- Транспозиция и сайт - специфическая рекомбинация

Содержание
- 2. Принципиальная схема транспозиции Транспозон должен содержать на своих концах инвертированные повторы (IR). Разрезание донорной и акцепторной
- 3. Вырезание транспозонов Еще до разрезания должен сформироваться так называемый синапс. Это происходит за счет формирования димера
- 4. Встраивание транспозонов Свободные концы транспозона при помощи транспозазы атакуют участок акцепторной ДНК с такой же последовательностью,
- 5. IS-элементы IS (Insersion Sequences)-элементы – самые маленькие и простые бактериальные транспозоны. Их длина обычно в диапазоне
- 6. Перемещение композитных транспозонов IS-элементы в составе композитного транспозона не могут перемещаться по отдельности, потому что: Часто
- 7. Сборка плазмид при помощи транспозонов Множество природных плазмид очевидным образом образовались вследствие активности транспозонов, зачастую –
- 8. Некомпозитные транспозоны Так называются транспозоны, не состоящие из двух одинаковых IS-элементов, и в то же время
- 9. Репликативная транспозиция на примере Tn3 Транспозон выпетливается, и транспозаза делает одноцепочечные разрывы в местах, обозначенных кружками.
- 10. Репликативная транспозиция на примере Tn3 Образуется вот такая жуткая штука из двух молекул ДНК, в каждой
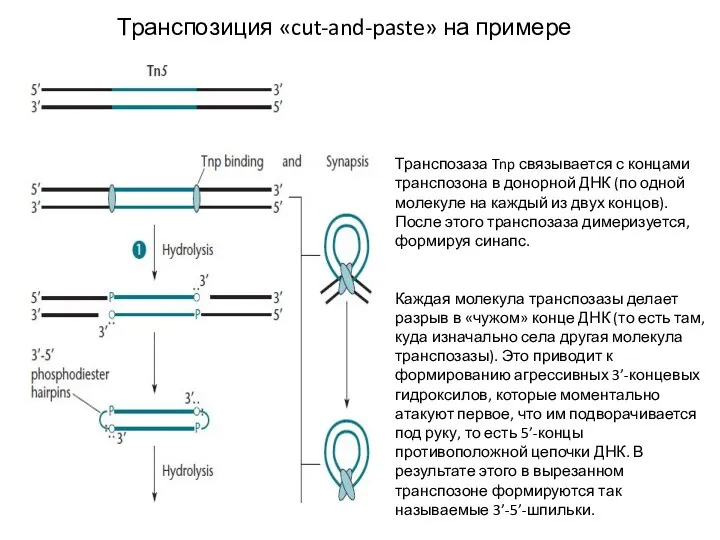
- 11. Транспозиция «cut-and-paste» на примере Tn5 Транспозаза Tnp связывается с концами транспозона в донорной ДНК (по одной
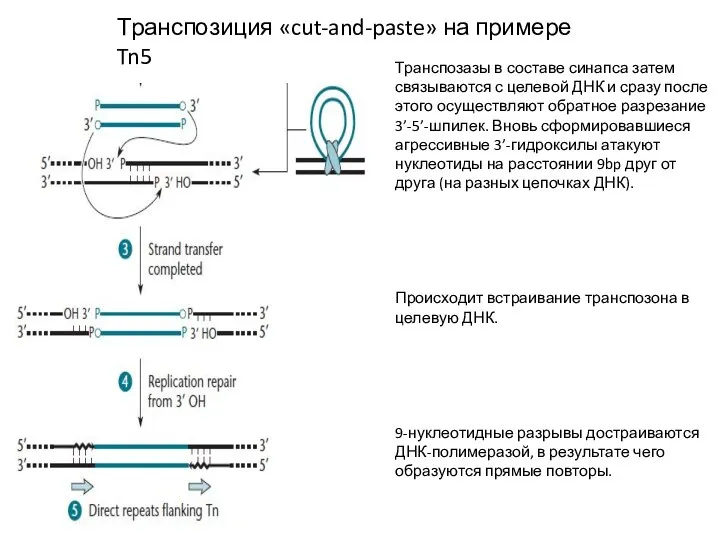
- 12. Транспозиция «cut-and-paste» на примере Tn5 Транспозазы в составе синапса затем связываются с целевой ДНК и сразу
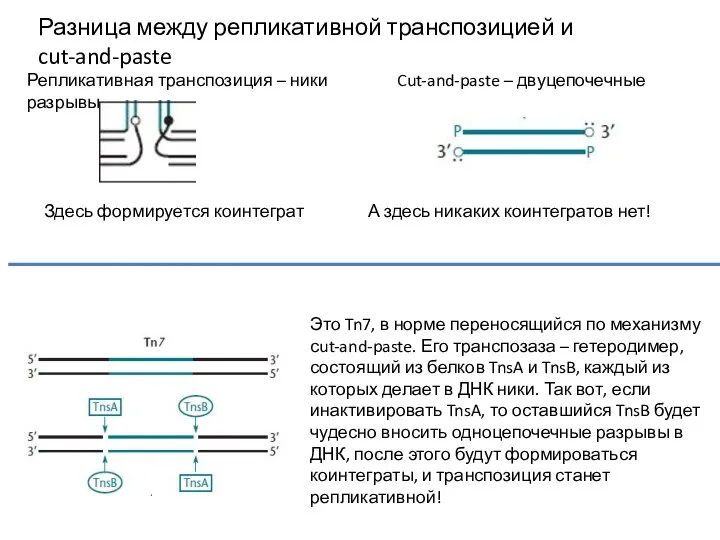
- 13. Разница между репликативной транспозицией и cut-and-paste Репликативная транспозиция – ники Cut-and-paste – двуцепочечные разрывы Здесь формируется
