- Синапсы. Электрическое взаимодействие клеток: электрический синапс. Глава 3

Содержание
- 2. Впервые ультраструктура межклеточных контактов была подробно изучена в 1963 г. М.Г. Фаркухаром (Farquhar M.G.) и Г.Е.
- 3. Рис. 3-1. Межклеточные контакты и структура щелевого контакта. А - типы межклеточных контактов. Б - модель
- 4. Структура коннексонов Коннексоны позволяют осуществлять обмен ионами и водорастворимыми молекулами с молекулярной массой до 1200-1500 Да
- 5. Рис. 3-2. Коннексоны и их молекулярная организация
- 6. Принцип межклеточного электротонического взаимодействия Для анализа факторов, определяющих условия передачи возбуждения от клетки к клетке, необходимо
- 7. Рис. 3-3. Эквивалентные электрические схемы клеточных контактов. А - эквивалентная электрическая схема клеточного контакта с узкой
- 8. Принципы обнаружения щелевого контакта Существует несколько принципов, лежащих в основе обнаружения щелевого контакта. 1. Электрофизиологический способ
- 9. Рис. 3-4. Определение электрической передачи между клетками. А - электрофизиологический способ определения наличия щелевого контакта и
- 10. Методы изучения щелевых контактов Рассмотрим пассивные электрические свойства сферической клетки и цилиндрического волокна, имеющие одинаковые свойства
- 11. Этот вопрос детально проанализирован при обсуждении механизма пассивного электротонического потенциала. Обсудим функциональное значение Rin. Если на
- 12. Рассмотрим теперь функциональное значение постоянной времени клетки. Обсудим только один из аспектов. Обычно для возбуждения даже
- 13. Рис. 3-5. Изучение электрической передачи между клетками методом двойного patch-clamp и измерения I-V характеристик. А -
- 14. Электрический синапс Принцип работы электрического синапса показан на рис. 3-6 А. Механизм передачи сигнала через электрический
- 15. Лиганд-рецепторное взаимодействие (основные определения) Лиганд - это молекула, которая может взаимодействовать с участком связывания (местами связывания)
- 16. Аффинность лиганда по отношению к участку связывания определяет, насколько вероятно то, что связанный лиганд покинет поверхность
- 17. Лиганды могут быть агонистами или антагонистами. Агонист - это лиганд, который взаимодействует с участком связывания и
- 18. Функциональный антагонизм может осуществляться с помощью разных механизмов в виде непрямого и физиологического антагонизма. Непрямой антагонизм
- 19. Среди аллостерических (аллотопических) модуляторов выделяют несколько видов. 1. Аллостерические (аллотопические) усилители (allosteric enhancers) - это модуляторы,
- 20. Аллостерическое (аллотопическое) взаимодействие - это взаимодействие между лигандами, которые связываются с отличающимися неперекрывающимися местами распознавания на
- 21. Химический синапс Химический синапс выполняет задачу передачи электрического сигнала от пресинаптического нейрона на постсинаптическую клетку. Для
- 22. Рис. 3-6. Химический синапс и принцип его работы
- 23. Аксональный транспорт Мембранные и цитоплазматические компоненты, которые образуются в биосинтезирующем аппарате сомы и проксимальной части дендритов,
- 24. Рис. 3-7. Синтез и рецикл синаптических везикул и их содержимого. А - Биосинтез нейротрансмиттеров. I. Везикулы,
- 25. Механизм высвобождения трансмиттера Серия потенциалов действия «пробегает» вдоль аксона, достигает нервного окончания и деполяризует пресинаптическую зону.
- 26. Рис. 3-8. Высвобождение трансмиттеров. А - белки, встроенные в мембрану везикулы. Б - взаимодействие везикулярных белков
- 27. Преобразование трансмиттера Трансмиттер может быть либо быстро химически расщеплен на неактивные компоненты, либо удален из синаптической
- 28. Рис. 3-9. Расщепление, удаление и обратный захват трансмиттера на примере ацетилхолина
- 29. Типы химических синапсов У части синапсов ионный канал, который при действии лиганда, т.е. благодаря связыванию с
- 30. Таблица 3-1. Перечень соединений, влияющие на никотиновые, мускариновые и холинергические синапсы
- 31. Рис. 3-10. Ионотропные и метаботропные синапсы
- 32. Типы трансмиттеров и механизмы их действия Ацетилхолин. Является медиатором для всех двигательных нейронов, многих интернейронов ЦНС,
- 33. γ-Аминомасляную кислоту выбрасывают в качестве трансмиттера многие тормозные интернейроны, имеющиеся практически во всех отделах центральной нервной
- 34. Норадреналин катехоламин, выполняющий функцию трансмиттера в ЦНС и синтезирующийся прежде всего нейронами locus coeruleus.Это ядро состоит
