Физика и структурная биология в начале XXI века И. Н. Сердюк Лаборатория физики нуклеопротеидов, Институт Белка РАН, г. Пущино. и Лаб
- Физика и структурная биология в начале XXI века И. Н. Сердюк Лаборатория физики нуклеопротеидов, Институт Белка РАН, г. Пущино. и Лаб

Содержание
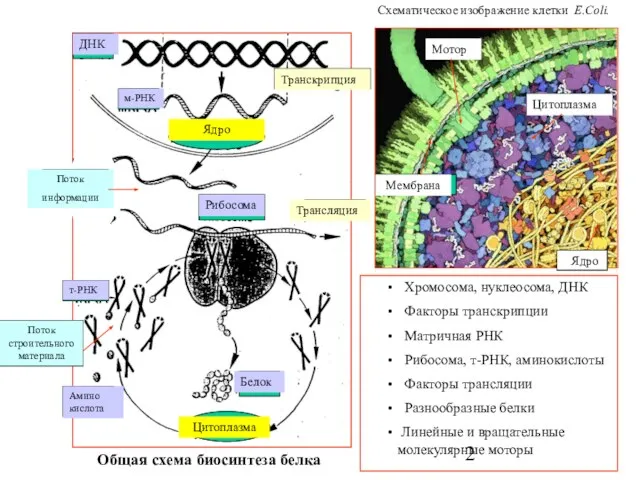
- 2. Транскрипция Поток информации ДНК м-РНК Рибосома Общая схема биосинтеза белка Поток строительного материала Трансляция Цитоплазма Ядро
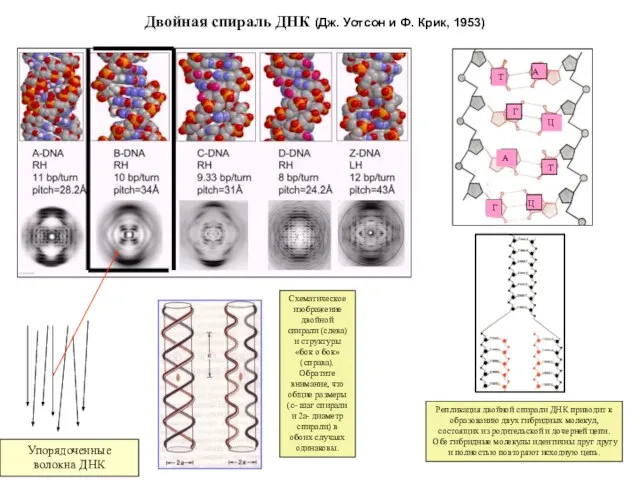
- 3. Двойная спираль ДНК (Дж. Уотсон и Ф. Крик, 1953) Репликация двойной спирали ДНК приводит к образованию
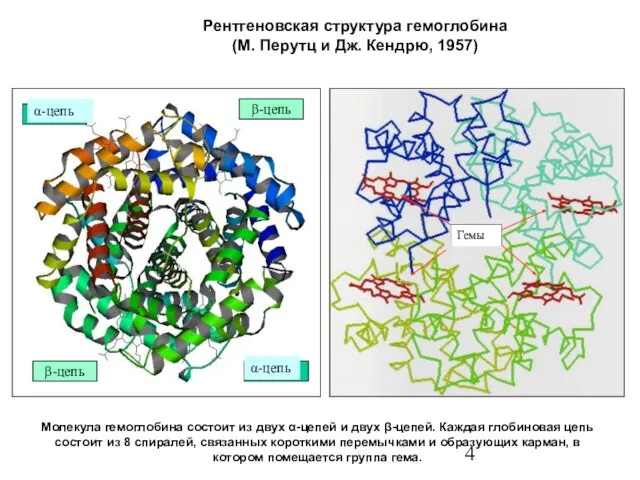
- 4. Рентгеновская структура гемоглобина (М. Перутц и Дж. Кендрю, 1957) Молекула гемоглобина состоит из двух α-цепей и
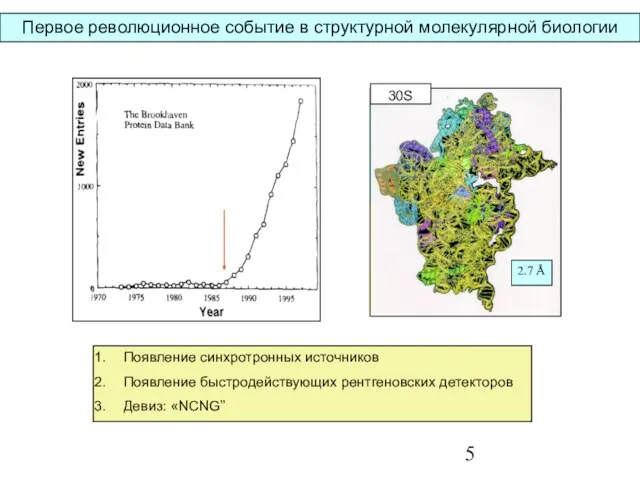
- 5. Первое революционное событие в структурной молекулярной биологии Появление синхротронных источников Появление быстродействующих рентгеновских детекторов Девиз: «NCNG”
- 6. Ондулятор Отклоняющий магнит Синхротронные источники третьего поколения Синхротрон Большое число отклоняющих магнитов располагаются по окружности синхротронного
- 7. Нейтронные источники Стационарные (с тратой энергии) Импульсные (с накачкой энергии) Ускоритель электронов + мишень из тяжелого
- 8. Трехмерная структура молекулы лизоцима и 157 молекул связанной с ним воды с разрешением 2 Å. Такое
- 9. Импульсная Фурье-ЯМР спектроскопия, многомерный и гетероядерный ЯМР, изощренные схемы селективного мечения белков и нуклеиновых кислот 2H,
- 10. Методы рентгеновской кристаллографии и ЯМР обладают двумя принципиальными недостатками: 1) работают при очень высокие концентрациях вещества
- 11. Исследование структуры и функционирования одиночных биологических макромолекул Детектирование одиночных флюоресцентно меченых молекул с помощью лазерной флюоресценции
- 12. Лазерная наведенная флюоресценция , В лазерной наведенной флюоресценции одиночная молекула периодически возбуждается с тем чтобы дать
- 13. Флюоресцентные метки, используемые в структурной биологии Внутренние метки: триптофан и тирозин в белках, а также минорные
- 14. Флюоресцентный перенос энергии. Флюоресцентная линейка В флюоресцентном переносе энергии (FRET), флюорофор, называемый донором (D), переносит часть
- 15. GFP как клеточный репортер Двухцветная флюоресцентная фотография клетки выявляющая распределение двух белков (Yellow-FP и Cyan-FP) в
- 16. Визуализация одиночных молекул требует радикального изменения наблюдаемого обьема вплоть до нескольких аттолитров Двухфотонное возбуждение в лазерной