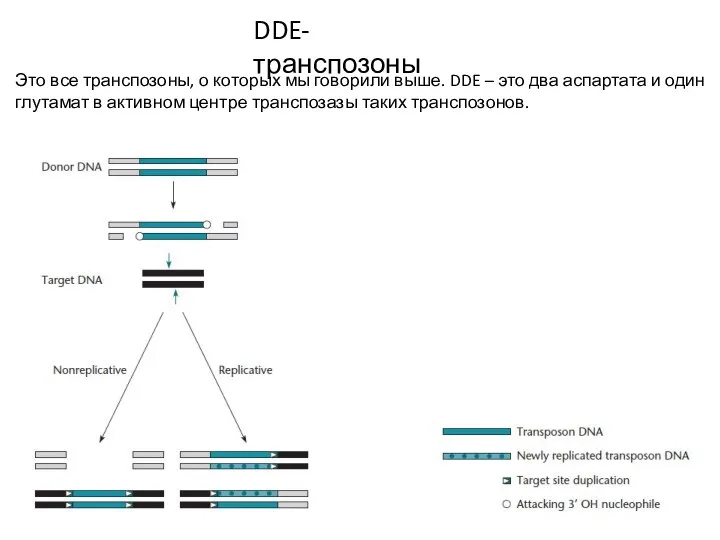
- 14. DDE-транспозоны Это все транспозоны, о которых мы говорили выше. DDE – это два аспартата и один
- 15. А бывают и другие транспозоны!
- 16. Сайт-специфическая рекомбинация Происходит по коротким участкам строго определенной последовательности. Хотя эти участки почти всегда содержат комплементарные
- 17. Интегроны Так называются участки крупных транспозонов, в которых собраны гены устойчивости к антибиотикам. Такие гены часто
- 18. Интегроны Интегроны встречаются не только в транспозонах, но и в хромосомах. Хороший пример – холерный вибрион,
- 19. ДНК-инвертазы Это почти то же самое, что и резольвазы (о которых мы специально говорить не будем,
- 20. Механизмы работы сайт-специфических рекомбиназ Все сайт-специфические рекомбиназы по механизму действия делятся на две группы: тирозиновые (Y)
- 21. Как работают Y-рекомбиназы (на примере Cre) 4 молекулы фермента связывают 2 рекомбинирующие молекулы, в каждой из
- 22. Как работают Y-рекомбиназы (на примере Cre) 3. В результате атаки нуклеофильными 5’-гидроксилами 3’-фосфорилтирозиновых связей создаются две
- 23. Как работают Y-рекомбиназы (на примере Cre) Изомеризация структуры Холидея каким-то образом подает сигнал двум пока незадействованным
- 24. Как работают S-рекомбиназы (на примере резольвазы транспозона γδ) 4 молекулы фермента связывают две молекулы ДНК по
- 25. Как работают S-рекомбиназы (на примере резольвазы транспозона γδ) В результате переворота происходит сближение фрагментов ДНК, принадлежавших
- 27. Скачать презентацию
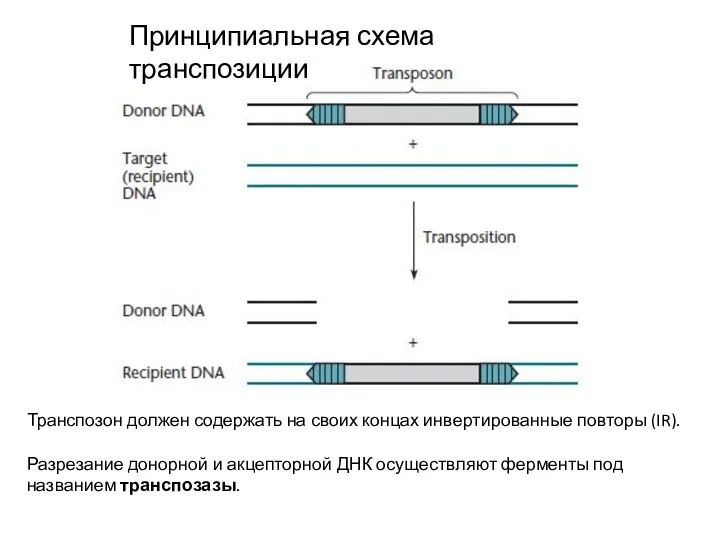
Слайд 2Принципиальная схема транспозиции
Транспозон должен содержать на своих концах инвертированные повторы (IR).
