- Цитология растений. Вегетативные органы растений

Содержание
- 2. Оглавление Цитология растений Ткани растений Вегетативные органы растений Генеративные органы растений
- 3. I. Цитология растений Общая характеристика растительной клетки Цитоплазма Пластиды Строение и функции ядра Вакуоль Клеточная оболочка
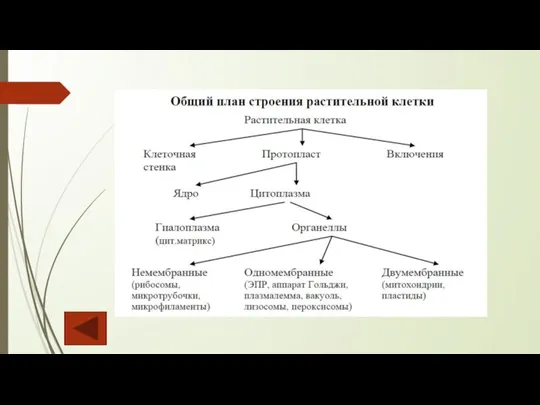
- 4. Общая характеристика растительной клетки Клетка представляет собой основную структурную и функциональную единицу всех живых существ и
- 6. Цитоплазма Цитоплазма — основная часть протопласта клетки, в которой проходят все процессы клеточного обмена веществ, кроме
- 7. Рибосомы Рибосомы — универсальные органеллы, содержащиеся во всех клетках. Функция рибосом — биосинтез белка. Каждая рибосома
- 9. Эндоплазматический ретикулум Эндоплазматический ретикулум (ЭР), или эндоплазматическая сеть, — непрерывно изменяющаяся трехмерная система субмикроскопических цистерн, канальцев
- 11. Аппарат Гольджи Аппарат Гольджи состоит из отдельных диктиосом (телец Гольджи) и пузырьков Гольджи. Диктиосомы представляют собой
- 13. Лизосомы Лизосомы — одномембранные органеллы округлой формы, в матриксе которых содержатся гидролитические ферменты — гидролазы, способные
- 15. Пероксисомы Пероксисомы (микротельца) встречаются в большинстве типов клеток растений и грибов. Они представляют собой мелкие (0,2—1,5
- 17. Митохондрии Митохондрии — постоянно перемещающиеся органеллы округлой, цилиндрической или нитевидной формы. Относительно крупный размер (длина —
- 19. Пластиды Пластиды встречаются только в растительных клетках. Выделяют три типа пластид (хлоро–, лейко– и хромопласты), которые
- 21. Лейкопласты – бесцветные мелкие пластиды, встречающиеся в запасающих органах растений (клубнях, корневищах, семенах и т. д.).
- 23. Схема взаимопревращение пластид
- 24. Строение и функции ядра Ядро — центральная органелла клетки, регулирующая все процессы ее жизнедеятельности. В ядре
- 25. Ядерная оболочка и кариолимфа Ядерная оболочка состоит из двух мембран, между которыми находится перинуклеарное пространство, заполненное
- 26. Ядрышко Ядрышко — плотное сферическое тельце, обычно образующееся в области вторичной перетяжки спутничных хромосом. В ядре
- 27. Хроматин Важнейшая часть ядра — хроматин, который представляет собой молекулы ДНК, заключенные в белковые футляры. Скручивание
- 29. Вакуоль Вакуоли содержатся почти во всех растительных клетках. Они представляют собой полости, заполненные клеточным соком и
- 31. Клеточная оболочка Клеточная оболочка – структурное образование на периферии клетки, придающее ей прочность, сохраняющее ее форму
- 32. Плазмодесмы Плазмодесмы присущи только растительным клеткам. Они представляют собой тонкие цитоплазматические тяжи, соединяющие соседние клетки. В
- 34. Поры Порами называют неутолщенные места оболочки (углубления), на которых отсутствует вторичная оболочка. Они содержат тончайшие отверстия,
- 35. Цитоплазматические включения Включения – это локальная концентрация некоторых продуктов обмена в определенных местах клетки. Крахмальные зерна
- 37. Ткани растений Понятие о ткани. Классификация растительных тканей Образовательные ткани Покровные ткани Всасывающие ткани Ассимиляционные ткани
- 38. Понятие о ткани. Классификация растительных тканей Ткани – это устойчивые, т.е. закономерно повторяющиеся, группы клеток, сходные
- 39. Образовательные ткани У взрослых растений образование новых клеток приурочено к определенным участкам – меристемам. Важная особенность
- 40. Апикальные (верхушечные) располагаются на верхушке побегов и на кончике всех молодых корешков и обеспечивают рост растения
- 42. Покровные ткани Покровные ткани защищают внутренние ткани растений от прямого влияния факторов внешней среды, регулируют испарение
- 44. Перидерма. Продолжительность жизни эпидермиса различна у разных видов и их органов. Например, на листьях и стеблях
- 46. Корка (ритидом) приходит на смену гладкой перидерме у некоторых древесных растений и состоит из чередующихся слоев
- 47. Всасывающие ткани Всасывающие ткани обеспечивают поступление в растение воды и растворенных в ней веществ. Они различны
- 48. Ассимиляционные ткани Главная функция ассимиляционных тканей — фотосинтез. Поэтому их еще называют фотосинтезирующими паренхимами. Располагаются они
- 49. Запасающие ткани К накоплению запасных веществ способны все живые клетки растения. Когда же запасающая функция является
- 50. Аэренхима В растениях довольно часто встречается ткань с большим числом очень крупных межклетников — аэренхима (рис.
- 52. Механические ткани Механические ткани обусловливают прочность растения. Стенки клеток, слагающих эти ткани, утолщены. Механические ткани чаще
- 53. Проводящие ткани Проводящие ткани выполняют функцию транспортировки по растению питательных веществ. Они образуют в теле растения
- 54. Сосуды характерны для покрытосеменных растений. Членики сосудов располагаются один под другим, образуя длинную полую трубку. Основное
- 55. Флоэма – это ткань сосудистых растений, проводящая органические вещества. Первичная флоэма, которую подразделяют на протофлоэму и
- 57. Выделительные ткани Секреторными (выделительными) тканями принято называть структуры, выделяющие терпены, полисахариды, соли, воду и другие вещества.
- 58. Перечислим внутренние секреторные ткани. 1. Секреторные идиобласты – выделительные клетки, рассеянные среди других тканей и накапливающие
- 60. Вегетативные органы растений Корень и корневая система Побег и система побегов
- 61. Корень и корневая система Определение и функции корня Зоны молодого корневого окончания Корневая система
- 62. Определение и функции корня Корень — основной вегетативный орган растения, выполняющий в типичном случае функцию почвенного
- 63. Зоны молодого корневого окончания Нежные клетки апикальной меристемы, словно живым наперстком, всегда прикрыты чехликом. Лишь в
- 64. Зоны молодого корневого окончания Вслед за этой зоной корень остается гладким, но становится светлым и как
- 66. Корневая система Корневая система – это совокупность всех корней одной особи. В состав корневых систем могут
- 68. Побег и система побегов Побег Строение и деятельность апикальной меристемы побега Морфологическое строение стебля Первичное строение
- 69. Побег Побег – это основной орган семенных растений, выполняющий функцию воздушного питания. Состоит из оси (стебля)
- 70. Строение и деятельность апикальной меристемы побега Нарастание побега осуществляется за счет апикальных меристем. Апекс побега отличается
