- Физиология человека

Содержание
- 2. Физиология: определение Физиология (от греч. φύσις — природа и греч. λόγος — знание) — наука о
- 3. Строение клетки Примерно половина объема клетки занята органеллами, изолированными от цитозоля мембранами. Общая поверхность мембран внутриклеточных
- 4. Строение плазматической мембраны м Животные клетки ограничены плазматической мембраной. Основной матрикс мембраны состоит из липидов, главным
- 5. Строение плазматической мембраны Главными функциональными элементами, погруженными в сравнительно инертный липидныи матрикс мембраны, являются белки. Белок
- 6. Строение клетки. Мембранные системы внутриклеточных органелл. Примерно половина объема клетки занята органеллами, изолированными от цитозоля мембранами.
- 7. Строение клетки. Цитоскелет. Цитоплазма, окружающая органеллы, не является аморфной; ее, пронизывает сеть цитоскелета. Цитоскелет состоит из
- 8. Строение клетки. Цитозоль. Состав цитозоля. Содержимое цитозоля. Около половины объема клетки занято цитозолем. Поскольку он приблизительно
- 10. Обмен веществ между клеткой и окружающей средой Клетка не является статичным образованием, поскольку здесь происходит постоянный
- 11. Диффузия через мембранные поры. Мембраны ионных каналов. Плазматическая мембрана (и другие клеточные мембраны) проницаемы не только
- 12. Мембранные ионные каналы За поведением мембранных ионных каналов легко наблюдать, поскольку возникающий при движении ионов ток
- 13. Активный транспорт, натриевый насос. Натрий / Калиевый (Na/К) насос. В предыдущем разделе описаны пассивная диффузия ионов
- 14. Эндоцитоз. Экзоцитоз. Для некоторых веществ, которые поступают в клетку или должны быть выведены из нее, транспортные
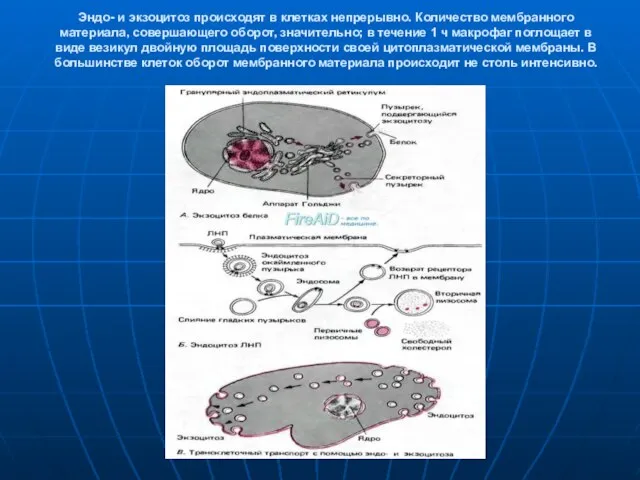
- 15. Эндо- и экзоцитоз происходят в клетках непрерывно. Количество мембранного материала, совершающего оборот, значительно; в течение 1
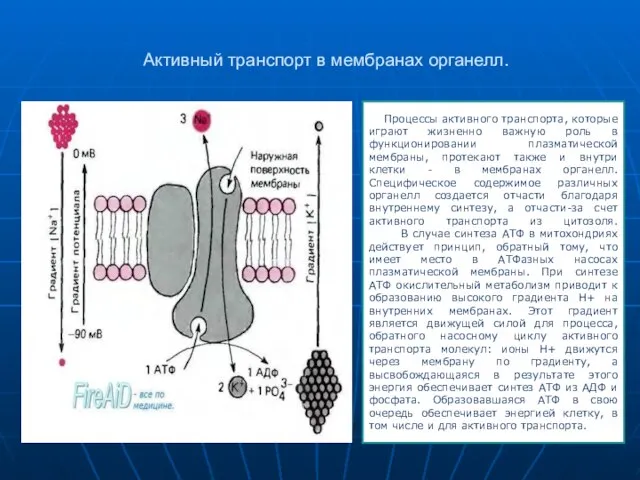
- 16. Активный транспорт в мембранах органелл. Процессы активного транспорта, которые играют жизненно важную роль в функционировании плазматической
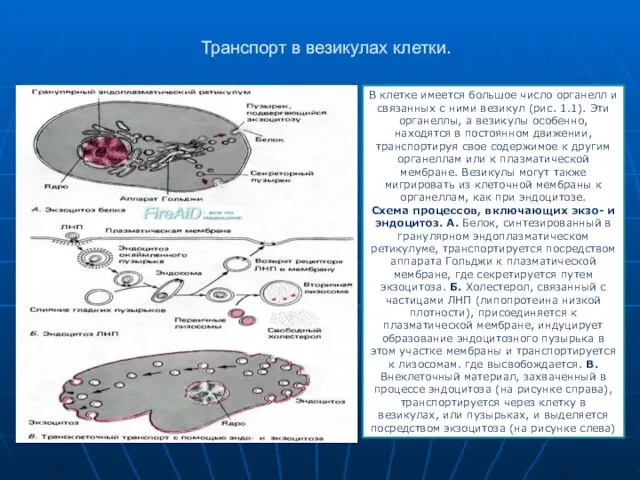
- 17. Транспорт в везикулах клетки. В клетке имеется большое число органелл и связанных с ними везикул (рис.
- 18. Процесс секреции белка представлен на рис. 1.12, А. Белок синтезируется вблизи ядра клетки на рибосомах, связанных
- 19. Регуляция клеточных функций. Регуляторные воздействия на клеточную мембрану. Мембранный потенциал. Поддержание индивидуальной клетки как функциональной единицы
- 20. Внеклеточные регуляторные вещества. Наиболее важным регуляторным механизмом с участием внеклеточных веществ является их взаимодействие со специфическими
- 21. Другие локальные агенты служат факторами роста. Наиболее известен фактор роста нервов (ФРН) для симпатических нейронов, который
- 23. Скачать презентацию