Разрезание донорной
Принципиальная схема транспозиции
Транспозон должен содержать на своих концах инвертированные повторы (IR).
Разрезание донорной
Слайд 3Вырезание транспозонов
Еще до разрезания должен сформироваться так называемый синапс. Это происходит за
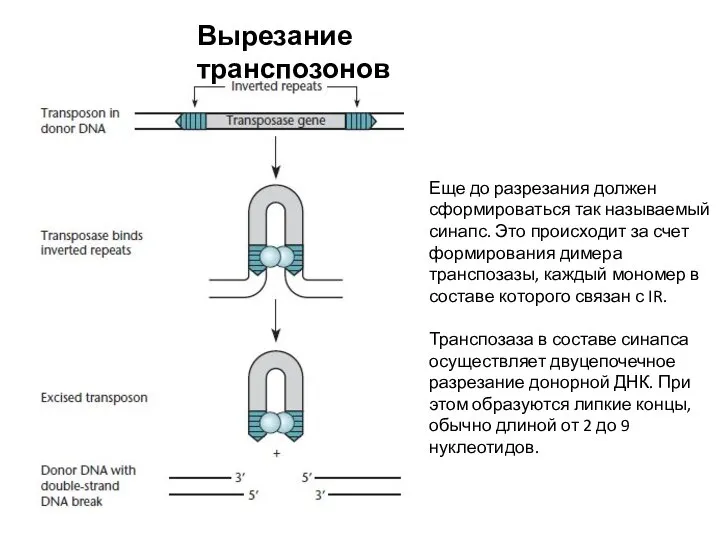
Вырезание транспозонов
Еще до разрезания должен сформироваться так называемый синапс. Это происходит за
Слайд 4Встраивание транспозонов
Свободные концы транспозона при помощи транспозазы атакуют участок акцепторной ДНК с
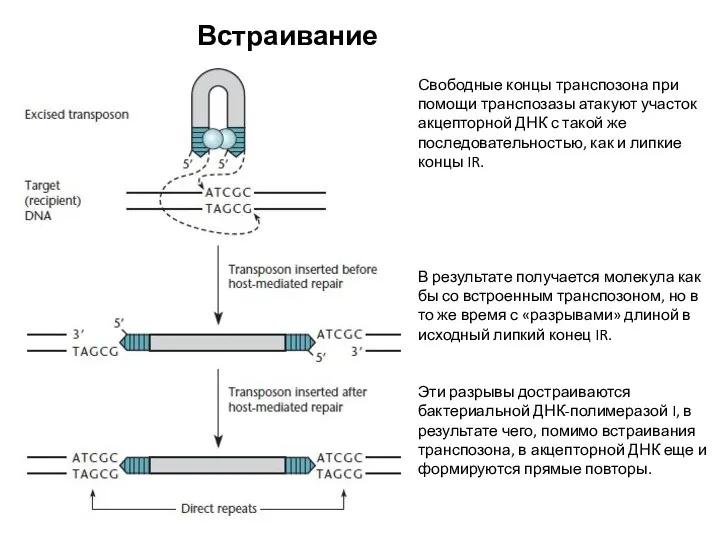
Встраивание транспозонов
Свободные концы транспозона при помощи транспозазы атакуют участок акцепторной ДНК с
Слайд 5IS-элементы
IS (Insersion Sequences)-элементы – самые маленькие и простые бактериальные транспозоны. Их длина
IS-элементы
IS (Insersion Sequences)-элементы – самые маленькие и простые бактериальные транспозоны. Их длина

Слайд 6Перемещение композитных транспозонов
IS-элементы в составе композитного транспозона не могут перемещаться по отдельности,
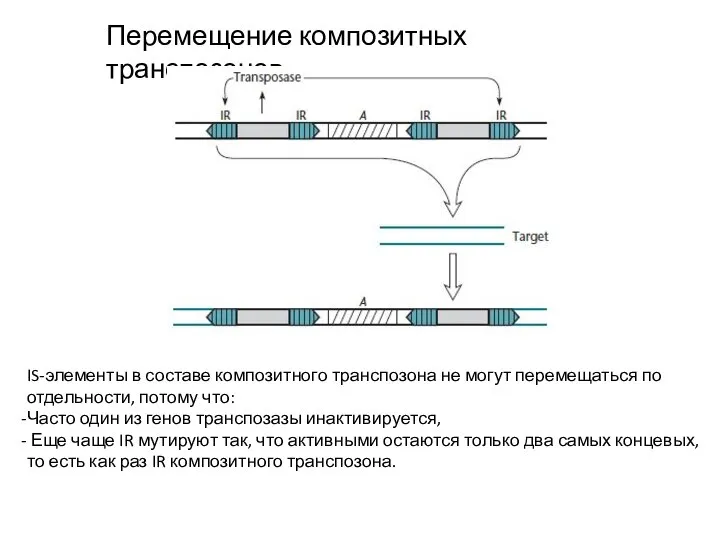
Перемещение композитных транспозонов
IS-элементы в составе композитного транспозона не могут перемещаться по отдельности,