- 35. Таблица 3-2. Примеры механизмов действия трансмиттеров на различные типы рецепторов
- 36. Типы ацетилхолиновных синапсов Среди синапсов с химическим механизмом передачи электрического сигнала особое место занимает так называемый
- 37. В ионотропном никотиновом АЦХ синапсе две молекулы свободного АЦХ связываются с рецепторным белком, представляющим собой одну
- 38. Рис. 3-11. Механизмы работы ионотропного синапса (А) и метаботропного синапса (Б)
- 39. Ионотропный никотиновый холинергический синапс На примере ионотропного никотинового холинергического синапса рассмотрим ацетилхолиновую передачу с участием ионотропного
- 40. Рис. 3-12. Структура n-холинорецептора
- 41. Глутаматный синапс Многие соединения используются в синапсах как трансмиттеры. Например, глутамат в мозге служит важнейшим трансмиттером
- 42. Рис. 3-13. Один из механизмов работы глутаматного синапса
- 43. Фосфорилирование и дефосфорилирование в глутаматном синапсе Глутаматный синапс содержит на постсинаптической мембране два типа ионотропных рецепторов.
- 44. Рис. 3-14. Глутамат и механизм работы синапса с этим трансмиттером через ионотропные рецепторы (лигандуправляемые катионные каналы)
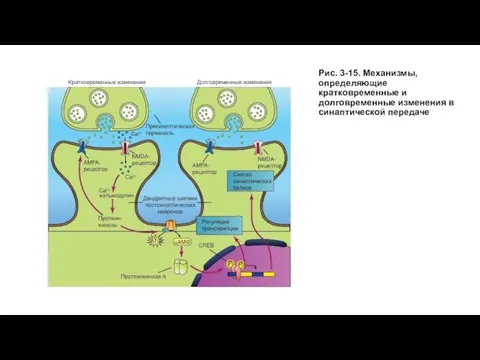
- 45. Механизм долговременной потенциации и долговременной синаптической депрессии На рисунке 3-15 показаны пути, приводящие к повышению уровня
- 46. Рис. 3-15. Механизмы, определяющие кратковременные и долговременные изменения в синаптической передаче
- 47. Рецепторы NMDA и AMPA определяют работу глутаматного синапса Некоторые из глутаматных синапсов, снабженные ионотропными рецепторами -
- 48. Рис. 3-16. Потенциалы, возникающие на постсинаптической мембране глутаматного синапса, которые определяются работой ионотропных рецепторов - лигандуправляемых
- 49. ГАМК как трансмиттер Многие тормозные интернейроны, присутствующие практически во всех отделах ЦНС, и аксоны клеток Пуркинье
- 50. Рис. 3-17. γ-Аминомасляная кислота как трансмиттер в синапсе: расщепление, удаление и обратный захват
- 51. Глицин как трансмиттер Аминокислота глицин выполняет функцию трансмиттера в тормозных синапсах и действует как нейромодулятор. Глицин
- 52. Рис. 3-18. Глицин: расщепление, удаление и обратный захват трансмиттера
- 53. NO как трансмиттер Оксид азота (NO) - это паракринный медиатор, выделяемый эндотелиальными клетками и некоторыми нейронами.
- 54. Рис. 3-19. Оксид азота (NO) в качестве трансмиттера, освобождаемого нейронами
- 55. Возбуждающий постсинаптический потенциал (ВПСП) Рассмотрим этот вопрос на примере АЦХ. Взаимодействуя с рецептором, молекулы АЦХ открывают
- 56. Тормозной постсинаптический потенциал (ТПСП) Решающими для ионного тока в мембране оказываются величина ее потенциала и количество
- 57. Рис. 3-20. Потенциалы и токи, возникающие на постсинаптической мембране. А - процессы освобождения трансмиттера и формирование
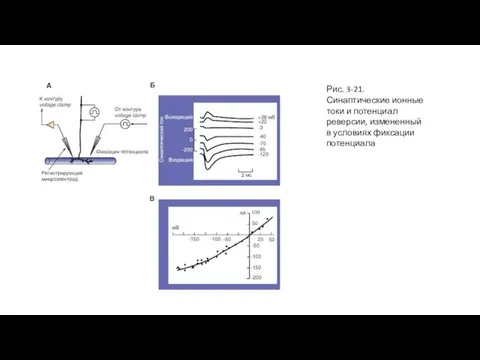
- 58. Синаптические ионные токи в условиях фиксации потенциала На панели (А) представлена схема регистрации токов концевой пластинки
- 59. Рис. 3-21. Синаптические ионные токи и потенциал реверсии, измененный в условиях фиксации потенциала
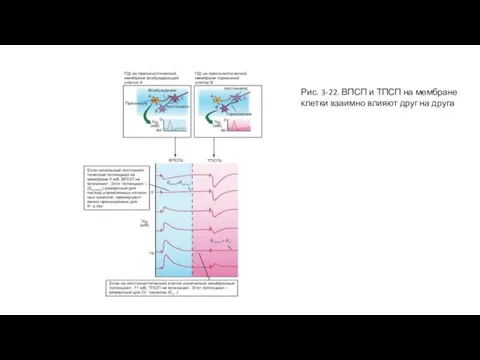
- 60. Взаимное влияние ВПСП и ТПСП Если на мембране клетки активируются одновременно возбуждающие и тормозные синапсы, то
- 61. Рис. 3-22. ВПСП и ТПСП на мембране клетки взаимно влияют друг на друга
- 62. Лиганд-управляемые ионные каналы. Их токи На примере двух различных типов лигандуправляемых ионных каналов (одиночного ионного канала,
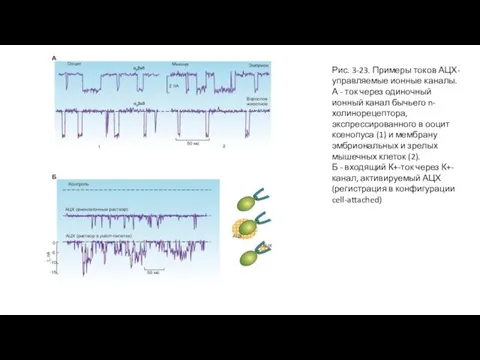
- 63. Рис. 3-23. Примеры токов АЦХ-управляемые ионные каналы. А - ток через одиночный ионный канал бычьего n-холинорецептора,
- 64. Patch-clamp для изучения канальных токов синапса При применении метода patch-clamp в конфигурации outside-out, продемонстрированной на рис.
- 65. Рис. 3-24. Активация ацетилхолинового рецептора - ионотропного одиночного ионного канала в нервномышечном синапсе
- 66. Соединения с синаптическим влиянием Наряду с трансмиттером, другие соединения тоже могут влиять на рецепторный белок. Если
- 67. Рис. 3-25. Свойства основных нейротрансмиттеров (см. таблицу)
- 69. Скачать презентацию
Слайд 2Впервые ультраструктура межклеточных контактов была подробно изучена в 1963 г. М.Г. Фаркухаром
Впервые ультраструктура межклеточных контактов была подробно изучена в 1963 г. М.Г. Фаркухаром

