- Биоинформатика, или молекулярная биология in silico

Содержание
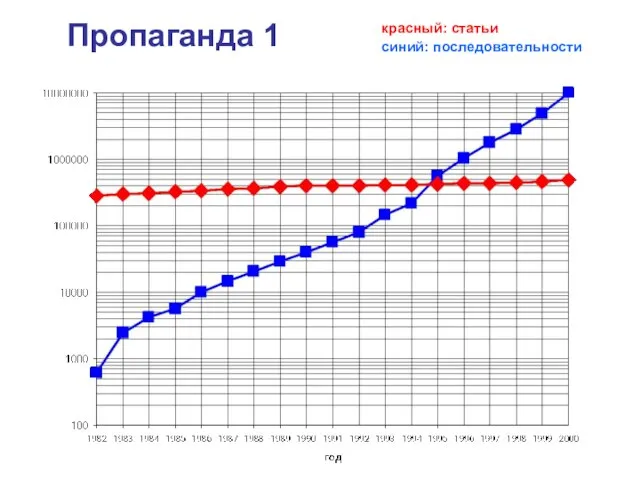
- 2. Пропаганда 1 красный: статьи синий: последовательности
- 3. Анализ индивидуальных генов Поиск родственных белков в банках последовательностей – перенос функции от гомологов Функциональные сайты
- 4. Анализ на уровне индивидуальных генов даёт возможность охарактеризовать 50-75% генов в новом геноме Но: ~100 универсально
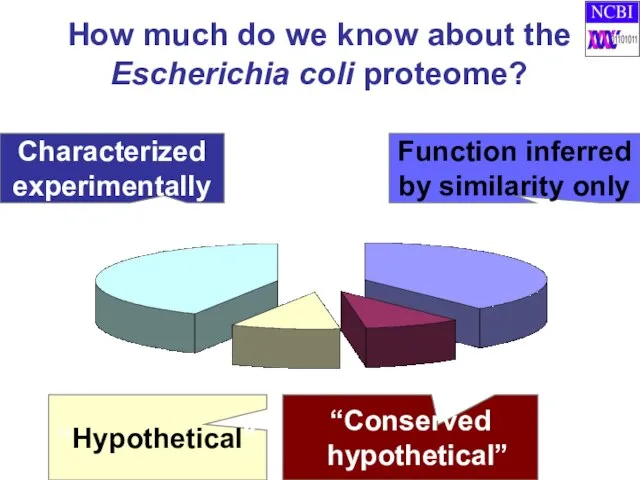
- 5. Characterized experimentally “Hypothetical” Function inferred by similarity only “Conserved hypothetical” How much do we know about
- 6. Пропаганда – 2 Полные геномы
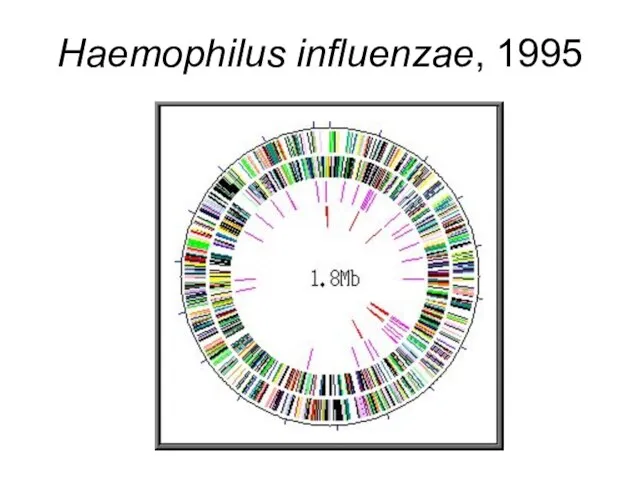
- 7. Haemophilus influenzae, 1995
- 8. Vibrio cholerae, 2000
- 9. Сравнительно-геномные подходы Positional clustering Phylogenetic profiling Gene fusions
- 10. Metabolic pathways
- 11. Functionally dependent genes tend to cluster on chromosomes in many different organisms
- 12. More genomes (stronger links) => highly significant clustering
- 13. … особенно в линейных путях (справа)
- 14. Распределение уровней связи (бимодальное для изоферментов, монотонное для субъединиц)
- 15. Phyletic profiles in the Phe/Tyr pathway
- 16. Arithmetics of phyletic patterns 3-dehydroquinate dehydratase (EC 4.2.1.10): Class I (AroD) COG0710 aompkzyq---lb-e----n---i-- Class II (AroQ)
- 17. STRING: trpB – fusions
- 18. Утилизация пектина E. chrysanthemi