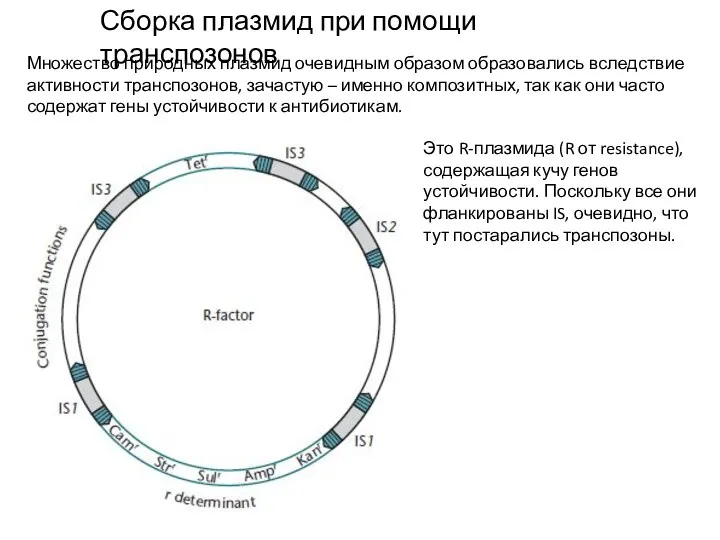
Слайд 7Сборка плазмид при помощи транспозонов
Множество природных плазмид очевидным образом образовались вследствие активности
Сборка плазмид при помощи транспозонов
Множество природных плазмид очевидным образом образовались вследствие активности
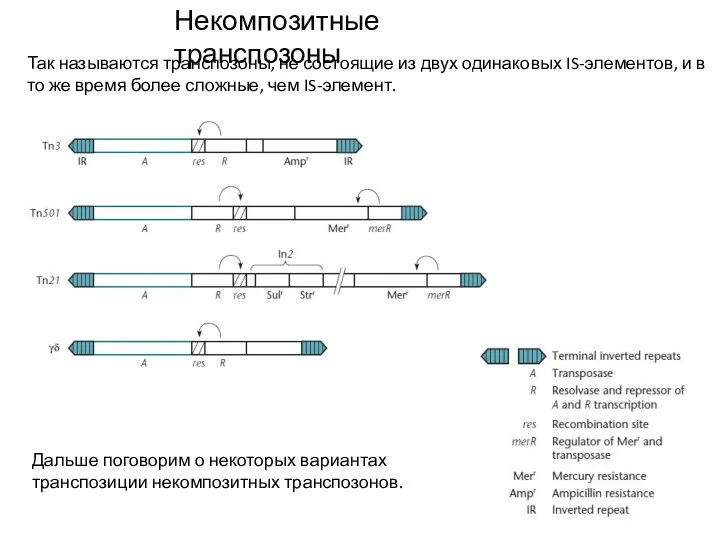
Слайд 8Некомпозитные транспозоны
Так называются транспозоны, не состоящие из двух одинаковых IS-элементов, и в
Некомпозитные транспозоны
Так называются транспозоны, не состоящие из двух одинаковых IS-элементов, и в
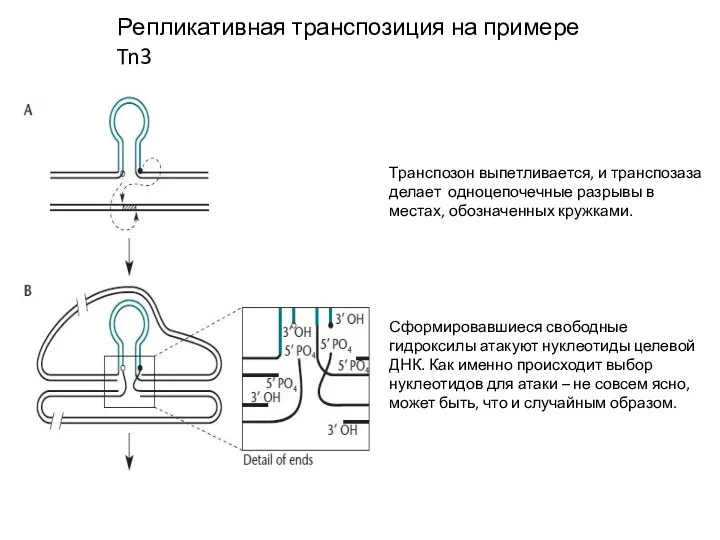
Слайд 9Репликативная транспозиция на примере Tn3
Транспозон выпетливается, и транспозаза делает одноцепочечные разрывы в
Репликативная транспозиция на примере Tn3
Транспозон выпетливается, и транспозаза делает одноцепочечные разрывы в
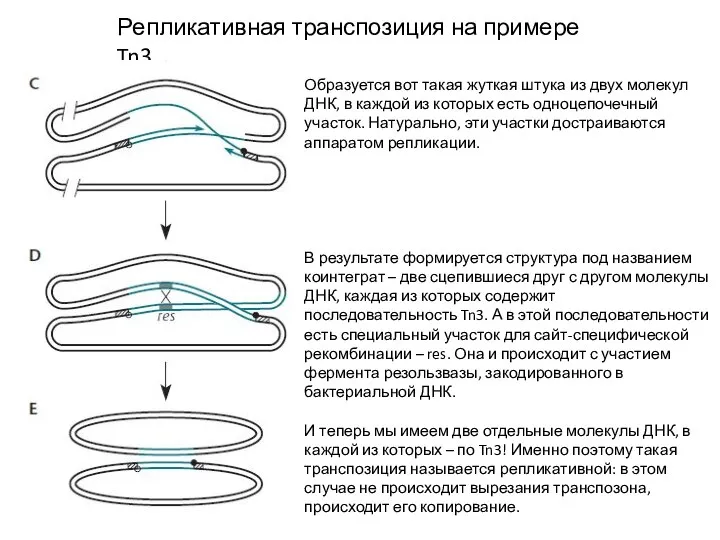
Слайд 10Репликативная транспозиция на примере Tn3
Образуется вот такая жуткая штука из двух молекул
Репликативная транспозиция на примере Tn3
Образуется вот такая жуткая штука из двух молекул
Слайд 11Транспозиция «cut-and-paste» на примере Tn5
Транспозаза Tnp связывается с концами транспозона в донорной
Транспозиция «cut-and-paste» на примере Tn5