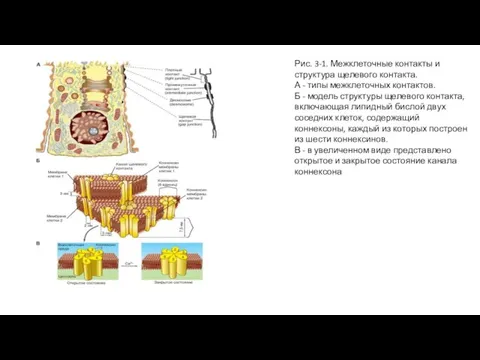
Слайд 3Рис. 3-1. Межклеточные контакты и структура щелевого контакта.
А - типы межклеточных контактов.
Рис. 3-1. Межклеточные контакты и структура щелевого контакта.
А - типы межклеточных контактов.
Слайд 4Структура коннексонов
Коннексоны позволяют осуществлять обмен ионами и водорастворимыми молекулами с молекулярной массой
Структура коннексонов
Коннексоны позволяют осуществлять обмен ионами и водорастворимыми молекулами с молекулярной массой

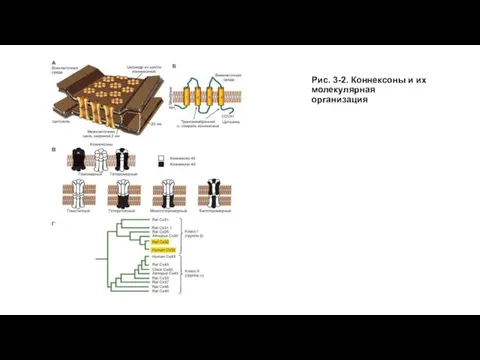
Слайд 5Рис. 3-2. Коннексоны и их молекулярная организация
Рис. 3-2. Коннексоны и их молекулярная организация
Слайд 6Принцип межклеточного электротонического взаимодействия
Для анализа факторов, определяющих условия передачи возбуждения от клетки
Принцип межклеточного электротонического взаимодействия
Для анализа факторов, определяющих условия передачи возбуждения от клетки

Слайд 7Рис. 3-3. Эквивалентные электрические схемы клеточных контактов.
А - эквивалентная электрическая схема клеточного
Рис. 3-3. Эквивалентные электрические схемы клеточных контактов.
А - эквивалентная электрическая схема клеточного

Слайд 8Принципы обнаружения щелевого контакта
Существует несколько принципов, лежащих в основе обнаружения щелевого контакта.
1.
Принципы обнаружения щелевого контакта
Существует несколько принципов, лежащих в основе обнаружения щелевого контакта.
1.