- 17. Двухфотонное возбуждение In 1931 M. Goppert-Mayer gave theoretical background of two-photon excitation in fluorescence. Предельно малый
- 18. Микроскопия ближнего поля Near-field scanning optical microscopy (NSOM). “Безлинзовая микроскопия” Техника изображения основанная на принципе «оптического
- 19. Флюоресцентная микроскопия при полном внутреннем отражении Характерная глубина проникновения света около 3000Å Характерный обьем возбуждения ~
- 20. Конфокальная микроскопия Формирование линзой изображения двух точек тонкого образца в отсутствии диафрагмы. Демонстрация получения конфокального изображения.
- 21. The main characteristic of five methods of single molecules optical detection.
- 22. П Е Р Е Р Ы В? Визуализация и манипулирование одиночными макромолекулами
- 23. Сканирующая туннельная микроскопия (G. Binning and H. Rohrer, 1980) Iodine atoms on the surface of platinum
- 24. Микроскопия силового поля (безлинзовая микроскопия) Микроскопия силового поля (по английски AFM), не содержит линзы как таковой
- 25. Изображение биологических макромолекул методом AFM Слева: изображение иммуноглобулина G методом AFM. Справа: кристаллическая структура Три последовательных
- 26. Изображение биологических структур методом AFM ДНК Фальш Кольцевая ДНК в B-форме. Диаметр спирали 24 Å
- 27. Наблюдение за ростом двумерных кристаллов методом AFM Рост кристаллов белка аннексина V на подложке из планарных
- 28. Манипуляция одиночными макромолекулами Бусинка с высоким показателем преломления преломления располагается в центре интенсивного лазерного пучка. Типичное
- 29. В оптическом твизере сильно сфокусированный лазерный луч является оптической ловушкой, которая втягивает в фокус маленькие коллоидальные
- 30. Метод AFM методе в режиме измерения силы. Типичный эксперимент по растягиванию одиночной молекулы в методе AFM
- 31. Белковая архитектура и механическая стабильность Красные линии указывают топологию критических H-связей, разрывающихся при приложении силы. Human
- 32. Природные конфигурации ДНК «Искусственные» конфигурации ДНК (Cлева S-DNA), полученная растяжением B-DNA с помощью оптического твизера (диаметр
- 33. Схематические диаграммы использования оптических твизеров при исследовании линейных молекулярных моторов
- 34. РНК-полимераза- вдоль ДНК Кинезин- вдоль микротрубочек Миозин - вдоль актиновых нитей Все линейные моторы движутся вдоль
- 35. Сравнение основных механических характеристик молекулярных моторов. Размер шага каждого линейного мотора зависит типа платформы: Реальная величина