- 19. … и транспорт олигогалактуронатов E. chrysanthemi Y. pestis K. pneumoniae
- 20. YpaA: транспортёр рибофлавина 5 предсказанных ТМ-сегментов => потенциальный транспортёр регуляторный RFN-элемент => ко-регуляция с генами метаболизма
- 21. Метаболическая реконструкция пути биосинтеза лизина: Идентификация пути ацетилированных интермедиатов в B. subtilis и родственных бактериях
- 22. Идентификация пути ацетилированных интермедиатов - 0 dapD (yquQ): ортолог известного гена E. coli
- 23. Идентификация пути ацетилированных интермедиатов - 1 patA: пиридоксаль-фосфат-зависимая аминотрансфераза (по гомологии) ко-локализуется и ко-регулируется с генами
- 24. Идентификация пути ацетилированных интермедиатов - 2 ykuR: N-ацил-L-аминокислота амидогидролаза (по гомологии) ко-локализуется и ко-регулируется с геном
- 25. Идентификация пути ацетилированных интермедиатов - 3 dapX: dapF отсутствует у некоторых бактерий (Staphylococcus aureus, Oenococcus oeni,
- 26. Сравнительная геномика систем утилизации цинка Две роли цинка в бактериях: Структурная в ДНК-полимеразах, праймазах, рибосомных белках
- 27. Регуляторы и сигналы nZUR-γ nZUR-α AdcR pZUR TTAACYRGTTAA GATATGTTATAACATATC GAAATGTTATANTATAACATTTC GTAATGTAATAACATTAC TAAATCGTAATNATTACGATTTA
- 28. Цинк и паралоги белков рибосом nZUR pZUR AdcR
- 29. (в скобках – мотив «цинковая лента») nZUR pZUR AdcR
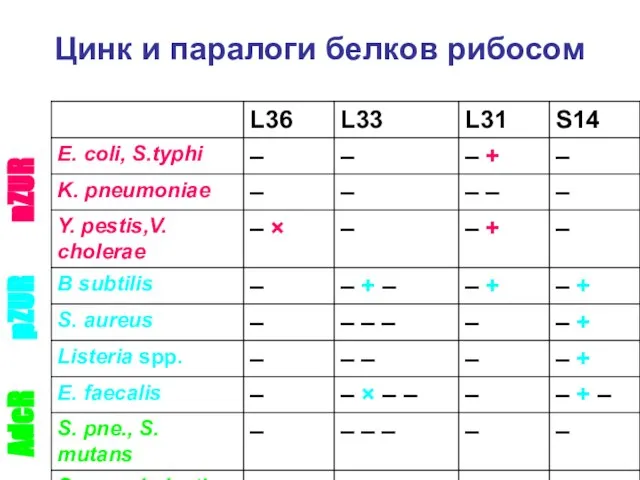
- 30. Сводка наблюдений: Makarova-Ponomarev-Koonin, 2001: L36, L33, L31, S14 – это единственные рибосомные белки, дуплицированные более, чем