Слайд 9Рис. 3-4. Определение электрической передачи между клетками.
А - электрофизиологический способ определения наличия
Рис. 3-4. Определение электрической передачи между клетками.
А - электрофизиологический способ определения наличия

Слайд 10Методы изучения щелевых контактов
Рассмотрим пассивные электрические свойства сферической клетки и цилиндрического волокна,
Методы изучения щелевых контактов
Рассмотрим пассивные электрические свойства сферической клетки и цилиндрического волокна,

Слайд 11Этот вопрос детально проанализирован при обсуждении механизма пассивного электротонического потенциала.
Обсудим функциональное значение Rin.
Этот вопрос детально проанализирован при обсуждении механизма пассивного электротонического потенциала.
Обсудим функциональное значение Rin.

Слайд 12Рассмотрим теперь функциональное значение постоянной времени клетки. Обсудим только один из аспектов.
Рассмотрим теперь функциональное значение постоянной времени клетки. Обсудим только один из аспектов.

Слайд 13Рис. 3-5. Изучение электрической передачи между клетками методом двойного patch-clamp и измерения
Рис. 3-5. Изучение электрической передачи между клетками методом двойного patch-clamp и измерения

Слайд 14Электрический синапс
Принцип работы электрического синапса показан на рис. 3-6 А. Механизм передачи
Электрический синапс
Принцип работы электрического синапса показан на рис. 3-6 А. Механизм передачи

Слайд 15Лиганд-рецепторное взаимодействие (основные определения)
Лиганд - это молекула, которая может взаимодействовать с участком
Лиганд-рецепторное взаимодействие (основные определения)
Лиганд - это молекула, которая может взаимодействовать с участком

Слайд 16Аффинность лиганда по отношению к участку связывания определяет, насколько вероятно то, что
Аффинность лиганда по отношению к участку связывания определяет, насколько вероятно то, что

Слайд 17Лиганды могут быть агонистами или антагонистами.
Агонист - это лиганд, который взаимодействует с
Лиганды могут быть агонистами или антагонистами.
Агонист - это лиганд, который взаимодействует с

Слайд 18Функциональный антагонизм может осуществляться с помощью разных механизмов в виде непрямого и
Функциональный антагонизм может осуществляться с помощью разных механизмов в виде непрямого и

Слайд 19Среди аллостерических (аллотопических) модуляторов выделяют несколько видов.
1. Аллостерические (аллотопические) усилители (allosteric enhancers)
Среди аллостерических (аллотопических) модуляторов выделяют несколько видов.
1. Аллостерические (аллотопические) усилители (allosteric enhancers)

Слайд 20Аллостерическое (аллотопическое) взаимодействие - это взаимодействие между лигандами, которые связываются с отличающимися
Аллостерическое (аллотопическое) взаимодействие - это взаимодействие между лигандами, которые связываются с отличающимися

Слайд 21Химический синапс
Химический синапс выполняет задачу передачи электрического сигнала от пресинаптического нейрона на
Химический синапс
Химический синапс выполняет задачу передачи электрического сигнала от пресинаптического нейрона на

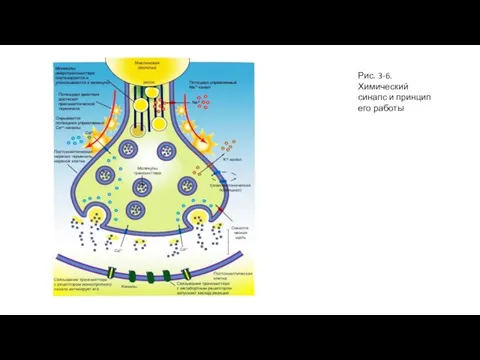
Слайд 22Рис. 3-6. Химический синапс и принцип его работы
Рис. 3-6. Химический синапс и принцип его работы
Слайд 23Аксональный транспорт
Мембранные и цитоплазматические компоненты, которые образуются в биосинтезирующем аппарате сомы и
Аксональный транспорт
Мембранные и цитоплазматические компоненты, которые образуются в биосинтезирующем аппарате сомы и

Слайд 24Рис. 3-7. Синтез и рецикл синаптических везикул и их содержимого.
А - Биосинтез
Рис. 3-7. Синтез и рецикл синаптических везикул и их содержимого.
А - Биосинтез

Слайд 25Механизм высвобождения трансмиттера
Серия потенциалов действия «пробегает» вдоль аксона, достигает нервного окончания и
Механизм высвобождения трансмиттера
Серия потенциалов действия «пробегает» вдоль аксона, достигает нервного окончания и

Слайд 26Рис. 3-8. Высвобождение трансмиттеров.
А - белки, встроенные в мембрану везикулы.
Б -
Рис. 3-8. Высвобождение трансмиттеров.
А - белки, встроенные в мембрану везикулы.
Б -

Слайд 27Преобразование трансмиттера
Трансмиттер может быть либо быстро химически расщеплен на неактивные компоненты, либо
Преобразование трансмиттера
Трансмиттер может быть либо быстро химически расщеплен на неактивные компоненты, либо

