых клеток Тип плоские черви. Класс сосальщики
Тип плоские черви. Класс сосальщики Транспорт веществ. Органы, участвующие в транспорте веществ у животных
Транспорт веществ. Органы, участвующие в транспорте веществ у животных Биотехнология 2
Биотехнология 2 Жилкование листьев
Жилкование листьев Лёгкие
Лёгкие LEKTsIYa__5_SD (1)
LEKTsIYa__5_SD (1) Лекция 5
Лекция 5 Обеспечение клеток энергией
Обеспечение клеток энергией Органы осязания, обоняния, вкуса
Органы осязания, обоняния, вкуса +Лекция Физиология сердца (1)
+Лекция Физиология сердца (1) Морфология леса
Морфология леса Рис
Рис Старение. Теории о процессе старения
Старение. Теории о процессе старения 1 апреля - День птиц
1 апреля - День птиц Мембранные органоиды клетки. Лекция №5
Мембранные органоиды клетки. Лекция №5 Как шум влияет на здоровье человека?
Как шум влияет на здоровье человека? Отдел Эвгленовые водоросли (Euglenophyta)
Отдел Эвгленовые водоросли (Euglenophyta) Дикие животные
Дикие животные День экологических знаний 15 апреля. Прощание с ботаникой 6 класс
День экологических знаний 15 апреля. Прощание с ботаникой 6 класс Поведенческий портрет бородатой неясыти
Поведенческий портрет бородатой неясыти Растительный организм и его особенности
Растительный организм и его особенности Исәнме, куянкай! Исәнме, керпе!
Исәнме, куянкай! Исәнме, керпе! Тип Саркожгутиконосцы. Класс Жгутиковые
Тип Саркожгутиконосцы. Класс Жгутиковые Презентация на тему Что такое биоинформатика?
Презентация на тему Что такое биоинформатика?  Животные и растения
Животные и растения Методика Е.П.Ильина. Теппинг-тест
Методика Е.П.Ильина. Теппинг-тест Размножение и развитие земноводных
Размножение и развитие земноводных