Супрамолекулярная химия в живой природе. Тетрапиррольные макроциклы. Кобаламин. Гемоглобин. ДНК. РНК. Синтез белка
- Супрамолекулярная химия в живой природе. Тетрапиррольные макроциклы. Кобаламин. Гемоглобин. ДНК. РНК. Синтез белка

Содержание
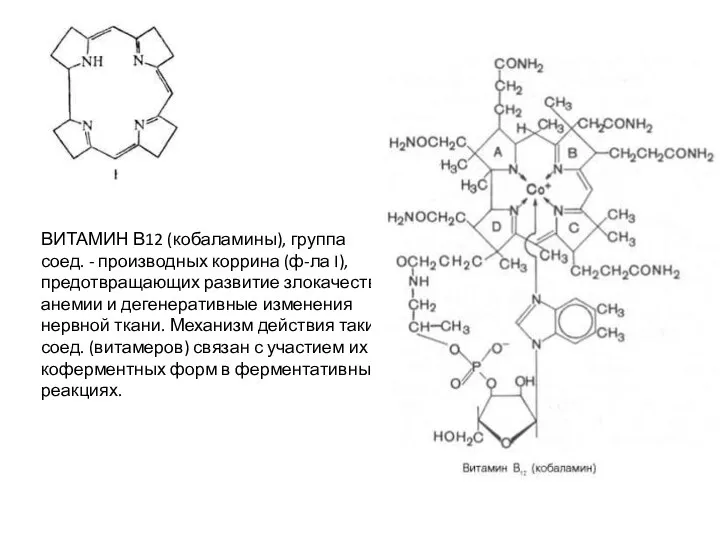
- 2. ВИТАМИН В12 (кобаламины), группа соед. - производных коррина (ф-ла I), предотвращающих развитие злокачеств. анемии и дегенеративные
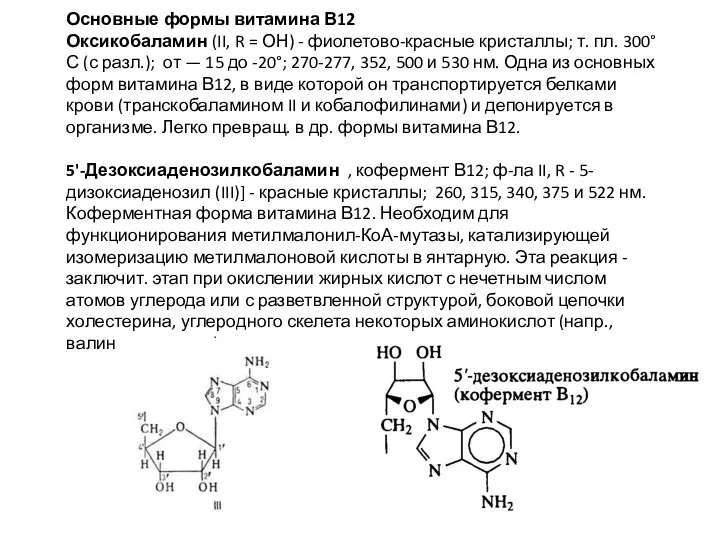
- 4. Основные формы витамина В12 Оксикобаламин (II, R = ОН) - фиолетово-красные кристаллы; т. пл. 300°С (с
- 5. Метилкобаламин (СН3-В12; ф-ла II, R = СН3) - в организме находится в меньших кол-вах, чем др.
- 6. В молекуле витамина В12 центральный атом кобальта соединен с атомами азота четырех восстановленных пиррольных колец, образующих
- 7. Биологическая роль. В 20-х годах ХХ века было замечено, что смертельно опасную пернициозную анемию можно вылечить
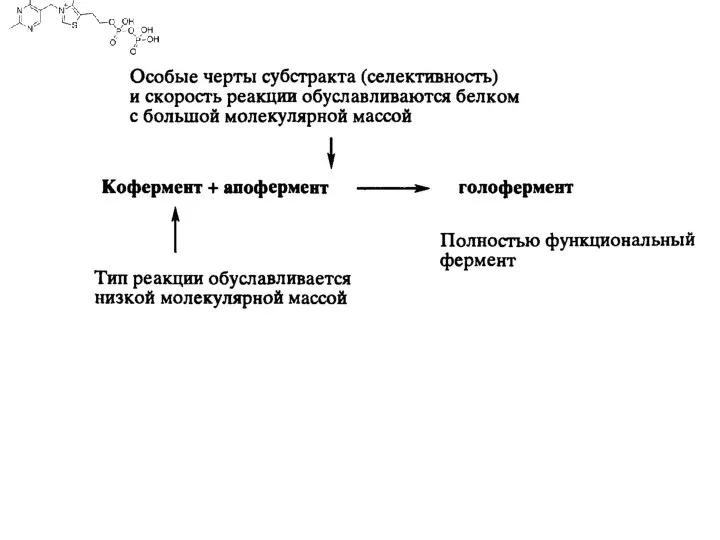
- 8. Для понимания биохимической роли кофермента В12 сначала необходимо дать определение понятию «кофермент». Коферменты, или коэнзимы —
- 10. Биологическая роль. Выявлены ферментные системы, в составе которых в качестве простетической группы участвуют не свободный витамин
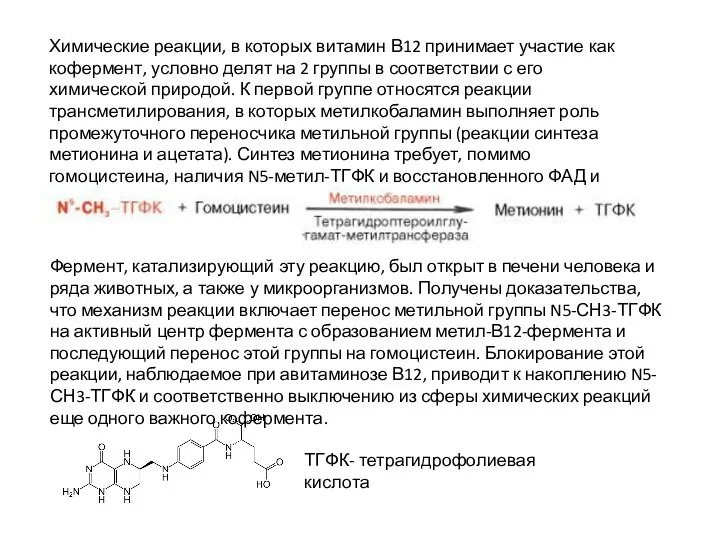
