- Транскрипция. Исследования транскрипции

Содержание
- 2. Исследования транскрипции В 1961 г. Франсуа Жакоб, Жак Моно предположили существование матричной РНК посредника. Триплетность кода:
- 3. Метод связывания триплетов В 1964 г. Ниренберг и Ледер разработали данный метод для установления точной последовательности
- 4. Использование повторяющихся кополимеров Гобинд Корана синтезировал протяженные молекулы РНК с заданной последовательностью, многократно повторяющейся. Из 2,
- 5. Wobble hypothesis В 1966 г. Ф.Крик сформулировал гипотезу качания (wobble hypothesis). Предположил, что для комплементации с
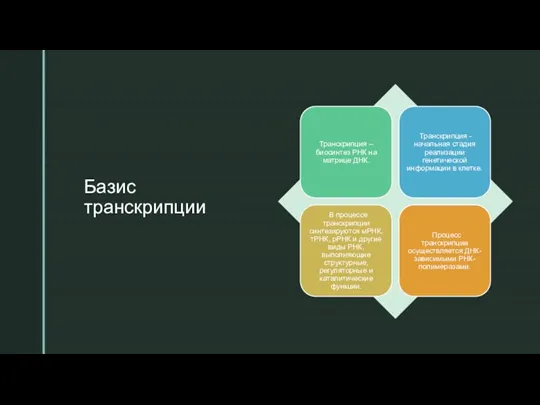
- 6. Базис транскрипции
- 7. РНК-полимераза РНК-полимераза - фермент, участвующий в синтезе РНК на ДНК-матрице. Использует в качестве субстрата рибонуклеозидтрифосфаты (NTP),
- 8. Виды РНК-полимераз
- 9. Базис транскрипции Единица транскрипции – транскриптон, участок ДНК, ограниченный промотором (зоной начала транскрипции) и терминатором (зоной
- 10. Последовательность транскрипции
- 11. Основные принципы транскрипции
- 13. Процессинг 1.Кэпирование 5'-конца – присоединении к этому концу иРНК так называемой шапочки (кэп-структуры – 7-метилгуанозина); 2.Полиаденилирование
- 14. Кэпирование Функции кэпа: защита 5'-конца транскрипта от экзонуклеаз; участие в сплайсинге; экспорт мРНК из ядра; участие
- 15. Сплайсинг В зависимости от специфичности механихма сплайсинга, интроны подразделяются на группы: Интроны, которые сами обладают ферментативной
- 16. Сплайсосома
- 17. Схема сплайсинга с сплайсосомой
- 18. Альтернативный сплайсинг
- 20. Скачать презентацию
Слайд 2Исследования транскрипции
В 1961 г. Франсуа Жакоб, Жак Моно предположили существование матричной РНК
Исследования транскрипции
В 1961 г. Франсуа Жакоб, Жак Моно предположили существование матричной РНК

Слайд 3Метод связывания триплетов
В 1964 г. Ниренберг и Ледер разработали данный метод для
Метод связывания триплетов
В 1964 г. Ниренберг и Ледер разработали данный метод для

Слайд 4Использование повторяющихся кополимеров
Гобинд Корана синтезировал протяженные молекулы РНК с заданной последовательностью, многократно
Использование повторяющихся кополимеров
Гобинд Корана синтезировал протяженные молекулы РНК с заданной последовательностью, многократно

Слайд 5Wobble hypothesis
В 1966 г. Ф.Крик сформулировал гипотезу качания (wobble hypothesis).
Предположил, что для
Wobble hypothesis
В 1966 г. Ф.Крик сформулировал гипотезу качания (wobble hypothesis).
Предположил, что для

Слайд 6Базис транскрипции
Базис транскрипции
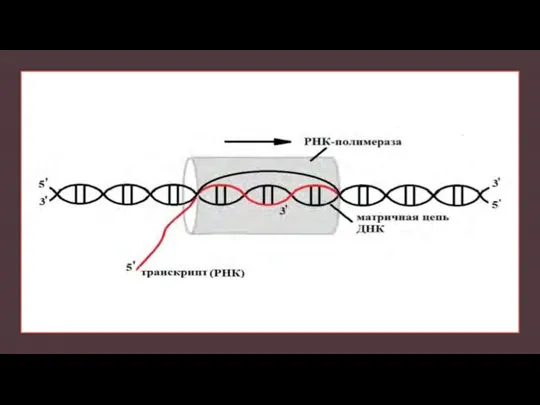
Слайд 7РНК-полимераза
РНК-полимераза - фермент, участвующий в синтезе РНК на ДНК-матрице.
Использует в качестве субстрата
РНК-полимераза
РНК-полимераза - фермент, участвующий в синтезе РНК на ДНК-матрице.
Использует в качестве субстрата