- 71. Морфологическое строение стебля Стебель – это осевая часть побега растений, состоящая из узлов и междоузлий и
- 72. Первичное строение стебля В стебле, имеющем первичное строение, различают эпидермис, первичную кору и стелу (центральный цилиндр).
- 73. Стелярная теория Стела – совокупность первичных проводящих пучков вместе с заключенными между ними прочими тканями и
- 75. Стелярная теория Дальнейшее эволюционное усложнение стелы связано с развитием крупных боковых органов (вай, листьев). В местах
- 76. Типы вторичных утолщений стебля Для большинства двудольных и голосеменных растений характерно вторичное утолщение стеблей. Это изменение
- 77. Лист и его функции Лист – это один из основных органов высших растений, занимающий боковое положение
- 78. Морфологическое строение листа Лист состоит из листовой пластинки и черешка. Место прикрепления листа к стеблю называют
- 79. Различают простые и сложные листья. Простые состоят из одной пластинки. Сложные имеют общий черешок (рахис), к
- 81. Форма, рассечение листовых пластинок простых листьев и листочков сложного листа могут быть весьма разнообразными. Форма основания
- 83. Форма листовой пластинки определяется соотношением ее длины и ширины и положением наиболее широкой ее части.
- 84. Край листа имеет вырезки разной глубины. В тех случаях, если они не заходят глубже чем на
- 85. Листья, вырезы края которых глубже 1/4 ширины полупластинки, называют расчлененными. Расчленение может быть тройчатым, пальчатым и
- 86. Одним из важных морфологических признаков листа является тип жилкования. Жилками условно называют проводящие пучки листа вместе
- 87. Анатомия листа Анатомическое строение листа определяется его главной функцией – фотосинтезом. Поэтому основной тканью листа является
- 88. В листе проводящие ткани формируют закрытые коллатеральные пучки, причем ксилема всегда повернута к верхней стороне, а
- 89. Генеративные органы растений Цветок Морфологическая характеристика соцветий Классификация соцветий Опыление растений Строение семени Прорастание семян Строение
- 90. Цветок Морфологические типы цветков Строение околоцветника Андроцей Строение гинецея Семязачаток Двойное оплодотворение и его значение Типы
- 91. Морфологические типы цветков Цветок – это высокоспециализированный орган полового и бесполого размножения покрытосеменных растений. В цветках
- 92. Строение околоцветника Околоцветник бывает простым (гомохламидным) и двойным (гетерохламидным). В случае отсутствия околоцветника цветок называют ахламидным.
- 94. Андроцей Андроцей – это совокупность тычинок в цветке. Их количество варьирует от одного (орхидные) до нескольких
- 96. Строение гинецея Гинецей – это совокупность плодолистиков, образующих один или несколько пестиков. Плодолистик состоит из стерильной
- 98. Место прикрепления семязачатка к завязи называют плацентой, а расположение плацент в завязи – плацентацией. В зависимости
- 99. Семязачаток Семязачаток – многоклеточное образование семенных растений, из которого развивается семя. Семязачаток состоит из нуцеллуса (мегаспорангия),
- 101. Опыление растений Опылением называется перенос пыльцы на рыльце пестика. Различают два основных типа опыления: автогамию, или
- 102. Агентами перекрестного опыления чаще всего выступают насекомые и ветер. Опыление с помощью насекомых называется энтомофилией. Растения
- 103. Двойное оплодотворение и его значение Пыльцевое зерно, попав на рыльце пестика, начинает прорастать. При этом его
- 104. Типы отклонений от нормального оплодотворения у цветковых растений 1. Апомиксис – формирование зародыша из неоплодотворенных элементов
- 105. Формула и диаграмма цветка Для краткого условного выражения строения цветков применяют формулы. При их составлении учитывают
- 106. Морфологическая характеристика соцветий Соцветие – это система видоизмененных побегов, несущих цветки. Образование соцветий является высокой специализацией
- 107. Классификация соцветий В основу общей морфологической классификации соцветий положены два признака: способ ветвления осей и степень
- 109. К сложным моноподиальным соцветиям относятся такие: сложный колос, на главной оси которого сидят элементарные колоски (пшеница);
- 111. Цимозные соцветия характеризуются симподиальным ветвлением, относятся к сложным закрытым и представлены следующими типами: • монохазий (ось
- 113. Строение семени Семя развивается из семязачатка после оплодотворения (в случае апомиксиса – без оплодотворения). Снаружи оно
- 114. Семенная кожура (спермодерма) выполняет защитную функцию. На кожуре есть небольшое отверстие – микропиле, способствующее проникновению первых
- 115. Перисперм – запасающая ткань, характерная для некоторых растений (перец, кувшинка, звездчатка, свекла) и возникающая из нуцеллуса
- 116. В семени злаков эндосперм занимает значительный объем, т. к. в нем откладываются запасные вещества. Он дифференцирован
- 118. Прорастание семян Для прорастания семян необходимы определенные условия: • наличие воды (ткани семян обезвожены и содержат
- 119. По скорости прорастания и сохранению всхожести можно выделить следующие типы семян: • с длительным, глубоким покоем
- 121. Строение и классификация плодов Плод – характерный орган покрытосеменных растений. Его функция заключается в защите и
- 122. Околоплодник (перикарпий) представляет собой разросшуюся стенку завязи. Иногда в его образовании принимают участие и другие части
- 123. Строение гинецея. Существует классификация, основанная на взаимосвязи плодолистиков друг с другом и другими частями цветка. При
- 125. Распространение плодов и семян Единицами распространения могут служить разные части растений, называемые диаспорами. Карпологические диаспоры представляют
- 126. 3. Анемохория – распространение с помощью ветра. Анемохорные виды характеризуются высокой семенной продуктивностью. • Мелкие семена
- 127. 5. Зоохория – распространение с помощью животных (птиц, млекопитающих, насекомых и др. групп). Она осуществляется тремя
- 130. Скачать презентацию
Слайд 2Оглавление
Цитология растений
Ткани растений
Вегетативные органы растений
Генеративные органы растений
Оглавление
Цитология растений
Ткани растений
Вегетативные органы растений
Генеративные органы растений
Слайд 3I. Цитология растений
Общая характеристика растительной клетки
Цитоплазма
Пластиды
Строение и функции ядра
Вакуоль
Клеточная оболочка
Цитоплазматические включения
I. Цитология растений
Общая характеристика растительной клетки
Цитоплазма
Пластиды
Строение и функции ядра
Вакуоль
Клеточная оболочка
Цитоплазматические включения

