- Геном про и эукариот

Содержание
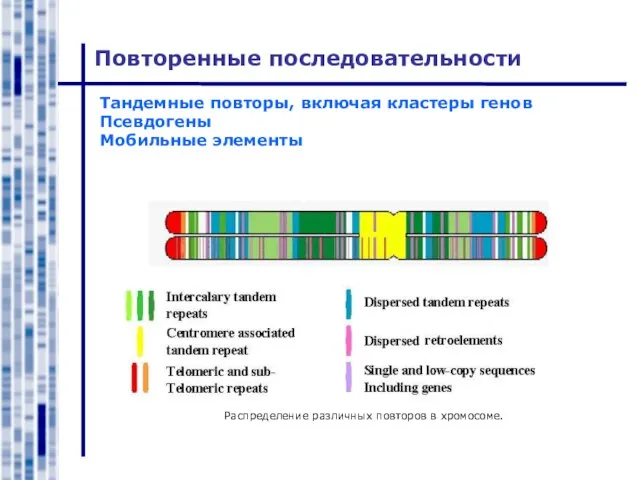
- 2. Повторенные последовательности Тандемные повторы, включая кластеры генов Псевдогены Мобильные элементы Распределение различных повторов в хромосоме.
- 3. Тандемы – сателлиты Сателлиты Drosophila melanogaster. Cателлиты – многократно повторенные короткие последовательности ДНК. Тандемные повторы с
- 4. Тандемы – сателлиты Наиболее часто встречающиеся короткие повторы, обнаруженные у крысы и собаки в различных хромосомах
- 5. Микросателлиты используются в качестве маркеров для популяционного анализа, а также в криминалистике и установлении отцовства (генетическая
- 6. Тандемы – кластеры генов Кластеры глобиновых генов в хромосомах 11 и 16 человека У эукариот гены,
- 7. Тандемы – кластеры генов Hox-кластер плодовой мушки и мыши. Считается, что дифференцировка частей тела проходит в
- 8. Тандемы – кластеры генов Предполагаемая эволюция Hox-кластера Кластеры генов возникают в результате дупликаций исходных генов и
- 9. Тандемы – кластеры генов Общая схема эволюции животных и их Hox-генов.
- 10. Тандемы – кластеры генов прокариот Однако не только у эукариот существуют кластеры генов. У бактерий обнаружены
- 11. Псевдогены Псевдогены – последовательности, сходные с обычными структурными генами, но, как правило, не экспрессирующиеся с образованием
- 12. Псевдогены Псевдогены образуются двумя путями: через дупликацию с последующей инактивацией копий мутациями и через интеграцию в
- 13. Мобильные элементы В геноме человека представлены non-LTR ретротранспозоны, LTR ретротранспозоны, эндогенные ретровирусы, ДНК транспозоны. В общей
- 14. Ретротранспозоны Среди ретротранспозонов выделяют три подкласса: - non-LTR ретротранспозоны - LTR ретротранспозоны - и SINE (short
- 15. Non-LTR ретротранспозоны Non-LTR ретротранспозоны используют праймер опосредованный механизм синтеза новой копии элемента (TPRT – target-primed reverse
- 16. LTR ретротранспозоны LTR ретротранспозоны используют механизм синтеза новой копии сходный с образованием новых копий кДНК ретровирусов.
- 17. SINE элементы SINE элементы используют ферментативную машину non-LTR ретротранспозонов для собственного перемещения. SINE элементы, чаще всего,
- 18. ДНК-транспозоны
- 19. ДНК-транспозоны Ранее считали, что ДНК-транспозоны эукариот делятся по механизму перемещения на два класса: размножающиеся с помощью
- 20. ДНК-транспозоны Основной механизм перемещения ДНК-транспозонов – вырезание/встраивание (cut-and-paste).
- 21. Мобильные элементы Мобильные элементы в геноме человека и мыши Состав и количество мобильных элементов сильно варьирует
- 22. Мобильные элементы - роль Мобильные элементы могут играть существенную роль в организации и эволюции генома
- 23. Мобильные элементы - роль Наиболее известный пример участия мобильных элементов в функционировании генома – сохранение теломер
- 24. Мобильные элементы - роль Различные варианты хромосомных перестроек в результате рекомбинаций между повторами, в частности –
- 26. Скачать презентацию
Слайд 2Повторенные последовательности
Тандемные повторы, включая кластеры генов
Псевдогены
Мобильные элементы
Распределение различных повторов в хромосоме.