Транспозаза Tnp связывается с концами транспозона в донорной
Слайд 12Транспозиция «cut-and-paste» на примере Tn5
Транспозазы в составе синапса затем связываются с целевой
Транспозиция «cut-and-paste» на примере Tn5
Транспозазы в составе синапса затем связываются с целевой
Слайд 13Разница между репликативной транспозицией и cut-and-paste
Репликативная транспозиция – ники Cut-and-paste – двуцепочечные
Разница между репликативной транспозицией и cut-and-paste
Репликативная транспозиция – ники Cut-and-paste – двуцепочечные
Слайд 14DDE-транспозоны
Это все транспозоны, о которых мы говорили выше. DDE – это два
DDE-транспозоны
Это все транспозоны, о которых мы говорили выше. DDE – это два
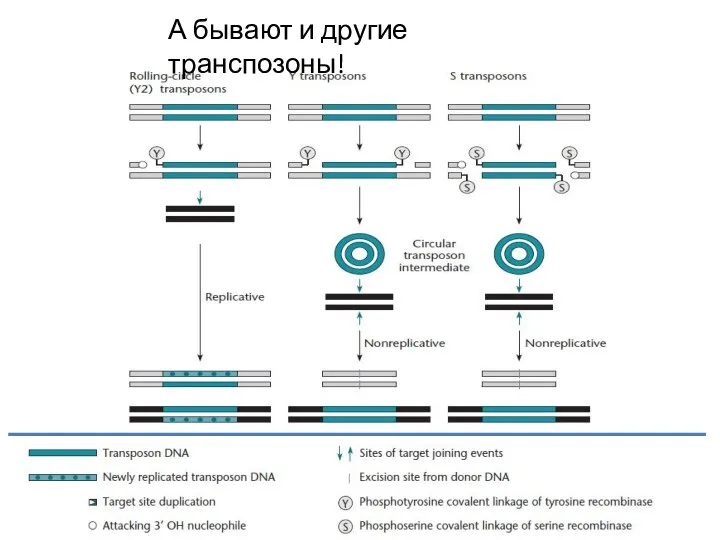
Слайд 15А бывают и другие транспозоны!
А бывают и другие транспозоны!
Слайд 16Сайт-специфическая рекомбинация
Происходит по коротким участкам строго определенной последовательности.
Хотя эти участки почти всегда
Сайт-специфическая рекомбинация
Происходит по коротким участкам строго определенной последовательности.
Хотя эти участки почти всегда

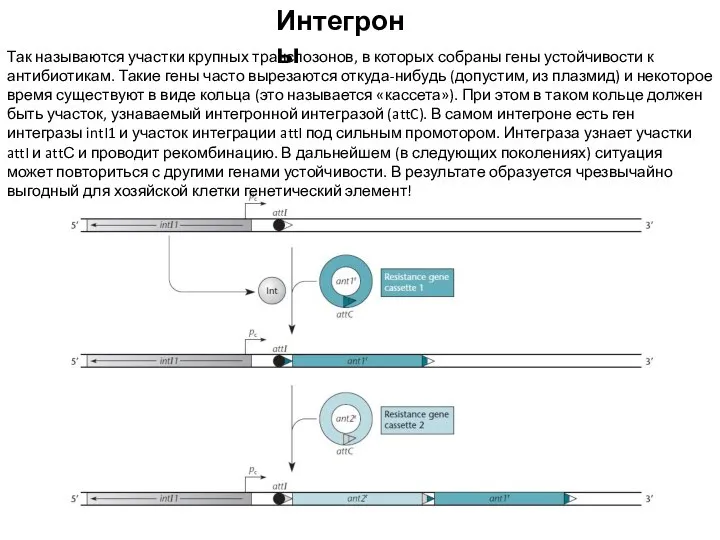
Слайд 17Интегроны
Так называются участки крупных транспозонов, в которых собраны гены устойчивости к антибиотикам.