- 36. F0F1 ATP фазный мотор двойного действия Молекулярная архитектура Na-насоса F0 мотора, генерирующего АТФ. Экспериментальная схема прямого
- 37. Молекулярная архитектура бактериальной флагеллы как вращающегося устройства Бактериальная флагелла состоит из мотора, располагающегося внутри клетки, втулки
- 38. I. Serdyuk N. Zaccai J. Zaccai “Physical methods in structural biology” Cambridge, 2005 Introduction. From thermodynamic
- 39. Курс лекций проф. И.Н. Сердюк “Физические методы в структурной биологии” Введение. От термодинамики к одиночным молекулам
- 41. Скачать презентацию
Слайд 2Транскрипция
Поток
информации
ДНК
м-РНК
Рибосома
Общая схема биосинтеза белка
Поток строительного материала
Трансляция
Цитоплазма
Ядро
т-РНК
Белок
Хромосома, нуклеосома, ДНК
Факторы
Транскрипция
Поток
информации
ДНК
м-РНК
Рибосома
Общая схема биосинтеза белка
Поток строительного материала
Трансляция
Цитоплазма
Ядро
т-РНК
Белок
Хромосома, нуклеосома, ДНК
Факторы
Слайд 3Двойная спираль ДНК (Дж. Уотсон и Ф. Крик, 1953)
Репликация двойной спирали ДНК
Двойная спираль ДНК (Дж. Уотсон и Ф. Крик, 1953)
Репликация двойной спирали ДНК
Слайд 4Рентгеновская структура гемоглобина
(М. Перутц и Дж. Кендрю, 1957)
Молекула гемоглобина состоит из двух
Рентгеновская структура гемоглобина
(М. Перутц и Дж. Кендрю, 1957)
Молекула гемоглобина состоит из двух
Слайд 5Первое революционное событие в структурной молекулярной биологии
Появление синхротронных источников
Появление быстродействующих рентгеновских детекторов
Девиз:
Первое революционное событие в структурной молекулярной биологии
Появление синхротронных источников
Появление быстродействующих рентгеновских детекторов
Девиз:
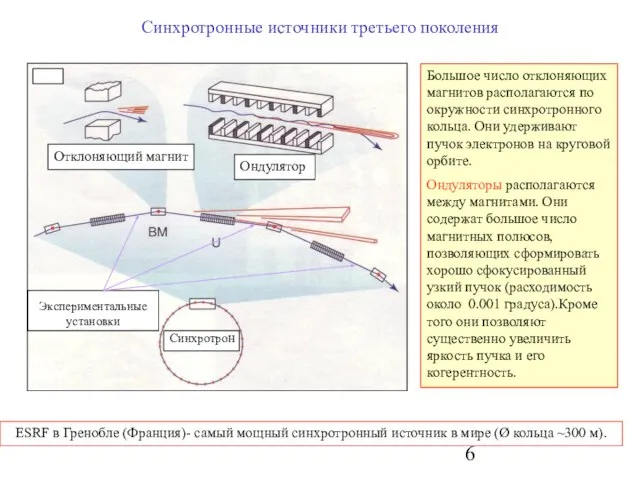
Слайд 6Ондулятор
Отклоняющий магнит
Синхротронные источники третьего поколения
Синхротрон
Большое число отклоняющих магнитов располагаются по окружности синхротронного
Ондулятор
Отклоняющий магнит
Синхротронные источники третьего поколения
Синхротрон
Большое число отклоняющих магнитов располагаются по окружности синхротронного
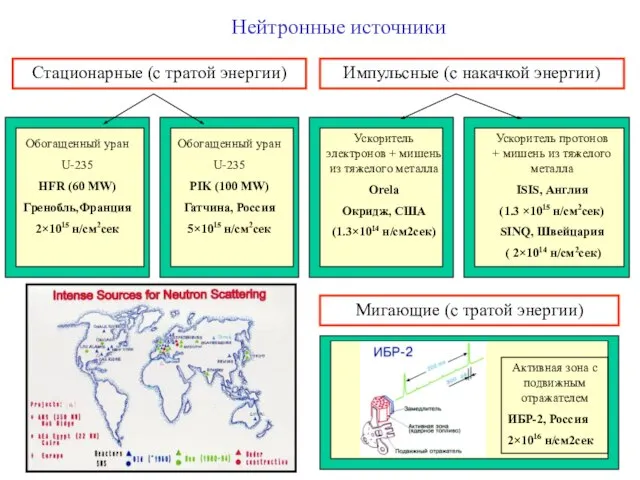
Слайд 7Нейтронные источники
Стационарные (с тратой энергии)
Импульсные (с накачкой энергии)
Ускоритель электронов + мишень из
Нейтронные источники
Стационарные (с тратой энергии)
Импульсные (с накачкой энергии)
Ускоритель электронов + мишень из
Слайд 8Трехмерная структура молекулы лизоцима и 157 молекул связанной с ним воды с
Трехмерная структура молекулы лизоцима и 157 молекул связанной с ним воды с