Повторенные последовательности
Тандемные повторы, включая кластеры генов
Псевдогены
Мобильные элементы
Распределение различных повторов в хромосоме.
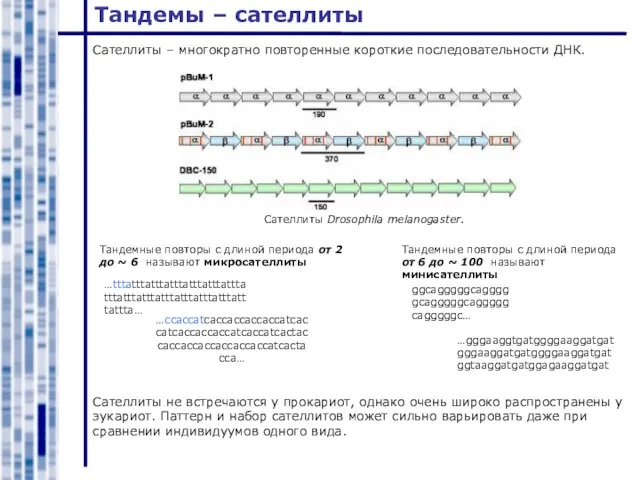
Слайд 3Тандемы – сателлиты
Сателлиты Drosophila melanogaster.
Cателлиты – многократно повторенные короткие последовательности ДНК.
Тандемные повторы
Тандемы – сателлиты
Сателлиты Drosophila melanogaster.
Cателлиты – многократно повторенные короткие последовательности ДНК.
Тандемные повторы
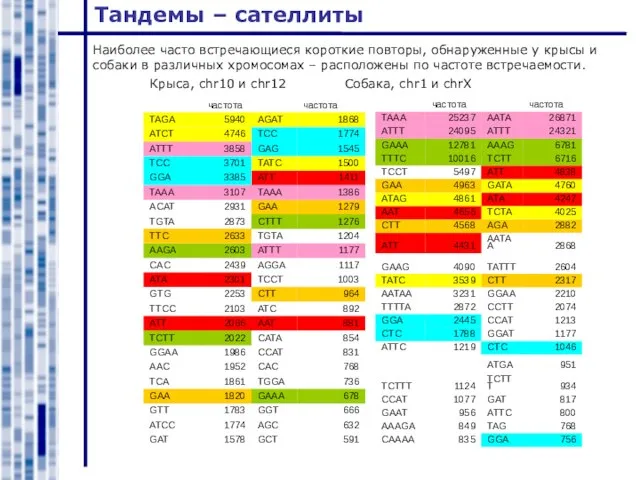
Слайд 4Тандемы – сателлиты
Наиболее часто встречающиеся короткие повторы, обнаруженные у крысы и собаки
Тандемы – сателлиты
Наиболее часто встречающиеся короткие повторы, обнаруженные у крысы и собаки
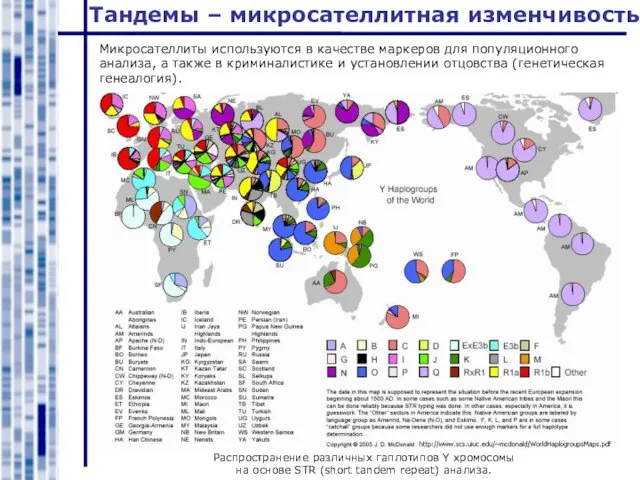
Слайд 5Микросателлиты используются в качестве маркеров для популяционного анализа, а также в криминалистике