- 11. Химические реакции, в которых витамин В12 принимает участие как кофермент, условно делят на 2 группы в
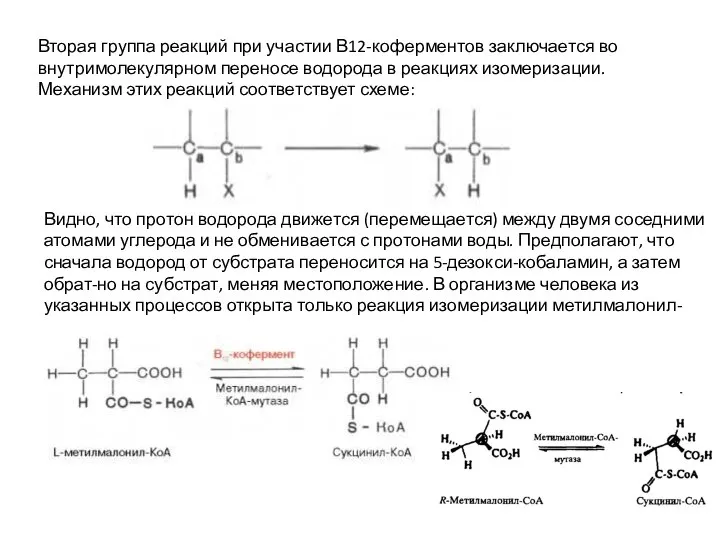
- 12. Вторая группа реакций при участии В12-коферментов заключается во внутримолекулярном переносе водорода в реакциях изомеризации. Механизм этих
- 13. Следует подчеркнуть, что реакция изомеризации метилмалонил-КоА требует наличия 5'-дезоксиаденозилкобаламина в качестве кофермента, в то время как
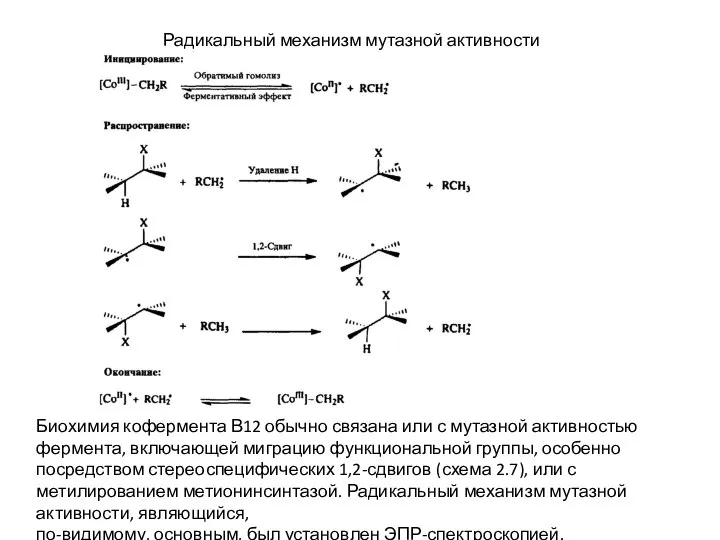
- 14. Радикальный механизм мутазной активности Биохимия кофермента В12 обычно связана или с мутазной активностью фермента, включающей миграцию
- 15. Распространение в природе и суточная потребность. Витамин В12 является единственным витамином, синтез которого осуществляется исключительно микроорганизмами;
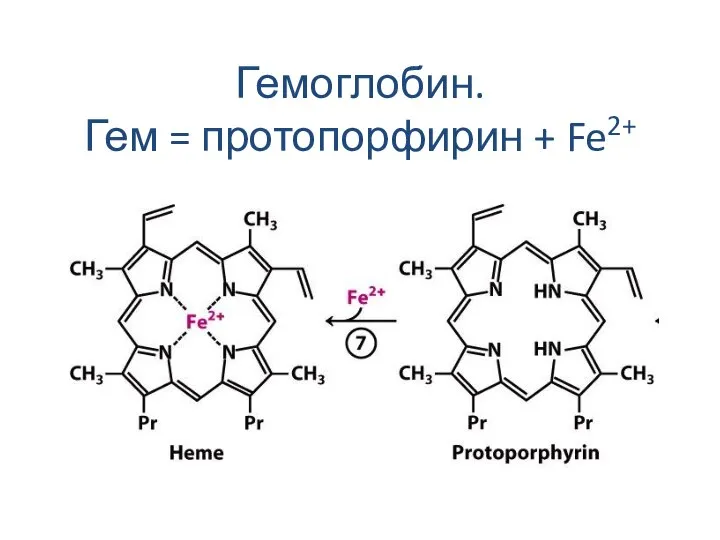
- 16. Гемоглобин. Гем = протопорфирин + Fe2+
- 17. Жизненно необходимой составляющей метаболизма высших организмов является кислород воздуха. Он метаболически окисляет сахара, такие, как глюкоза
- 18. Для того чтобы аэробные организмы могли использовать реакционноспособ- ный О2 в процессе, противоположном фотосинтезу, необходимо поглотить
- 20. Главной проблемой реакции металлических центров с О2 является его склонность к необратимому окислительному взаимодействию с металлическим