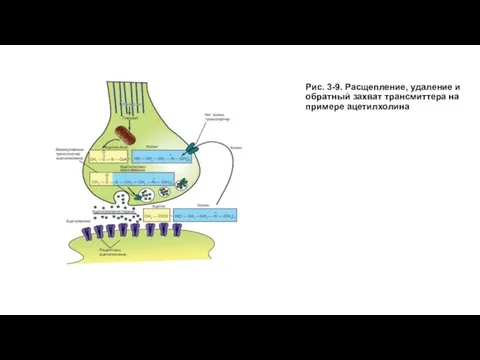
Слайд 28Рис. 3-9. Расщепление, удаление и обратный захват трансмиттера на примере ацетилхолина
Рис. 3-9. Расщепление, удаление и обратный захват трансмиттера на примере ацетилхолина
Слайд 29Типы химических синапсов
У части синапсов ионный канал, который при действии лиганда, т.е.
Типы химических синапсов
У части синапсов ионный канал, который при действии лиганда, т.е.

Слайд 30Таблица 3-1. Перечень соединений, влияющие на никотиновые, мускариновые и холинергические синапсы
Таблица 3-1. Перечень соединений, влияющие на никотиновые, мускариновые и холинергические синапсы

Слайд 31Рис. 3-10. Ионотропные и метаботропные синапсы
Рис. 3-10. Ионотропные и метаботропные синапсы

Слайд 32Типы трансмиттеров и механизмы их действия
Ацетилхолин. Является медиатором для всех двигательных нейронов,
Типы трансмиттеров и механизмы их действия
Ацетилхолин. Является медиатором для всех двигательных нейронов,

Слайд 33γ-Аминомасляную кислоту выбрасывают в качестве трансмиттера многие тормозные интернейроны, имеющиеся практически во
γ-Аминомасляную кислоту выбрасывают в качестве трансмиттера многие тормозные интернейроны, имеющиеся практически во

Слайд 34Норадреналин катехоламин, выполняющий функцию трансмиттера в ЦНС и синтезирующийся прежде всего нейронами
Норадреналин катехоламин, выполняющий функцию трансмиттера в ЦНС и синтезирующийся прежде всего нейронами

Слайд 35Таблица 3-2. Примеры механизмов действия трансмиттеров на различные типы рецепторов
Таблица 3-2. Примеры механизмов действия трансмиттеров на различные типы рецепторов

Слайд 36Типы ацетилхолиновных синапсов
Среди синапсов с химическим механизмом передачи электрического сигнала особое место
Типы ацетилхолиновных синапсов
Среди синапсов с химическим механизмом передачи электрического сигнала особое место

Слайд 37В ионотропном никотиновом АЦХ синапсе две молекулы свободного АЦХ связываются с рецепторным
В ионотропном никотиновом АЦХ синапсе две молекулы свободного АЦХ связываются с рецепторным

Слайд 38Рис. 3-11. Механизмы работы ионотропного синапса (А) и метаботропного синапса (Б)
Рис. 3-11. Механизмы работы ионотропного синапса (А) и метаботропного синапса (Б)
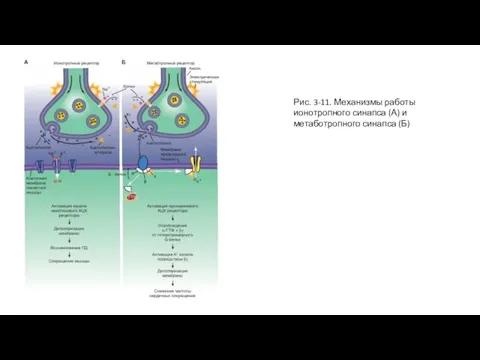
Слайд 39Ионотропный никотиновый холинергический синапс
На примере ионотропного никотинового холинергического синапса рассмотрим ацетилхолиновую передачу
Ионотропный никотиновый холинергический синапс
На примере ионотропного никотинового холинергического синапса рассмотрим ацетилхолиновую передачу

Слайд 40Рис. 3-12. Структура n-холинорецептора
Рис. 3-12. Структура n-холинорецептора
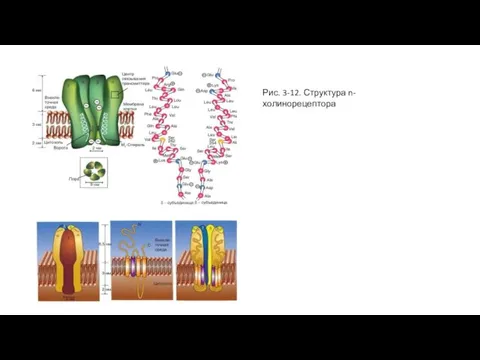
Слайд 41Глутаматный синапс
Многие соединения используются в синапсах как трансмиттеры. Например, глутамат в мозге
Глутаматный синапс
Многие соединения используются в синапсах как трансмиттеры. Например, глутамат в мозге