Слайд 3Строение клетки
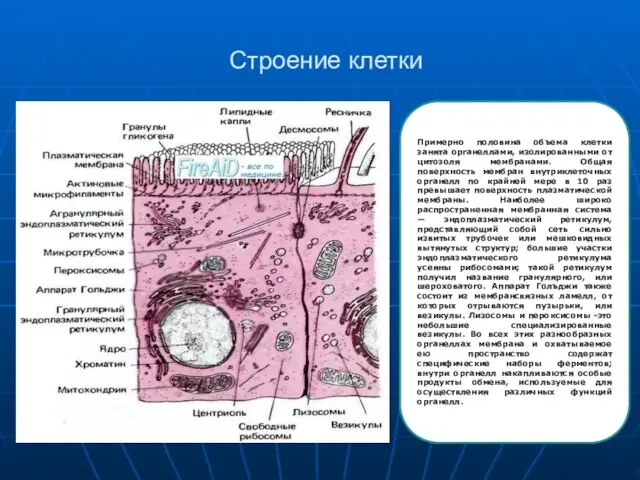
Примерно половина объема клетки занята органеллами, изолированными от цитозоля мембранами. Общая
Строение клетки
Примерно половина объема клетки занята органеллами, изолированными от цитозоля мембранами. Общая
Слайд 4Строение плазматической мембраны
м
Животные клетки ограничены плазматической мембраной.
Основной матрикс мембраны состоит из
Строение плазматической мембраны
м
Животные клетки ограничены плазматической мембраной.
Основной матрикс мембраны состоит из
Слайд 5Строение плазматической мембраны
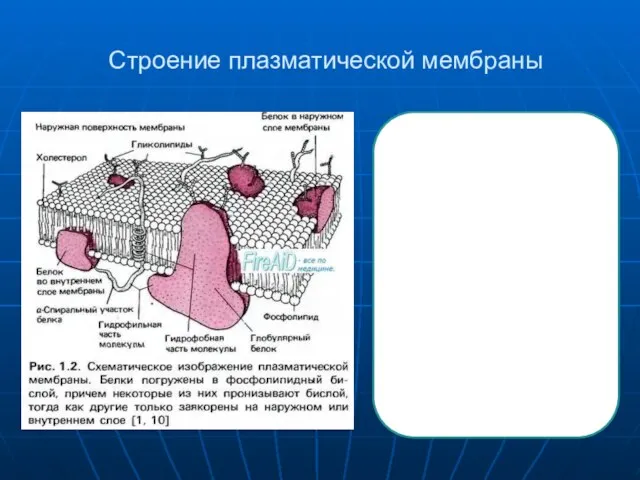
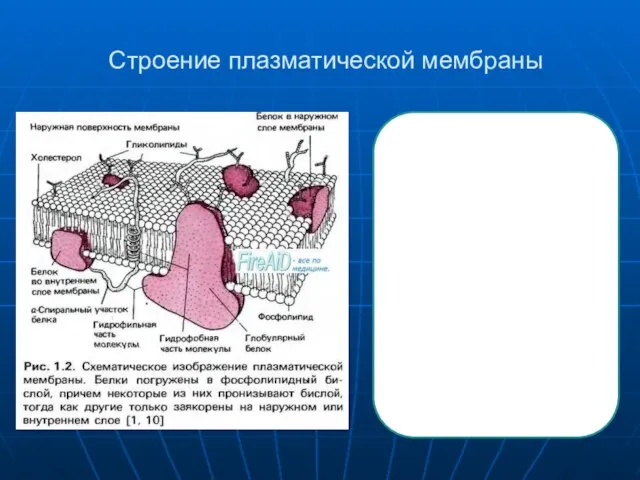
Главными функциональными элементами, погруженными в сравнительно инертный липидныи матрикс мембраны,
Строение плазматической мембраны
Главными функциональными элементами, погруженными в сравнительно инертный липидныи матрикс мембраны,
