- Цитоплазма

Содержание
- 2. ЦИТОПЛАЗМА Цитоплазма составляет основную массу клетки - это все ее внутреннее содержание, за исключением ядра Содержит
- 3. Цитозоль это структурированный коллоид, состоящий из сложной смеси растворенных в воде органических макромолекул - белков, жиров,
- 4. Химический состав и свойства цитозоля: Неорганические вещества: вода, соли, газы. Органические вещества: белки, углеводы, липиды, нуклеотиды.
- 5. Цитоскелет. К элементам цитоскелета относят белковые фибриллярные структуры, расположенные в цитоплазме клетки: микротрубочки, актиновые и промежуточные
- 6. Цитоскелет эукариот. Актиновые микрофиламенты окрашены в красный, микротрубочки — в зелёный, ядра клеток — в голубой
- 7. Структура микротрубочки
- 8. Структура молекулы миозина и комплекса актин-тропомиозин
- 9. Функции цитоскелета: 1. Поддержка объема и формы клеток 2. Изменение формы клеток 3. Передвижение органелл и
- 10. ВКЛЮЧЕНИЯ В цитоплазме клеток также включения - непостоянные компоненты, выполняющие функцию запасания питательных веществ (капли жира,
- 11. ВКЛЮЧЕНИЯ ГЛИКОГЕНА В КЛЕТКАХ ПЕЧЕНИ Окраска кармином с докраской ядер гематоксилином. Включения гликогена (красное окрашивание), ядра
- 12. ЖИРОВЫЕ ВКЛЮЧЕНИЯ В КЛЕТКАХ ПЕЧЕНИ Окраска оксидом осмия с докраской ядер сафранином жировые включения (черные шарики)
- 13. Органеллы - постоянные компоненты цитоплазмы, имеющие определенную структуру и выполняющие определенные функции. Классификация органелл по наличию
- 14. Классификация органелл по выполняемой функции: А) Органеллы общего назначения: митохондрии, лизосомы, пероксисомы, комплекс Гольджи, эндоплазматическая сеть,
- 15. Эндоплазматический ретикулум (эндоплазматическая сеть, ЭПР или ЭПС) - система переходящих друг в друга мембранных отсеков (трубок
- 16. ОДНОМЕМБРАННЫЕ ОРГАНЕЛЛЫ Эндоплазматическая сеть Агранулярная Функции гладкой ЭПС: 1. Синтез фосфолипидов и углеводов; 2. Накопление и
- 17. Ганулярная ЭПС Функции зернистой ЭПС: 1. Участие в процессе синтеза белков; 2. Накопление и модификация белков,
- 18. Комплекс Гольжи (Аппарат Гольджи) - стопка плоских мембранных цистерн. В цистернах созревают белки, синтезированные на мембранах
- 19. Функции комлекса Гольджи: 1. Накопление и модификация синтезированных в ЭПС макромолекул; 2. Образование сложных секретов и
- 20. Цис- и транс- компартменты аппарата Гольджи
- 21. Лизосома небольшая органелла, ограниченная одной мембраной. В ней находятся литические ферменты, способные расщепить все биополимеры. Первичные
- 22. Функции лизосом: 1. Переваривание веществ, поступающих в клетку извне в процессе фагоцитоза. 2. Переваривание поврежденных внутриклеточных
- 23. Пероксисома – небольшая органелла, ограниченная одной мембраной, содержащая большое количество ферментов, катализирующих окислительно-восстановительные реакции (оксидазы D-аминокислот,
- 24. Вакуоли (только в растительной клетке) Они отделены от цитоплазмы одинарной мембраной - тонопластом. Полость вакуоли заполнена
- 25. Двумембранные органеллы Митохондрии. Внутренний просвет митохондрий, называемый матриксом, отграничен от цитоплазмы двумя мембранами, наружной и внутренней,
- 26. Митохондрии. Схема строения. Электронограмма.
- 27. 1 – наружная мембрана, 2 – внутренняя мембрана, 3 – кристы, 4- матрикс Схема строения митохондрии
- 28. Пластиды - органеллы растений и некоторых фотосинтезирующих простейших. Покрыты двойной мембраной и имеют в своём составе
- 29. Пластиды : хлоропласты, хромопласты та лейкопласти. Хлоропласты
- 30. Схема строения хлоропласта 1- граны, 2- наружная мембрана, 3- строма, 4 – ламеллы, 5- включения (зерна
- 31. Пространственная модель хлоропласта
- 32. Рибосома - немембранная органелла сферической или эллипсоидной формы, состоящая из большой и малой субъединиц. Синтез рибосом
- 33. Схема строения рибосомы В эукариотических клетках рибосомы располагаются на мембранах эндоплазматического ретикулума и в цитоплазме. Нередко
- 34. Рибосомы Схема строения прокариотической рибосомы Схема строения эукариотической рибосомы
- 35. Клеточный центр представлен двумя центриолями ориентированными перпендикулярно друг к другу. Центриоли представляют собой цилиндрические белковые структуры,
- 36. Клеточный центр.
- 37. Базальные тельца лежат в цитоплазме в основе ресничек и жгутиков. Каждое кинетосома собой цилиндр, образованным девятью
- 38. Реснички и жгутики
- 40. Клеточное ядро
- 41. Ядро – центральный информационный аппарат клетки. Структура интерфазного ядра. Хроматин: эухроматин, гетерохроматин. Термин «ядро» впервые был
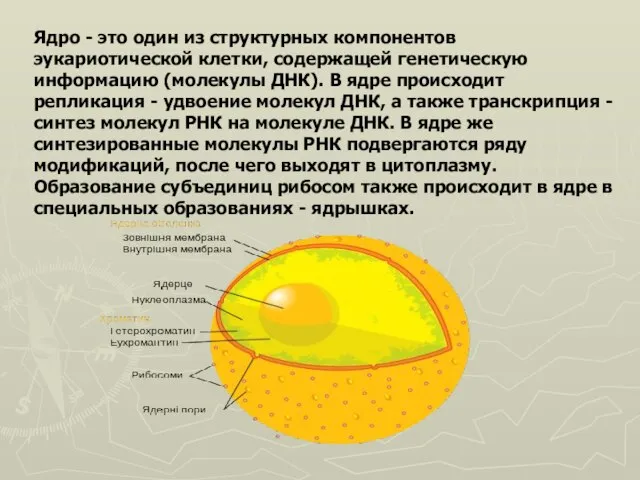
- 42. Ядро - это один из структурных компонентов эукариотической клетки, содержащей генетическую информацию (молекулы ДНК). В ядре
- 43. История открытия
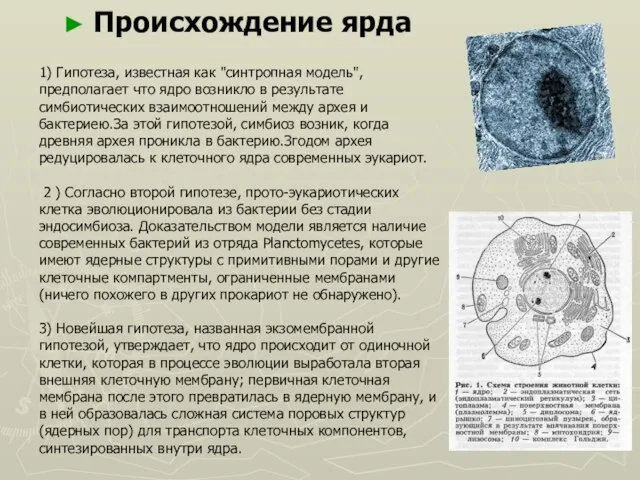
- 44. 1) Гипотеза, известная как "синтропная модель", предполагает что ядро возникло в результате симбиотических взаимоотношений между архея
- 45. . Ядро и некоторые органеллы эукариотической клетки. Электронограмма.
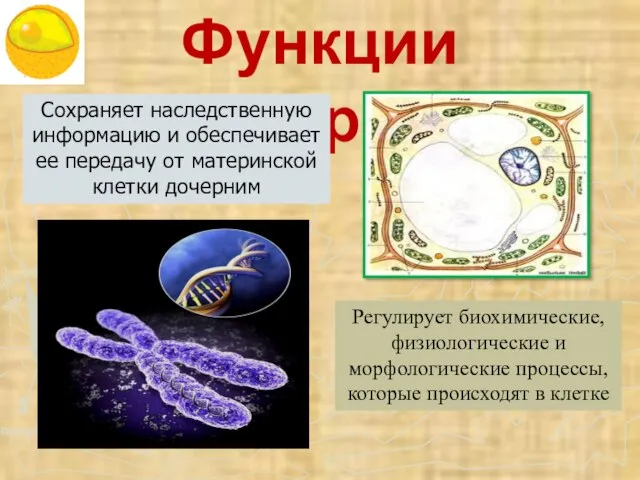
- 46. Функции ядра Сохраняет наследственную информацию и обеспечивает ее передачу от материнской клетки дочерним Регулирует биохимические, физиологические
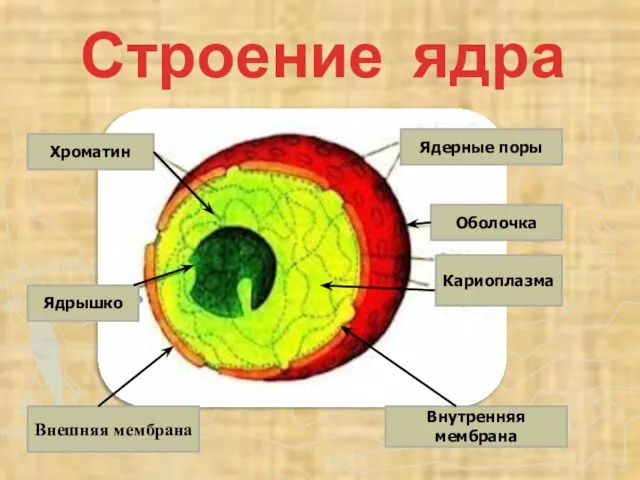
- 47. Строение ядра Ядрышко Внешняя мембрана Оболочка Ядерные поры Хроматин Внутренняя мембрана Кариоплазма
- 48. Ядро отделено от цитоплазмы ядерной оболочкой, образованной. Ядерная оболочка (кариолемма) состоит из двух биологических мембран (внешней
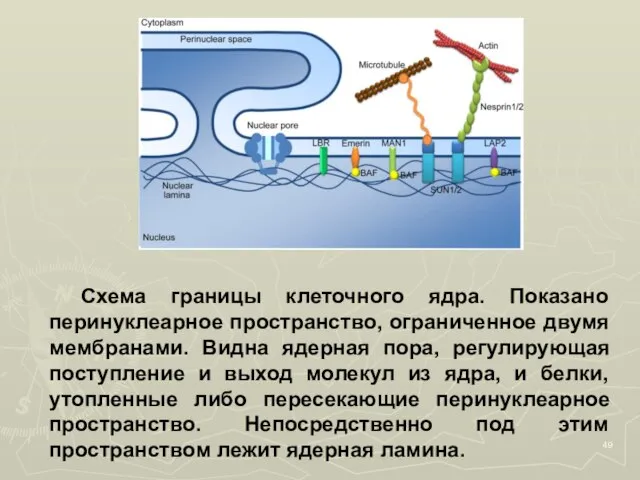
- 49. Схема границы клеточного ядра. Показано перинуклеарное пространство, ограниченное двумя мембранами. Видна ядерная пора, регулирующая поступление и
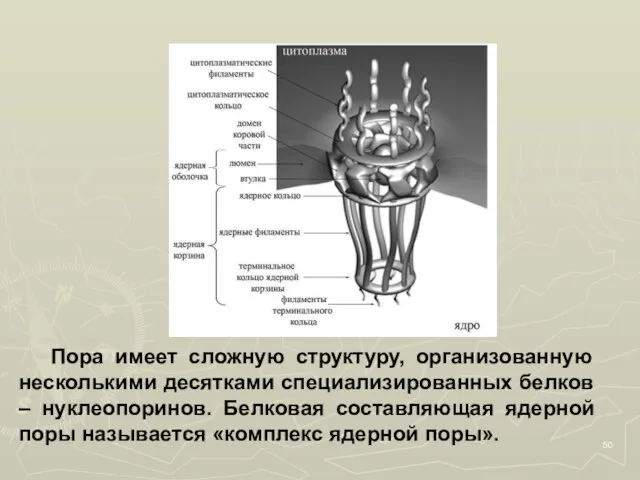
- 50. Пора имеет сложную структуру, организованную несколькими десятками специализированных белков – нуклеопоринов. Белковая составляющая ядерной поры называется
- 51. Функции ядерной оболочки: разграничение содержимого ядра и цитоплазмы, транспорт веществ между ядром и цитоплазмой. Ядерная оболочка