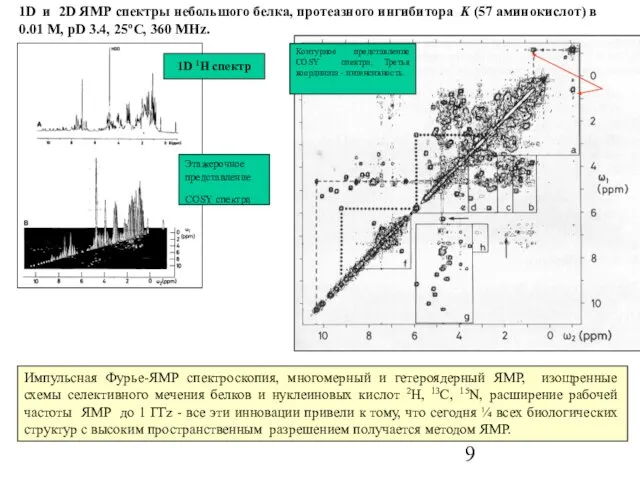
Слайд 9Импульсная Фурье-ЯМР спектроскопия, многомерный и гетероядерный ЯМР, изощренные схемы селективного мечения белков
Импульсная Фурье-ЯМР спектроскопия, многомерный и гетероядерный ЯМР, изощренные схемы селективного мечения белков
Слайд 10 Методы рентгеновской кристаллографии и ЯМР обладают двумя принципиальными недостатками:
1) работают
Методы рентгеновской кристаллографии и ЯМР обладают двумя принципиальными недостатками:
1) работают

Слайд 11
Исследование структуры и функционирования одиночных биологических макромолекул
Детектирование одиночных флюоресцентно меченых молекул
Исследование структуры и функционирования одиночных биологических макромолекул
Детектирование одиночных флюоресцентно меченых молекул

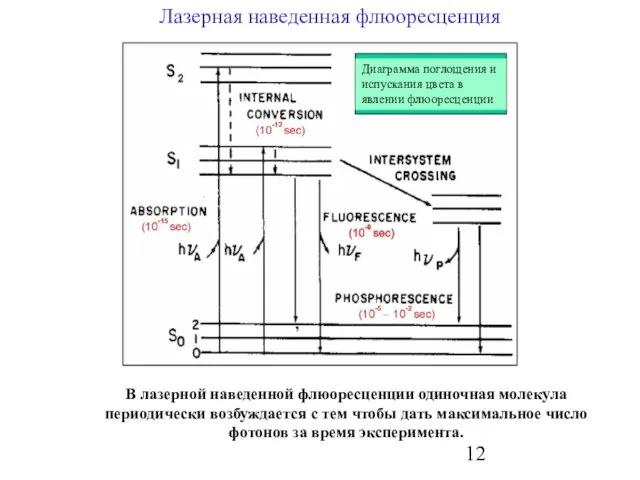
Слайд 12Лазерная наведенная флюоресценция
,
В лазерной наведенной флюоресценции одиночная молекула периодически возбуждается с тем
Лазерная наведенная флюоресценция
,
В лазерной наведенной флюоресценции одиночная молекула периодически возбуждается с тем
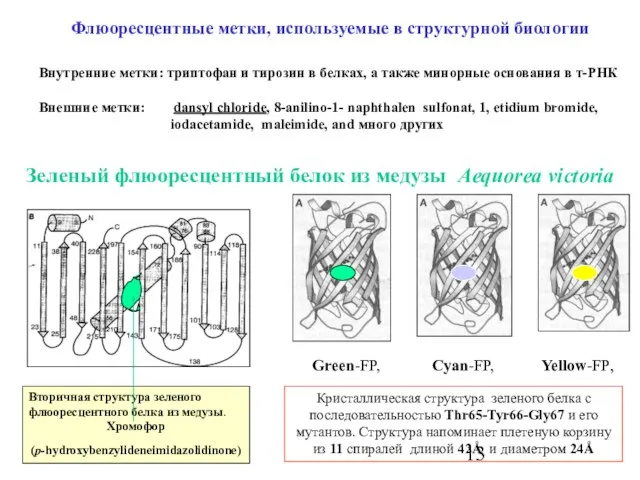
Слайд 13Флюоресцентные метки, используемые в структурной биологии
Внутренние метки: триптофан и тирозин в белках,
Флюоресцентные метки, используемые в структурной биологии
Внутренние метки: триптофан и тирозин в белках,
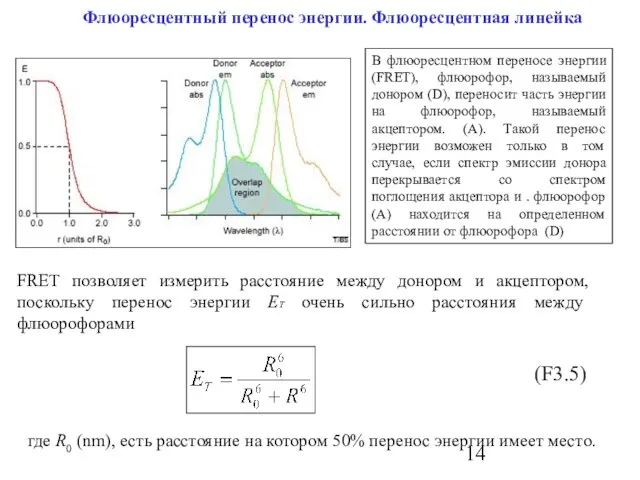
Слайд 14 Флюоресцентный перенос энергии. Флюоресцентная линейка
В флюоресцентном переносе энергии (FRET), флюорофор, называемый
Флюоресцентный перенос энергии. Флюоресцентная линейка
В флюоресцентном переносе энергии (FRET), флюорофор, называемый
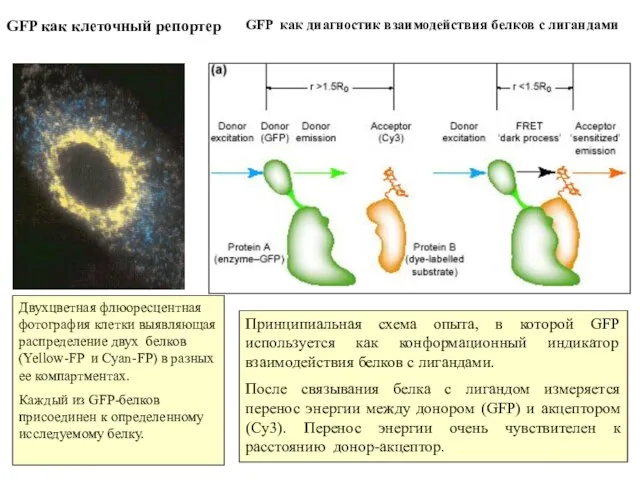
Слайд 15GFP как клеточный репортер
Двухцветная флюоресцентная фотография клетки выявляющая распределение двух белков
GFP как клеточный репортер
Двухцветная флюоресцентная фотография клетки выявляющая распределение двух белков
Слайд 16Визуализация одиночных молекул требует радикального изменения наблюдаемого обьема вплоть до нескольких аттолитров
Двухфотонное
Визуализация одиночных молекул требует радикального изменения наблюдаемого обьема вплоть до нескольких аттолитров
Двухфотонное

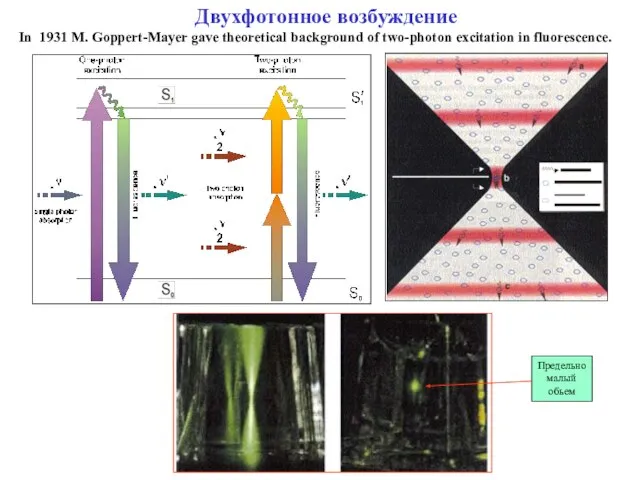
Слайд 17Двухфотонное возбуждение
In 1931 M. Goppert-Mayer gave theoretical background of two-photon excitation in
Двухфотонное возбуждение
In 1931 M. Goppert-Mayer gave theoretical background of two-photon excitation in
Слайд 18Микроскопия ближнего поля
Near-field scanning optical microscopy (NSOM).