Слайд 42Рис. 3-13. Один из механизмов работы глутаматного синапса
Рис. 3-13. Один из механизмов работы глутаматного синапса
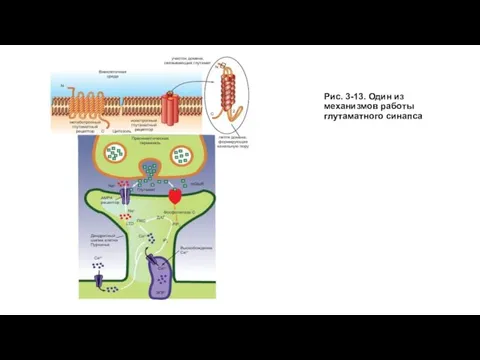
Слайд 43Фосфорилирование и дефосфорилирование в глутаматном синапсе
Глутаматный синапс содержит на постсинаптической мембране два
Фосфорилирование и дефосфорилирование в глутаматном синапсе
Глутаматный синапс содержит на постсинаптической мембране два

Слайд 44Рис. 3-14. Глутамат и механизм работы синапса с этим трансмиттером через ионотропные
Рис. 3-14. Глутамат и механизм работы синапса с этим трансмиттером через ионотропные
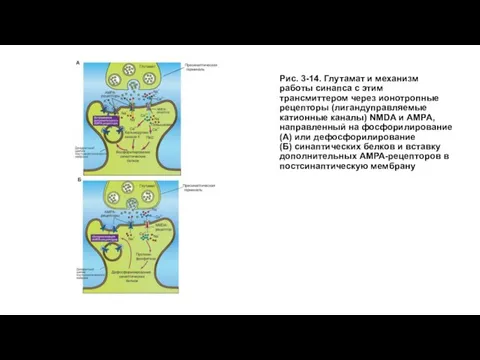
Слайд 45Механизм долговременной потенциации и долговременной синаптической депрессии
На рисунке 3-15 показаны пути, приводящие
Механизм долговременной потенциации и долговременной синаптической депрессии
На рисунке 3-15 показаны пути, приводящие

Слайд 46Рис. 3-15. Механизмы, определяющие кратковременные и долговременные изменения в синаптической передаче
Рис. 3-15. Механизмы, определяющие кратковременные и долговременные изменения в синаптической передаче
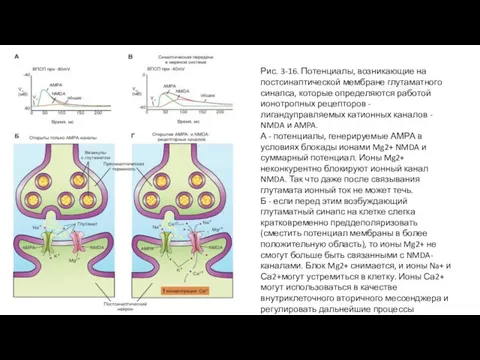
Слайд 47Рецепторы NMDA и AMPA определяют работу глутаматного синапса
Некоторые из глутаматных синапсов, снабженные
Рецепторы NMDA и AMPA определяют работу глутаматного синапса
Некоторые из глутаматных синапсов, снабженные

Слайд 48Рис. 3-16. Потенциалы, возникающие на постсинаптической мембране глутаматного синапса, которые определяются работой
Рис. 3-16. Потенциалы, возникающие на постсинаптической мембране глутаматного синапса, которые определяются работой
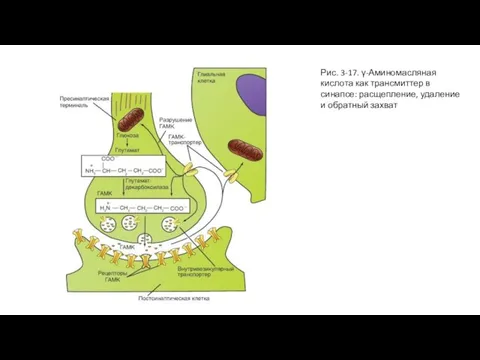
Слайд 49ГАМК как трансмиттер
Многие тормозные интернейроны, присутствующие практически во всех отделах ЦНС, и
ГАМК как трансмиттер
Многие тормозные интернейроны, присутствующие практически во всех отделах ЦНС, и

Слайд 50Рис. 3-17. γ-Аминомасляная кислота как трансмиттер в синапсе: расщепление, удаление и обратный
Рис. 3-17. γ-Аминомасляная кислота как трансмиттер в синапсе: расщепление, удаление и обратный
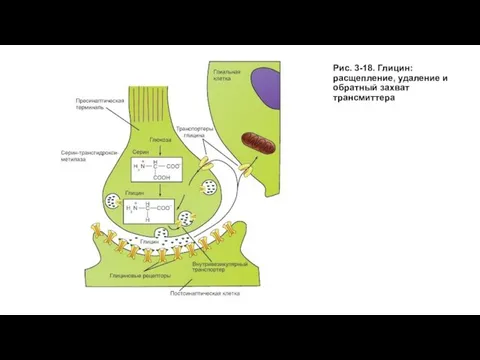
Слайд 51Глицин как трансмиттер
Аминокислота глицин выполняет функцию трансмиттера в тормозных синапсах и действует
Глицин как трансмиттер
Аминокислота глицин выполняет функцию трансмиттера в тормозных синапсах и действует