Слайд 6Строение клетки. Мембранные системы внутриклеточных органелл.
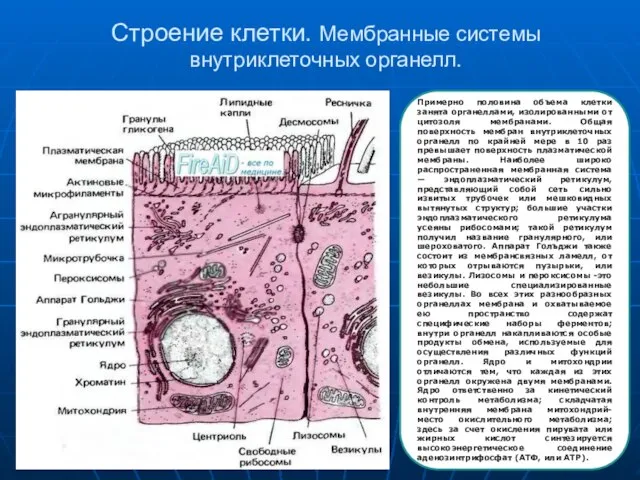
Примерно половина объема клетки занята органеллами, изолированными
Строение клетки. Мембранные системы внутриклеточных органелл.
Примерно половина объема клетки занята органеллами, изолированными
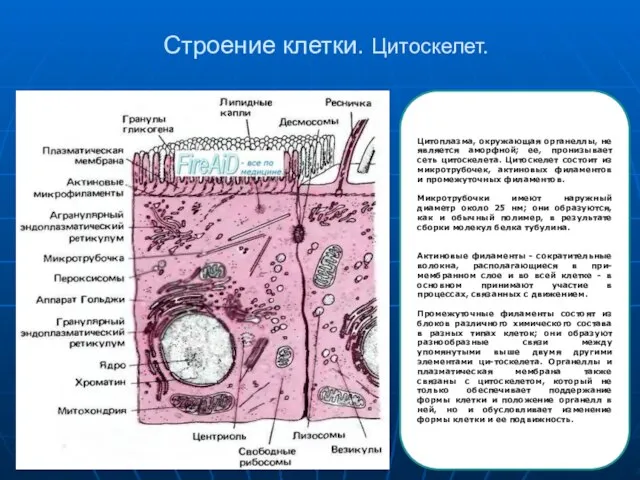
Слайд 7Строение клетки. Цитоскелет.
Цитоплазма, окружающая органеллы, не является аморфной; ее, пронизывает сеть цитоскелета.
Строение клетки. Цитоскелет.
Цитоплазма, окружающая органеллы, не является аморфной; ее, пронизывает сеть цитоскелета.
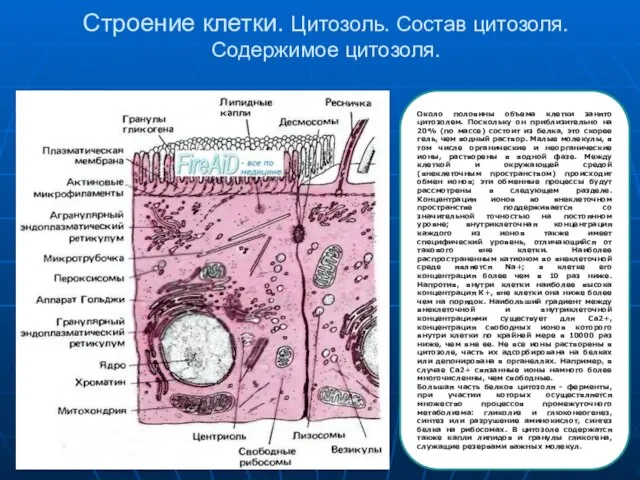
Слайд 8Строение клетки. Цитозоль. Состав цитозоля. Содержимое цитозоля.
Около половины объема клетки занято цитозолем.
Строение клетки. Цитозоль. Состав цитозоля. Содержимое цитозоля.
Около половины объема клетки занято цитозолем.