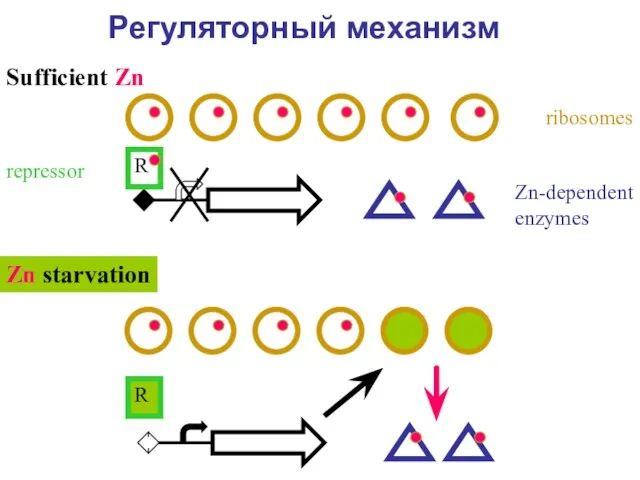
- 31. Плохой сценарий достаточно цинка недостаточно цинка: весь цинк потреблен рибосомами, ферменты голодают
- 32. Хороший сценарий достаточно цинка недостаточно цинка: часть рибосом включает белки, не содержащие цинка – остается для
- 33. Регуляторный механизм ribosomes Zn-dependent enzymes R Sufficient Zn Zn starvation R repressor
- 34. Предсказание … (Proc Natl Acad Sci U S A. 2003 Aug 19;100(17):9912-7.) … и подтверждения (Mol
- 35. Регуляторная система «с нуля под ключ» Консервативный сигнал перед генами рибонуклеотид-редуктаз Потенциальный регулятор (через филогенетический паттерн
- 36. Другие члены регулона Реутилизация дезоксирибонуклеотидов Репликация (ДНК-лигазы, топоизомеразы, ДНК-полимеразы
- 37. Как регулируется: репрессия в результате кооперативного связывания
- 39. Скачать презентацию
Слайд 3Анализ индивидуальных генов
Поиск родственных белков в банках последовательностей – перенос функции от
Анализ индивидуальных генов
Поиск родственных белков в банках последовательностей – перенос функции от

Слайд 4Анализ на уровне индивидуальных генов даёт возможность охарактеризовать 50-75% генов в новом
Анализ на уровне индивидуальных генов даёт возможность охарактеризовать 50-75% генов в новом

Слайд 5Characterized
experimentally
“Hypothetical”
Function inferred
by similarity only
“Conserved
hypothetical”
How much do we know about the
Characterized