- 21. В соответствии с моделью Вайсса Fe3+-центр является Низкоспиновым/ Это должно приводить к парамагнетизму как от одного
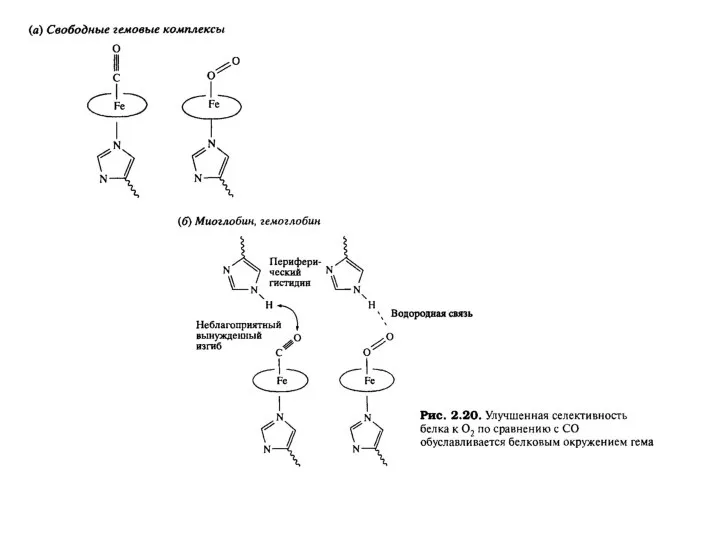
- 23. Окисленный гем и его непосредственное белковое окружение, включающее стабилизирующие водородные связи.
- 24. Хорошо известно, что такие газы, как СО, или легкоадсорбирующиеся соли, например CN-, крайне токсичны. Это происходит
- 26. ДНК, РНК и биосинтез белка. Нуклеиновые кислоты как самособирающиеся супрамолекулярные системы. Ученые никогда не переставали удивляться
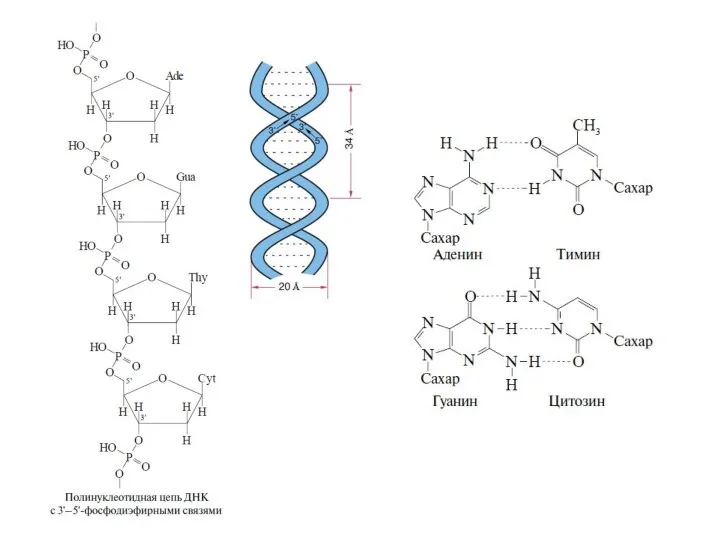
- 27. ДНК (дезоксирибонуклеиновая кислота) хорошо известна как молекула, несущая в себе всю генетическую информацию, необходимую для построения
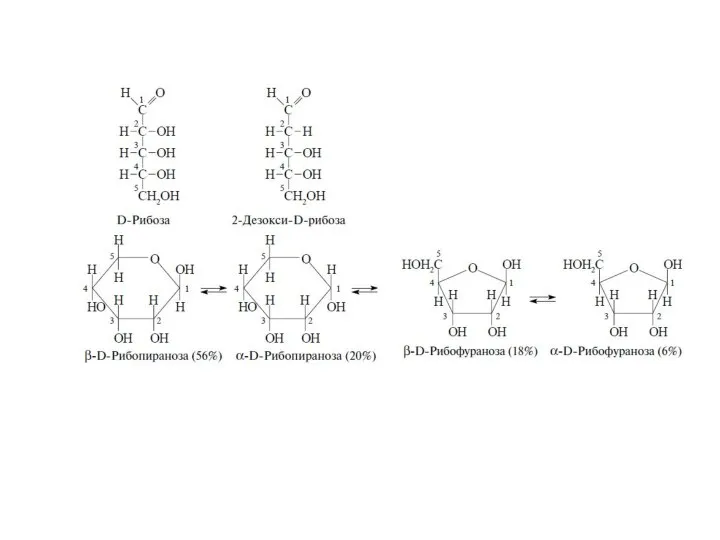
- 29. Как и большинство других моносахаридов, рибоза существует не в линейной, а в циклической полуацетальной форме. Последняя
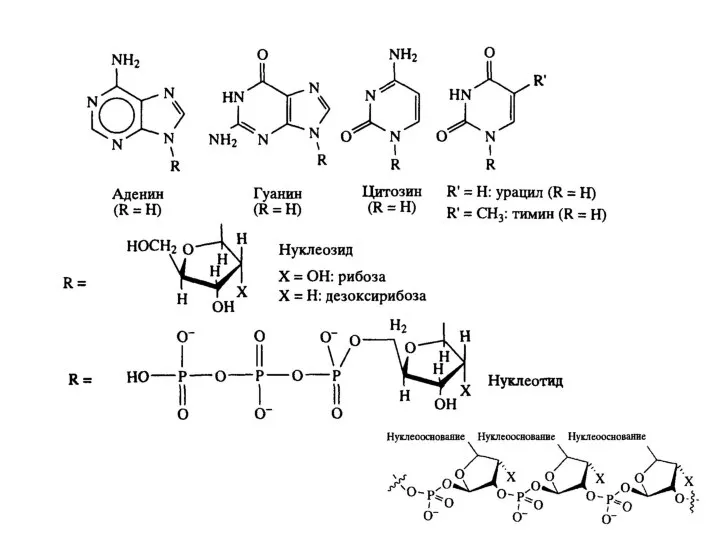
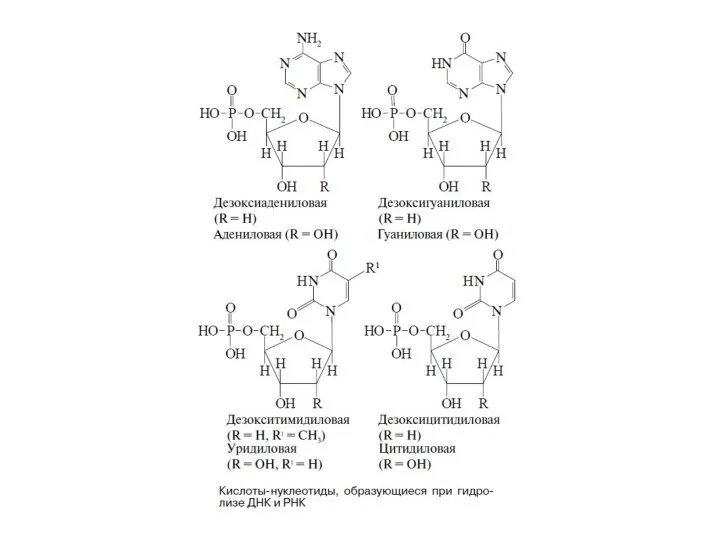
- 31. С помощью ферментативного гидролиза нуклеиновых кислот было установлено, что элементарными звеньями, из которых они состоят, являются
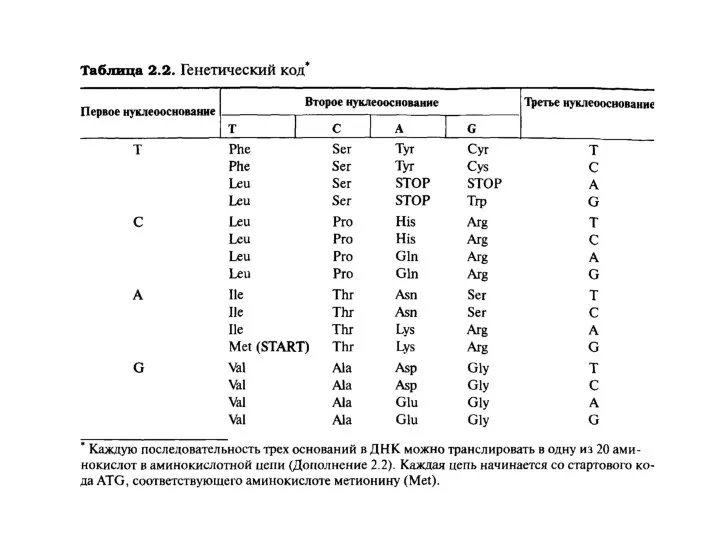
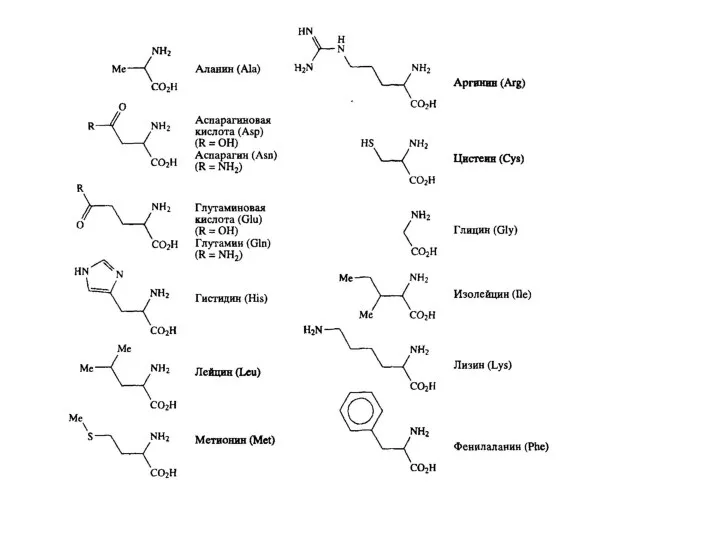
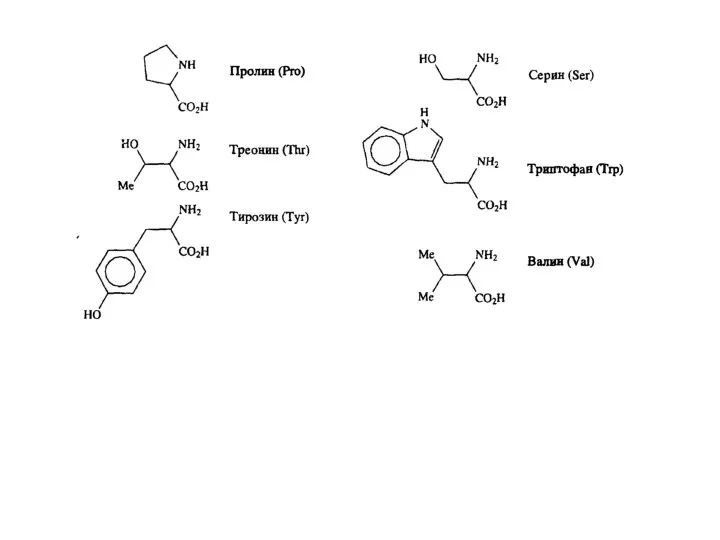