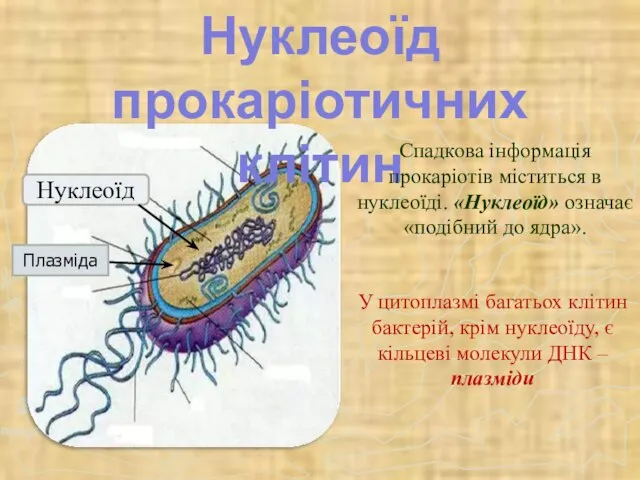
- 52. Нуклеоїд прокаріотичних клітин Спадкова інформація прокаріотів міститься в нуклеоїді. «Нуклеоїд» означає «подібний до ядра». У цитоплазмі
- 53. Между структурными компонентами ядра заключена жидкая фаза клеточного ядра, кариоплазма, в которой протекают процессы ядерного метаболизма
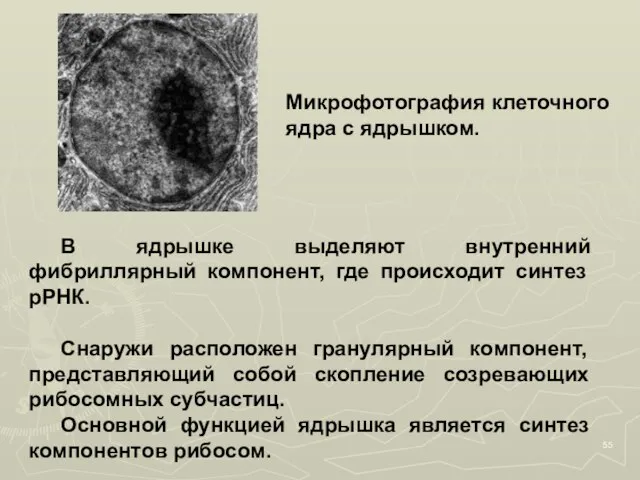
- 54. Ядрышко находится внутри ядра, не имеет собственной мембранной оболочки, хорошо различимо под световым и электронным микроскопом.
- 55. В ядрышке выделяют внутренний фибриллярный компонент, где происходит синтез рРНК. Снаружи расположен гранулярный компонент, представляющий собой
- 56. Хроматин. В интерфазном ядре выявляются зоны плотного вещества, которое хорошо воспринимает красители. Благодаря способности хорошо окрашиваться
- 58. Функции гистоновых белков: - создание специфической укладки ДНК - защита ДНК - препятствование транскрипции Функции негистоновых
- 59. Гистоны Н2А, Н2В, Н3, Н4 входят в состав нуклеосом, Н1 находится между нуклеосомами. Нуклеосома состоит из
- 60. Вокруг одной нуклеосомы лежат 165-200 нуклеотидных пар, а ДНК делает 1,67 - 1,8 витков. В среднем
- 61. Схема строения нуклеосом и дезоксирибонуклеопротеидного тяжа (нуклеофиламента).
- 62. В разные моменты существования клетки хроматин имеет разную пространственную организацию. Он способен в деконденсации (деспирализации) и
- 63. Эухроматин функционально активен. С него происходит считывание информации (транскрипция). Гетерохроматин функционально не активен. Информация с него
- 64. 2. Хромосомный, геномный и генный уровни организации наследственного материала. Хромосомный уровень организации наследственного материала. Перед началом
- 65. Этапы спирализации хроматина.
- 66. Хромосома имеет вид буквы X, так как две хроматиды соединены между собой в районе центромеры. Центромера
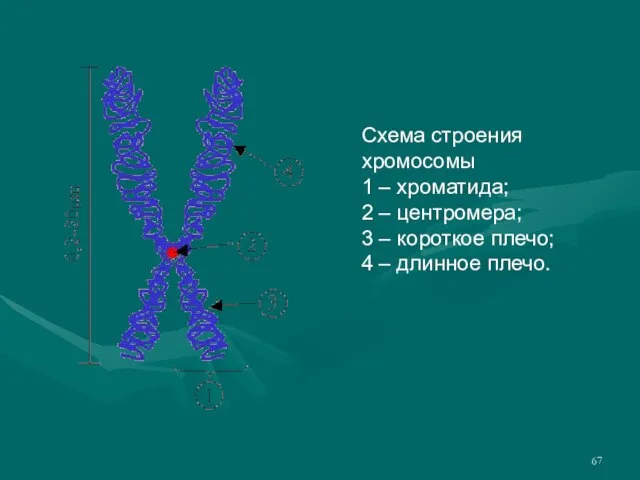
- 67. Схема строения хромосомы 1 – хроматида; 2 – центромера; 3 – короткое плечо; 4 – длинное
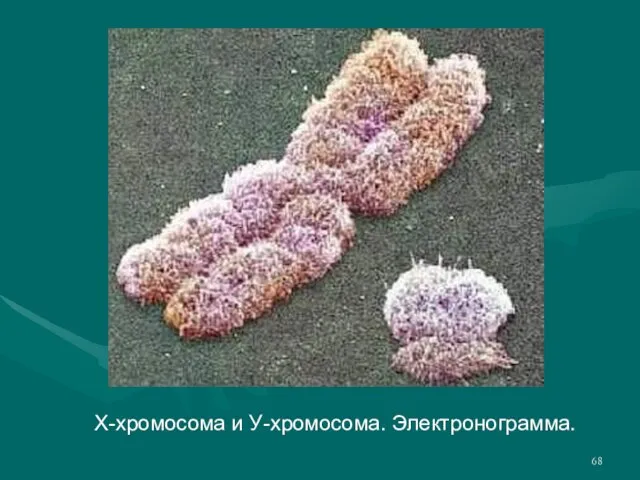
- 68. Х-хромосома и У-хромосома. Электронограмма.
- 69. По строению различают четыре типа хромосом: телоцентрические (палочковидные хромосомы с центромерой, расположенной на проксимальном конце); акроцентрические
- 70. Кариотип – совокупность признаков (число, размеры, форма) полного набора хромосом клеток данного биологического вида (видовой кариотип),
- 71. Нормальный кариотип (кариограмма) мужчины. Идиограмма нормального кариотипа мужчины.
- 72. Нормальный кариотип (кариограмма) женщины Идиограмма нормального кариотипа женщины.
- 73. Основы существующей унифицированной классификации хромосом были заложены в 1960 году в Денвере. В основу классификации положены
- 74. Группа D включает три акроцентрические хромосомы среднего размера: 13, 14 и 15. Группа Е также три
- 75. Гаплоидный набор хромосом мужчины. Хромосомы разделены на 7 групп по Денверской классификации (1960 год). Х и
- 76. Геномный уровень организации наследственного материала. Генотип – совокупность генов данного организма. Генотип, в отличие от понятий
- 77. Геном – совокупность всех генов организма. Термин «геном» был предложен Гансом Винклером в 1920 г. для
- 78. 3. Молекулярные основы наследственности. Реализация наследственной информации. К нуклеиновым кислотам относят дезоксирибонуклеиновую кислоту (ДНК) и рибонуклеиновую
- 79. В 1953 году Фрэнсис Крик и Джеймс Уотсон описали строение ДНК. Они обнаружили, что: - ДНК
- 80. Значительный вклад в изучение закономерностей строения ДНК внес Эрвин Чаргаф. Правила Чаргафа. - сумма пиримидиновых оснований
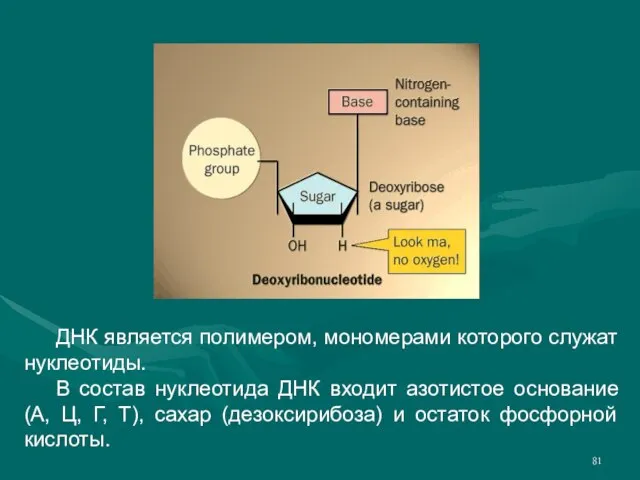
- 81. ДНК является полимером, мономерами которого служат нуклеотиды. В состав нуклеотида ДНК входит азотистое основание (А, Ц,
- 82. Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы и фосфатной группы. Эти связи называются фосфодиэфирными.
- 83. Асимметричные концы цепи ДНК называются 3' (три прим) и 5' (пять прим). Полярность цепи играет важную
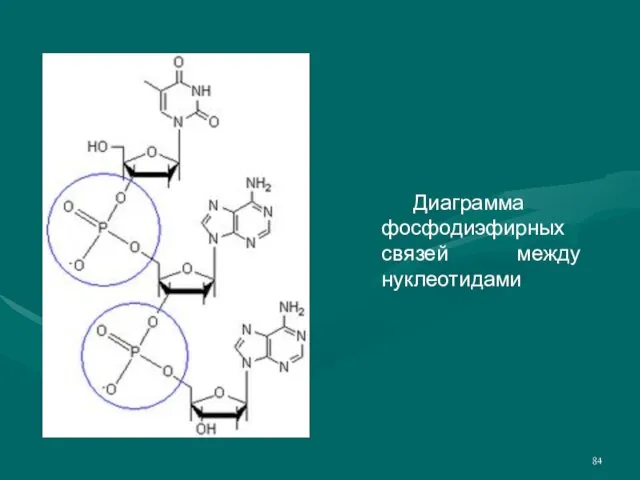
- 84. Диаграмма фосфодиэфирных связей между нуклеотидами
- 85. Структура молекулы ДНК получила название «двойной спирали». Азотистые основания одной из цепей соединены с азотистыми основаниями
- 86. В природе существует полиморфизм молекул ДНК. Основными конформациями ДНК являются следующие: В-форма: обычная регулярная правозакрученная двойная
- 87. Полиморфизм молекул ДНК.
- 88. Последовательность нуклеотидов ДНК позволяет «кодировать» информацию о различных типах РНК, таких как информационные, или матричные (м
- 89. Основное свойство ДНК – способность к редупликации. Это процесс биосинтеза ДНК, в результате которого из одной
- 90. К каждой нити по принципу комплементарности достраивается вторая нить. Это полуконсервативный способ репликации. В нем участвуют
- 91. Фрагменты Оказаки (Okazaki fragment) — это относительно короткие фрагменты ДНК (с РНК-праймером на 5' конце), которые
- 92. Репликация ДНК.
- 93. Схематическое изображение процесса репликации, цифрами отмечены: (1) запаздывающая нить, (2) лидирующая нить, (3) ДНК полимераза (Polα),
- 94. В процессе жизнедеятельности возможны повреждения ДКН под влиянием физических, химических, биологических факторов. Повреждения возникают вследствие ошибок
- 95. Такие изменения в ДНК приводят к изменчивости. Изменчивость - свойство живых организмов, заключающееся в изменении генов
- 96. Генные мутации происходят в результате замены, вставки или выпадения нуклеотидов или их блоком (размер до 5
- 97. Для устранения этих ошибок и поддержания стабильности цепи ДНК существуют механизмы, устраняющие эти ошибки – репарации.
- 98. Темновая репарация - свойство клеток ликвидировать повреждения в ДНК без участия видимого света. Механизм темновой репарации:
- 99. Пострепликативная репарация состоит в устранении ошибок репликации. Она включает следующие компоненты: - фермент, "узнающий" химически изменённые