experimentally
“Hypothetical”
Function inferred
by similarity only
“Conserved
hypothetical”
How much do we know about the
Слайд 6Пропаганда – 2 Полные геномы
Пропаганда – 2 Полные геномы
Слайд 7Haemophilus influenzae, 1995
Haemophilus influenzae, 1995
Слайд 8Vibrio cholerae, 2000
Vibrio cholerae, 2000
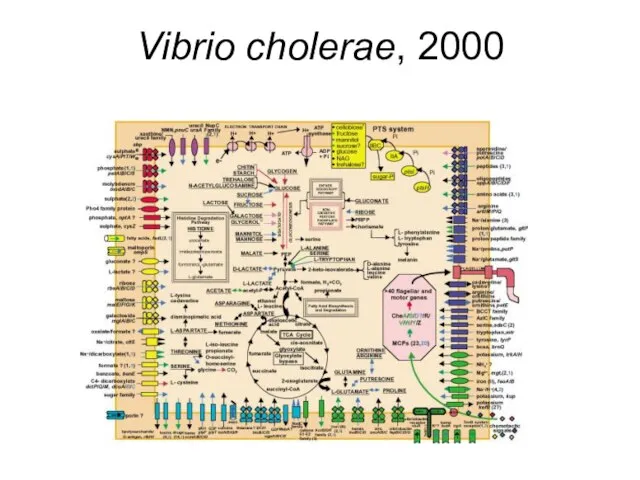
Слайд 9Сравнительно-геномные подходы
Positional clustering
Phylogenetic profiling
Gene fusions
Сравнительно-геномные подходы
Positional clustering
Phylogenetic profiling
Gene fusions

Слайд 10Metabolic pathways
Metabolic pathways
Слайд 11Functionally dependent genes tend to cluster on chromosomes in many different organisms
Functionally dependent genes tend to cluster on chromosomes in many different organisms
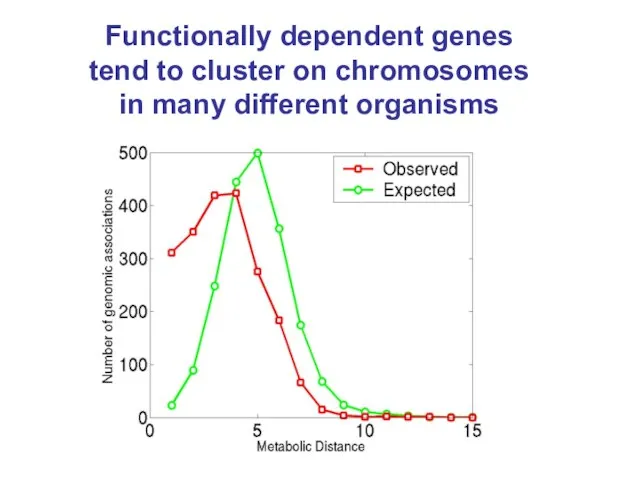
Слайд 12More genomes (stronger links)
=> highly significant clustering
More genomes (stronger links)
=> highly significant clustering
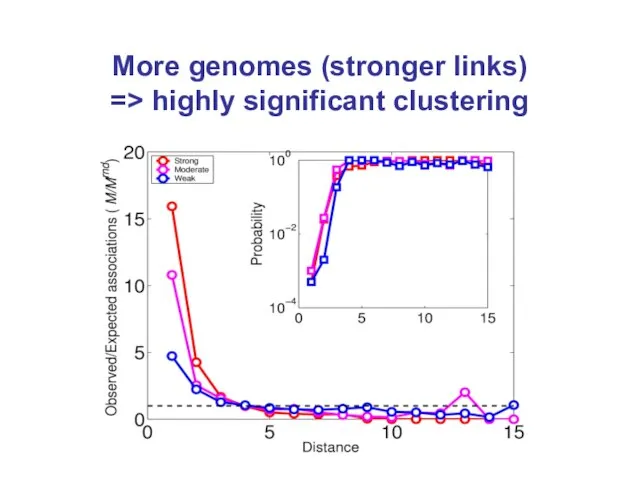
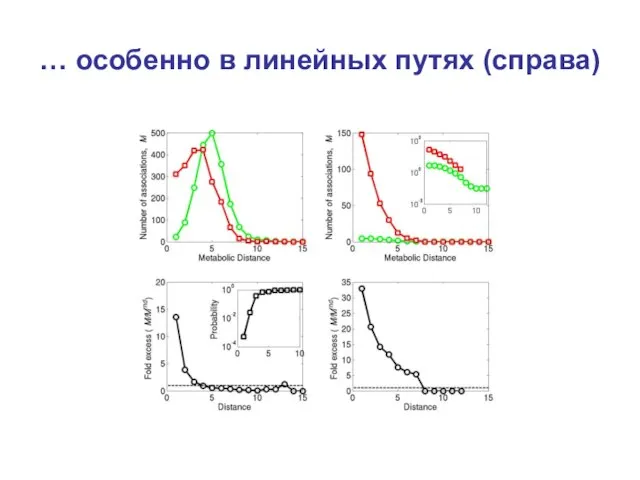
Слайд 13… особенно в линейных путях (справа)
… особенно в линейных путях (справа)
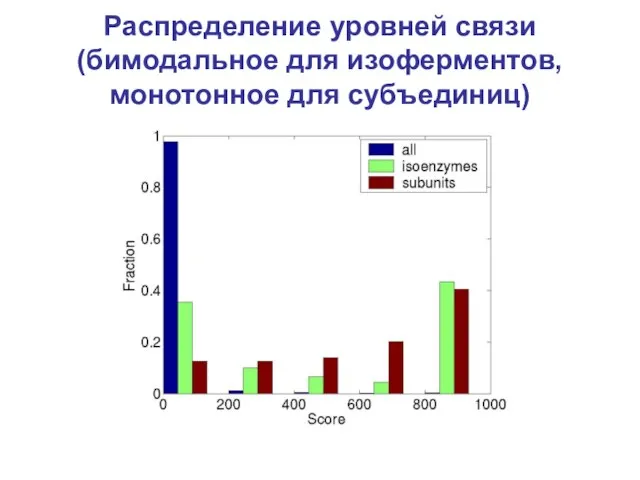
Слайд 14Распределение уровней связи
(бимодальное для изоферментов, монотонное для субъединиц)
Распределение уровней связи
(бимодальное для изоферментов, монотонное для субъединиц)
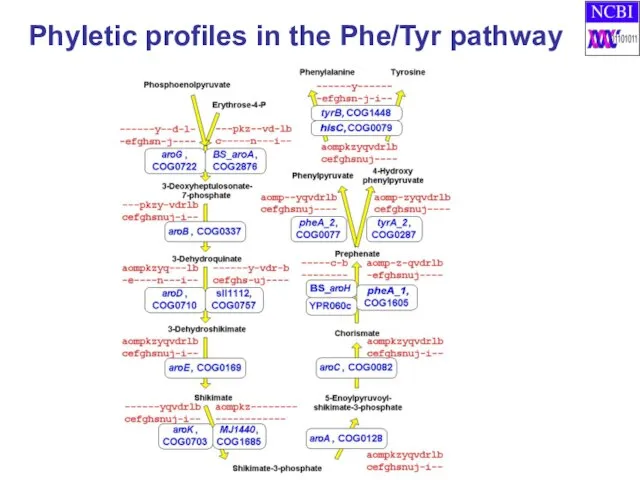
Слайд 15Phyletic profiles in the Phe/Tyr pathway
Phyletic profiles in the Phe/Tyr pathway