- 39. Биосинтез белка происходит в два этапа. В первый этап входит транскрипция и процессинг РНК, второй этап
- 40. Между транскрипцией и трансляцией молекула мРНК претерпевает ряд последовательных изменений, которые обеспечивают созревание функционирующей матрицы для
- 41. Сплайсинг (от англ. splice — сращивать или склеивать концы чего-либо) — процесс вырезания определенных нуклеотидных последовательностей
- 42. Экзон [от английского ex(pressi)on — выражение, выразительность], участок гена (ДНК) эукариот, несущий генетическую информацию, кодирующую синтез
- 43. Трансляция У прокариот мРНК может считываться рибосомами в аминокислотную последовательность белков сразу после транскрипции, а у
- 45. Скачать презентацию
Слайд 2ВИТАМИН В12 (кобаламины), группа соед. - производных коррина (ф-ла I), предотвращающих развитие
ВИТАМИН В12 (кобаламины), группа соед. - производных коррина (ф-ла I), предотвращающих развитие
Слайд 4Основные формы витамина В12
Оксикобаламин (II, R = ОН) - фиолетово-красные кристаллы; т.
Основные формы витамина В12
Оксикобаламин (II, R = ОН) - фиолетово-красные кристаллы; т.
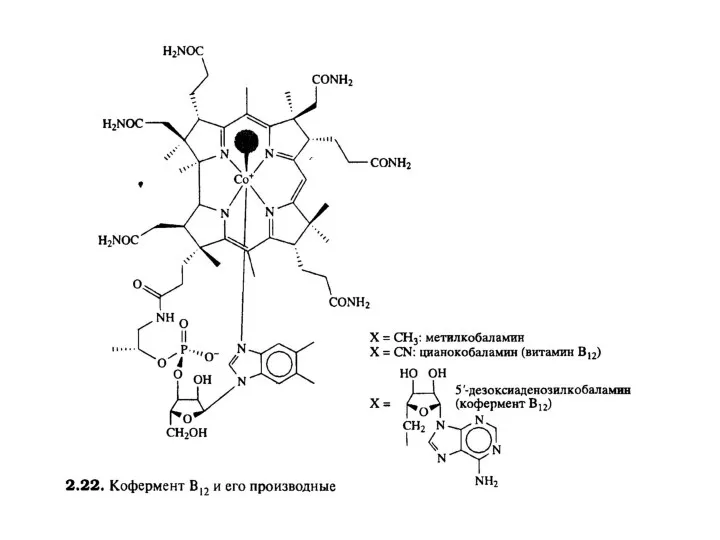
Слайд 5Метилкобаламин (СН3-В12; ф-ла II, R = СН3) - в организме находится в
Метилкобаламин (СН3-В12; ф-ла II, R = СН3) - в организме находится в