- 100. Кроме репараций стабильность фенотипа обеспечивают антимутационные барьеры. Они не позволят уже произошедшей, но не устраненной мутации,
- 101. РНК является полимером, мономерами которого служат нуклеотиды. В состав нуклеотида РНК входит азотистое основание (А, Ц,
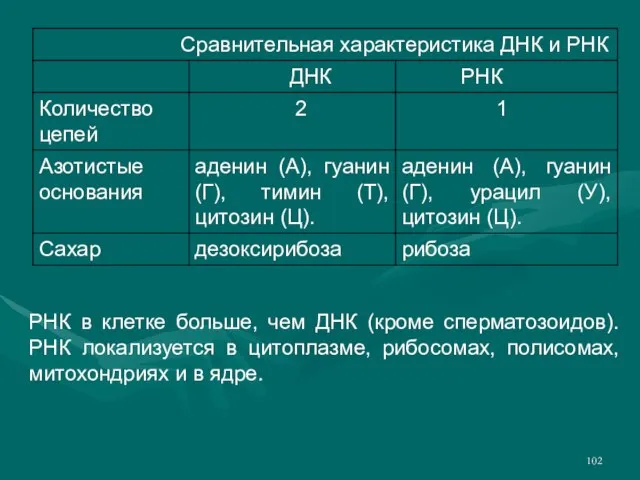
- 102. РНК в клетке больше, чем ДНК (кроме сперматозоидов). РНК локализуется в цитоплазме, рибосомах, полисомах, митохондриях и
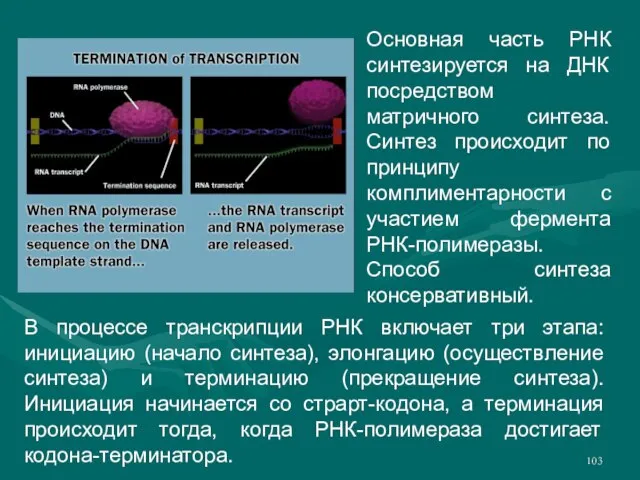
- 103. Основная часть РНК синтезируется на ДНК посредством матричного синтеза. Синтез происходит по принципу комплиментарности с участием
- 104. Выделяют три вида РНК – рибосомальная (р-РНК), транспортная (т-РНК), информационная (и-РНК). р-РНК составляет до 80% всей
- 105. т-РНК составляет до 15% всей РНК клетки. Содержит 75-90 нуклеотидов. т-РНК Характерна пространственная организация в виде
- 106. Ген - структурно-функциональная единица наследственной информации, представляющая собой участок ДНК, который через синтез белка или пула
- 107. Гену также присущи такие свойства как: - лабильность — способность многократно мутировать; - множественный аллелизм —
- 108. Функции генов близки к функциям ДНК: - хранение наследственной информации, - воспроизведение наследственной информации, - передача
- 109. Классификаций генов множество. Выделяют аллельные и неаллельные гены, доминантные и рецессивные гены, гены аутосом и половых
- 111. Генетический код - это последовательное расположение нуклеотидов ДНК, определяющее порядок расположения аминокислот в белке. Теория генетического
- 112. Непрерывность - между триплетами нет знаков препинания, информация считывается непрерывно. Неперекрываемость - один и тот же
- 113. В последнее время подвергаются сомнению некоторые свойства генетического кода: триплетность (третий нуклеотид триплета неравнозначен двум первым),
- 114. Таблица генетического кода. Секторный вариант записи генетического кода, внутренний круг — 1-е основание кодона (от 5'-конца).
- 115. Для обеспечения процессов онтогенеза необходима реализация генетической информации. Основным механизмом ее реализации является биосинтез белка. Биосинтез
- 116. Транскрипция - это процесс переписывания информации с ДНК на и-РНК. Происходит в ядре, по принципу комплементарности,
- 117. У эукариотов ДНК содержит экзоны (участки, несущие информацию и интроны (неинформационные участки). У эукариотот на базе
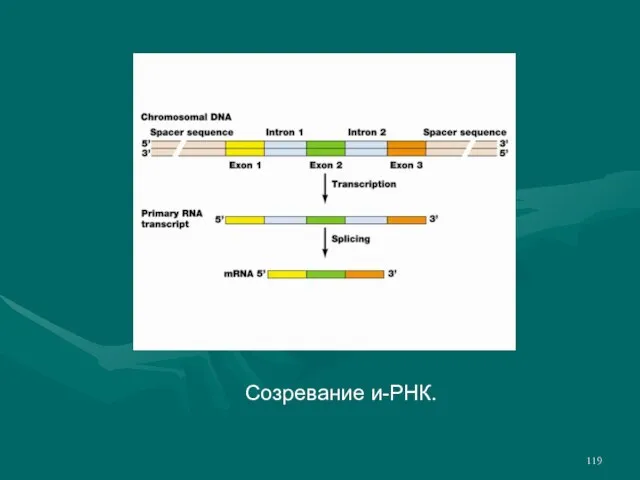
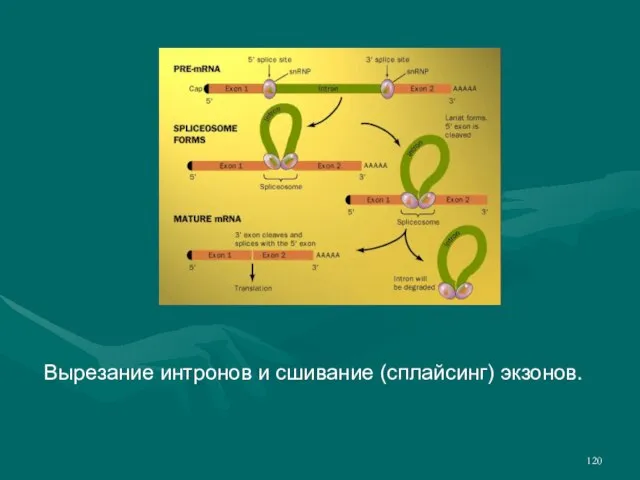
- 118. В процессе сплайсинга из пре-мРНК удаляются некодирующие белок последовательности (интроны). На 5' конец молекулы и-РНК добавляется
- 119. Созревание и-РНК.
- 120. Вырезание интронов и сшивание (сплайсинг) экзонов.
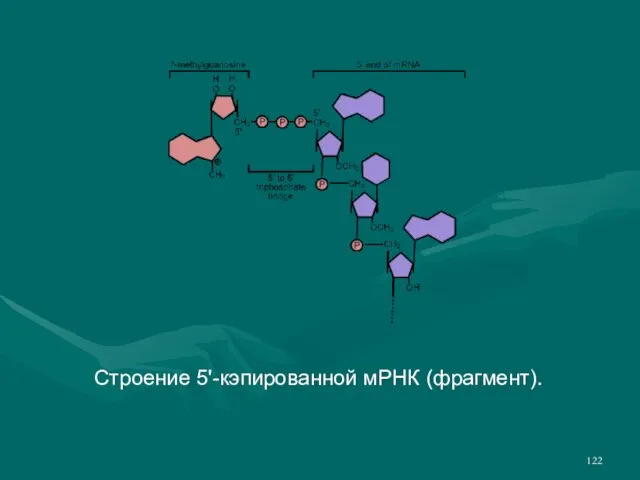
- 121. Зрелая и-РНК состоит из нескольких участков: 5'-кэп, 5'-нетранслируемая область, кодирующая (транслируемая) область, 3'-нетранслируемая область и 3'-полиадениновый
- 122. Строение 5'-кэпированной мРНК (фрагмент).
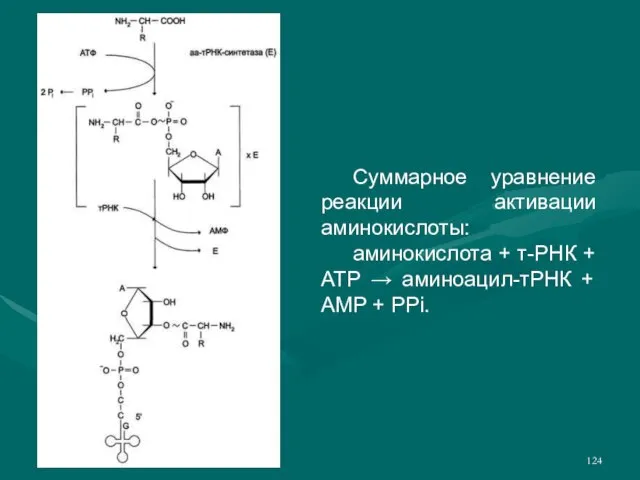
- 123. Рекогниция - процесс узнавания т-РНК аминокислоты, активация ее и транспорт к месту сборки полипептидной последовательность. Активация
- 124. Суммарное уравнение реакции активации аминокислоты: аминокислота + т-РНК + ATP → аминоацил-тРНК + AMP + PPi.
- 125. Трансляция - процесс построения полипептида на базе информации и-РНК. Происходит в рибосомах. Аминокислотная последовательность белка выстраивается
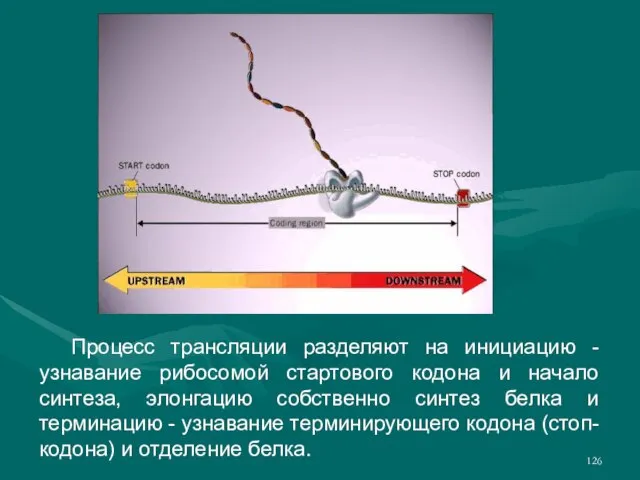
- 126. Процесс трансляции разделяют на инициацию - узнавание рибосомой стартового кодона и начало синтеза, элонгацию собственно синтез
- 127. Инициация трансляции начинается со стартового кодона AUG-кодона, кодирующего метионин. Происходит узнавание рибосомой этого кодона и привлечение
- 128. Терминация осуществляется, когда в А-сайте рибосомы оказывается один из стоп-кодонов - UAG, UAA, UGA. Из-за отсутствия
- 129. Готовая белковая молекула отщепляется от рибосомы и транспортируется в нужное место клетки. Для достижения своего активного
- 130. Считывание информации c ДНК носит название экспрессия генов. В каждый момент времени информация считывается только с
- 131. Оперон - участок структурных генов (цистронов), объединенных общей регуляторной зоной. Начинается и заканчивается оперон регуляторными областями
- 132. Примером оперона генома прокариот является лактозный оперон (lac-оперон) - полицистронный оперон бактерий, кодирующий гены метаболизма лактозы.
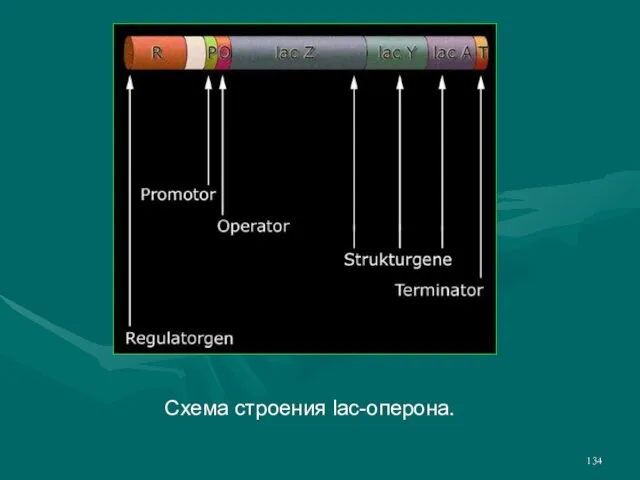
- 133. Лактозный оперон (lac-operon) состоит из трех структурных генов (гены lacZ, lacY и lacA), промотора, оператора и
- 134. Схема строения lac-оперона.
- 135. Промотор узнает, активирует и направляет РНК-полимеразу на оператор, оператор пропускает или не пропускает РНК-полимеразу на структурные
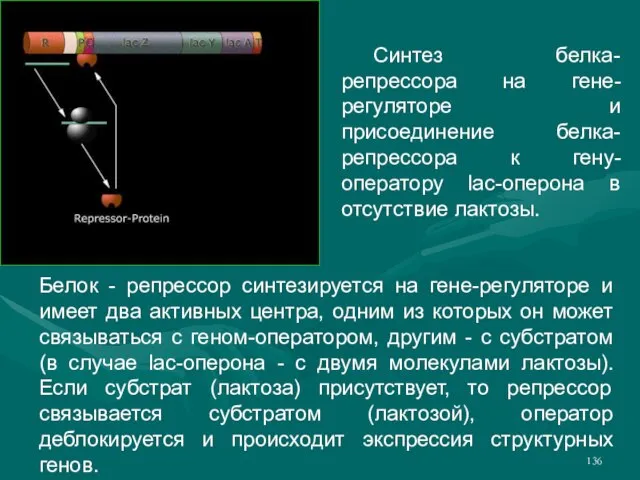
- 136. Белок - репрессор синтезируется на гене-регуляторе и имеет два активных центра, одним из которых он может
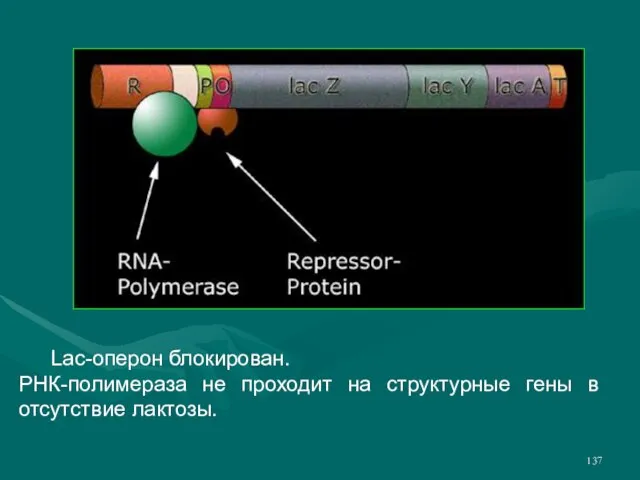
- 137. Lac-оперон блокирован. РНК-полимераза не проходит на структурные гены в отсутствие лактозы.
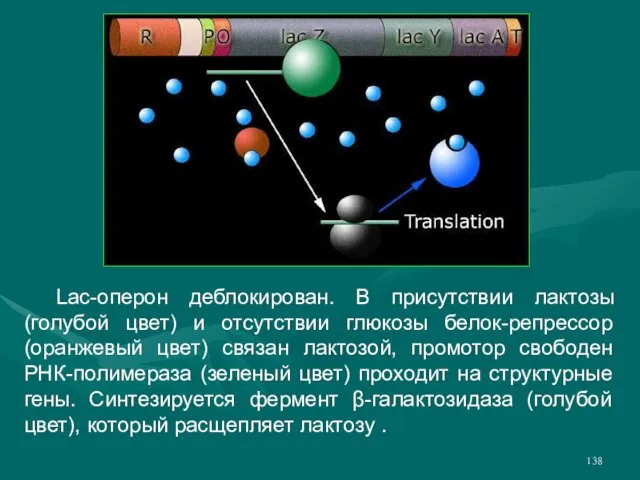
- 138. Lac-оперон деблокирован. В присутствии лактозы (голубой цвет) и отсутствии глюкозы белок-репрессор (оранжевый цвет) связан лактозой, промотор
- 139. В регуляции активности lac-оперона принимает участие специфический белок - белок, активирующий катаболизм (САР, catabolism activating protein),
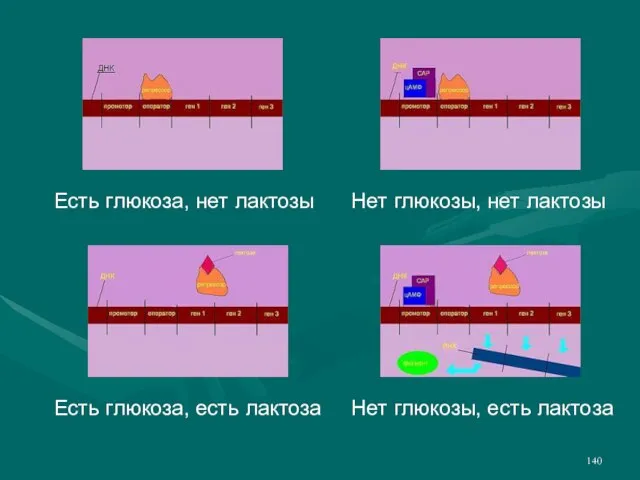
- 140. Есть глюкоза, нет лактозы Есть глюкоза, есть лактоза Нет глюкозы, нет лактозы Нет глюкозы, есть лактоза
- 141. У эукариот регуляция экспрессии генов происходит сложнее. РНК-полимеразы эукариот сами по себе не способны узнать промотор,
- 142. Усилители и репрессоры могут быть расположены далеко (на расстоянии 1000 нуклеотидных пар и более от старта
- 143. У эукариот регуляция осуществляется на этапе созревания про-и-РНК в зрелую и-РНК. У эукариот регуляция экспрессии генов
- 145. Скачать презентацию