Интегроны
Так называются участки крупных транспозонов, в которых собраны гены устойчивости к антибиотикам.
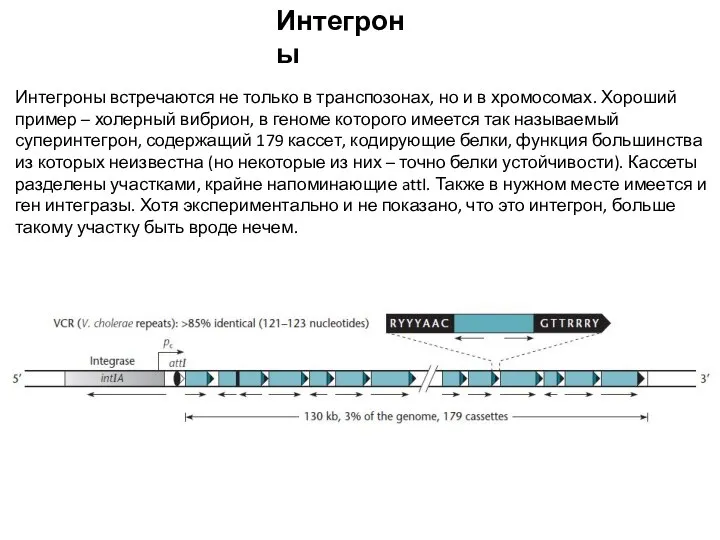
Слайд 18Интегроны
Интегроны встречаются не только в транспозонах, но и в хромосомах. Хороший пример
Интегроны
Интегроны встречаются не только в транспозонах, но и в хромосомах. Хороший пример
Слайд 19ДНК-инвертазы
Это почти то же самое, что и резольвазы (о которых мы специально
ДНК-инвертазы
Это почти то же самое, что и резольвазы (о которых мы специально

Слайд 20Механизмы работы сайт-специфических рекомбиназ
Все сайт-специфические рекомбиназы по механизму действия делятся на две
Механизмы работы сайт-специфических рекомбиназ
Все сайт-специфические рекомбиназы по механизму действия делятся на две

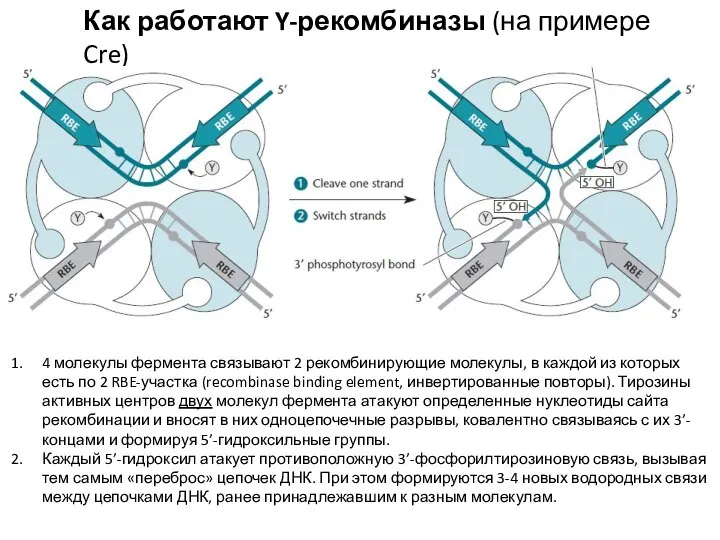
Слайд 21Как работают Y-рекомбиназы (на примере Cre)
4 молекулы фермента связывают 2 рекомбинирующие молекулы,
Как работают Y-рекомбиназы (на примере Cre)
4 молекулы фермента связывают 2 рекомбинирующие молекулы,
Слайд 22Как работают Y-рекомбиназы (на примере Cre)
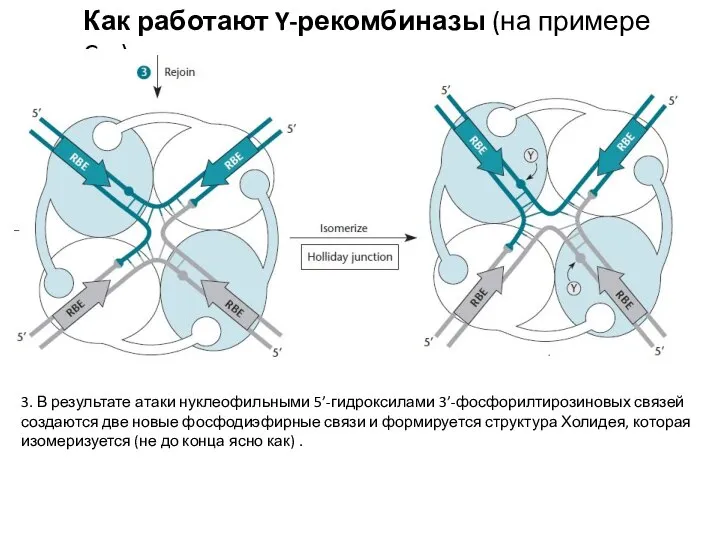
3. В результате атаки нуклеофильными 5’-гидроксилами 3’-фосфорилтирозиновых
Как работают Y-рекомбиназы (на примере Cre)
3. В результате атаки нуклеофильными 5’-гидроксилами 3’-фосфорилтирозиновых