Слайд 6В молекуле витамина В12 центральный атом кобальта соединен с атомами азота четырех
В молекуле витамина В12 центральный атом кобальта соединен с атомами азота четырех

Слайд 7Биологическая роль.
В 20-х годах ХХ века было замечено, что смертельно опасную пернициозную
Биологическая роль.
В 20-х годах ХХ века было замечено, что смертельно опасную пернициозную

Слайд 8Для понимания биохимической роли кофермента В12 сначала необходимо дать определение понятию «кофермент».
Для понимания биохимической роли кофермента В12 сначала необходимо дать определение понятию «кофермент».

Слайд 10Биологическая роль. Выявлены ферментные системы, в составе которых в качестве простетической группы
Биологическая роль. Выявлены ферментные системы, в составе которых в качестве простетической группы

Слайд 11Химические реакции, в которых витамин В12 принимает участие как кофермент, условно делят
Химические реакции, в которых витамин В12 принимает участие как кофермент, условно делят
Слайд 12Вторая группа реакций при участии В12-коферментов заключается во внутримолекулярном переносе водорода в
Вторая группа реакций при участии В12-коферментов заключается во внутримолекулярном переносе водорода в
Слайд 13Следует подчеркнуть, что реакция изомеризации метилмалонил-КоА требует наличия 5'-дезоксиаденозилкобаламина в качестве кофермента,
Следует подчеркнуть, что реакция изомеризации метилмалонил-КоА требует наличия 5'-дезоксиаденозилкобаламина в качестве кофермента,