Слайд 3Цитозоль
это структурированный коллоид, состоящий из сложной смеси растворенных в воде органических
Цитозоль
это структурированный коллоид, состоящий из сложной смеси растворенных в воде органических

Слайд 4Химический состав и свойства цитозоля:
Неорганические вещества: вода, соли, газы.
Органические вещества: белки, углеводы,
Химический состав и свойства цитозоля:
Неорганические вещества: вода, соли, газы.
Органические вещества: белки, углеводы,

Слайд 5Цитоскелет.
К элементам цитоскелета относят белковые фибриллярные структуры, расположенные в цитоплазме клетки:
Цитоскелет.
К элементам цитоскелета относят белковые фибриллярные структуры, расположенные в цитоплазме клетки:

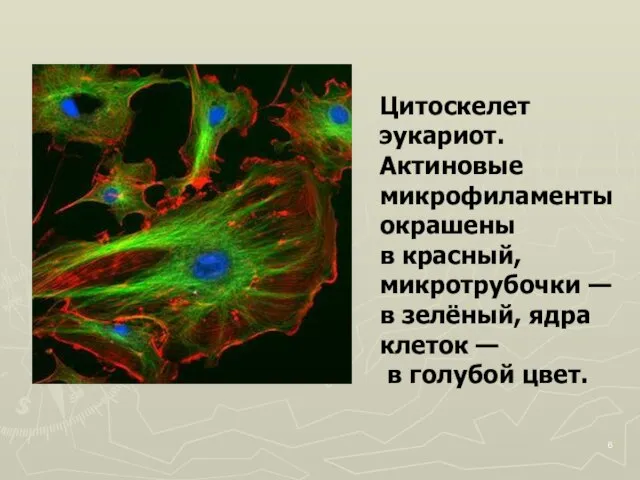
Слайд 6Цитоскелет эукариот. Актиновые микрофиламенты окрашены
в красный, микротрубочки — в зелёный, ядра
Цитоскелет эукариот. Актиновые микрофиламенты окрашены
в красный, микротрубочки — в зелёный, ядра
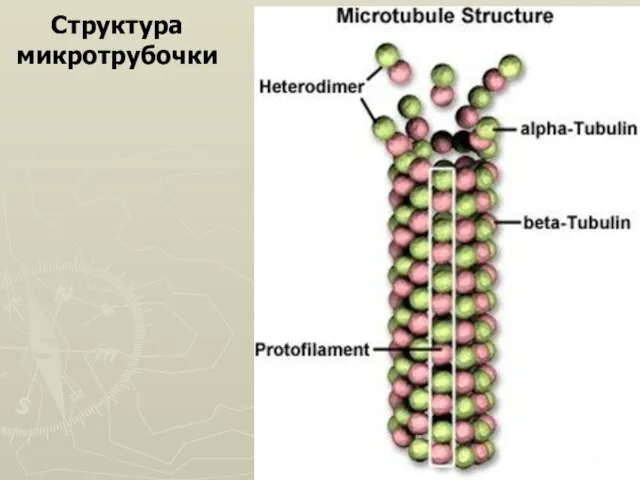
Слайд 7Структура микротрубочки
Структура микротрубочки
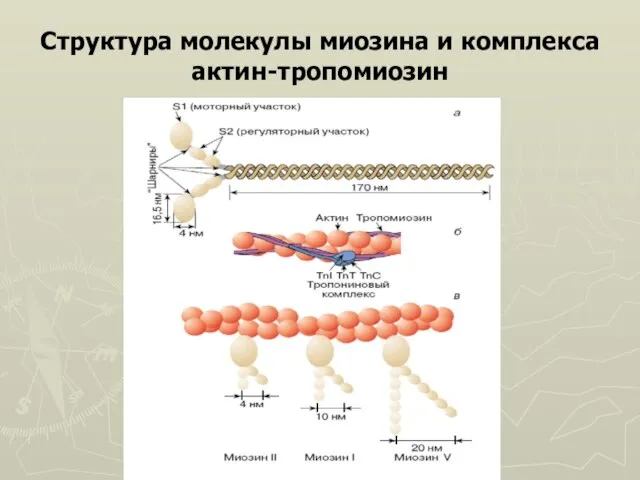
Слайд 8Структура молекулы миозина и комплекса
актин-тропомиозин
Структура молекулы миозина и комплекса
актин-тропомиозин
Слайд 9Функции цитоскелета:
1. Поддержка объема и формы клеток
2. Изменение формы клеток
3. Передвижение органелл
Функции цитоскелета:
1. Поддержка объема и формы клеток
2. Изменение формы клеток
3. Передвижение органелл

Слайд 10ВКЛЮЧЕНИЯ
В цитоплазме клеток также включения - непостоянные компоненты, выполняющие функцию запасания питательных
ВКЛЮЧЕНИЯ
В цитоплазме клеток также включения - непостоянные компоненты, выполняющие функцию запасания питательных

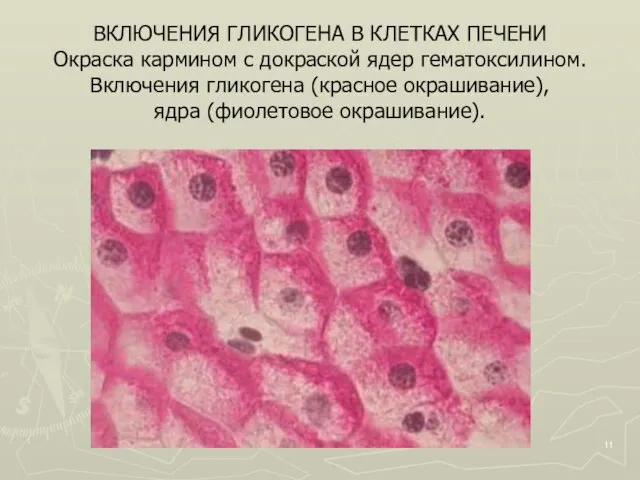
Слайд 11ВКЛЮЧЕНИЯ ГЛИКОГЕНА В КЛЕТКАХ ПЕЧЕНИ
Окраска кармином с докраской ядер гематоксилином.
Включения гликогена
ВКЛЮЧЕНИЯ ГЛИКОГЕНА В КЛЕТКАХ ПЕЧЕНИ Окраска кармином с докраской ядер гематоксилином. Включения гликогена
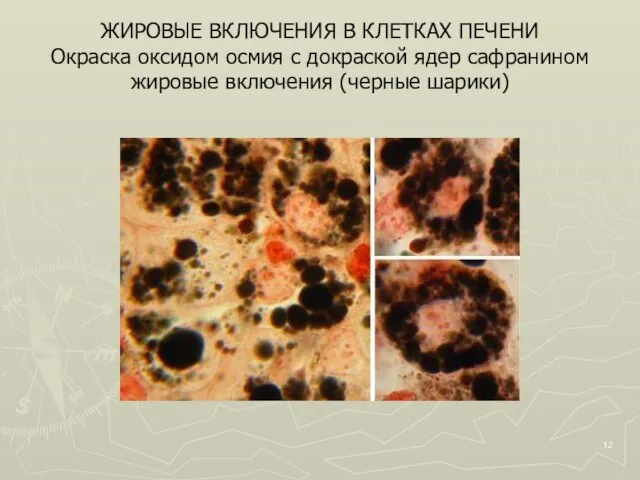
Слайд 12ЖИРОВЫЕ ВКЛЮЧЕНИЯ В КЛЕТКАХ ПЕЧЕНИ
Окраска оксидом осмия с докраской ядер сафранином
жировые
ЖИРОВЫЕ ВКЛЮЧЕНИЯ В КЛЕТКАХ ПЕЧЕНИ Окраска оксидом осмия с докраской ядер сафранином жировые
Слайд 13 Органеллы - постоянные компоненты цитоплазмы, имеющие определенную структуру и выполняющие определенные функции.
Классификация
Органеллы - постоянные компоненты цитоплазмы, имеющие определенную структуру и выполняющие определенные функции.
Классификация