Слайд 16Arithmetics of phyletic patterns
3-dehydroquinate dehydratase (EC 4.2.1.10):
Class I (AroD) COG0710 aompkzyq---lb-e----n---i--
Class
Arithmetics of phyletic patterns
3-dehydroquinate dehydratase (EC 4.2.1.10):
Class I (AroD) COG0710 aompkzyq---lb-e----n---i--
Class

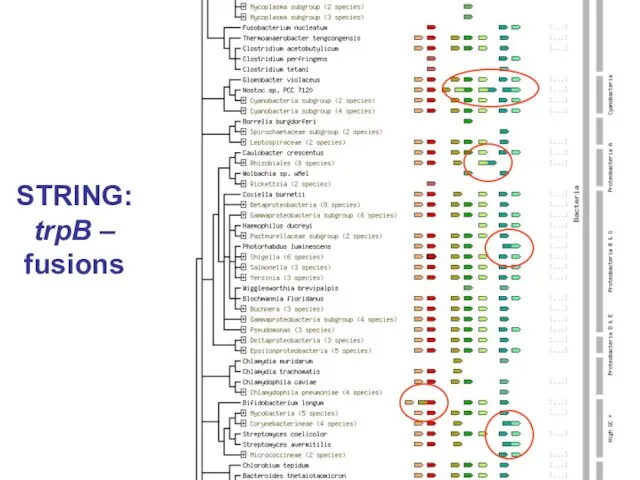
Слайд 17STRING: trpB – fusions
STRING: trpB – fusions
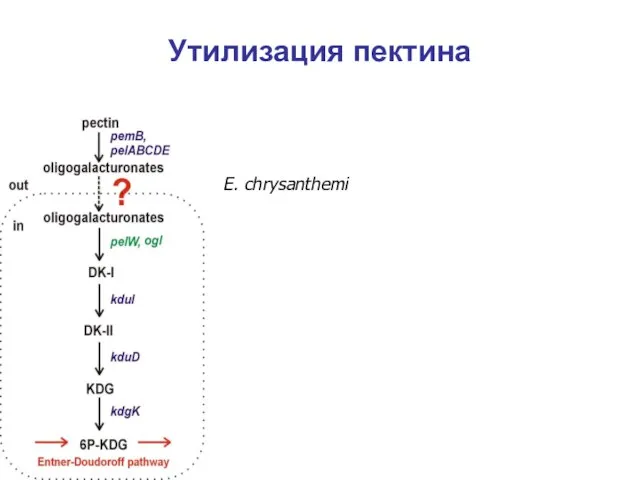
Слайд 18Утилизация пектина
E. chrysanthemi
Утилизация пектина
E. chrysanthemi
Слайд 19… и транспорт олигогалактуронатов
E. chrysanthemi
Y. pestis
K. pneumoniae
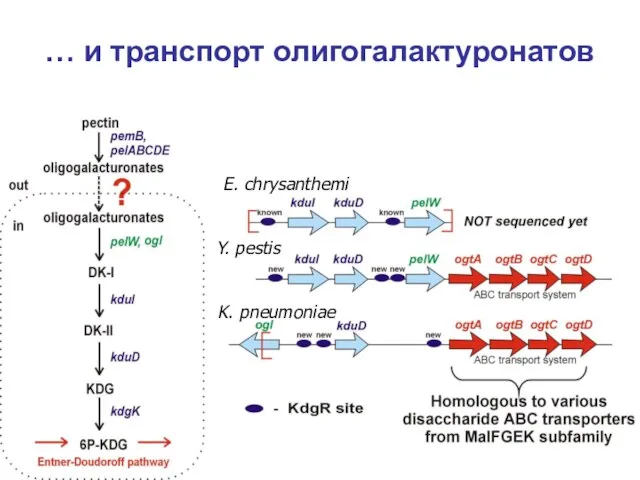
… и транспорт олигогалактуронатов
E. chrysanthemi
Y. pestis
K. pneumoniae
Слайд 20YpaA: транспортёр рибофлавина
5 предсказанных ТМ-сегментов => потенциальный транспортёр
регуляторный RFN-элемент => ко-регуляция с
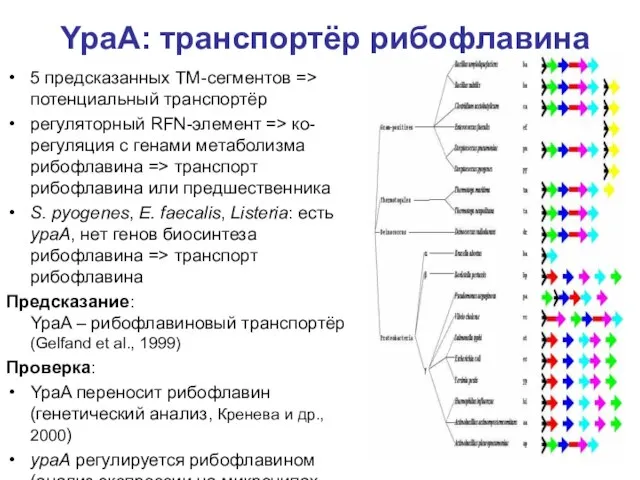
YpaA: транспортёр рибофлавина
5 предсказанных ТМ-сегментов => потенциальный транспортёр
регуляторный RFN-элемент => ко-регуляция с
Слайд 21Метаболическая реконструкция пути биосинтеза лизина: Идентификация пути ацетилированных интермедиатов в B. subtilis
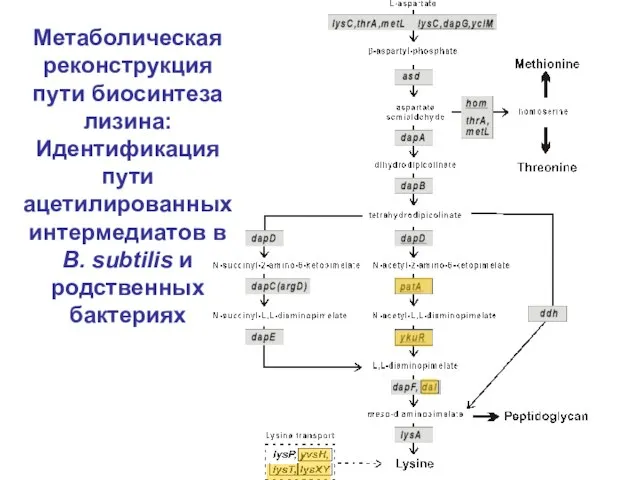
Метаболическая реконструкция пути биосинтеза лизина: Идентификация пути ацетилированных интермедиатов в B. subtilis
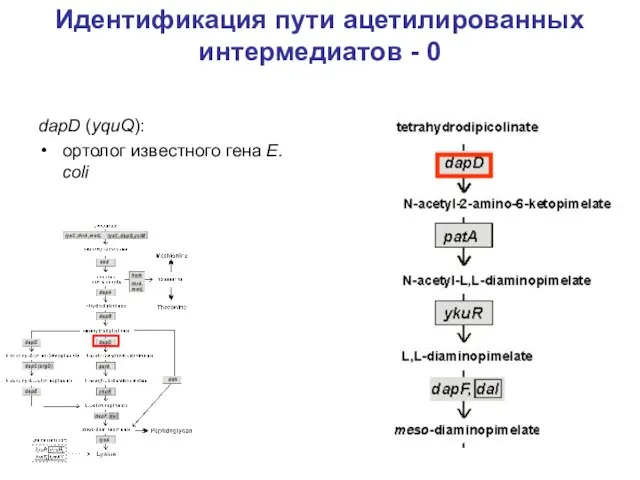
Слайд 22Идентификация пути ацетилированных интермедиатов - 0
dapD (yquQ):
ортолог известного гена E. coli
Идентификация пути ацетилированных интермедиатов - 0