Слайд 10Обмен веществ между клеткой и окружающей средой
Клетка не является статичным образованием, поскольку
Обмен веществ между клеткой и окружающей средой
Клетка не является статичным образованием, поскольку

Слайд 11Диффузия через мембранные поры. Мембраны ионных каналов.
Плазматическая мембрана (и другие клеточные мембраны)
Диффузия через мембранные поры. Мембраны ионных каналов.
Плазматическая мембрана (и другие клеточные мембраны)
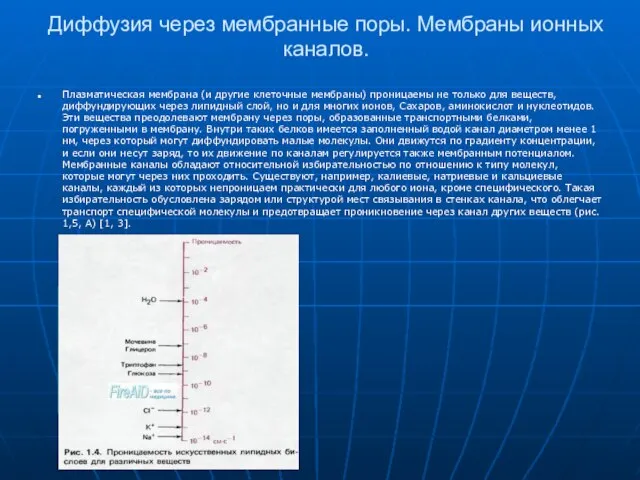
Слайд 12Мембранные ионные каналы
За поведением мембранных ионных каналов легко наблюдать, поскольку возникающий при
Мембранные ионные каналы
За поведением мембранных ионных каналов легко наблюдать, поскольку возникающий при

Слайд 13Активный транспорт, натриевый насос. Натрий / Калиевый (Na/К) насос.
В предыдущем разделе
Активный транспорт, натриевый насос. Натрий / Калиевый (Na/К) насос.
В предыдущем разделе
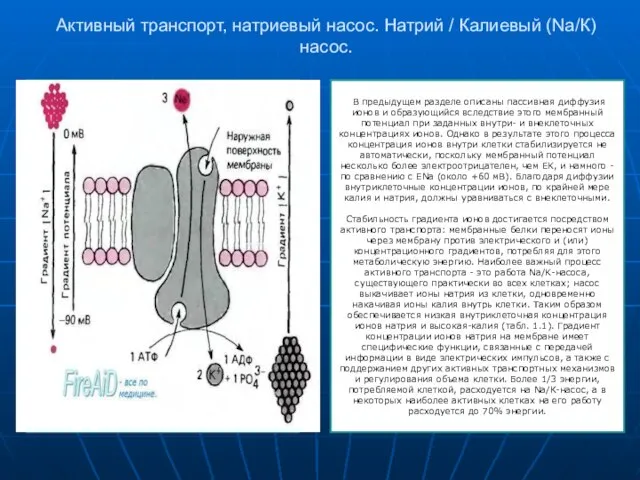
Слайд 14Эндоцитоз. Экзоцитоз.
Для некоторых веществ, которые поступают в клетку или должны быть
Эндоцитоз. Экзоцитоз.
Для некоторых веществ, которые поступают в клетку или должны быть