Слайд 23Как работают Y-рекомбиназы (на примере Cre)
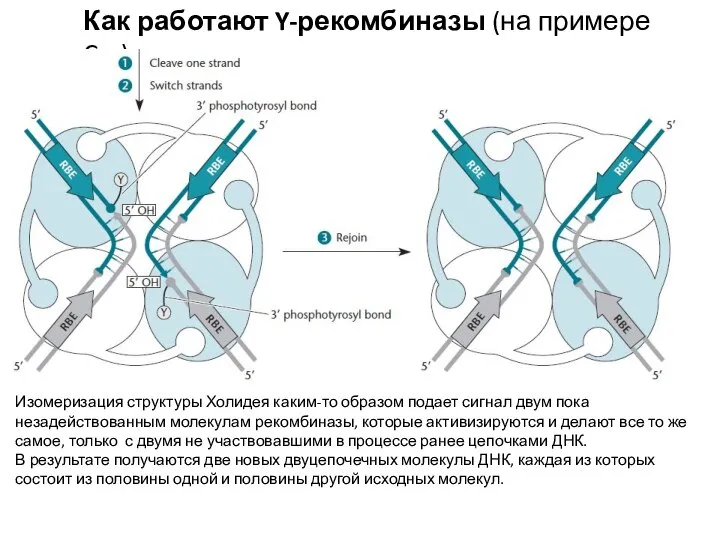
Изомеризация структуры Холидея каким-то образом подает сигнал
Как работают Y-рекомбиназы (на примере Cre)
Изомеризация структуры Холидея каким-то образом подает сигнал
Слайд 24Как работают S-рекомбиназы (на примере резольвазы транспозона γδ)
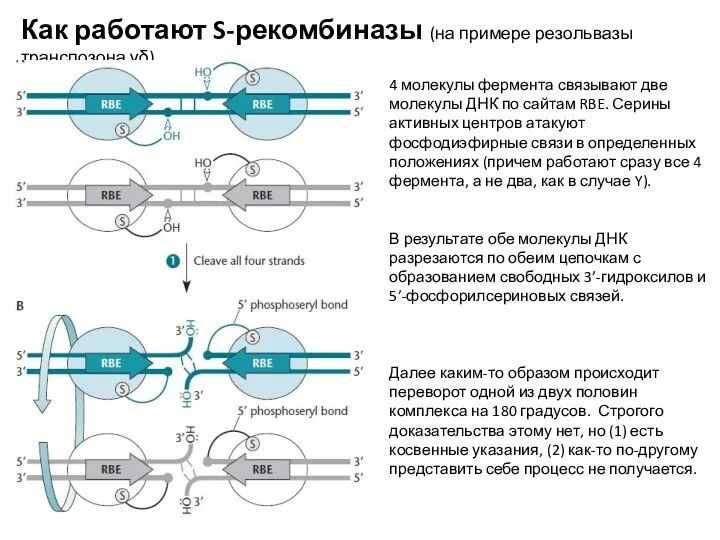
4 молекулы фермента связывают две
Как работают S-рекомбиназы (на примере резольвазы транспозона γδ)
4 молекулы фермента связывают две
Слайд 25Как работают S-рекомбиназы (на примере резольвазы транспозона γδ)
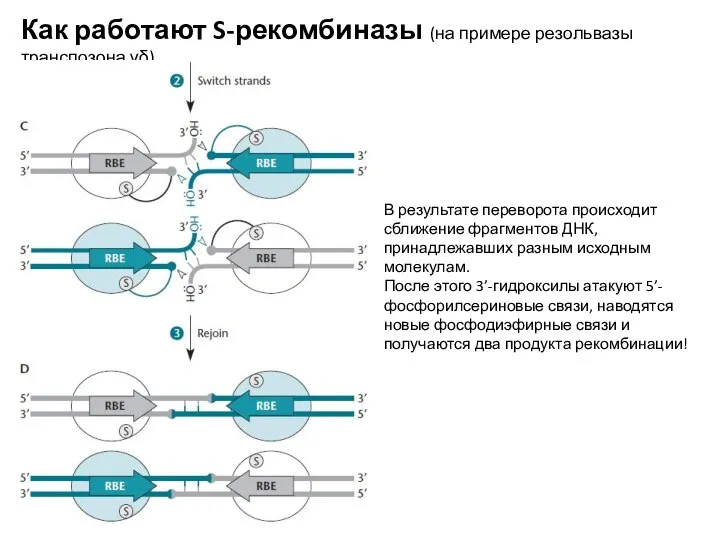
В результате переворота происходит сближение
Как работают S-рекомбиназы (на примере резольвазы транспозона γδ)
В результате переворота происходит сближение
 Отряд Лососеобразные
Отряд Лососеобразные Редкие паукообразные, амфибии и рептилии национального парка Нижняя Кама
Редкие паукообразные, амфибии и рептилии национального парка Нижняя Кама Дыхание и кровообращение
Дыхание и кровообращение Адам ағзасы
Адам ағзасы Ваши права и обязанности
Ваши права и обязанности Красная книга Самарской области
Красная книга Самарской области 832648_№3-realizaciya-nasledstvennoi-informacii
832648_№3-realizaciya-nasledstvennoi-informacii Как растет живое