Слайд 8Виды РНК-полимераз
Виды РНК-полимераз

Слайд 9Базис транскрипции
Единица транскрипции – транскриптон, участок ДНК, ограниченный промотором (зоной начала транскрипции)
Базис транскрипции
Единица транскрипции – транскриптон, участок ДНК, ограниченный промотором (зоной начала транскрипции)

Слайд 10Последовательность транскрипции
Последовательность транскрипции

Слайд 11Основные принципы транскрипции
Основные принципы транскрипции

Слайд 13Процессинг
1.Кэпирование 5'-конца – присоединении к этому концу иРНК так называемой шапочки (кэп-структуры
Процессинг
1.Кэпирование 5'-конца – присоединении к этому концу иРНК так называемой шапочки (кэп-структуры

Слайд 14Кэпирование
Функции кэпа: защита 5'-конца транскрипта от экзонуклеаз; участие в сплайсинге; экспорт мРНК
Кэпирование
Функции кэпа: защита 5'-конца транскрипта от экзонуклеаз; участие в сплайсинге; экспорт мРНК

Слайд 15Сплайсинг
В зависимости от специфичности механихма сплайсинга, интроны подразделяются на группы:
Интроны, которые сами
Сплайсинг
В зависимости от специфичности механихма сплайсинга, интроны подразделяются на группы:
Интроны, которые сами

Слайд 16Сплайсосома
Сплайсосома
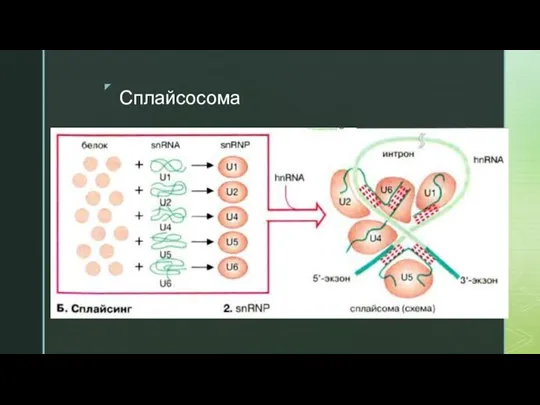
Слайд 17Схема сплайсинга с сплайсосомой
Схема сплайсинга с сплайсосомой

Слайд 18Альтернативный сплайсинг
Альтернативный сплайсинг

 Нервная система трубчатого типа
Нервная система трубчатого типа Жгутики эукариот
Жгутики эукариот нервная ткань
нервная ткань Презентация на тему Учение Ч. Дарвина об искусственном отборе
Презентация на тему Учение Ч. Дарвина об искусственном отборе  Приспособленность организмов к среде обитания, как результат действия естественного отбора
Приспособленность организмов к среде обитания, как результат действия естественного отбора Гормональная регуляция обменных процессов: белков, воды, натрия, калия, кальция, фосфатов. Лекция № 22
Гормональная регуляция обменных процессов: белков, воды, натрия, калия, кальция, фосфатов. Лекция № 22 Бесполое размножение
Бесполое размножение Саранча - насекомые из семейства Acridiodea
Саранча - насекомые из семейства Acridiodea Микробы вокруг нас
Микробы вокруг нас Движение крови по сосудам
Движение крови по сосудам Динамика численности рыжего соснового пилильщика
Динамика численности рыжего соснового пилильщика Учение В.И.вернадского о биосфере. 9 класс
Учение В.И.вернадского о биосфере. 9 класс Анатомно-физиологические особенности человека в подростковом возрасте
Анатомно-физиологические особенности человека в подростковом возрасте 03. Одномембранные органоиды. Цитотплазма ЛИЦЕЙ
03. Одномембранные органоиды. Цитотплазма ЛИЦЕЙ 12. Покрытосеменные растения (1)
12. Покрытосеменные растения (1) Шипов лес - самый крупный дубовый лес в степи
Шипов лес - самый крупный дубовый лес в степи Витамины и здоровье
Витамины и здоровье Таблиця Різноманість птахів
Таблиця Різноманість птахів Грибы
Грибы Учёт численности сообщества настоящих тюленей семейства Phocidae в заливе Пильтун (остров Сахалин) c применением БПЛА в 2020 г
Учёт численности сообщества настоящих тюленей семейства Phocidae в заливе Пильтун (остров Сахалин) c применением БПЛА в 2020 г Fruits
Fruits Близнецовый метод изучения генетики
Близнецовый метод изучения генетики Принцип восприятия стимула волосковыми рецепторами
Принцип восприятия стимула волосковыми рецепторами Класс Рептилии, 7 класс
Класс Рептилии, 7 класс Викторина по биологии "Что? Где? Когда?"
Викторина по биологии "Что? Где? Когда?" Становление человеческого общества
Становление человеческого общества Палеозой эрасы
Палеозой эрасы Регенера́ция (восстановление)
Регенера́ция (восстановление)