Слайд 4Общая характеристика растительной клетки
Клетка представляет собой основную структурную и функциональную единицу всех
Общая характеристика растительной клетки
Клетка представляет собой основную структурную и функциональную единицу всех

Слайд 6Цитоплазма
Цитоплазма — основная часть протопласта клетки, в которой проходят все процессы клеточного
Цитоплазма
Цитоплазма — основная часть протопласта клетки, в которой проходят все процессы клеточного

Слайд 7Рибосомы
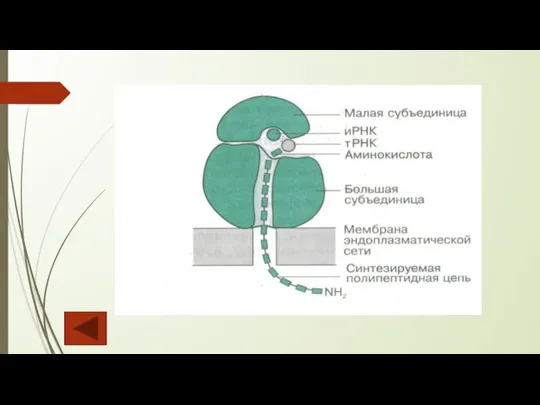
Рибосомы — универсальные органеллы, содержащиеся во всех клетках. Функция рибосом — биосинтез
Рибосомы
Рибосомы — универсальные органеллы, содержащиеся во всех клетках. Функция рибосом — биосинтез

Слайд 9Эндоплазматический ретикулум
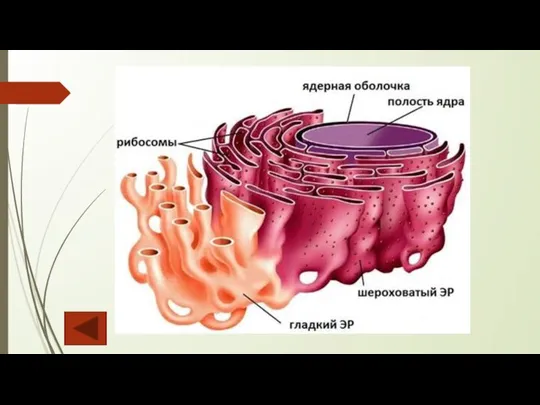
Эндоплазматический ретикулум (ЭР), или эндоплазматическая сеть, — непрерывно изменяющаяся трехмерная система
Эндоплазматический ретикулум
Эндоплазматический ретикулум (ЭР), или эндоплазматическая сеть, — непрерывно изменяющаяся трехмерная система

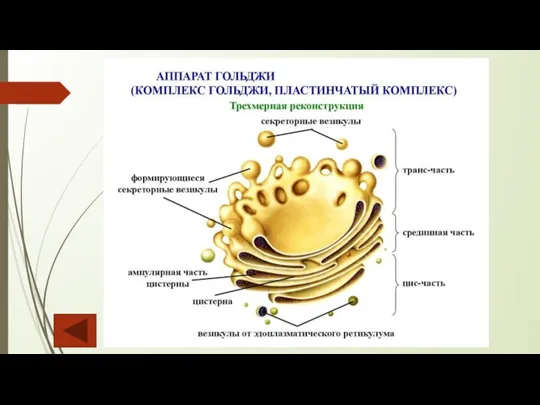
Слайд 11Аппарат Гольджи
Аппарат Гольджи состоит из отдельных диктиосом (телец Гольджи) и пузырьков Гольджи.
Аппарат Гольджи
Аппарат Гольджи состоит из отдельных диктиосом (телец Гольджи) и пузырьков Гольджи.

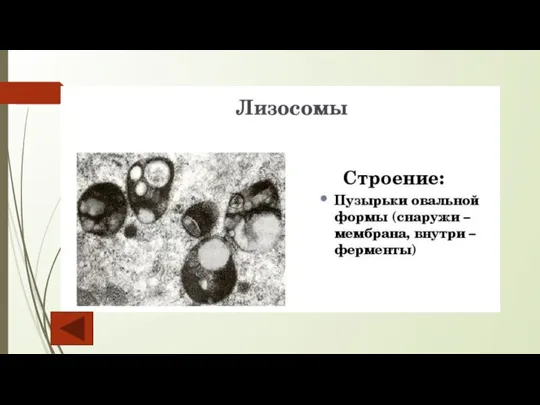
Слайд 13Лизосомы
Лизосомы — одномембранные органеллы округлой формы, в матриксе которых содержатся гидролитические ферменты
Лизосомы
Лизосомы — одномембранные органеллы округлой формы, в матриксе которых содержатся гидролитические ферменты

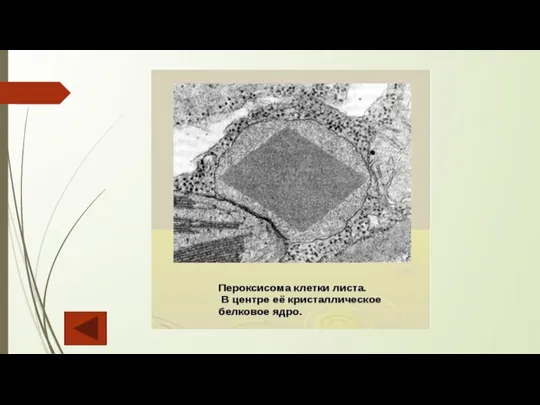
Слайд 15Пероксисомы
Пероксисомы (микротельца) встречаются в большинстве типов клеток растений и грибов. Они представляют
Пероксисомы
Пероксисомы (микротельца) встречаются в большинстве типов клеток растений и грибов. Они представляют

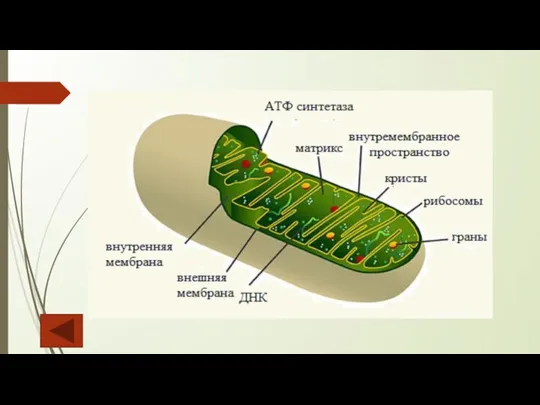
Слайд 17Митохондрии
Митохондрии — постоянно перемещающиеся органеллы округлой, цилиндрической или нитевидной формы. Относительно крупный
Митохондрии
Митохондрии — постоянно перемещающиеся органеллы округлой, цилиндрической или нитевидной формы. Относительно крупный

Слайд 19Пластиды
Пластиды встречаются только в растительных клетках. Выделяют три типа пластид (хлоро–, лейко–
Пластиды
Пластиды встречаются только в растительных клетках. Выделяют три типа пластид (хлоро–, лейко–

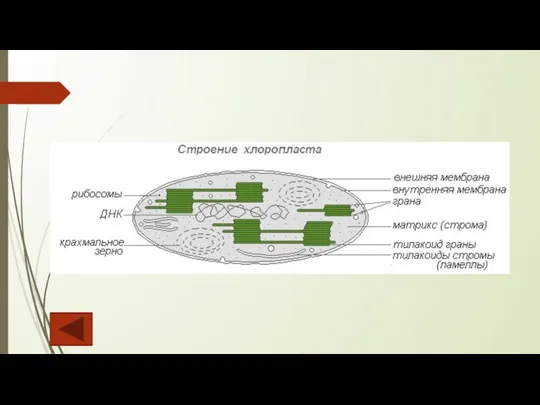
Слайд 21Лейкопласты – бесцветные мелкие пластиды, встречающиеся в запасающих органах растений (клубнях, корневищах,
Лейкопласты – бесцветные мелкие пластиды, встречающиеся в запасающих органах растений (клубнях, корневищах,