Слайд 14Классификация органелл по выполняемой функции:
А) Органеллы общего назначения: митохондрии, лизосомы, пероксисомы, комплекс
Классификация органелл по выполняемой функции:
А) Органеллы общего назначения: митохондрии, лизосомы, пероксисомы, комплекс

Слайд 15Эндоплазматический ретикулум (эндоплазматическая сеть, ЭПР или ЭПС) - система переходящих друг в
Эндоплазматический ретикулум (эндоплазматическая сеть, ЭПР или ЭПС) - система переходящих друг в

Слайд 16ОДНОМЕМБРАННЫЕ ОРГАНЕЛЛЫ
Эндоплазматическая сеть
Агранулярная
Функции гладкой ЭПС:
1. Синтез фосфолипидов и углеводов;
2. Накопление и модификация
ОДНОМЕМБРАННЫЕ ОРГАНЕЛЛЫ
Эндоплазматическая сеть
Агранулярная
Функции гладкой ЭПС:
1. Синтез фосфолипидов и углеводов;
2. Накопление и модификация

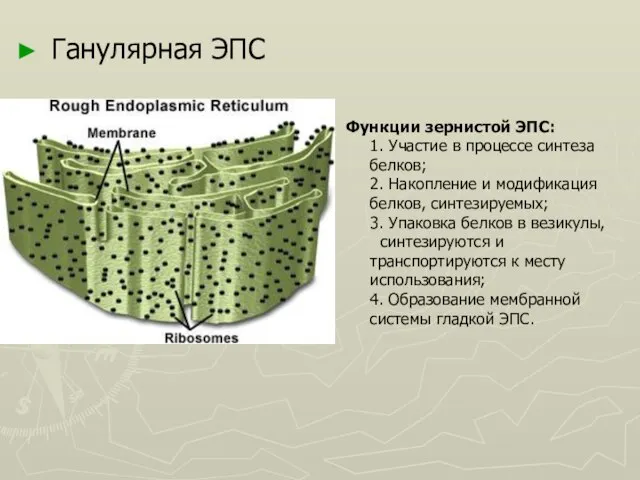
Слайд 17Ганулярная ЭПС
Функции зернистой ЭПС:
1. Участие в процессе синтеза белков;
2. Накопление и модификация
Ганулярная ЭПС
Функции зернистой ЭПС: 1. Участие в процессе синтеза белков; 2. Накопление и модификация
Слайд 18 Комплекс Гольжи (Аппарат Гольджи) - стопка плоских мембранных цистерн.
В цистернах созревают
Комплекс Гольжи (Аппарат Гольджи) - стопка плоских мембранных цистерн.
В цистернах созревают

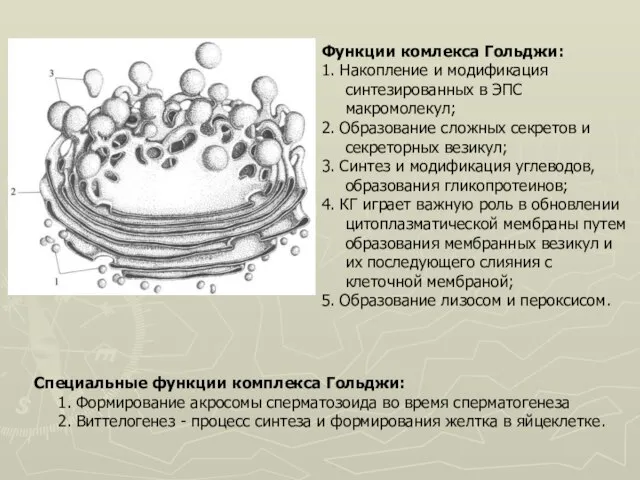
Слайд 19Функции комлекса Гольджи:
1. Накопление и модификация синтезированных в ЭПС макромолекул;
2. Образование сложных
Функции комлекса Гольджи:
1. Накопление и модификация синтезированных в ЭПС макромолекул;
2. Образование сложных
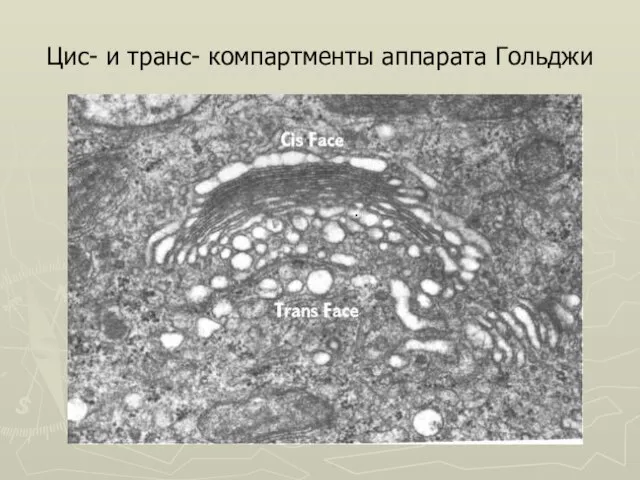
Слайд 20Цис- и транс- компартменты аппарата Гольджи
Цис- и транс- компартменты аппарата Гольджи
Слайд 21Лизосома небольшая органелла, ограниченная одной мембраной. В ней находятся литические ферменты, способные
Лизосома небольшая органелла, ограниченная одной мембраной. В ней находятся литические ферменты, способные

Слайд 22Функции лизосом:
1. Переваривание веществ, поступающих в клетку извне в процессе фагоцитоза.
2. Переваривание
Функции лизосом:
1. Переваривание веществ, поступающих в клетку извне в процессе фагоцитоза.
2. Переваривание

Слайд 23 Пероксисома – небольшая органелла, ограниченная одной мембраной, содержащая большое количество ферментов, катализирующих
Пероксисома – небольшая органелла, ограниченная одной мембраной, содержащая большое количество ферментов, катализирующих

Слайд 24Вакуоли (только в растительной клетке)
Они отделены от цитоплазмы одинарной мембраной - тонопластом.

Слайд 25Двумембранные органеллы
Митохондрии. Внутренний просвет митохондрий, называемый матриксом, отграничен от цитоплазмы двумя мембранами,
Двумембранные органеллы
Митохондрии. Внутренний просвет митохондрий, называемый матриксом, отграничен от цитоплазмы двумя мембранами,

Слайд 26Митохондрии.
Схема строения. Электронограмма.
Митохондрии.
Схема строения. Электронограмма.
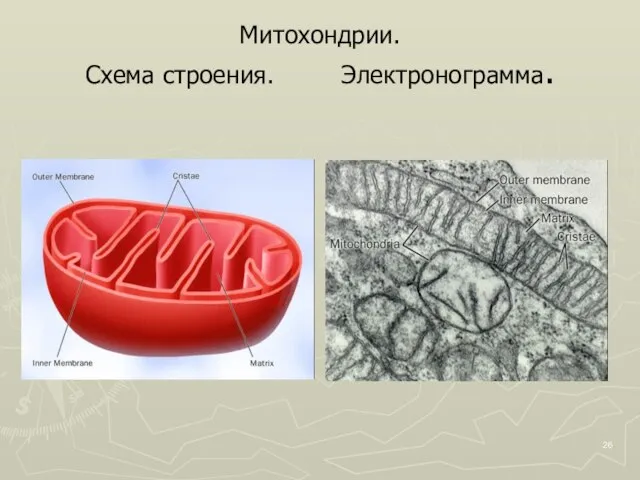
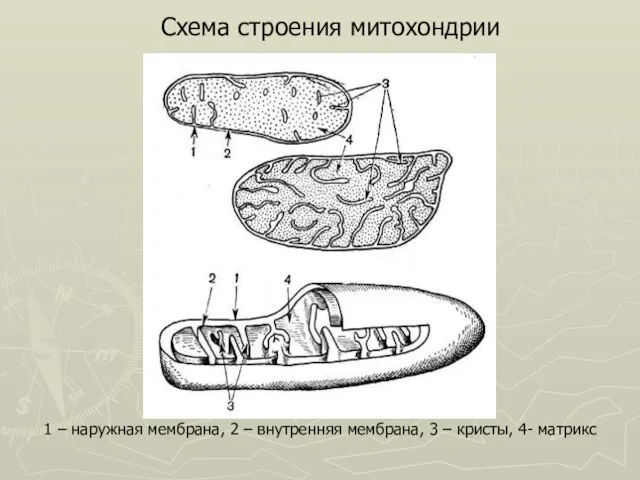
Слайд 271 – наружная мембрана, 2 – внутренняя мембрана, 3 – кристы, 4-
1 – наружная мембрана, 2 – внутренняя мембрана, 3 – кристы, 4-
Слайд 28 Пластиды - органеллы растений и некоторых фотосинтезирующих простейших. Покрыты двойной мембраной и
Пластиды - органеллы растений и некоторых фотосинтезирующих простейших. Покрыты двойной мембраной и

Слайд 29Пластиды : хлоропласты, хромопласты та лейкопласти.
Хлоропласты
Пластиды : хлоропласты, хромопласты та лейкопласти.
Хлоропласты

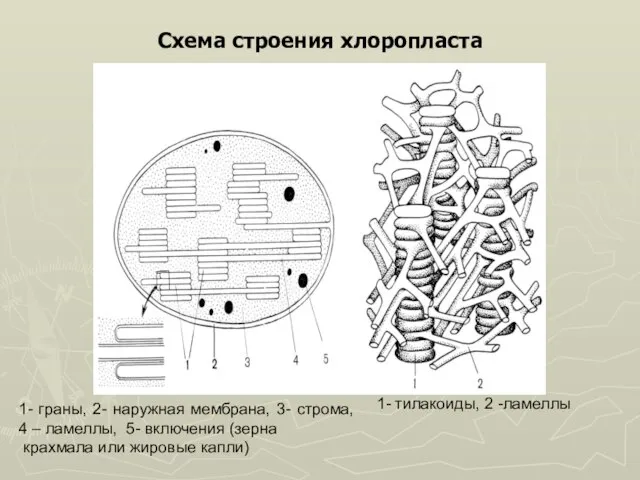
Слайд 30Схема строения хлоропласта
1- граны, 2- наружная мембрана, 3- строма, 4 – ламеллы,
Схема строения хлоропласта
1- граны, 2- наружная мембрана, 3- строма, 4 – ламеллы,
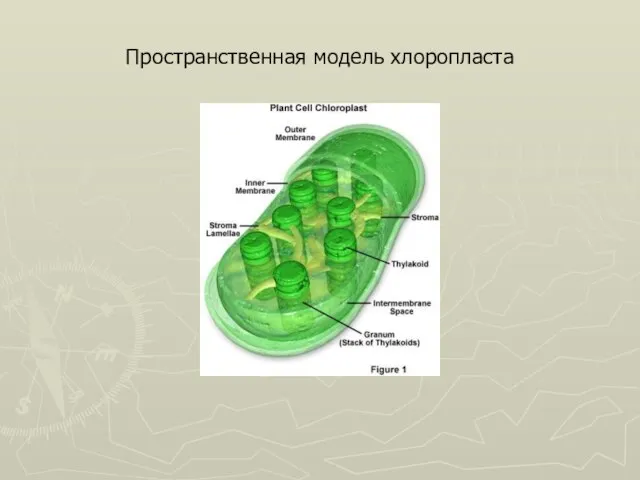
Слайд 31Пространственная модель хлоропласта
Пространственная модель хлоропласта
Слайд 32Рибосома - немембранная органелла сферической или эллипсоидной формы, состоящая из большой и
Рибосома - немембранная органелла сферической или эллипсоидной формы, состоящая из большой и