Слайд 52Рис. 3-18. Глицин: расщепление, удаление и обратный захват трансмиттера
Рис. 3-18. Глицин: расщепление, удаление и обратный захват трансмиттера
Слайд 53NO как трансмиттер
Оксид азота (NO) - это паракринный медиатор, выделяемый эндотелиальными клетками
NO как трансмиттер
Оксид азота (NO) - это паракринный медиатор, выделяемый эндотелиальными клетками

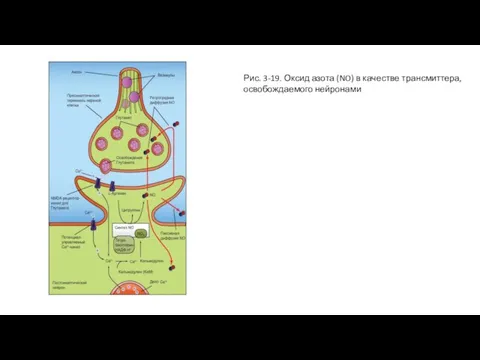
Слайд 54Рис. 3-19. Оксид азота (NO) в качестве трансмиттера, освобождаемого нейронами
Рис. 3-19. Оксид азота (NO) в качестве трансмиттера, освобождаемого нейронами
Слайд 55Возбуждающий постсинаптический потенциал (ВПСП)
Рассмотрим этот вопрос на примере АЦХ. Взаимодействуя с рецептором,
Возбуждающий постсинаптический потенциал (ВПСП)
Рассмотрим этот вопрос на примере АЦХ. Взаимодействуя с рецептором,

Слайд 56Тормозной постсинаптический потенциал (ТПСП)
Решающими для ионного тока в мембране оказываются величина ее
Тормозной постсинаптический потенциал (ТПСП)
Решающими для ионного тока в мембране оказываются величина ее

Слайд 57Рис. 3-20. Потенциалы и токи, возникающие на постсинаптической мембране.
А - процессы освобождения
Рис. 3-20. Потенциалы и токи, возникающие на постсинаптической мембране.
А - процессы освобождения

Слайд 58Синаптические ионные токи
в условиях фиксации потенциала
На панели (А) представлена схема регистрации токов
Синаптические ионные токи
в условиях фиксации потенциала
На панели (А) представлена схема регистрации токов

Слайд 59Рис. 3-21. Синаптические ионные токи и потенциал реверсии, измененный в условиях фиксации
Рис. 3-21. Синаптические ионные токи и потенциал реверсии, измененный в условиях фиксации
Слайд 60Взаимное влияние ВПСП и ТПСП
Если на мембране клетки активируются одновременно возбуждающие и
Взаимное влияние ВПСП и ТПСП
Если на мембране клетки активируются одновременно возбуждающие и

Слайд 61Рис. 3-22. ВПСП и ТПСП на мембране клетки взаимно влияют друг на
Рис. 3-22. ВПСП и ТПСП на мембране клетки взаимно влияют друг на
Слайд 62Лиганд-управляемые ионные каналы. Их токи
На примере двух различных типов лигандуправляемых ионных каналов
Лиганд-управляемые ионные каналы. Их токи
На примере двух различных типов лигандуправляемых ионных каналов

Слайд 63Рис. 3-23. Примеры токов АЦХ-управляемые ионные каналы.
А - ток через одиночный ионный
Рис. 3-23. Примеры токов АЦХ-управляемые ионные каналы.
А - ток через одиночный ионный
Слайд 64Patch-clamp для изучения канальных токов синапса
При применении метода patch-clamp в конфигурации outside-out,
Patch-clamp для изучения канальных токов синапса
При применении метода patch-clamp в конфигурации outside-out,

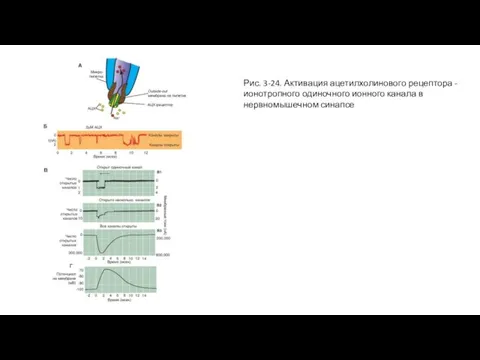
Слайд 65Рис. 3-24. Активация ацетилхолинового рецептора - ионотропного одиночного ионного канала в нервномышечном
Рис. 3-24. Активация ацетилхолинового рецептора - ионотропного одиночного ионного канала в нервномышечном
Слайд 66Соединения с синаптическим влиянием
Наряду с трансмиттером, другие соединения тоже могут влиять на
Соединения с синаптическим влиянием
Наряду с трансмиттером, другие соединения тоже могут влиять на