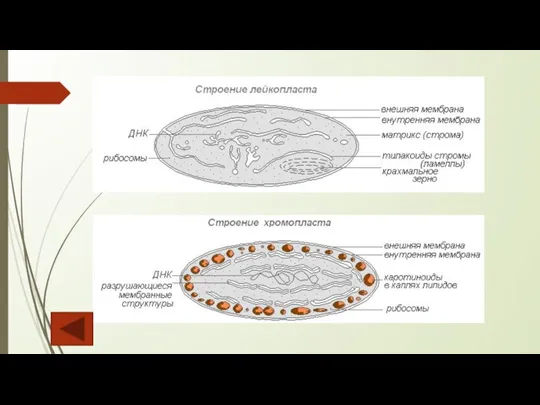
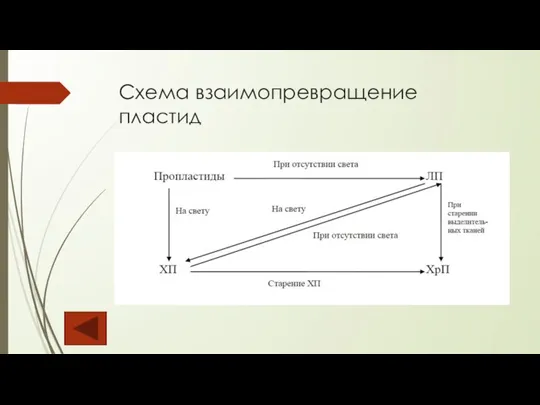
Слайд 23Схема взаимопревращение пластид
Схема взаимопревращение пластид
Слайд 24Строение и функции ядра
Ядро — центральная органелла клетки, регулирующая все процессы ее
Строение и функции ядра
Ядро — центральная органелла клетки, регулирующая все процессы ее

Слайд 25Ядерная оболочка и кариолимфа
Ядерная оболочка состоит из двух мембран, между которыми находится
Ядерная оболочка и кариолимфа
Ядерная оболочка состоит из двух мембран, между которыми находится

Слайд 26Ядрышко
Ядрышко — плотное сферическое тельце, обычно образующееся в области вторичной перетяжки спутничных
Ядрышко
Ядрышко — плотное сферическое тельце, обычно образующееся в области вторичной перетяжки спутничных

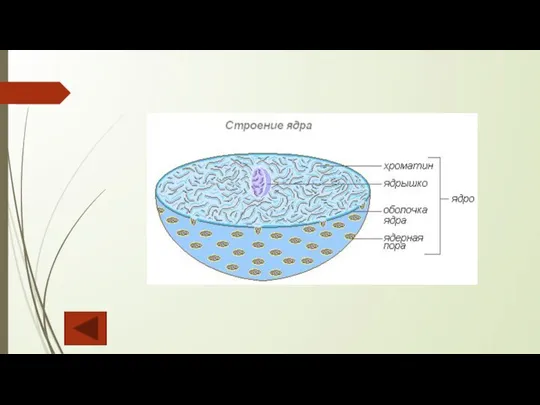
Слайд 27Хроматин
Важнейшая часть ядра — хроматин, который представляет собой молекулы ДНК, заключенные в
Хроматин
Важнейшая часть ядра — хроматин, который представляет собой молекулы ДНК, заключенные в

Слайд 29Вакуоль
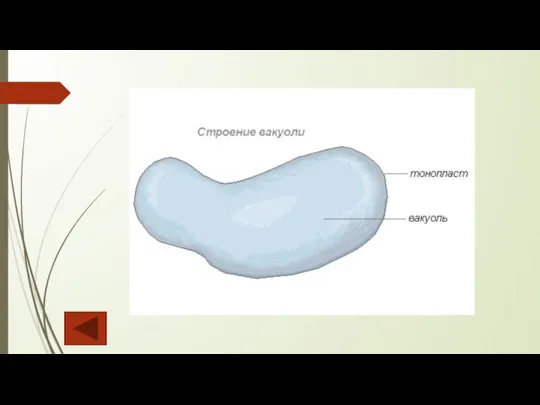
Вакуоли содержатся почти во всех растительных клетках. Они представляют собой полости, заполненные
Вакуоль
Вакуоли содержатся почти во всех растительных клетках. Они представляют собой полости, заполненные

Слайд 31Клеточная оболочка
Клеточная оболочка – структурное образование на периферии клетки, придающее ей прочность,
Клеточная оболочка
Клеточная оболочка – структурное образование на периферии клетки, придающее ей прочность,

Слайд 32Плазмодесмы
Плазмодесмы присущи только растительным клеткам. Они представляют собой тонкие цитоплазматические тяжи, соединяющие
Плазмодесмы
Плазмодесмы присущи только растительным клеткам. Они представляют собой тонкие цитоплазматические тяжи, соединяющие


Слайд 34Поры
Порами называют неутолщенные места оболочки (углубления), на которых отсутствует вторичная оболочка. Они
Поры
Порами называют неутолщенные места оболочки (углубления), на которых отсутствует вторичная оболочка. Они

Слайд 35Цитоплазматические включения
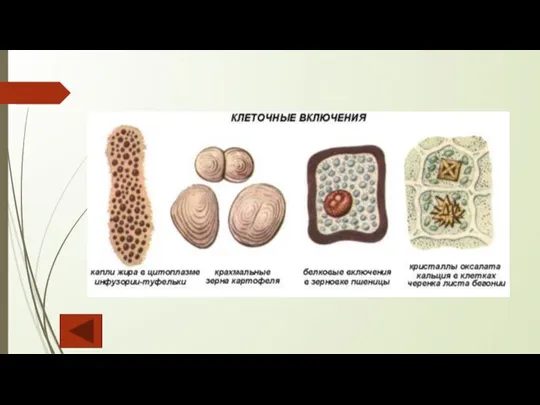
Включения – это локальная концентрация некоторых продуктов обмена в определенных местах
Цитоплазматические включения
Включения – это локальная концентрация некоторых продуктов обмена в определенных местах

Слайд 37Ткани растений
Понятие о ткани. Классификация растительных тканей
Образовательные ткани
Покровные ткани
Всасывающие ткани
Ассимиляционные ткани
Запасающие ткани
Аэренхима
Механические
Ткани растений
Понятие о ткани. Классификация растительных тканей
Образовательные ткани
Покровные ткани
Всасывающие ткани
Ассимиляционные ткани
Запасающие ткани
Аэренхима
Механические

Слайд 38Понятие о ткани. Классификация растительных тканей
Ткани – это устойчивые, т.е. закономерно повторяющиеся,
Понятие о ткани. Классификация растительных тканей
Ткани – это устойчивые, т.е. закономерно повторяющиеся,