Слайд 14Радикальный механизм мутазной активности
Биохимия кофермента В12 обычно связана или с мутазной активностью
Радикальный механизм мутазной активности
Биохимия кофермента В12 обычно связана или с мутазной активностью
Слайд 15Распространение в природе и суточная потребность. Витамин В12 является единственным витамином, синтез
Распространение в природе и суточная потребность. Витамин В12 является единственным витамином, синтез

Слайд 16Гемоглобин.
Гем = протопорфирин + Fe2+
Гемоглобин.
Гем = протопорфирин + Fe2+
Слайд 17Жизненно необходимой составляющей метаболизма высших
организмов является кислород воздуха. Он метаболически окисляет
Жизненно необходимой составляющей метаболизма высших
организмов является кислород воздуха. Он метаболически окисляет

Слайд 18Для того чтобы аэробные организмы могли использовать реакционноспособ-
ный О2 в процессе,
Для того чтобы аэробные организмы могли использовать реакционноспособ-
ный О2 в процессе,
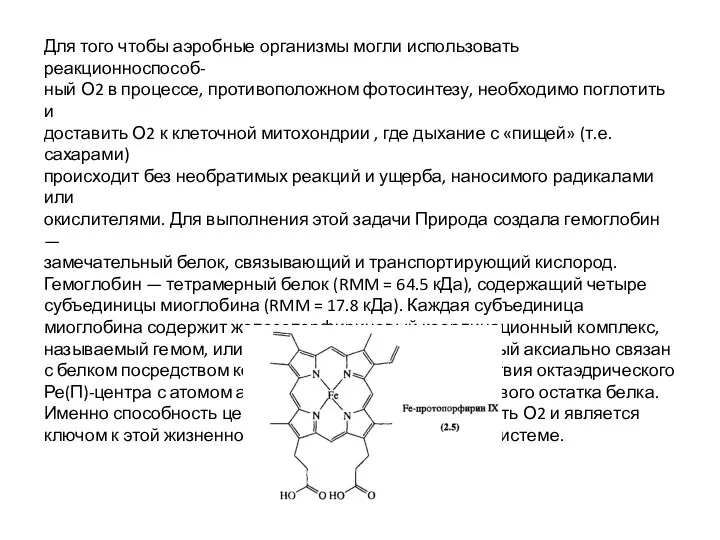
Слайд 20Главной проблемой реакции металлических центров с О2 является его
склонность к необратимому
Главной проблемой реакции металлических центров с О2 является его
склонность к необратимому

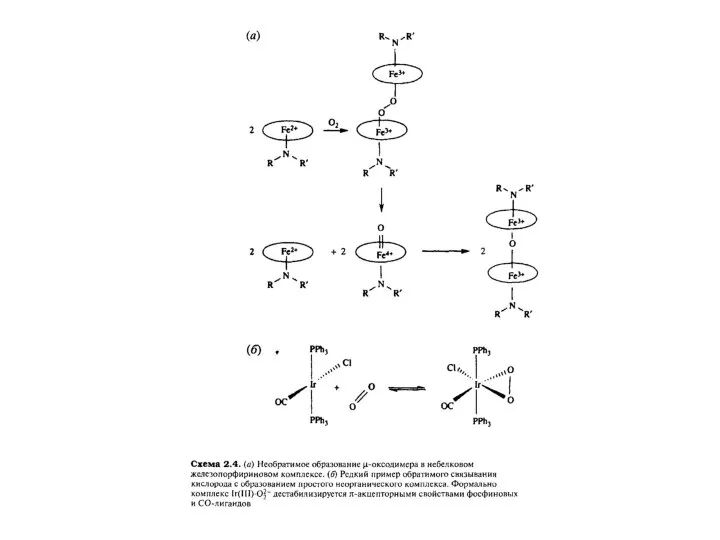
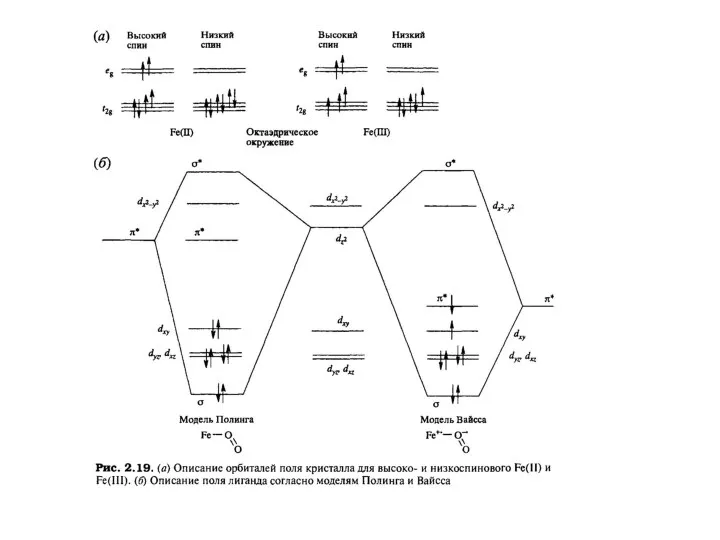
Слайд 21В соответствии с моделью Вайсса Fe3+-центр является
Низкоспиновым/ Это должно приводить к
В соответствии с моделью Вайсса Fe3+-центр является
Низкоспиновым/ Это должно приводить к