Слайд 33Схема строения рибосомы
В эукариотических клетках рибосомы располагаются на мембранах эндоплазматического ретикулума
Схема строения рибосомы
В эукариотических клетках рибосомы располагаются на мембранах эндоплазматического ретикулума
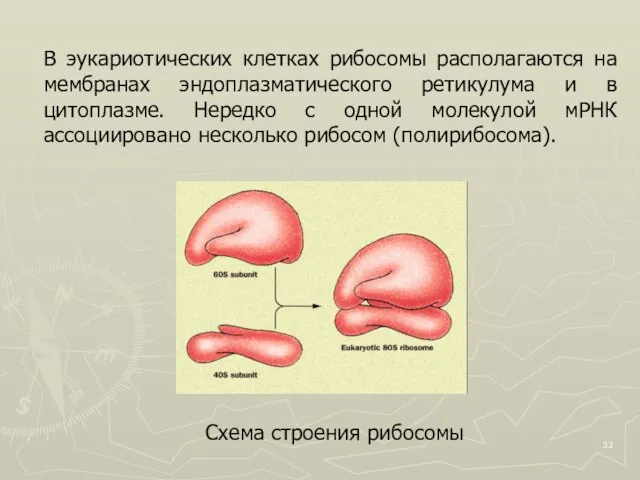
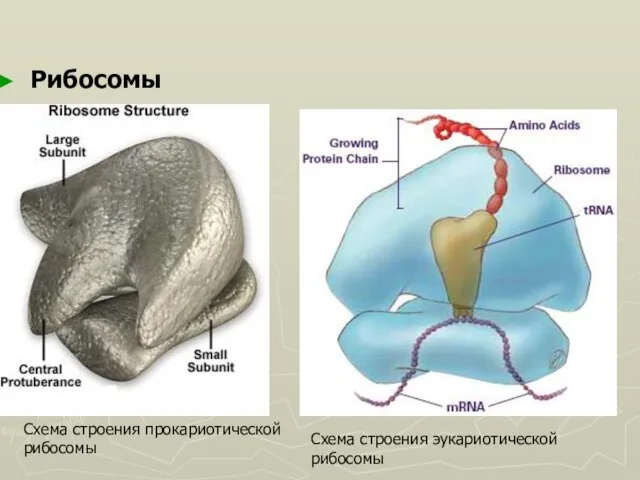
Слайд 34Рибосомы
Схема строения прокариотической
рибосомы
Схема строения эукариотической
рибосомы
Рибосомы
Схема строения прокариотической
рибосомы
Схема строения эукариотической
рибосомы
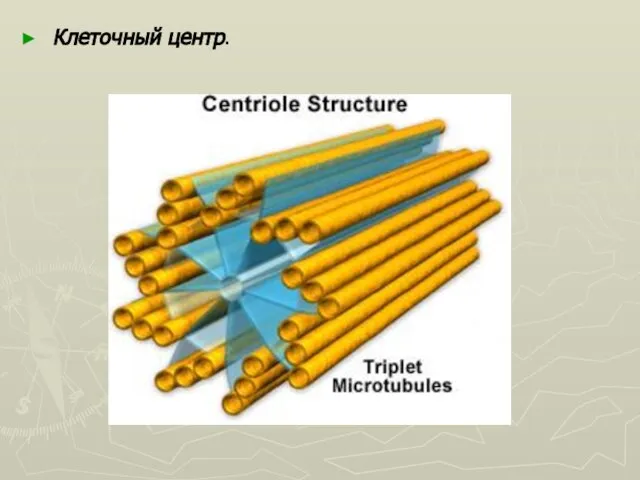
Слайд 35Клеточный центр представлен двумя центриолями ориентированными перпендикулярно друг к другу. Центриоли представляют
Клеточный центр представлен двумя центриолями ориентированными перпендикулярно друг к другу. Центриоли представляют

Слайд 36Клеточный центр.
Клеточный центр.
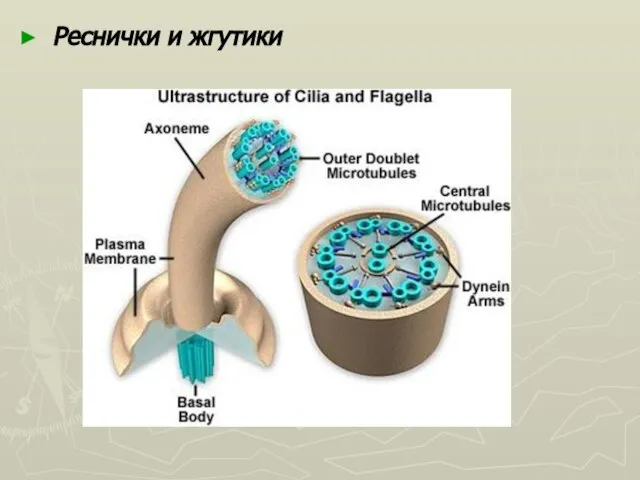
Слайд 37Базальные тельца лежат в цитоплазме в основе ресничек и жгутиков. Каждое кинетосома
Базальные тельца лежат в цитоплазме в основе ресничек и жгутиков. Каждое кинетосома

Слайд 38Реснички и жгутики
Реснички и жгутики

Слайд 40Клеточное ядро
Клеточное ядро

Слайд 41 Ядро – центральный информационный аппарат клетки. Структура интерфазного ядра. Хроматин: эухроматин,
Ядро – центральный информационный аппарат клетки. Структура интерфазного ядра. Хроматин: эухроматин,

Слайд 42Ядро - это один из структурных компонентов эукариотической клетки, содержащей генетическую информацию
Ядро - это один из структурных компонентов эукариотической клетки, содержащей генетическую информацию
Слайд 43История открытия
История открытия

Слайд 441) Гипотеза, известная как "синтропная модель", предполагает что ядро возникло в результате
1) Гипотеза, известная как "синтропная модель", предполагает что ядро возникло в результате
Слайд 45 .
Ядро и некоторые органеллы эукариотической клетки. Электронограмма.
.
Ядро и некоторые органеллы эукариотической клетки. Электронограмма.

Слайд 46Функции ядра
Сохраняет наследственную информацию и обеспечивает ее передачу от материнской клетки дочерним
Регулирует
Функции ядра
Сохраняет наследственную информацию и обеспечивает ее передачу от материнской клетки дочерним
Регулирует
Слайд 47Строение ядра
Ядрышко
Внешняя мембрана
Оболочка
Ядерные поры
Хроматин
Внутренняя мембрана
Кариоплазма
Строение ядра
Ядрышко
Внешняя мембрана
Оболочка
Ядерные поры
Хроматин
Внутренняя мембрана
Кариоплазма
Слайд 48 Ядро отделено от цитоплазмы ядерной оболочкой, образованной.
Ядерная оболочка (кариолемма) состоит из
Ядро отделено от цитоплазмы ядерной оболочкой, образованной.
Ядерная оболочка (кариолемма) состоит из

Слайд 49 Схема границы клеточного ядра. Показано перинуклеарное пространство, ограниченное двумя мембранами. Видна ядерная
Схема границы клеточного ядра. Показано перинуклеарное пространство, ограниченное двумя мембранами. Видна ядерная
Слайд 50 Пора имеет сложную структуру, организованную несколькими десятками специализированных белков – нуклеопоринов. Белковая
Пора имеет сложную структуру, организованную несколькими десятками специализированных белков – нуклеопоринов. Белковая
Слайд 51 Функции ядерной оболочки: разграничение содержимого ядра и цитоплазмы, транспорт веществ между ядром
Функции ядерной оболочки: разграничение содержимого ядра и цитоплазмы, транспорт веществ между ядром

Слайд 52Нуклеоїд прокаріотичних клітин
Спадкова інформація прокаріотів міститься в нуклеоїді. «Нуклеоїд» означає «подібний до
Нуклеоїд прокаріотичних клітин
Спадкова інформація прокаріотів міститься в нуклеоїді. «Нуклеоїд» означає «подібний до
Слайд 53 Между структурными компонентами ядра заключена жидкая фаза клеточного ядра, кариоплазма, в которой
Между структурными компонентами ядра заключена жидкая фаза клеточного ядра, кариоплазма, в которой

Слайд 54 Ядрышко находится внутри ядра, не имеет собственной мембранной оболочки, хорошо различимо под
Ядрышко находится внутри ядра, не имеет собственной мембранной оболочки, хорошо различимо под

Слайд 55 В ядрышке выделяют внутренний фибриллярный компонент, где происходит синтез рРНК.
Снаружи расположен
В ядрышке выделяют внутренний фибриллярный компонент, где происходит синтез рРНК.
Снаружи расположен
Слайд 56 Хроматин. В интерфазном ядре выявляются зоны плотного вещества, которое хорошо воспринимает красители.
Хроматин. В интерфазном ядре выявляются зоны плотного вещества, которое хорошо воспринимает красители.

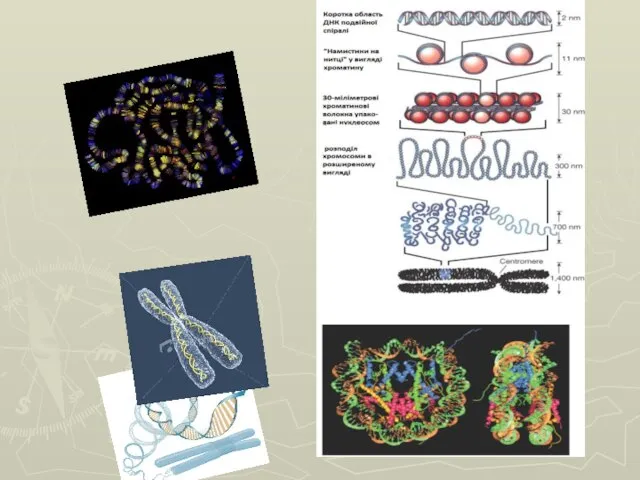
Слайд 58 Функции гистоновых белков:
- создание специфической укладки ДНК
- защита ДНК
- препятствование транскрипции
Функции негистоновых
Функции гистоновых белков:
- создание специфической укладки ДНК
- защита ДНК
- препятствование транскрипции
Функции негистоновых

Слайд 59 Гистоны Н2А, Н2В, Н3, Н4 входят в состав нуклеосом, Н1 находится между
Гистоны Н2А, Н2В, Н3, Н4 входят в состав нуклеосом, Н1 находится между

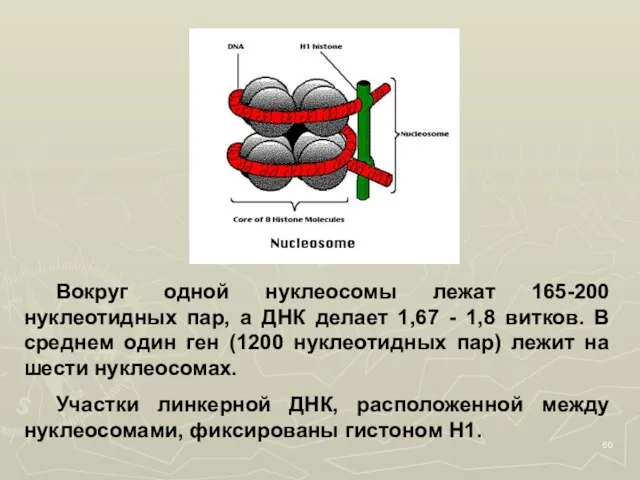
Слайд 60 Вокруг одной нуклеосомы лежат 165-200 нуклеотидных пар, а ДНК делает 1,67 -
Вокруг одной нуклеосомы лежат 165-200 нуклеотидных пар, а ДНК делает 1,67 -
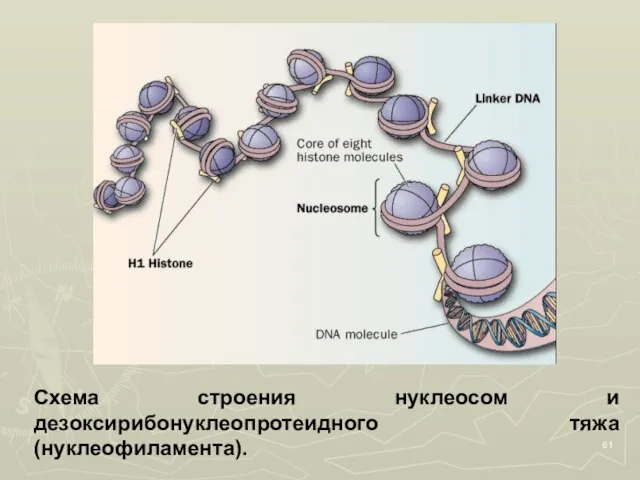
Слайд 61Схема строения нуклеосом и дезоксирибонуклеопротеидного тяжа (нуклеофиламента).
Схема строения нуклеосом и дезоксирибонуклеопротеидного тяжа (нуклеофиламента).
Слайд 62 В разные моменты существования клетки хроматин имеет разную пространственную организацию.
Он способен
В разные моменты существования клетки хроматин имеет разную пространственную организацию.
Он способен

Слайд 63 Эухроматин функционально активен. С него происходит считывание информации (транскрипция).
Гетерохроматин функционально не активен.
Эухроматин функционально активен. С него происходит считывание информации (транскрипция).
Гетерохроматин функционально не активен.