“Безлинзовая микроскопия”
Техника изображения основанная
Микроскопия ближнего поля
Near-field scanning optical microscopy (NSOM).
“Безлинзовая микроскопия”
Техника изображения основанная

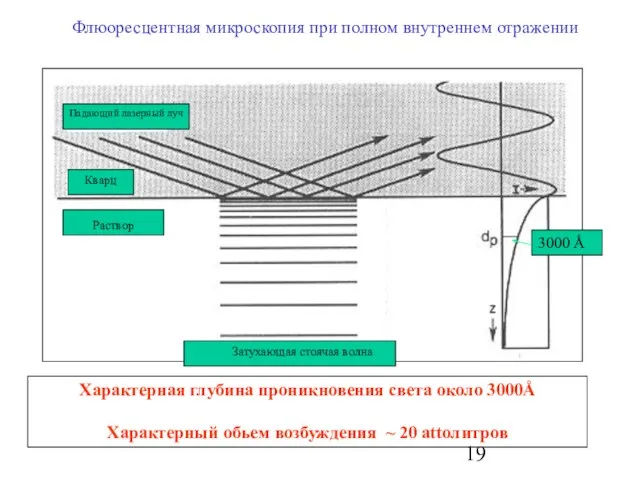
Слайд 19Флюоресцентная микроскопия при полном внутреннем отражении
Характерная глубина проникновения света около 3000Å
Характерный обьем
Флюоресцентная микроскопия при полном внутреннем отражении
Характерная глубина проникновения света около 3000Å
Характерный обьем
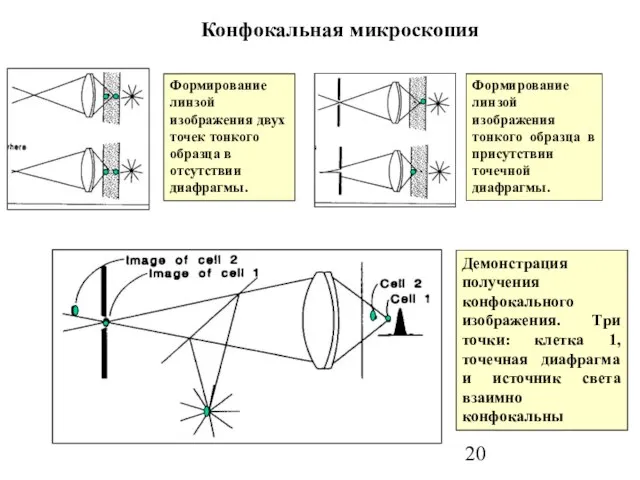
Слайд 20Конфокальная микроскопия
Формирование линзой изображения двух точек тонкого образца в отсутствии диафрагмы.
Конфокальная микроскопия
Формирование линзой изображения двух точек тонкого образца в отсутствии диафрагмы.
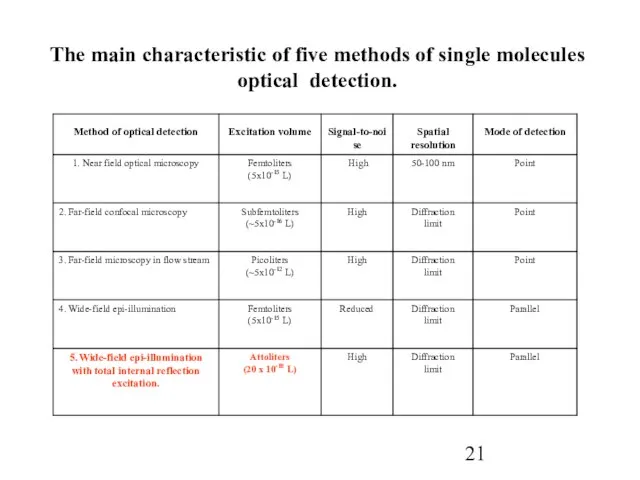
Слайд 21The main characteristic of five methods of single molecules optical detection.
The main characteristic of five methods of single molecules optical detection.
Слайд 22П Е Р Е Р Ы В?
Визуализация и манипулирование одиночными макромолекулами
П Е Р Е Р Ы В?