dapD (yquQ):
ортолог известного гена E. coli
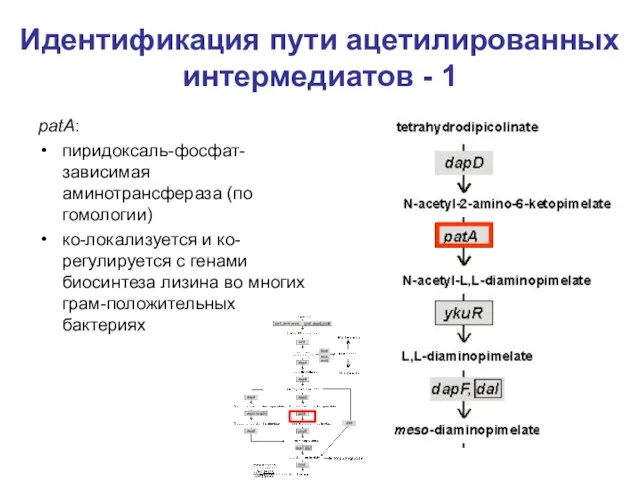
Слайд 23Идентификация пути ацетилированных интермедиатов - 1
patA:
пиридоксаль-фосфат-зависимая аминотрансфераза (по гомологии)
ко-локализуется и ко-регулируется
Идентификация пути ацетилированных интермедиатов - 1
patA:
пиридоксаль-фосфат-зависимая аминотрансфераза (по гомологии)
ко-локализуется и ко-регулируется
Слайд 24Идентификация пути ацетилированных интермедиатов - 2
ykuR:
N-ацил-L-аминокислота амидогидролаза (по гомологии)
ко-локализуется и ко-регулируется
Идентификация пути ацетилированных интермедиатов - 2
ykuR:
N-ацил-L-аминокислота амидогидролаза (по гомологии)
ко-локализуется и ко-регулируется

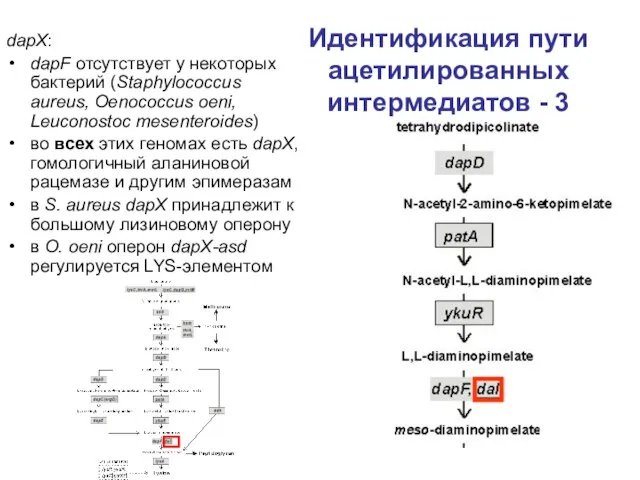
Слайд 25Идентификация пути ацетилированных интермедиатов - 3
dapX:
dapF отсутствует у некоторых бактерий (Staphylococcus
Идентификация пути ацетилированных интермедиатов - 3
dapX:
dapF отсутствует у некоторых бактерий (Staphylococcus
Слайд 26Сравнительная геномика систем утилизации цинка
Две роли цинка в бактериях:
Структурная в ДНК-полимеразах,
Сравнительная геномика систем утилизации цинка
Две роли цинка в бактериях:
Структурная в ДНК-полимеразах,

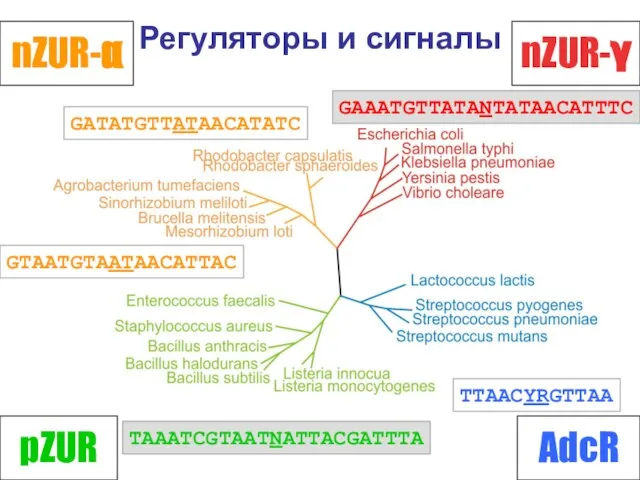
Слайд 27Регуляторы и сигналы
nZUR-γ
nZUR-α
AdcR
pZUR
TTAACYRGTTAA
GATATGTTATAACATATC
GAAATGTTATANTATAACATTTC
GTAATGTAATAACATTAC
TAAATCGTAATNATTACGATTTA
Регуляторы и сигналы
nZUR-γ
nZUR-α
AdcR
pZUR
TTAACYRGTTAA