Слайд 64 2. Хромосомный, геномный и генный уровни организации наследственного материала.
Хромосомный уровень организации
2. Хромосомный, геномный и генный уровни организации наследственного материала.
Хромосомный уровень организации

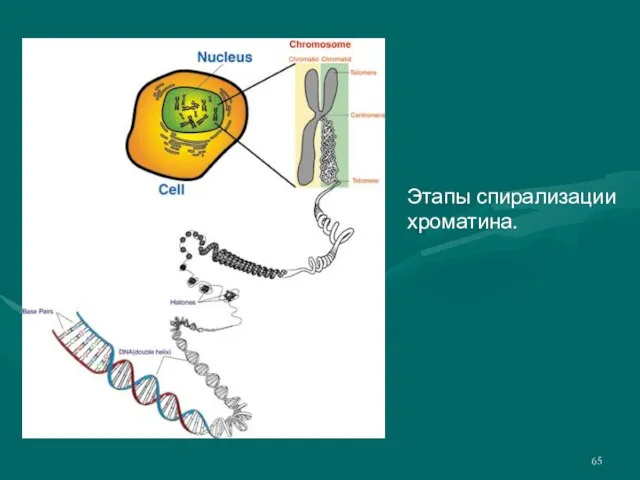
Слайд 65Этапы спирализации хроматина.
Этапы спирализации хроматина.
Слайд 66 Хромосома имеет вид буквы X, так как две хроматиды соединены между собой
Хромосома имеет вид буквы X, так как две хроматиды соединены между собой

Слайд 67Схема строения хромосомы
1 – хроматида;
2 – центромера;
3 – короткое
Схема строения хромосомы
1 – хроматида;
2 – центромера;
3 – короткое
Слайд 68Х-хромосома и У-хромосома. Электронограмма.
Х-хромосома и У-хромосома. Электронограмма.
Слайд 69 По строению различают четыре типа хромосом:
телоцентрические (палочковидные хромосомы с центромерой, расположенной
По строению различают четыре типа хромосом:
телоцентрические (палочковидные хромосомы с центромерой, расположенной

Слайд 70 Кариотип – совокупность признаков (число, размеры, форма) полного набора хромосом клеток данного
Кариотип – совокупность признаков (число, размеры, форма) полного набора хромосом клеток данного

Слайд 71Нормальный кариотип (кариограмма) мужчины.
Идиограмма нормального кариотипа мужчины.
Нормальный кариотип (кариограмма) мужчины.
Идиограмма нормального кариотипа мужчины.

Слайд 72Нормальный кариотип (кариограмма) женщины
Идиограмма нормального кариотипа женщины.
Нормальный кариотип (кариограмма) женщины
Идиограмма нормального кариотипа женщины.

Слайд 73 Основы существующей унифицированной классификации хромосом были заложены в 1960 году в Денвере.
Основы существующей унифицированной классификации хромосом были заложены в 1960 году в Денвере.

Слайд 74 Группа D включает три акроцентрические хромосомы среднего размера: 13, 14 и 15.
Группа
Группа D включает три акроцентрические хромосомы среднего размера: 13, 14 и 15.
Группа

Слайд 75 Гаплоидный набор хромосом мужчины.
Хромосомы разделены на 7 групп по Денверской классификации (1960
Гаплоидный набор хромосом мужчины.
Хромосомы разделены на 7 групп по Денверской классификации (1960

Слайд 76Геномный уровень организации наследственного материала.
Генотип – совокупность генов данного организма.
Генотип, в отличие
Геномный уровень организации наследственного материала.
Генотип – совокупность генов данного организма.
Генотип, в отличие

Слайд 77 Геном – совокупность всех генов организма.
Термин «геном» был предложен Гансом Винклером в
Геном – совокупность всех генов организма.
Термин «геном» был предложен Гансом Винклером в

Слайд 78 3. Молекулярные основы наследственности. Реализация наследственной информации.
К нуклеиновым кислотам относят дезоксирибонуклеиновую кислоту
3. Молекулярные основы наследственности. Реализация наследственной информации.
К нуклеиновым кислотам относят дезоксирибонуклеиновую кислоту

Слайд 79 В 1953 году Фрэнсис Крик и Джеймс Уотсон описали строение ДНК. Они
В 1953 году Фрэнсис Крик и Джеймс Уотсон описали строение ДНК. Они

Слайд 80 Значительный вклад в изучение закономерностей строения ДНК внес Эрвин Чаргаф.
Правила Чаргафа.
- сумма
Значительный вклад в изучение закономерностей строения ДНК внес Эрвин Чаргаф.
Правила Чаргафа.
- сумма

Слайд 81 ДНК является полимером, мономерами которого служат нуклеотиды.
В состав нуклеотида ДНК входит азотистое
ДНК является полимером, мономерами которого служат нуклеотиды.
В состав нуклеотида ДНК входит азотистое
Слайд 82 Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы и фосфатной группы.
Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы и фосфатной группы.

Слайд 83 Асимметричные концы цепи ДНК называются 3' (три прим) и 5' (пять прим).
Асимметричные концы цепи ДНК называются 3' (три прим) и 5' (пять прим).

Слайд 84 Диаграмма фосфодиэфирных связей между нуклеотидами
Диаграмма фосфодиэфирных связей между нуклеотидами
Слайд 85 Структура молекулы ДНК получила название «двойной спирали». Азотистые основания одной из цепей
Структура молекулы ДНК получила название «двойной спирали». Азотистые основания одной из цепей

Слайд 86 В природе существует полиморфизм молекул ДНК. Основными конформациями ДНК являются следующие:
В-форма: обычная
В природе существует полиморфизм молекул ДНК. Основными конформациями ДНК являются следующие:
В-форма: обычная

Слайд 87Полиморфизм молекул ДНК.
Полиморфизм молекул ДНК.

Слайд 88 Последовательность нуклеотидов ДНК позволяет «кодировать» информацию о различных типах РНК, таких как
Последовательность нуклеотидов ДНК позволяет «кодировать» информацию о различных типах РНК, таких как

Слайд 89 Основное свойство ДНК – способность к редупликации.
Это процесс биосинтеза ДНК, в
Основное свойство ДНК – способность к редупликации.
Это процесс биосинтеза ДНК, в

Слайд 90 К каждой нити по принципу комплементарности достраивается вторая нить. Это полуконсервативный способ
К каждой нити по принципу комплементарности достраивается вторая нить. Это полуконсервативный способ

Слайд 91 Фрагменты Оказаки (Okazaki fragment) — это относительно короткие фрагменты ДНК (с РНК-праймером
Фрагменты Оказаки (Okazaki fragment) — это относительно короткие фрагменты ДНК (с РНК-праймером

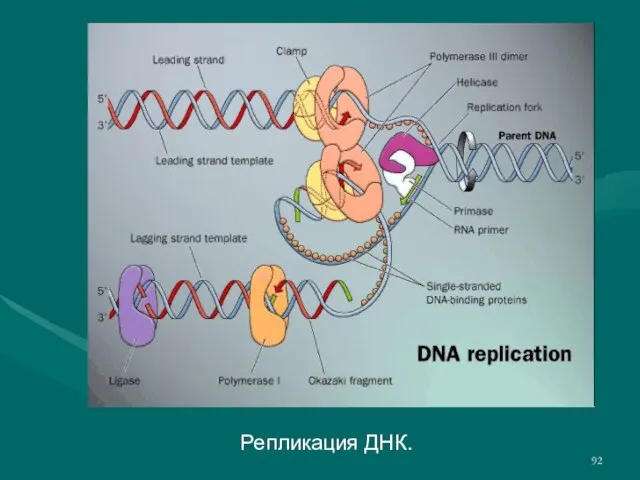
Слайд 92Репликация ДНК.
Репликация ДНК.
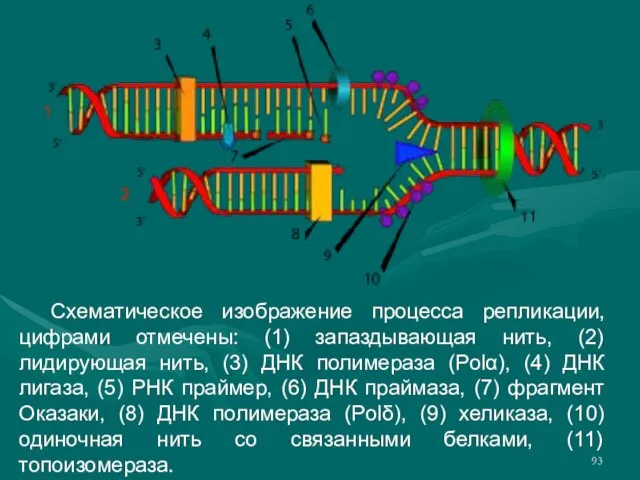
Слайд 93 Схематическое изображение процесса репликации, цифрами отмечены: (1) запаздывающая нить, (2) лидирующая нить,
Схематическое изображение процесса репликации, цифрами отмечены: (1) запаздывающая нить, (2) лидирующая нить,
Слайд 94 В процессе жизнедеятельности возможны повреждения ДКН под влиянием физических, химических, биологических факторов.
В процессе жизнедеятельности возможны повреждения ДКН под влиянием физических, химических, биологических факторов.