Визуализация и манипулирование одиночными макромолекулами

Слайд 23Сканирующая туннельная микроскопия
(G. Binning and H. Rohrer, 1980)
Iodine atoms on the
Сканирующая туннельная микроскопия
(G. Binning and H. Rohrer, 1980)
Iodine atoms on the

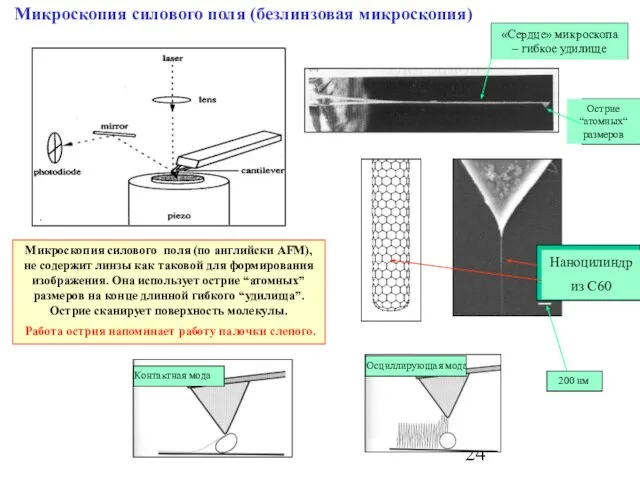
Слайд 24Микроскопия силового поля (безлинзовая микроскопия)
Микроскопия силового поля (по английски AFM), не содержит
Микроскопия силового поля (безлинзовая микроскопия)
Микроскопия силового поля (по английски AFM), не содержит
Слайд 25Изображение биологических макромолекул методом AFM
Слева: изображение иммуноглобулина G методом AFM. Справа: кристаллическая
Изображение биологических макромолекул методом AFM
Слева: изображение иммуноглобулина G методом AFM. Справа: кристаллическая

Слайд 26Изображение биологических структур методом AFM
ДНК
Фальш
Кольцевая ДНК в B-форме. Диаметр спирали
Изображение биологических структур методом AFM
ДНК
Фальш
Кольцевая ДНК в B-форме. Диаметр спирали

Слайд 27Наблюдение за ростом двумерных кристаллов методом AFM
Рост кристаллов белка аннексина V на
Наблюдение за ростом двумерных кристаллов методом AFM
Рост кристаллов белка аннексина V на

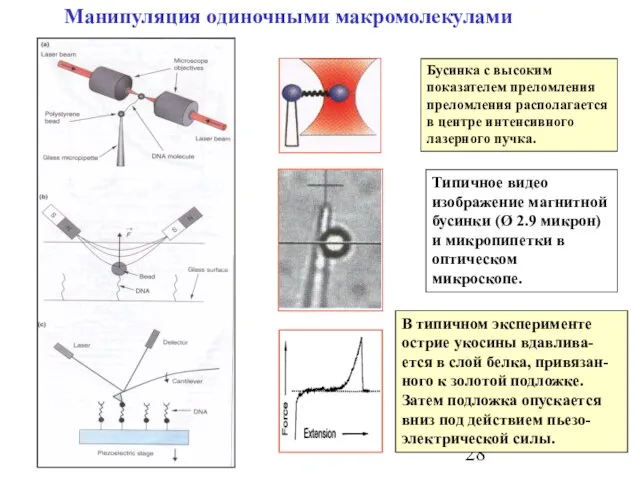
Слайд 28Манипуляция одиночными макромолекулами
Бусинка с высоким показателем преломления преломления располагается в центре интенсивного
Манипуляция одиночными макромолекулами
Бусинка с высоким показателем преломления преломления располагается в центре интенсивного
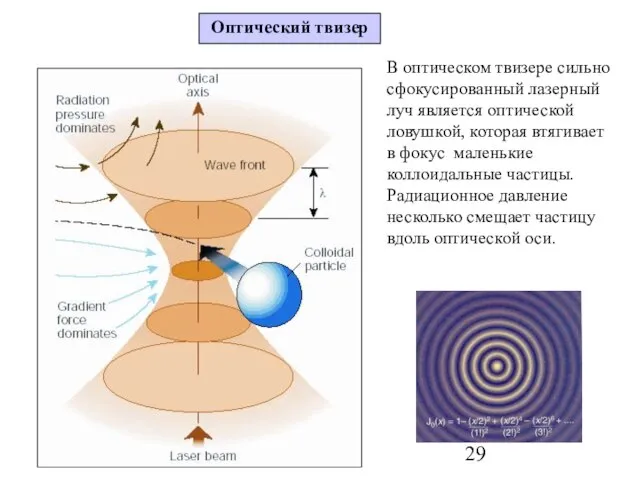
Слайд 29В оптическом твизере сильно сфокусированный лазерный луч является оптической ловушкой, которая втягивает
В оптическом твизере сильно сфокусированный лазерный луч является оптической ловушкой, которая втягивает
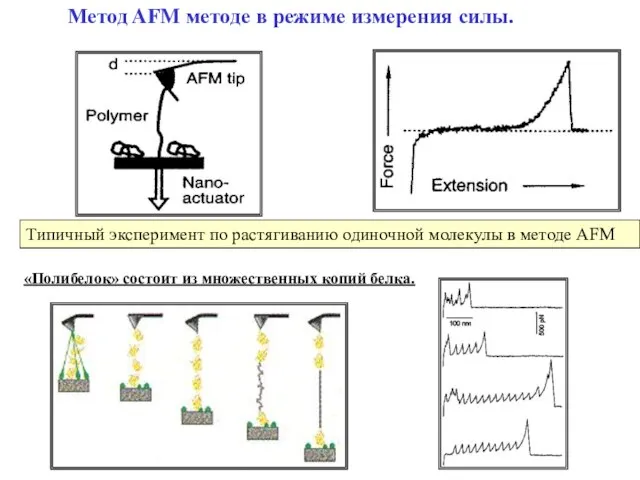
Слайд 30Метод AFM методе в режиме измерения силы.
Типичный эксперимент по растягиванию одиночной
Метод AFM методе в режиме измерения силы.