Микросателлиты используются в качестве маркеров для популяционного анализа, а также в криминалистике
Слайд 6Тандемы – кластеры генов
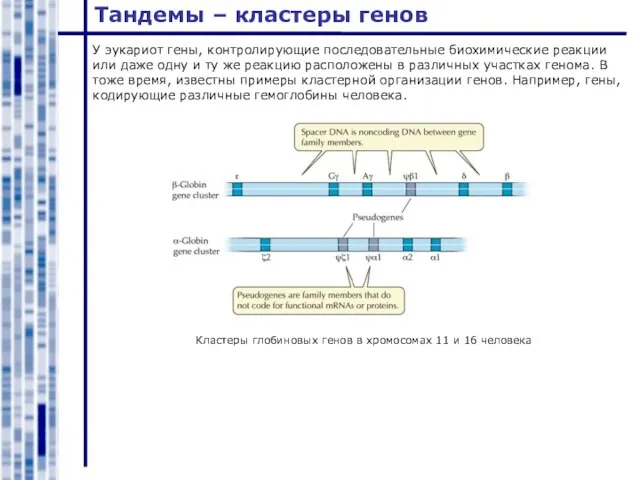
Кластеры глобиновых генов в хромосомах 11 и 16 человека
У
Тандемы – кластеры генов
Кластеры глобиновых генов в хромосомах 11 и 16 человека
У
Слайд 7Тандемы – кластеры генов
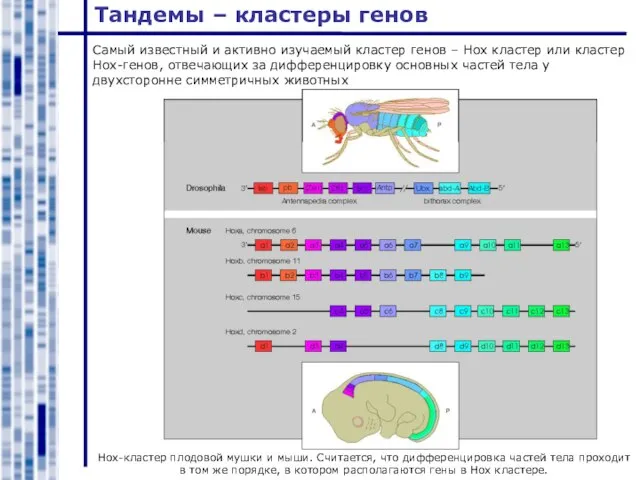
Hox-кластер плодовой мушки и мыши. Считается, что дифференцировка частей
Тандемы – кластеры генов
Hox-кластер плодовой мушки и мыши. Считается, что дифференцировка частей
Слайд 8Тандемы – кластеры генов
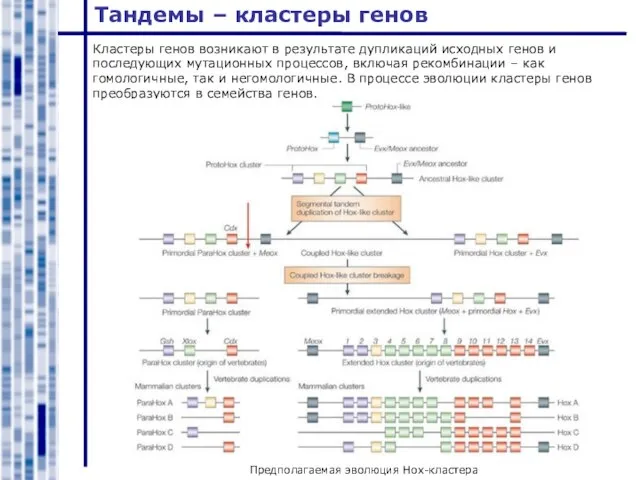
Предполагаемая эволюция Hox-кластера
Кластеры генов возникают в результате дупликаций исходных
Тандемы – кластеры генов
Предполагаемая эволюция Hox-кластера
Кластеры генов возникают в результате дупликаций исходных
Слайд 9Тандемы – кластеры генов
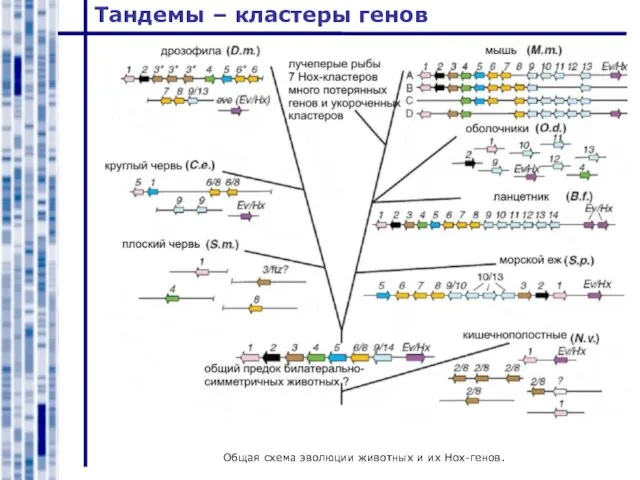
Общая схема эволюции животных и их Hox-генов.