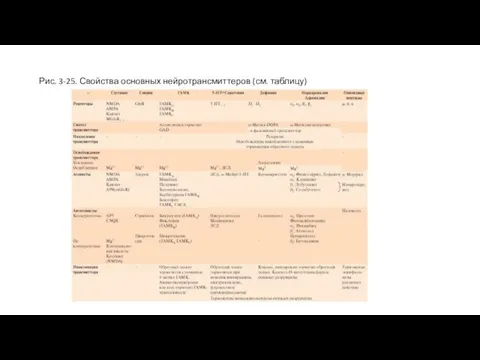
Слайд 67Рис. 3-25. Свойства основных нейротрансмиттеров (см. таблицу)
Рис. 3-25. Свойства основных нейротрансмиттеров (см. таблицу)
 Биосфера
Биосфера ЛЕКЦИЯ 3 ВИРУСОЛОГИЯ НОВ
ЛЕКЦИЯ 3 ВИРУСОЛОГИЯ НОВ Божья коровка. 2 часть
Божья коровка. 2 часть Процессы видообразования
Процессы видообразования Физиология питания
Физиология питания Строение клетки. Роберт Гук
Строение клетки. Роберт Гук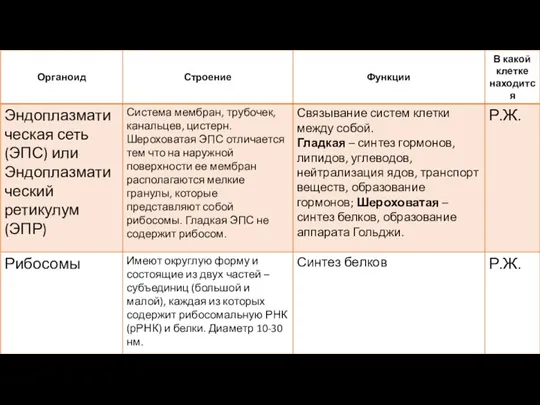 Органоиды клетки. Таблица
Органоиды клетки. Таблица Морфологические структуры грибов
Морфологические структуры грибов нервная ткань
нервная ткань Однодольные и двудольные растения. Бинарная номенклатура
Однодольные и двудольные растения. Бинарная номенклатура Энтомология
Энтомология Отношения организмов в сообществе
Отношения организмов в сообществе Виды цветочного оформления
Виды цветочного оформления Строение и образ жизни кишечнополостных
Строение и образ жизни кишечнополостных Fruits
Fruits Рыба язь
Рыба язь Занятие 5. Проводящие пучки
Занятие 5. Проводящие пучки Жизненный цикл Лягушачьей многоустки – Polystoma integerrimum
Жизненный цикл Лягушачьей многоустки – Polystoma integerrimum онтогенез урок
онтогенез урок Популяционная динамика бортевой пчелы (Apis mellifera mellifera L.) в заповеднике Шульган-Таш за полвека наблюдений
Популяционная динамика бортевой пчелы (Apis mellifera mellifera L.) в заповеднике Шульган-Таш за полвека наблюдений Пищеварительная система Пищеварительный тракт
Пищеварительная система Пищеварительный тракт Отряды птиц
Отряды птиц Строение клеток прокариот и эукариот
Строение клеток прокариот и эукариот Роль теории эволюции в науке
Роль теории эволюции в науке Ванадий в организме человека
Ванадий в организме человека Управление мужскими желаниями
Управление мужскими желаниями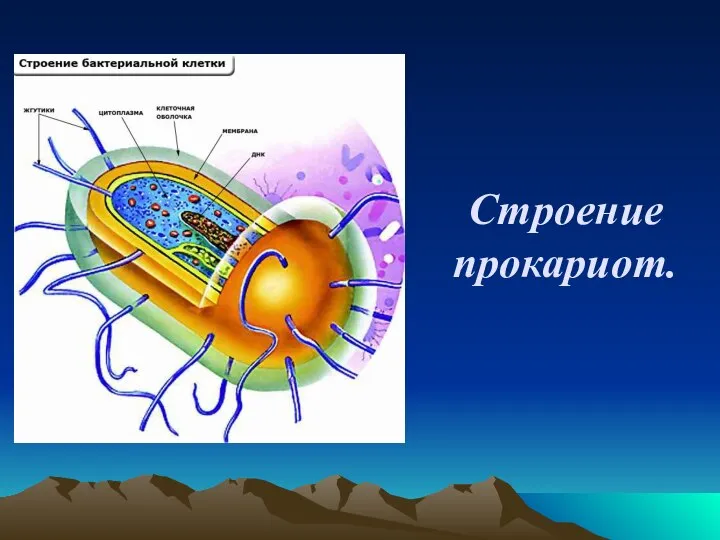 Строение прокариот
Строение прокариот Презентация на тему Деревья которые нас удивляют
Презентация на тему Деревья которые нас удивляют