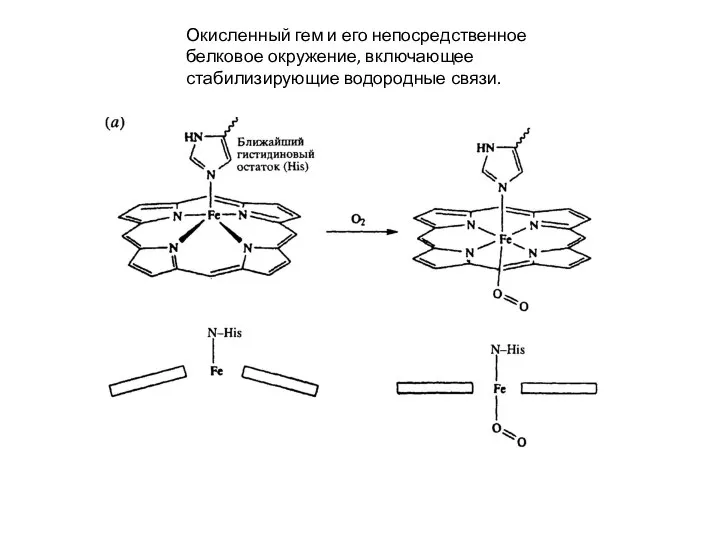
Слайд 23Окисленный гем и его непосредственное белковое окружение, включающее
стабилизирующие водородные связи.
Окисленный гем и его непосредственное белковое окружение, включающее
стабилизирующие водородные связи.
Слайд 24Хорошо известно, что такие газы, как СО, или легкоадсорбирующиеся соли,
например CN-,
Хорошо известно, что такие газы, как СО, или легкоадсорбирующиеся соли,
например CN-,

Слайд 26ДНК, РНК и биосинтез белка.
Нуклеиновые кислоты как самособирающиеся супрамолекулярные системы.
Ученые никогда
ДНК, РНК и биосинтез белка.
Нуклеиновые кислоты как самособирающиеся супрамолекулярные системы.
Ученые никогда

Слайд 27ДНК (дезоксирибонуклеиновая кислота) хорошо известна как молекула, несущая в себе всю генетическую
ДНК (дезоксирибонуклеиновая кислота) хорошо известна как молекула, несущая в себе всю генетическую

Слайд 29Как и большинство других моносахаридов, рибоза существует не в линейной, а в

Слайд 31С помощью ферментативного гидролиза нуклеиновых кислот было установлено, что элементарными звеньями, из
С помощью ферментативного гидролиза нуклеиновых кислот было установлено, что элементарными звеньями, из