Слайд 15Эндо- и экзоцитоз происходят в клетках непрерывно. Количество мембранного материала, совершающего оборот,
Эндо- и экзоцитоз происходят в клетках непрерывно. Количество мембранного материала, совершающего оборот,
Слайд 16Активный транспорт в мембранах органелл.
Процессы активного транспорта, которые играют жизненно важную
Активный транспорт в мембранах органелл.
Процессы активного транспорта, которые играют жизненно важную
Слайд 17Транспорт в везикулах клетки.
В клетке имеется большое число органелл и связанных с
Транспорт в везикулах клетки.
В клетке имеется большое число органелл и связанных с
Слайд 18Процесс секреции белка представлен на рис. 1.12, А. Белок синтезируется вблизи ядра
Процесс секреции белка представлен на рис. 1.12, А. Белок синтезируется вблизи ядра

Слайд 19Регуляция клеточных функций. Регуляторные воздействия на клеточную мембрану. Мембранный потенциал.
Поддержание индивидуальной клетки
Регуляция клеточных функций. Регуляторные воздействия на клеточную мембрану. Мембранный потенциал.
Поддержание индивидуальной клетки

Слайд 20Внеклеточные регуляторные вещества. Наиболее важным регуляторным механизмом с участием внеклеточных веществ является
Внеклеточные регуляторные вещества. Наиболее важным регуляторным механизмом с участием внеклеточных веществ является

Слайд 21Другие локальные агенты служат факторами роста. Наиболее известен фактор роста нервов (ФРН)
Другие локальные агенты служат факторами роста. Наиболее известен фактор роста нервов (ФРН)

 Natur und Umwelt Ukraine
Natur und Umwelt Ukraine Теперешнее продолжительное время в английском
Теперешнее продолжительное время в английском HR-БРЕНДИНГ
HR-БРЕНДИНГ Печи и печная утварь XIX - начала XX веков
Печи и печная утварь XIX - начала XX веков Компания по продаже компьютеров Non Stop
Компания по продаже компьютеров Non Stop Olympic games (Олимпийские игры)
Olympic games (Олимпийские игры) Учебный центр Softline
Учебный центр Softline Корпоративная социальная ответственность бизнеса
Корпоративная социальная ответственность бизнеса  Управление проектамиЛекция 3.Календарное планирование и оперативное управление
Управление проектамиЛекция 3.Календарное планирование и оперативное управление Презентация проекта«Дидактическое пособиедля учеников 6 класса пофранцузскому языку.Les Curiosites de Paris.»Выполнила: Ильина АннаРук
Презентация проекта«Дидактическое пособиедля учеников 6 класса пофранцузскому языку.Les Curiosites de Paris.»Выполнила: Ильина АннаРук Презентация на тему Наука и культура в 1917-1945 гг
Презентация на тему Наука и культура в 1917-1945 гг  Финансовая грамотность населения России: проблемы и пути повышения Национальное агентство финансовых исследований 17 сентября 2009.
Финансовая грамотность населения России: проблемы и пути повышения Национальное агентство финансовых исследований 17 сентября 2009. Мне есть за что благодарить Творца
Мне есть за что благодарить Творца Молочная кринка
Молочная кринка Общая физическая подготовка в волейболе
Общая физическая подготовка в волейболе 8fac0b10a31b6bed2d5911b38d2c0d23
8fac0b10a31b6bed2d5911b38d2c0d23 Анализ взаимодействия и дорожная карта
Анализ взаимодействия и дорожная карта Технологическая эволюция рынка труда
Технологическая эволюция рынка труда История России в произведениях А. С. Пушкина
История России в произведениях А. С. Пушкина Урок презентация на тему Орфограммы в корне
Урок презентация на тему Орфограммы в корне Основы теории принятия решений. Лекция 3
Основы теории принятия решений. Лекция 3 Об особенностях внедрения ФГОС с этнокультурной составляющей Республики Бурятия
Об особенностях внедрения ФГОС с этнокультурной составляющей Республики Бурятия Простейшие элементы электронных схем
Простейшие элементы электронных схем Презентация на тему Афганская война
Презентация на тему Афганская война  Проблемное обучение в преподавании физики
Проблемное обучение в преподавании физики Презентация на тему Вечно холодные, вчено свободные
Презентация на тему Вечно холодные, вчено свободные ДЯТЕЛ ТЕТЕРЕВ ГЛУХАРЬ РЯБЧИК ГОЛУБЬ ФИЛИН СОВА.
ДЯТЕЛ ТЕТЕРЕВ ГЛУХАРЬ РЯБЧИК ГОЛУБЬ ФИЛИН СОВА. Игры народов мира
Игры народов мира