Слайд 39Образовательные ткани
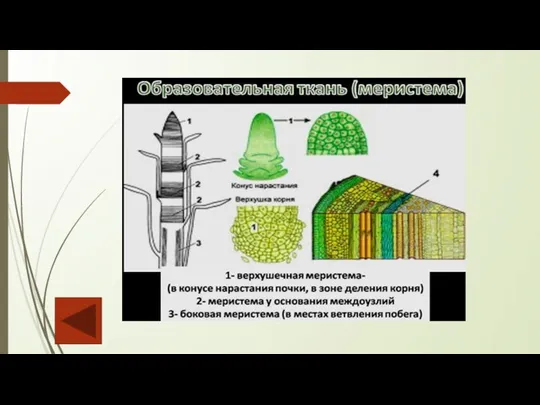
У взрослых растений образование новых клеток приурочено к определенным участкам –
Образовательные ткани
У взрослых растений образование новых клеток приурочено к определенным участкам –

Слайд 40Апикальные (верхушечные) располагаются на верхушке побегов и на кончике всех молодых корешков
Апикальные (верхушечные) располагаются на верхушке побегов и на кончике всех молодых корешков

Слайд 42Покровные ткани
Покровные ткани защищают внутренние ткани растений от прямого влияния факторов внешней
Покровные ткани
Покровные ткани защищают внутренние ткани растений от прямого влияния факторов внешней


Слайд 44Перидерма. Продолжительность жизни эпидермиса различна у разных видов и их органов. Например,
Перидерма. Продолжительность жизни эпидермиса различна у разных видов и их органов. Например,


Слайд 46Корка (ритидом) приходит на смену гладкой перидерме у некоторых древесных растений и
Корка (ритидом) приходит на смену гладкой перидерме у некоторых древесных растений и

Слайд 47Всасывающие ткани
Всасывающие ткани обеспечивают поступление в растение воды и растворенных в ней
Всасывающие ткани
Всасывающие ткани обеспечивают поступление в растение воды и растворенных в ней

Слайд 48Ассимиляционные ткани
Главная функция ассимиляционных тканей — фотосинтез. Поэтому их еще называют фотосинтезирующими
Ассимиляционные ткани
Главная функция ассимиляционных тканей — фотосинтез. Поэтому их еще называют фотосинтезирующими

Слайд 49Запасающие ткани
К накоплению запасных веществ способны все живые клетки растения. Когда же
Запасающие ткани
К накоплению запасных веществ способны все живые клетки растения. Когда же

Слайд 50Аэренхима
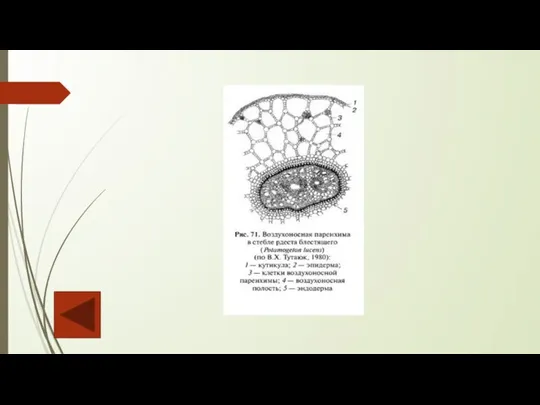
В растениях довольно часто встречается ткань с большим числом очень крупных межклетников
Аэренхима
В растениях довольно часто встречается ткань с большим числом очень крупных межклетников

Слайд 52Механические ткани
Механические ткани обусловливают прочность растения. Стенки клеток, слагающих эти ткани, утолщены.
Механические ткани
Механические ткани обусловливают прочность растения. Стенки клеток, слагающих эти ткани, утолщены.

Слайд 53Проводящие ткани
Проводящие ткани выполняют функцию транспортировки по растению питательных веществ. Они образуют
Проводящие ткани
Проводящие ткани выполняют функцию транспортировки по растению питательных веществ. Они образуют

Слайд 54Сосуды характерны для покрытосеменных растений. Членики сосудов располагаются один под другим, образуя
Сосуды характерны для покрытосеменных растений. Членики сосудов располагаются один под другим, образуя

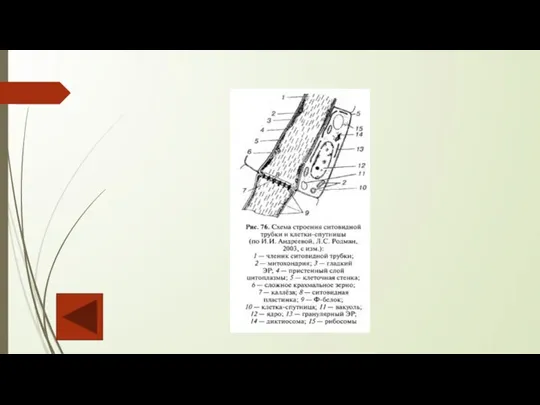
Слайд 55Флоэма – это ткань сосудистых растений, проводящая органические вещества. Первичная флоэма, которую
Флоэма – это ткань сосудистых растений, проводящая органические вещества. Первичная флоэма, которую

Слайд 57Выделительные ткани
Секреторными (выделительными) тканями принято называть структуры, выделяющие терпены, полисахариды, соли, воду
Выделительные ткани
Секреторными (выделительными) тканями принято называть структуры, выделяющие терпены, полисахариды, соли, воду

Слайд 58Перечислим внутренние секреторные ткани.
1. Секреторные идиобласты – выделительные клетки, рассеянные среди других
Перечислим внутренние секреторные ткани.
1. Секреторные идиобласты – выделительные клетки, рассеянные среди других


Слайд 60Вегетативные органы растений
Корень и корневая система
Побег и система побегов
Вегетативные органы растений
Корень и корневая система
Побег и система побегов

Слайд 61Корень и корневая система
Определение и функции корня
Зоны молодого корневого окончания
Корневая система
Корень и корневая система
Определение и функции корня
Зоны молодого корневого окончания
Корневая система

Слайд 62Определение и функции корня
Корень — основной вегетативный орган растения, выполняющий в типичном
Определение и функции корня
Корень — основной вегетативный орган растения, выполняющий в типичном

Слайд 63Зоны молодого корневого окончания
Нежные клетки апикальной меристемы, словно живым наперстком, всегда
Зоны молодого корневого окончания
Нежные клетки апикальной меристемы, словно живым наперстком, всегда

Слайд 64Зоны молодого корневого окончания
Вслед за этой зоной корень остается гладким, но становится
Зоны молодого корневого окончания
Вслед за этой зоной корень остается гладким, но становится


Слайд 66Корневая система
Корневая система – это совокупность всех корней одной особи. В состав
Корневая система
Корневая система – это совокупность всех корней одной особи. В состав


Слайд 68Побег и система побегов
Побег
Строение и деятельность апикальной меристемы побега
Морфологическое строение стебля
Первичное строение
Побег и система побегов
Побег
Строение и деятельность апикальной меристемы побега
Морфологическое строение стебля
Первичное строение

Слайд 69Побег
Побег – это основной орган семенных растений, выполняющий функцию воздушного питания. Состоит
Побег
Побег – это основной орган семенных растений, выполняющий функцию воздушного питания. Состоит

Слайд 70Строение и деятельность апикальной меристемы побега
Нарастание побега осуществляется за счет апикальных меристем.
Строение и деятельность апикальной меристемы побега
Нарастание побега осуществляется за счет апикальных меристем.