Слайд 95 Такие изменения в ДНК приводят к изменчивости. Изменчивость - свойство живых организмов,
Такие изменения в ДНК приводят к изменчивости. Изменчивость - свойство живых организмов,

Слайд 96 Генные мутации происходят в результате замены, вставки или выпадения нуклеотидов или их
Генные мутации происходят в результате замены, вставки или выпадения нуклеотидов или их

Слайд 97 Для устранения этих ошибок и поддержания стабильности цепи ДНК существуют механизмы, устраняющие
Для устранения этих ошибок и поддержания стабильности цепи ДНК существуют механизмы, устраняющие

Слайд 98 Темновая репарация - свойство клеток ликвидировать повреждения в ДНК без участия видимого
Темновая репарация - свойство клеток ликвидировать повреждения в ДНК без участия видимого

Слайд 99 Пострепликативная репарация состоит в устранении ошибок репликации.
Она включает следующие компоненты:
- фермент,
Пострепликативная репарация состоит в устранении ошибок репликации.
Она включает следующие компоненты:
- фермент,

Слайд 100 Кроме репараций стабильность фенотипа обеспечивают антимутационные барьеры.
Они не позволят уже произошедшей,
Кроме репараций стабильность фенотипа обеспечивают антимутационные барьеры.
Они не позволят уже произошедшей,

Слайд 101 РНК является полимером, мономерами которого служат нуклеотиды.
В состав нуклеотида РНК входит азотистое
РНК является полимером, мономерами которого служат нуклеотиды.
В состав нуклеотида РНК входит азотистое

Слайд 102РНК в клетке больше, чем ДНК (кроме сперматозоидов). РНК локализуется в цитоплазме,
РНК в клетке больше, чем ДНК (кроме сперматозоидов). РНК локализуется в цитоплазме,
Слайд 103Основная часть РНК синтезируется на ДНК посредством матричного синтеза. Синтез происходит по
Основная часть РНК синтезируется на ДНК посредством матричного синтеза. Синтез происходит по
Слайд 104 Выделяют три вида РНК – рибосомальная (р-РНК), транспортная (т-РНК), информационная (и-РНК).
р-РНК составляет
Выделяют три вида РНК – рибосомальная (р-РНК), транспортная (т-РНК), информационная (и-РНК).
р-РНК составляет

Слайд 105 т-РНК составляет до 15% всей РНК клетки. Содержит 75-90 нуклеотидов.
т-РНК Характерна
т-РНК составляет до 15% всей РНК клетки. Содержит 75-90 нуклеотидов.
т-РНК Характерна

Слайд 106 Ген - структурно-функциональная единица наследственной информации, представляющая собой участок ДНК, который через
Ген - структурно-функциональная единица наследственной информации, представляющая собой участок ДНК, который через

Слайд 107 Гену также присущи такие свойства как:
- лабильность — способность многократно мутировать;
- множественный
Гену также присущи такие свойства как:
- лабильность — способность многократно мутировать;
- множественный

Слайд 108 Функции генов близки к функциям ДНК:
- хранение наследственной информации,
- воспроизведение наследственной
Функции генов близки к функциям ДНК:
- хранение наследственной информации,
- воспроизведение наследственной

Слайд 109 Классификаций генов множество. Выделяют аллельные и неаллельные гены, доминантные и рецессивные гены,
Классификаций генов множество. Выделяют аллельные и неаллельные гены, доминантные и рецессивные гены,


Слайд 111
Генетический код - это последовательное расположение нуклеотидов ДНК, определяющее порядок расположения аминокислот
Генетический код - это последовательное расположение нуклеотидов ДНК, определяющее порядок расположения аминокислот

Слайд 112 Непрерывность - между триплетами нет знаков препинания, информация считывается непрерывно.
Неперекрываемость - один
Непрерывность - между триплетами нет знаков препинания, информация считывается непрерывно.
Неперекрываемость - один

Слайд 113В последнее время подвергаются сомнению некоторые свойства генетического кода:
триплетность (третий нуклеотид
В последнее время подвергаются сомнению некоторые свойства генетического кода:
триплетность (третий нуклеотид

Слайд 114Таблица генетического кода.
Секторный вариант записи генетического кода, внутренний круг — 1-е
Таблица генетического кода.
Секторный вариант записи генетического кода, внутренний круг — 1-е

Слайд 115 Для обеспечения процессов онтогенеза необходима реализация генетической информации.
Основным механизмом ее реализации
Для обеспечения процессов онтогенеза необходима реализация генетической информации.
Основным механизмом ее реализации

Слайд 116 Транскрипция - это процесс переписывания информации с ДНК на и-РНК.
Происходит в
Транскрипция - это процесс переписывания информации с ДНК на и-РНК.
Происходит в

Слайд 117 У эукариотов ДНК содержит экзоны (участки, несущие информацию и интроны (неинформационные участки).
У
У эукариотов ДНК содержит экзоны (участки, несущие информацию и интроны (неинформационные участки).
У

Слайд 118 В процессе сплайсинга из пре-мРНК удаляются некодирующие белок последовательности (интроны).
На 5'
В процессе сплайсинга из пре-мРНК удаляются некодирующие белок последовательности (интроны).
На 5'

Слайд 119Созревание и-РНК.
Созревание и-РНК.
Слайд 120Вырезание интронов и сшивание (сплайсинг) экзонов.
Вырезание интронов и сшивание (сплайсинг) экзонов.
Слайд 121Зрелая и-РНК состоит из нескольких участков: 5'-кэп, 5'-нетранслируемая область, кодирующая (транслируемая) область,
Зрелая и-РНК состоит из нескольких участков: 5'-кэп, 5'-нетранслируемая область, кодирующая (транслируемая) область,

Слайд 122Строение 5'-кэпированной мРНК (фрагмент).
Строение 5'-кэпированной мРНК (фрагмент).
Слайд 123 Рекогниция - процесс узнавания т-РНК аминокислоты, активация ее и транспорт к месту
Рекогниция - процесс узнавания т-РНК аминокислоты, активация ее и транспорт к месту

Слайд 124 Суммарное уравнение реакции активации аминокислоты:
аминокислота + т-РНК + ATP → аминоацил-тРНК +
Суммарное уравнение реакции активации аминокислоты:
аминокислота + т-РНК + ATP → аминоацил-тРНК +
Слайд 125 Трансляция - процесс построения полипептида на базе информации и-РНК. Происходит в рибосомах.
Аминокислотная
Трансляция - процесс построения полипептида на базе информации и-РНК. Происходит в рибосомах.
Аминокислотная

Слайд 126 Процесс трансляции разделяют на инициацию - узнавание рибосомой стартового кодона и начало
Процесс трансляции разделяют на инициацию - узнавание рибосомой стартового кодона и начало
Слайд 127Инициация трансляции начинается со стартового кодона AUG-кодона, кодирующего метионин. Происходит узнавание рибосомой
Инициация трансляции начинается со стартового кодона AUG-кодона, кодирующего метионин. Происходит узнавание рибосомой

Слайд 128Терминация осуществляется, когда в А-сайте рибосомы оказывается один из стоп-кодонов - UAG,
Терминация осуществляется, когда в А-сайте рибосомы оказывается один из стоп-кодонов - UAG,

Слайд 129Готовая белковая молекула отщепляется от рибосомы и транспортируется в нужное место клетки.
Готовая белковая молекула отщепляется от рибосомы и транспортируется в нужное место клетки.

Слайд 130 Считывание информации c ДНК носит название экспрессия генов.
В каждый момент времени информация
Считывание информации c ДНК носит название экспрессия генов.
В каждый момент времени информация

Слайд 131 Оперон - участок структурных генов (цистронов), объединенных общей регуляторной зоной.
Начинается и заканчивается
Оперон - участок структурных генов (цистронов), объединенных общей регуляторной зоной.
Начинается и заканчивается

Слайд 132 Примером оперона генома прокариот является лактозный оперон (lac-оперон) - полицистронный оперон бактерий,
Примером оперона генома прокариот является лактозный оперон (lac-оперон) - полицистронный оперон бактерий,

Слайд 133 Лактозный оперон (lac-operon) состоит из трех структурных генов (гены lacZ, lacY и
Лактозный оперон (lac-operon) состоит из трех структурных генов (гены lacZ, lacY и

Слайд 134Схема строения lac-оперона.
Схема строения lac-оперона.
Слайд 135 Промотор узнает, активирует и направляет РНК-полимеразу на оператор, оператор пропускает или не
Промотор узнает, активирует и направляет РНК-полимеразу на оператор, оператор пропускает или не

Слайд 136Белок - репрессор синтезируется на гене-регуляторе и имеет два активных центра, одним
Белок - репрессор синтезируется на гене-регуляторе и имеет два активных центра, одним
Слайд 137 Lac-оперон блокирован.
РНК-полимераза не проходит на структурные гены в отсутствие лактозы.
Lac-оперон блокирован.
РНК-полимераза не проходит на структурные гены в отсутствие лактозы.
Слайд 138 Lac-оперон деблокирован. В присутствии лактозы (голубой цвет) и отсутствии глюкозы белок-репрессор (оранжевый
Lac-оперон деблокирован. В присутствии лактозы (голубой цвет) и отсутствии глюкозы белок-репрессор (оранжевый
Слайд 139В регуляции активности lac-оперона принимает участие специфический белок - белок, активирующий катаболизм
В регуляции активности lac-оперона принимает участие специфический белок - белок, активирующий катаболизм

Слайд 140Есть глюкоза, нет лактозы
Есть глюкоза, есть лактоза
Нет глюкозы, нет лактозы
Есть глюкоза, нет лактозы
Есть глюкоза, есть лактоза
Нет глюкозы, нет лактозы
Слайд 141 У эукариот регуляция экспрессии генов происходит сложнее.
РНК-полимеразы эукариот сами по себе не
У эукариот регуляция экспрессии генов происходит сложнее.
РНК-полимеразы эукариот сами по себе не

Слайд 142 Усилители и репрессоры могут быть расположены далеко (на расстоянии 1000 нуклеотидных пар
Усилители и репрессоры могут быть расположены далеко (на расстоянии 1000 нуклеотидных пар

Слайд 143 У эукариот регуляция осуществляется на этапе созревания про-и-РНК в зрелую и-РНК.
У эукариот
У эукариот регуляция осуществляется на этапе созревания про-и-РНК в зрелую и-РНК.
У эукариот

 Первобытное искусство. Наскальные рисунки и скульптура
Первобытное искусство. Наскальные рисунки и скульптура МОУ «Ивано-Лисичанская СОШ»Лагерь «Мечта»
МОУ «Ивано-Лисичанская СОШ»Лагерь «Мечта» Изображать можно линией
Изображать можно линией Тип Моллюски (1)
Тип Моллюски (1) День России
День России Администрирование информационных систем
Администрирование информационных систем Кого мы воспитаем или какими будут наши дети?
Кого мы воспитаем или какими будут наши дети? ИСТОРИЧЕСКАЯ СПРАВКА о школьной форме
ИСТОРИЧЕСКАЯ СПРАВКА о школьной форме Береги зрение!
Береги зрение! МАРКЕТИНГОВЫЕ ИССЛЕДОВАНИЯ ДЛЯ БАНКОВСКОГО СЕКТОРА ВОЗМОЖНОСТИ И РЕШАЕМЫЕ ЗАДАЧИ
МАРКЕТИНГОВЫЕ ИССЛЕДОВАНИЯ ДЛЯ БАНКОВСКОГО СЕКТОРА ВОЗМОЖНОСТИ И РЕШАЕМЫЕ ЗАДАЧИ Эффект проливного дождя
Эффект проливного дождя Русские хороводы
Русские хороводы ОСОБЕННОСТИ ПЕДАГОГИЧЕСКИХ ИССЛЕДОВАНИЙ
ОСОБЕННОСТИ ПЕДАГОГИЧЕСКИХ ИССЛЕДОВАНИЙ Современные машиностроительные технологии
Современные машиностроительные технологии Государственный Фонд развития промышленности Рязанской области (ГФРП РО)
Государственный Фонд развития промышленности Рязанской области (ГФРП РО) Резервы, условные обязательства и условные активы
Резервы, условные обязательства и условные активы «Будь здоров»
«Будь здоров» КРОВООБРАЩЕНИЕ
КРОВООБРАЩЕНИЕ +
+ Фокус на Христе
Фокус на Христе Презентация (1) (1)
Презентация (1) (1) Хеллоуин. Кропивницкий
Хеллоуин. Кропивницкий Проект«Возможно ли в наше время сохранение единого русского литературного языка?»
Проект«Возможно ли в наше время сохранение единого русского литературного языка?» МОУ СОШ п. Тарбагатай «Сияющая радуга» - открытая школа, школа сотрудничества и диалога
МОУ СОШ п. Тарбагатай «Сияющая радуга» - открытая школа, школа сотрудничества и диалога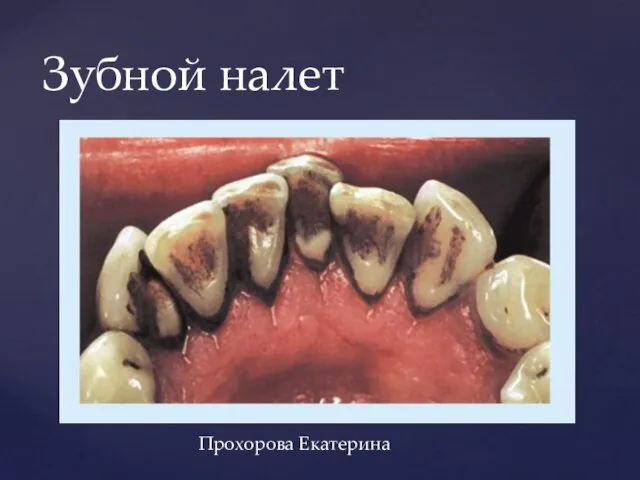 Презентация на тему Зубной налет
Презентация на тему Зубной налет Меню услуг Школы управления продажами
Меню услуг Школы управления продажами Король. Шахматы в школе
Король. Шахматы в школе Акционерное общество
Акционерное общество