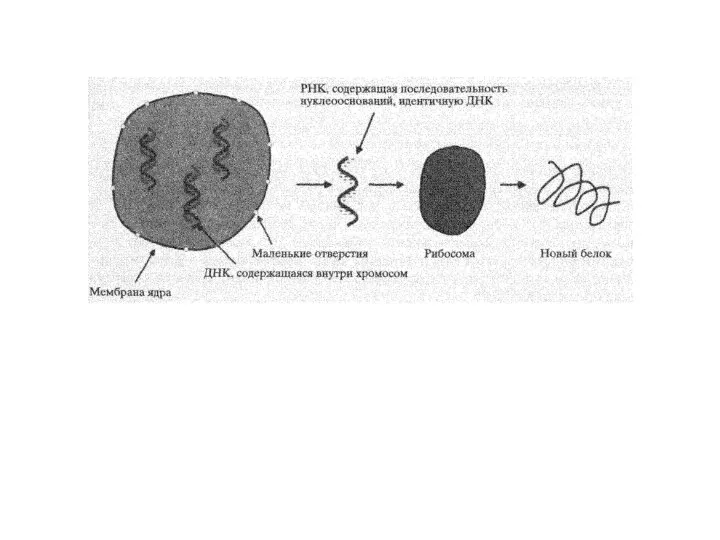
Слайд 39Биосинтез белка происходит в два этапа. В первый этап входит транскрипция и
Биосинтез белка происходит в два этапа. В первый этап входит транскрипция и
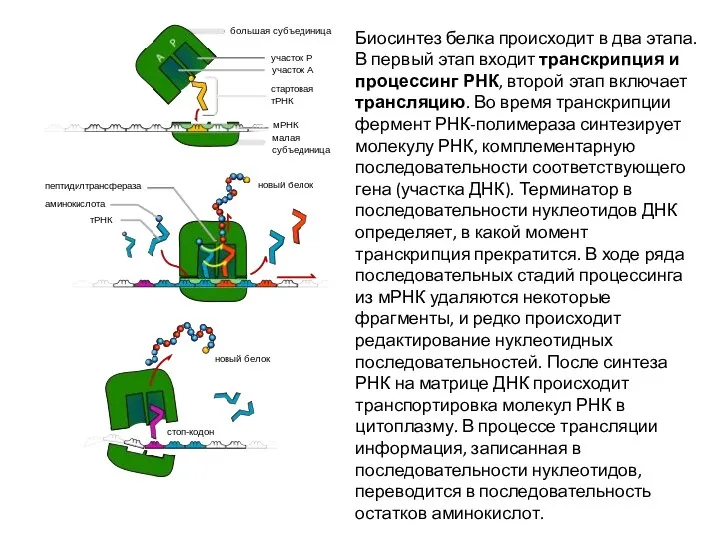
Слайд 40Между транскрипцией и трансляцией молекула мРНК претерпевает ряд последовательных изменений, которые обеспечивают
Между транскрипцией и трансляцией молекула мРНК претерпевает ряд последовательных изменений, которые обеспечивают
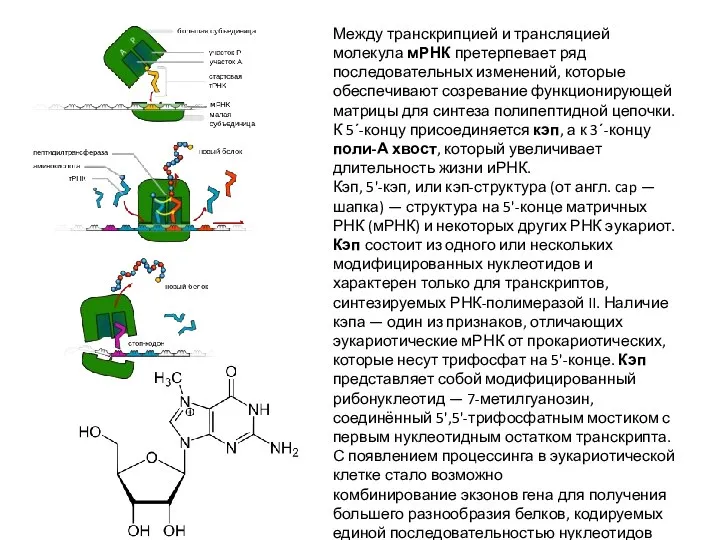
Слайд 41Сплайсинг (от англ. splice — сращивать или склеивать концы чего-либо) — процесс
Сплайсинг (от англ. splice — сращивать или склеивать концы чего-либо) — процесс
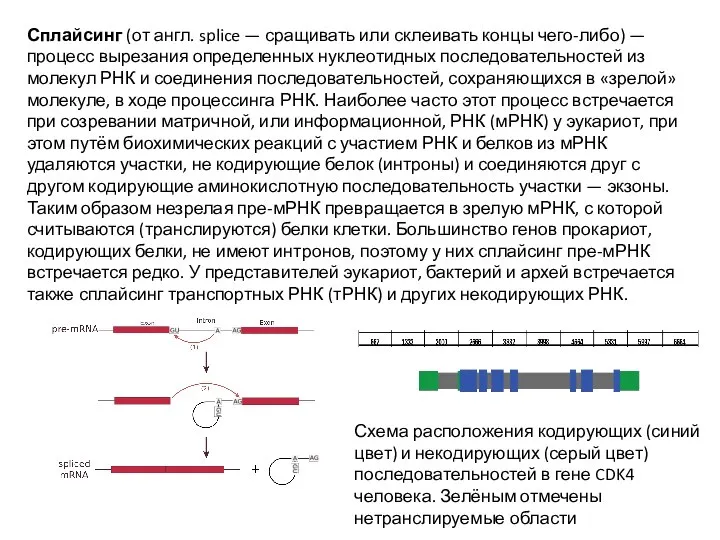
Слайд 42Экзон [от английского ex(pressi)on — выражение, выразительность], участок гена (ДНК) эукариот, несущий генетическую
Экзон [от английского ex(pressi)on — выражение, выразительность], участок гена (ДНК) эукариот, несущий генетическую
![Экзон [от английского ex(pressi)on — выражение, выразительность], участок гена (ДНК) эукариот, несущий](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1165626/slide-41.jpg)
Слайд 43Трансляция
У прокариот мРНК может считываться рибосомами в аминокислотную последовательность белков сразу после
Трансляция
У прокариот мРНК может считываться рибосомами в аминокислотную последовательность белков сразу после

 Соцветия
Соцветия Хромосомная и генная инженерия
Хромосомная и генная инженерия Природа вокруг нас
Природа вокруг нас Общая характеристика птиц
Общая характеристика птиц Природа. Царство растений
Природа. Царство растений Нуклеиновые кислоты НК
Нуклеиновые кислоты НК Электронный плакат, птица Гаичка
Электронный плакат, птица Гаичка Изучение возрастной изменчивости
Изучение возрастной изменчивости Органы растений. Строение побега
Органы растений. Строение побега Опорно - двигательная система. Интересные факты
Опорно - двигательная система. Интересные факты Размножение самшита полуодревесневшими черенками с использованием регуляторов роста
Размножение самшита полуодревесневшими черенками с использованием регуляторов роста Вечерница малая
Вечерница малая Щука - речная хищница
Щука - речная хищница Освещенность
Освещенность Лисица
Лисица Строение, функции и возрастные особенности развития опорно-двигательного аппарата. Лекция 2
Строение, функции и возрастные особенности развития опорно-двигательного аппарата. Лекция 2 Животный мир тундры
Животный мир тундры Агрессивные иммигранты в мире растений
Агрессивные иммигранты в мире растений Торф. Виды торфа
Торф. Виды торфа Презентация на тему Гипертония
Презентация на тему Гипертония  Альвеолярно-капиллярная мембрана
Альвеолярно-капиллярная мембрана Высшие споровые растения: мхи
Высшие споровые растения: мхи Опорно-двигательный аппарат. Часть 2
Опорно-двигательный аппарат. Часть 2 Царство грибы
Царство грибы Презентация на тему МОНОГИБРИДНОЕ СКРЕЩИВАНИЕ
Презентация на тему МОНОГИБРИДНОЕ СКРЕЩИВАНИЕ  Из гусеницы в бабочку. 3 класс
Из гусеницы в бабочку. 3 класс Презентация на тему Естественный отбор. Формы естественного отбора
Презентация на тему Естественный отбор. Формы естественного отбора  Вирусы, как форма жизни
Вирусы, как форма жизни