Типичный эксперимент по растягиванию одиночной
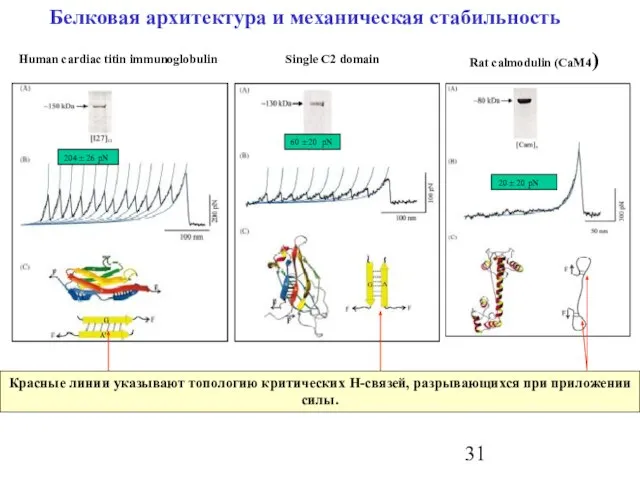
Слайд 31Белковая архитектура и механическая стабильность
Красные линии указывают топологию критических H-связей, разрывающихся при
Белковая архитектура и механическая стабильность
Красные линии указывают топологию критических H-связей, разрывающихся при
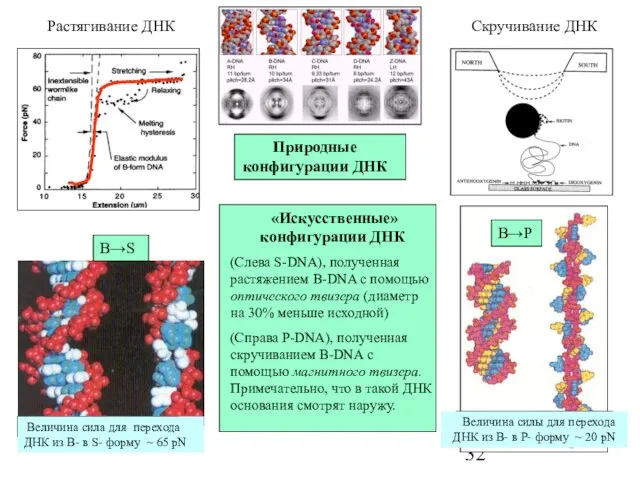
Слайд 32Природные конфигурации ДНК
«Искусственные» конфигурации ДНК
(Cлева S-DNA), полученная растяжением B-DNA с
Природные конфигурации ДНК
«Искусственные» конфигурации ДНК
(Cлева S-DNA), полученная растяжением B-DNA с
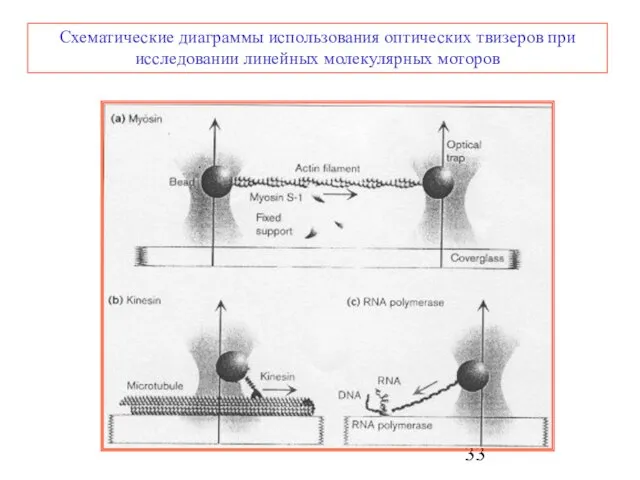
Слайд 33Схематические диаграммы использования оптических твизеров при исследовании линейных молекулярных моторов
Схематические диаграммы использования оптических твизеров при исследовании линейных молекулярных моторов
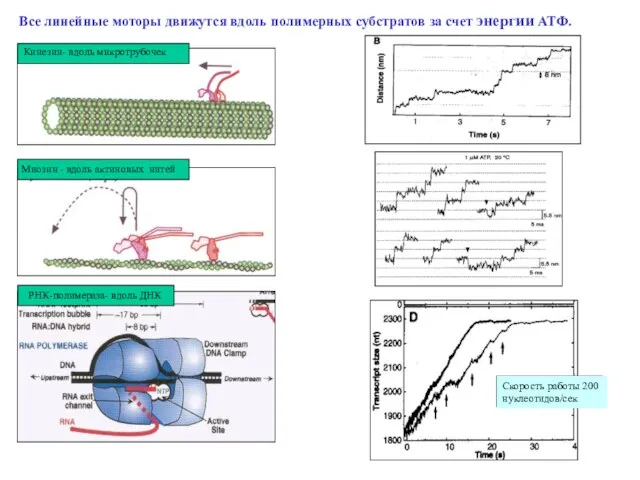
Слайд 34РНК-полимераза- вдоль ДНК
Кинезин- вдоль микротрубочек
Миозин - вдоль актиновых нитей
Все линейные моторы
РНК-полимераза- вдоль ДНК
Кинезин- вдоль микротрубочек
Миозин - вдоль актиновых нитей
Все линейные моторы
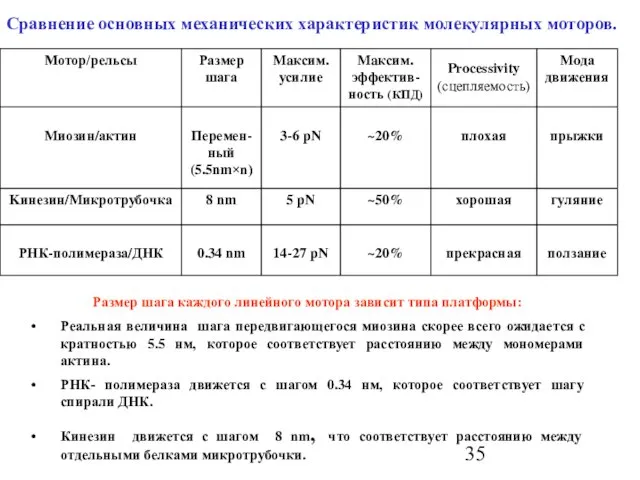
Слайд 35Сравнение основных механических характеристик молекулярных моторов.
Размер шага каждого линейного мотора зависит типа
Сравнение основных механических характеристик молекулярных моторов.