GATATGTTATAACATATC
GAAATGTTATANTATAACATTTC
GTAATGTAATAACATTAC
TAAATCGTAATNATTACGATTTA
Слайд 28Цинк и паралоги белков рибосом
nZUR
pZUR
AdcR
Цинк и паралоги белков рибосом
nZUR
pZUR
AdcR
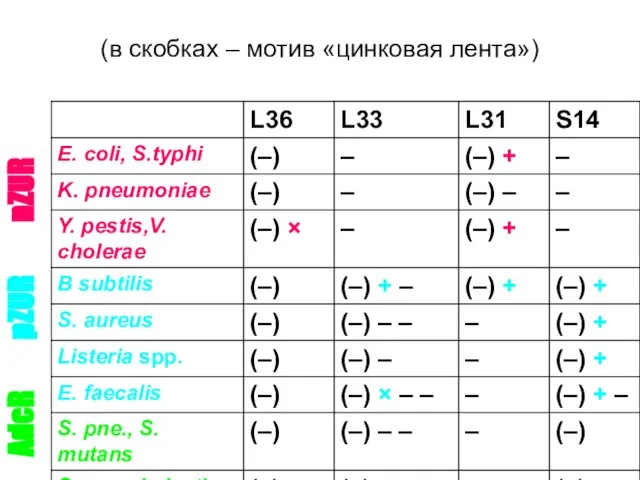
Слайд 29(в скобках – мотив «цинковая лента»)
nZUR
pZUR
AdcR
(в скобках – мотив «цинковая лента»)
nZUR
pZUR
AdcR
Слайд 30Сводка наблюдений:
Makarova-Ponomarev-Koonin, 2001:
L36, L33, L31, S14 – это единственные рибосомные белки, дуплицированные
Сводка наблюдений:
Makarova-Ponomarev-Koonin, 2001:
L36, L33, L31, S14 – это единственные рибосомные белки, дуплицированные

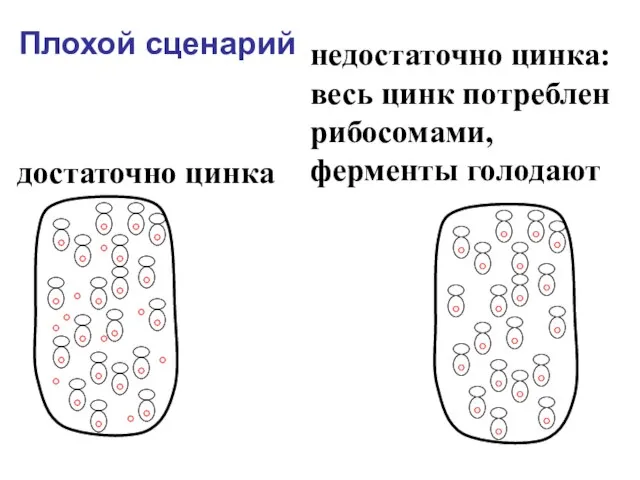
Слайд 31Плохой сценарий
достаточно цинка
недостаточно цинка: весь цинк потреблен рибосомами, ферменты голодают
Плохой сценарий
достаточно цинка
недостаточно цинка: весь цинк потреблен рибосомами, ферменты голодают
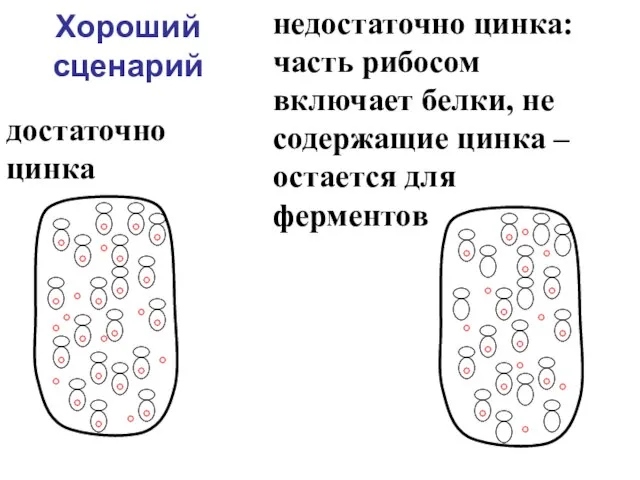
Слайд 32Хороший сценарий
достаточно цинка
недостаточно цинка: часть рибосом включает белки, не содержащие цинка –
Хороший сценарий
достаточно цинка
недостаточно цинка: часть рибосом включает белки, не содержащие цинка –
Слайд 33Регуляторный механизм
ribosomes
Zn-dependent
enzymes
R
Sufficient Zn
Zn starvation
R
repressor
Регуляторный механизм
ribosomes
Zn-dependent
enzymes
R
Sufficient Zn
Zn starvation
R
repressor
Слайд 34Предсказание …
(Proc Natl Acad Sci U S A. 2003 Aug 19;100(17):9912-7.)
…
Предсказание …
(Proc Natl Acad Sci U S A. 2003 Aug 19;100(17):9912-7.)