Слайд 71Морфологическое строение стебля
Стебель – это осевая часть побега растений, состоящая из узлов
Морфологическое строение стебля
Стебель – это осевая часть побега растений, состоящая из узлов

Слайд 72Первичное строение стебля
В стебле, имеющем первичное строение, различают эпидермис, первичную кору и
Первичное строение стебля
В стебле, имеющем первичное строение, различают эпидермис, первичную кору и

Слайд 73Стелярная теория
Стела – совокупность первичных проводящих пучков вместе с заключенными между ними
Стелярная теория
Стела – совокупность первичных проводящих пучков вместе с заключенными между ними


Слайд 75Стелярная теория
Дальнейшее эволюционное усложнение стелы связано с развитием крупных боковых органов (вай,
Стелярная теория
Дальнейшее эволюционное усложнение стелы связано с развитием крупных боковых органов (вай,

Слайд 76Типы вторичных утолщений стебля
Для большинства двудольных и голосеменных растений характерно вторичное утолщение
Типы вторичных утолщений стебля
Для большинства двудольных и голосеменных растений характерно вторичное утолщение

Слайд 77Лист и его функции
Лист – это один из основных органов высших растений,
Лист и его функции
Лист – это один из основных органов высших растений,

Слайд 78Морфологическое строение листа
Лист состоит из листовой пластинки и черешка. Место прикрепления листа
Морфологическое строение листа
Лист состоит из листовой пластинки и черешка. Место прикрепления листа

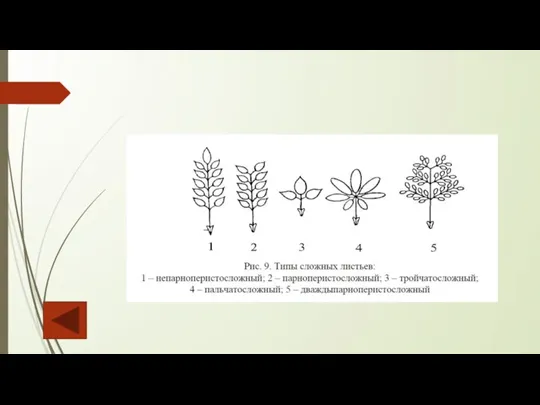
Слайд 79Различают простые и сложные листья. Простые состоят из одной пластинки. Сложные имеют
Различают простые и сложные листья. Простые состоят из одной пластинки. Сложные имеют

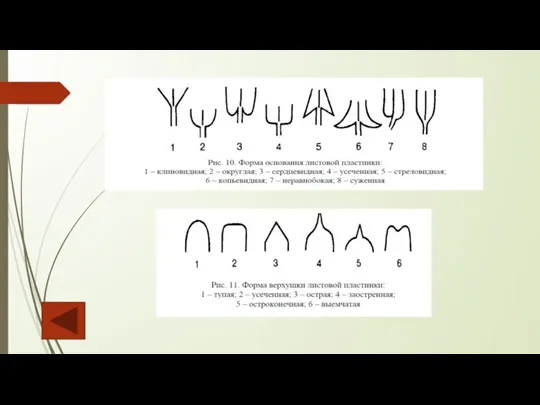
Слайд 81Форма, рассечение листовых пластинок простых листьев и листочков сложного листа могут быть
Форма, рассечение листовых пластинок простых листьев и листочков сложного листа могут быть

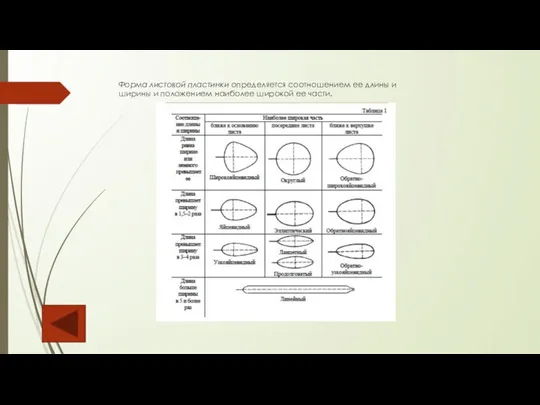
Слайд 83Форма листовой пластинки определяется соотношением ее длины и
ширины и положением наиболее широкой
Форма листовой пластинки определяется соотношением ее длины и ширины и положением наиболее широкой
Слайд 84Край листа имеет вырезки разной глубины. В тех случаях, если они не
заходят
Край листа имеет вырезки разной глубины. В тех случаях, если они не заходят
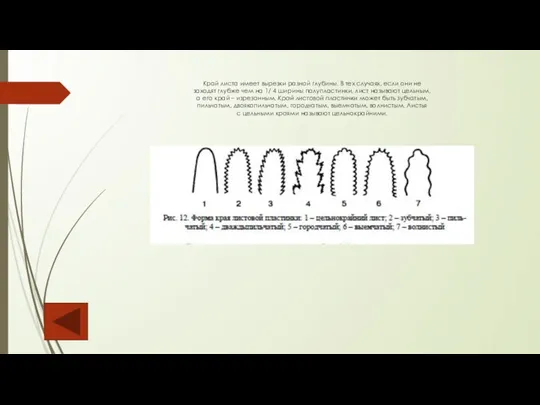
Слайд 85Листья, вырезы края которых глубже 1/4 ширины полупластинки, называют расчлененными.
Расчленение может быть
Листья, вырезы края которых глубже 1/4 ширины полупластинки, называют расчлененными. Расчленение может быть
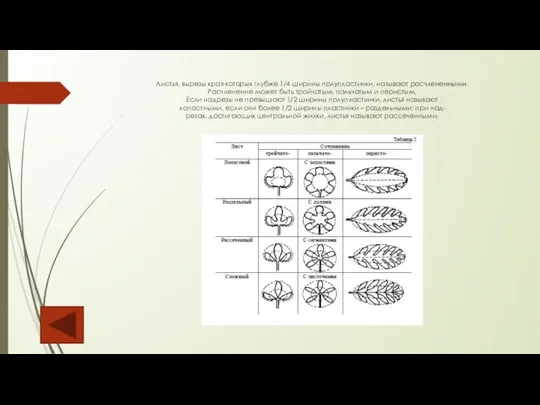
Слайд 86Одним из важных морфологических признаков листа является тип жилкования. Жилками условно называют
Одним из важных морфологических признаков листа является тип жилкования. Жилками условно называют

Слайд 87Анатомия листа
Анатомическое строение листа определяется его главной функцией – фотосинтезом. Поэтому основной
Анатомия листа
Анатомическое строение листа определяется его главной функцией – фотосинтезом. Поэтому основной

Слайд 88В листе проводящие ткани формируют закрытые коллатеральные пучки, причем ксилема всегда повернута
В листе проводящие ткани формируют закрытые коллатеральные пучки, причем ксилема всегда повернута

Слайд 89Генеративные органы растений
Цветок
Морфологическая характеристика соцветий
Классификация соцветий
Опыление растений
Строение семени
Прорастание семян
Строение и классификации плодов
Распространение
Генеративные органы растений
Цветок
Морфологическая характеристика соцветий
Классификация соцветий
Опыление растений
Строение семени
Прорастание семян
Строение и классификации плодов
Распространение