Как растет живое Насекомые сада и огорода
Насекомые сада и огорода Обмен углеводов. Гликолиз
Обмен углеводов. Гликолиз Мир насекомых. Фоторабота Чумаколенко Тимура
Мир насекомых. Фоторабота Чумаколенко Тимура Мир животных
Мир животных Жизнь паукообразных
Жизнь паукообразных Хвойные растения
Хвойные растения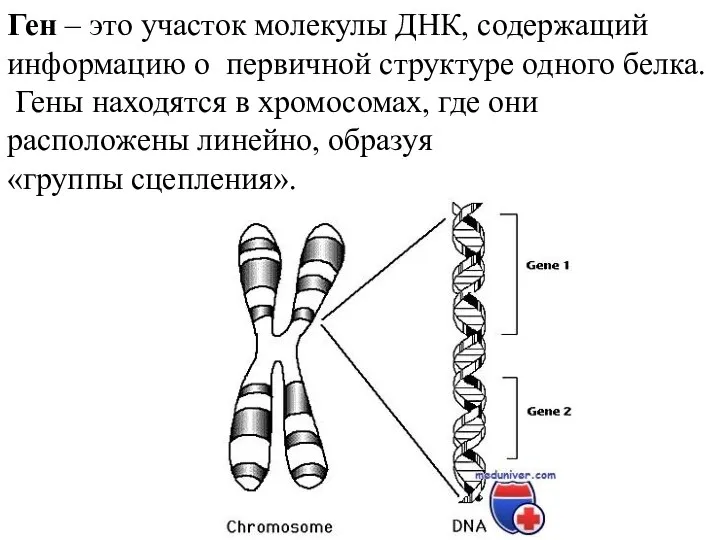 Решение задач по генетике
Решение задач по генетике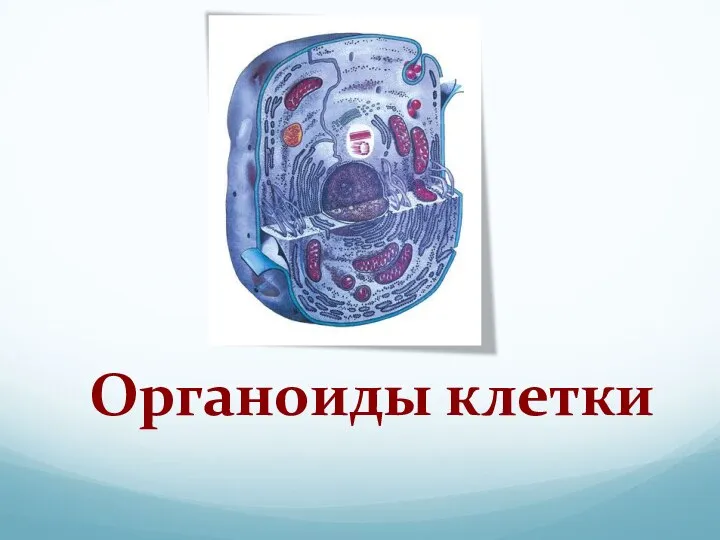 Органоиды клетки
Органоиды клетки Презентация на тему Животные Австралии
Презентация на тему Животные Австралии  Вегетативные органы растений
Вегетативные органы растений Микробиология основных пищевых продуктов
Микробиология основных пищевых продуктов Назовите ученых. Вставьте пропущенные слова
Назовите ученых. Вставьте пропущенные слова Кожа – наружный покровный орган
Кожа – наружный покровный орган Гаметогенез. Биологическое значение оплодотворения
Гаметогенез. Биологическое значение оплодотворения Речная выдра
Речная выдра Редкие растения России
Редкие растения России Деревья
Деревья Обмен веществ и превращение энергии
Обмен веществ и превращение энергии Цикл Кребса. Дыхательная цепь растений
Цикл Кребса. Дыхательная цепь растений Area of distributijn of red wolves
Area of distributijn of red wolves