Размер шага каждого линейного мотора зависит типа
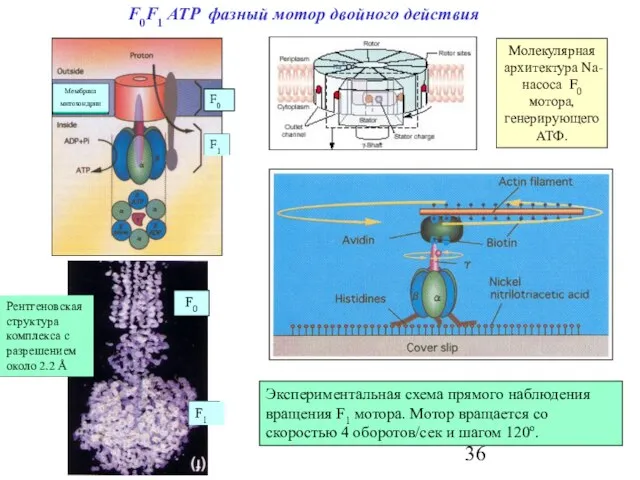
Слайд 36F0F1 ATP фазный мотор двойного действия
Молекулярная архитектура Na-насоса F0 мотора, генерирующего АТФ.
Экспериментальная
F0F1 ATP фазный мотор двойного действия
Молекулярная архитектура Na-насоса F0 мотора, генерирующего АТФ.
Экспериментальная
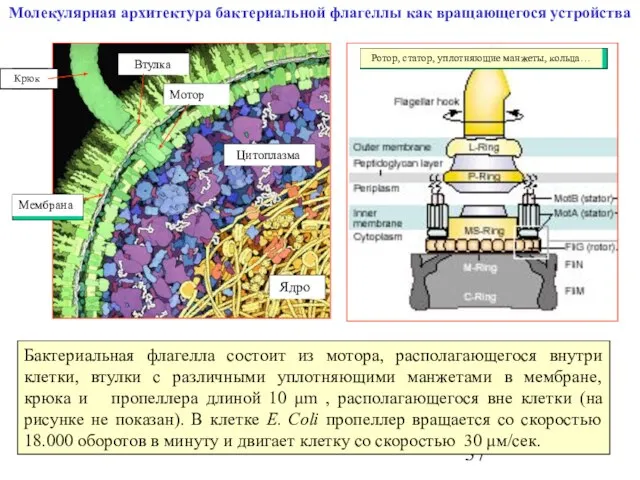
Слайд 37Молекулярная архитектура бактериальной флагеллы как вращающегося устройства
Бактериальная флагелла состоит из мотора, располагающегося
Молекулярная архитектура бактериальной флагеллы как вращающегося устройства
Бактериальная флагелла состоит из мотора, располагающегося
Слайд 38I. Serdyuk N. Zaccai J. Zaccai
“Physical methods in structural biology”
Cambridge, 2005
Introduction.
I. Serdyuk N. Zaccai J. Zaccai
“Physical methods in structural biology”
Cambridge, 2005
Introduction.

Слайд 39Курс лекций
проф. И.Н. Сердюк
“Физические методы в структурной биологии”
Введение. От термодинамики к одиночным
Курс лекций
проф. И.Н. Сердюк
“Физические методы в структурной биологии”
Введение. От термодинамики к одиночным

 Каркасное моделирование
Каркасное моделирование  Азбука в загадках-добавлялках
Азбука в загадках-добавлялках Оплата труда на предприятии
Оплата труда на предприятии Вышивка швом узелок и рококо
Вышивка швом узелок и рококо Или неиспользованный потенциал Информационные технологии в PR
Или неиспользованный потенциал Информационные технологии в PR Презентация на тему Физиология питания
Презентация на тему Физиология питания  РАСЧЕТ СТОИМОСТИ БЮДЖЕТНОЙ ОБРАЗОВАТЕЛЬНОЙ УСЛУГИ В БАЗОВОМ ОБРАЗОВАТЕЛЬНОМ УЧРЕЖДЕНИИ ПО МОДЕЛЬНОЙ МЕТОДИКЕ, РЕКОМЕНДОВАННОЙ М
РАСЧЕТ СТОИМОСТИ БЮДЖЕТНОЙ ОБРАЗОВАТЕЛЬНОЙ УСЛУГИ В БАЗОВОМ ОБРАЗОВАТЕЛЬНОМ УЧРЕЖДЕНИИ ПО МОДЕЛЬНОЙ МЕТОДИКЕ, РЕКОМЕНДОВАННОЙ М Кто курит табак – тот себе враг.
Кто курит табак – тот себе враг. п.Советский. 2008г.
п.Советский. 2008г. ТЕМА УРОКА
ТЕМА УРОКА ботулизм
ботулизм Использование технологии проектной деятельности при подготовке конкурсных работ учащимися объединения хранители природы
Использование технологии проектной деятельности при подготовке конкурсных работ учащимися объединения хранители природы сочинение по Лермонтову
сочинение по Лермонтову Опыт работы МОУ Курагинской СОШ №1
Опыт работы МОУ Курагинской СОШ №1 Структура доходов и расходов бюджета
Структура доходов и расходов бюджета Самооценка готовности к профессионально-личностному саморазвитию будущих педагогов
Самооценка готовности к профессионально-личностному саморазвитию будущих педагогов Оценка эффективности системы товародвижения
Оценка эффективности системы товародвижения Роман в стихах А.С. Пушкина « Евгений Онегин»
Роман в стихах А.С. Пушкина « Евгений Онегин» Рождество Христово
Рождество Христово Фазировка. Косвенные методы фазировки
Фазировка. Косвенные методы фазировки Русь в XIII-XV веках
Русь в XIII-XV веках Формирование смысложизненных ориентаций школьников
Формирование смысложизненных ориентаций школьников Наибольший общий делитель 6 класс
Наибольший общий делитель 6 класс Семь жемчужин Египта
Семь жемчужин Египта Программы для ГРВкомпании «МедЭО»
Программы для ГРВкомпании «МедЭО» Презентация на тему Защита организма от излучения
Презентация на тему Защита организма от излучения Что такое фотография?
Что такое фотография? Староакульшетская школа
Староакульшетская школа