Слайд 90Цветок
Морфологические типы цветков
Строение околоцветника
Андроцей
Строение гинецея
Семязачаток
Двойное оплодотворение и его значение
Типы отклонений от нормального
Цветок
Морфологические типы цветков
Строение околоцветника
Андроцей
Строение гинецея
Семязачаток
Двойное оплодотворение и его значение
Типы отклонений от нормального

Слайд 91Морфологические типы цветков
Цветок – это высокоспециализированный орган полового и бесполого размножения покрытосеменных
Морфологические типы цветков
Цветок – это высокоспециализированный орган полового и бесполого размножения покрытосеменных

Слайд 92Строение околоцветника
Околоцветник бывает простым (гомохламидным) и двойным (гетерохламидным). В случае отсутствия околоцветника
Строение околоцветника
Околоцветник бывает простым (гомохламидным) и двойным (гетерохламидным). В случае отсутствия околоцветника

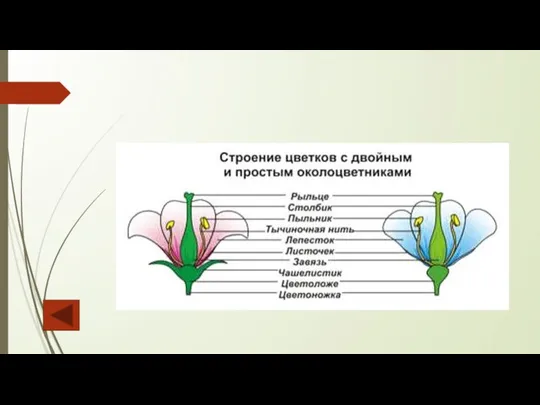
Слайд 94Андроцей
Андроцей – это совокупность тычинок в цветке. Их количество варьирует от одного
Андроцей
Андроцей – это совокупность тычинок в цветке. Их количество варьирует от одного

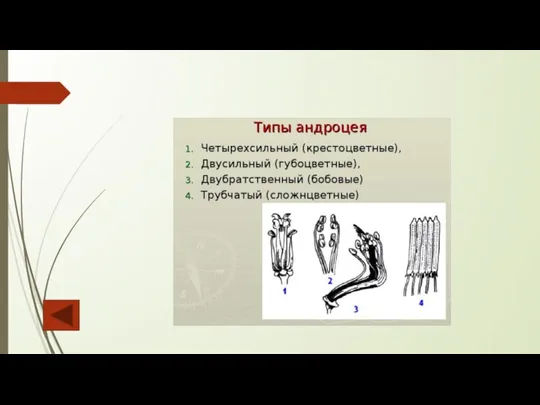
Слайд 96Строение гинецея
Гинецей – это совокупность плодолистиков, образующих один или несколько пестиков.
Плодолистик состоит
Строение гинецея
Гинецей – это совокупность плодолистиков, образующих один или несколько пестиков.
Плодолистик состоит


Слайд 98Место прикрепления семязачатка к завязи называют плацентой, а расположение плацент в завязи
Место прикрепления семязачатка к завязи называют плацентой, а расположение плацент в завязи

Слайд 99Семязачаток
Семязачаток – многоклеточное образование семенных растений, из которого развивается семя. Семязачаток состоит
Семязачаток
Семязачаток – многоклеточное образование семенных растений, из которого развивается семя. Семязачаток состоит


Слайд 101 Опыление растений
Опылением называется перенос пыльцы на рыльце пестика.
Различают два основных типа
Опыление растений
Опылением называется перенос пыльцы на рыльце пестика.
Различают два основных типа

Слайд 102Агентами перекрестного опыления чаще всего выступают насекомые и ветер. Опыление с помощью
Агентами перекрестного опыления чаще всего выступают насекомые и ветер. Опыление с помощью

Слайд 103Двойное оплодотворение и его значение
Пыльцевое зерно, попав на рыльце пестика, начинает прорастать.
Двойное оплодотворение и его значение
Пыльцевое зерно, попав на рыльце пестика, начинает прорастать.

Слайд 104Типы отклонений от нормального оплодотворения у цветковых растений
1. Апомиксис – формирование зародыша
Типы отклонений от нормального оплодотворения у цветковых растений
1. Апомиксис – формирование зародыша

Слайд 105Формула и диаграмма цветка
Для краткого условного выражения строения цветков применяют формулы. При
Формула и диаграмма цветка
Для краткого условного выражения строения цветков применяют формулы. При

Слайд 106Морфологическая характеристика соцветий
Соцветие – это система видоизмененных побегов, несущих цветки. Образование соцветий
Морфологическая характеристика соцветий
Соцветие – это система видоизмененных побегов, несущих цветки. Образование соцветий

Слайд 107Классификация соцветий
В основу общей морфологической классификации соцветий положены два признака: способ ветвления
Классификация соцветий
В основу общей морфологической классификации соцветий положены два признака: способ ветвления

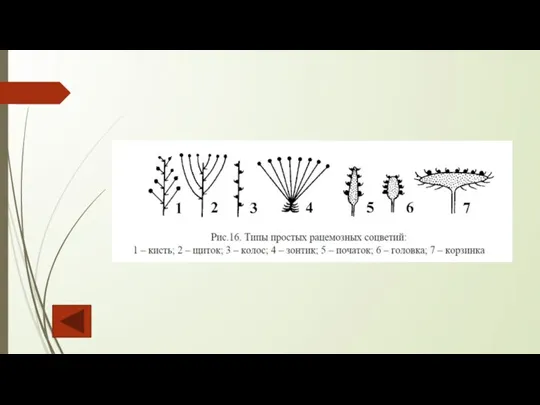
Слайд 109 К сложным моноподиальным соцветиям относятся такие:
сложный колос, на главной оси которого
К сложным моноподиальным соцветиям относятся такие:
сложный колос, на главной оси которого

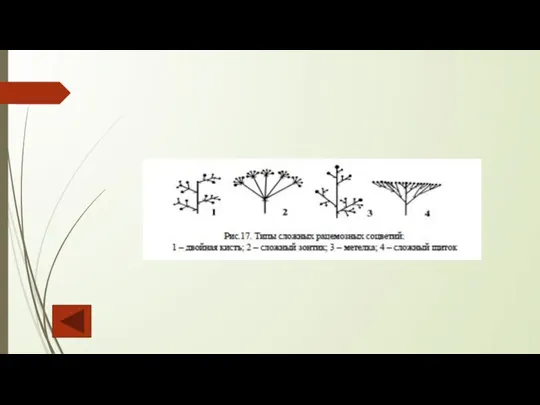
Слайд 111Цимозные соцветия характеризуются симподиальным ветвлением, относятся к сложным закрытым и представлены следующими
Цимозные соцветия характеризуются симподиальным ветвлением, относятся к сложным закрытым и представлены следующими


Слайд 113 Строение семени
Семя развивается из семязачатка после оплодотворения (в случае апомиксиса –
Строение семени
Семя развивается из семязачатка после оплодотворения (в случае апомиксиса –

Слайд 114Семенная кожура (спермодерма) выполняет защитную функцию. На кожуре есть небольшое отверстие –
Семенная кожура (спермодерма) выполняет защитную функцию. На кожуре есть небольшое отверстие –