…

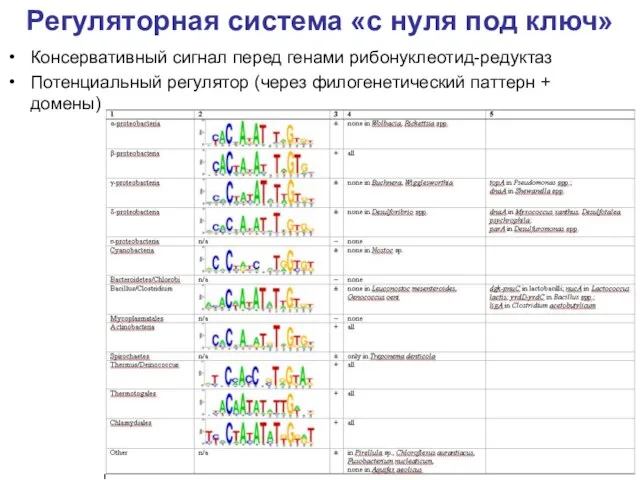
Слайд 35Регуляторная система «с нуля под ключ»
Консервативный сигнал перед генами рибонуклеотид-редуктаз
Потенциальный регулятор (через
Регуляторная система «с нуля под ключ»
Консервативный сигнал перед генами рибонуклеотид-редуктаз
Потенциальный регулятор (через
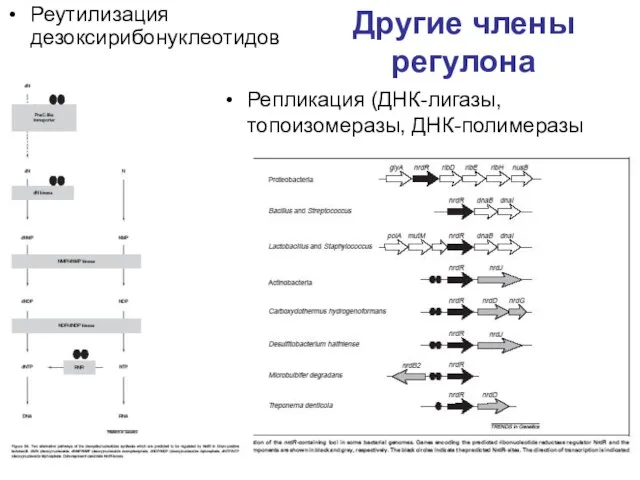
Слайд 36Другие члены регулона
Реутилизация дезоксирибонуклеотидов
Репликация (ДНК-лигазы, топоизомеразы, ДНК-полимеразы
Другие члены регулона
Реутилизация дезоксирибонуклеотидов
Репликация (ДНК-лигазы, топоизомеразы, ДНК-полимеразы
Слайд 37Как регулируется: репрессия в результате кооперативного связывания
Как регулируется: репрессия в результате кооперативного связывания

 Результаты исследования «SEO-альманах» ТопЭксперт.РФ
Результаты исследования «SEO-альманах» ТопЭксперт.РФ Решение текстовых задач арифметическим способом по УМК Г.В.Дорофеева
Решение текстовых задач арифметическим способом по УМК Г.В.Дорофеева Важность правильного позиционирования брендав социальных медиа
Важность правильного позиционирования брендав социальных медиа Элиста
Элиста Авторская песня и ее особенности
Авторская песня и ее особенности Экологические праздники
Экологические праздники UK rivers
UK rivers Государственная символика
Государственная символика Круговорот кислорода в природе
Круговорот кислорода в природе Правовые основы воинской службы
Правовые основы воинской службы Профессия Повар-кондитер
Профессия Повар-кондитер Ради жизни на земле
Ради жизни на земле Праздник Суздальского огурца
Праздник Суздальского огурца Решение задач на уроках математики с использованием статистических данных по правонарушениям
Решение задач на уроках математики с использованием статистических данных по правонарушениям Энергия потока людей
Энергия потока людей Спортивная зима
Спортивная зима Самые опасные животные
Самые опасные животные Безопасность информации электронного правительства
Безопасность информации электронного правительства Презентация на тему 12 апреля –День космонавтики
Презентация на тему 12 апреля –День космонавтики Наши клиенты
Наши клиенты Презентация на тему Решение задач по нахождению величин по сумме и разности
Презентация на тему Решение задач по нахождению величин по сумме и разности Первая украинская горная экспедиция в Иран Сложные экспедиции в малодоступные уголки планеты
Первая украинская горная экспедиция в Иран Сложные экспедиции в малодоступные уголки планеты ЭФФЕКТИВНОСТЬ И ПЕРСПЕКТИВЫ РАЗВИТИЯ ТЕПЛОФИКАЦИИ В СОВРЕМЕННЫХ ЭКОНОМИЧЕСКИХ УСЛОВИЯХ
ЭФФЕКТИВНОСТЬ И ПЕРСПЕКТИВЫ РАЗВИТИЯ ТЕПЛОФИКАЦИИ В СОВРЕМЕННЫХ ЭКОНОМИЧЕСКИХ УСЛОВИЯХ Входной контроль по вопросам перехода к Федеральному государственному образовательному стандарту начального общего образования
Входной контроль по вопросам перехода к Федеральному государственному образовательному стандарту начального общего образования МО учителей естественно-географического цикла
МО учителей естественно-географического цикла Раздел «Система мониторинга достижений детьми планируемых результатов освоения Образовательной программы»
Раздел «Система мониторинга достижений детьми планируемых результатов освоения Образовательной программы» Органические вещества клетки (10 класс)
Органические вещества клетки (10 класс) khram_moego_detstva
khram_moego_detstva