Слайд 115Перисперм – запасающая ткань, характерная для некоторых растений (перец, кувшинка, звездчатка, свекла)
Перисперм – запасающая ткань, характерная для некоторых растений (перец, кувшинка, звездчатка, свекла)

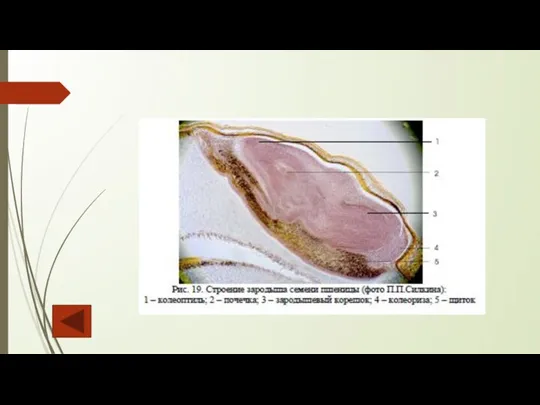
Слайд 116В семени злаков эндосперм занимает значительный объем, т. к. в нем откладываются
В семени злаков эндосперм занимает значительный объем, т. к. в нем откладываются

Слайд 118Прорастание семян
Для прорастания семян необходимы определенные условия:
• наличие воды (ткани семян обезвожены
Прорастание семян
Для прорастания семян необходимы определенные условия:
• наличие воды (ткани семян обезвожены

Слайд 119По скорости прорастания и сохранению всхожести можно выделить следующие типы семян:
• с
По скорости прорастания и сохранению всхожести можно выделить следующие типы семян:
• с


Слайд 121Строение и классификация плодов
Плод – характерный орган покрытосеменных растений. Его функция заключается
Строение и классификация плодов
Плод – характерный орган покрытосеменных растений. Его функция заключается

Слайд 122Околоплодник (перикарпий) представляет собой разросшуюся стенку завязи. Иногда в его образовании принимают
Околоплодник (перикарпий) представляет собой разросшуюся стенку завязи. Иногда в его образовании принимают

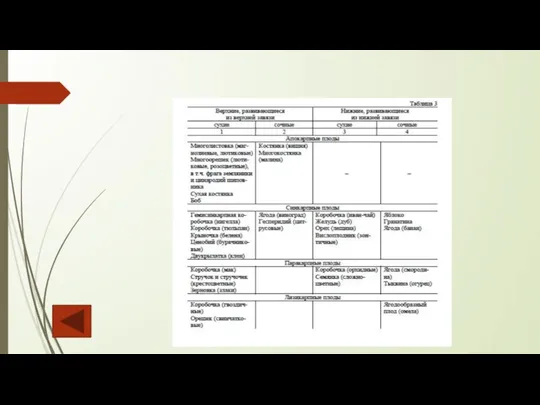
Слайд 123Строение гинецея. Существует классификация, основанная на взаимосвязи плодолистиков друг с другом и
Строение гинецея. Существует классификация, основанная на взаимосвязи плодолистиков друг с другом и

Слайд 125Распространение плодов и семян
Единицами распространения могут служить разные части растений, называемые диаспорами.
Распространение плодов и семян
Единицами распространения могут служить разные части растений, называемые диаспорами.

Слайд 1263. Анемохория – распространение с помощью ветра. Анемохорные виды характеризуются высокой семенной
3. Анемохория – распространение с помощью ветра. Анемохорные виды характеризуются высокой семенной

Слайд 1275. Зоохория – распространение с помощью животных (птиц, млекопитающих, насекомых и др.
5. Зоохория – распространение с помощью животных (птиц, млекопитающих, насекомых и др.


 Автор проекта: Гафурова Алия Раильевна Воспитатель МДОУ №3 «Звездочка» г.Ершова. Саратовской области 2010 год.
Автор проекта: Гафурова Алия Раильевна Воспитатель МДОУ №3 «Звездочка» г.Ершова. Саратовской области 2010 год. Цветы первоцветы
Цветы первоцветы Теории происхождения человека
Теории происхождения человека Китообразные (происхождение)
Китообразные (происхождение) Мейоз
Мейоз Характеристика грибов. Питание грибов
Характеристика грибов. Питание грибов Синтез белка
Синтез белка Скаты
Скаты Тип споровики
Тип споровики Цветковые растения. Однолетники и многолетники
Цветковые растения. Однолетники и многолетники Автоволновые процессы в активновозбудимых средах
Автоволновые процессы в активновозбудимых средах Эволюция. Антропогенез
Эволюция. Антропогенез Кафедра биологии и методики ее преподавания
Кафедра биологии и методики ее преподавания Белки - состав, строение, свойства
Белки - состав, строение, свойства Натрий в организме собаки
Натрий в организме собаки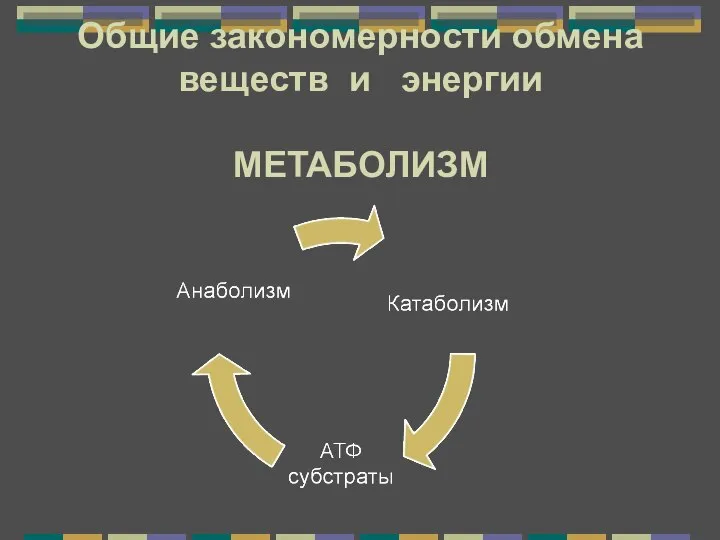 Метаболизм. Энергетический обмен
Метаболизм. Энергетический обмен Животные левши или правши?
Животные левши или правши? Моя Отчизна - Русь. Владимир Иванович Даль
Моя Отчизна - Русь. Владимир Иванович Даль Пищеварительная система. Часть 2
Пищеварительная система. Часть 2 Презентация на тему "Виды и формы иммунитета" - презентации по Биологии
Презентация на тему "Виды и формы иммунитета" - презентации по Биологии Пищеварительная система человека
Пищеварительная система человека Единый государственный экзамен
Единый государственный экзамен Презентация на тему Осторожно: клещи!
Презентация на тему Осторожно: клещи!  XI Пара ЧМН (n. accessorius)
XI Пара ЧМН (n. accessorius) Salna un sarma
Salna un sarma Кожно-мышечная чувствительность. Обоняние. Вкус
Кожно-мышечная чувствительность. Обоняние. Вкус Биогеноценоз и экосистемы - сходства и различия
Биогеноценоз и экосистемы - сходства и различия Презентация на тему БАБОЧКИ
Презентация на тему БАБОЧКИ