Тандемы – кластеры генов
Общая схема эволюции животных и их Hox-генов.
Слайд 10Тандемы – кластеры генов прокариот
Однако не только у эукариот существуют кластеры генов.
Тандемы – кластеры генов прокариот
Однако не только у эукариот существуют кластеры генов.

Слайд 11Псевдогены
Псевдогены – последовательности, сходные с обычными структурными генами, но, как правило, не
Псевдогены
Псевдогены – последовательности, сходные с обычными структурными генами, но, как правило, не
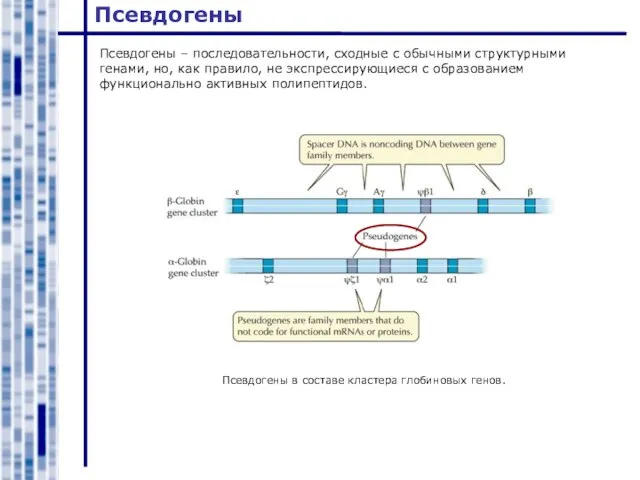
Слайд 12Псевдогены
Псевдогены образуются двумя путями: через дупликацию с последующей инактивацией копий мутациями и
Псевдогены
Псевдогены образуются двумя путями: через дупликацию с последующей инактивацией копий мутациями и
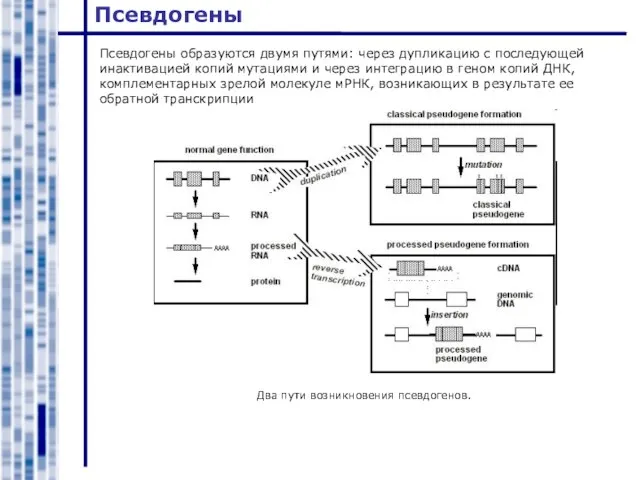
Слайд 13Мобильные элементы
В геноме человека представлены non-LTR ретротранспозоны, LTR ретротранспозоны, эндогенные ретровирусы, ДНК
Мобильные элементы
В геноме человека представлены non-LTR ретротранспозоны, LTR ретротранспозоны, эндогенные ретровирусы, ДНК

Слайд 14Ретротранспозоны
Среди ретротранспозонов выделяют три подкласса:
- non-LTR ретротранспозоны
- LTR ретротранспозоны
- и SINE
Ретротранспозоны
Среди ретротранспозонов выделяют три подкласса:
- non-LTR ретротранспозоны
- LTR ретротранспозоны
- и SINE
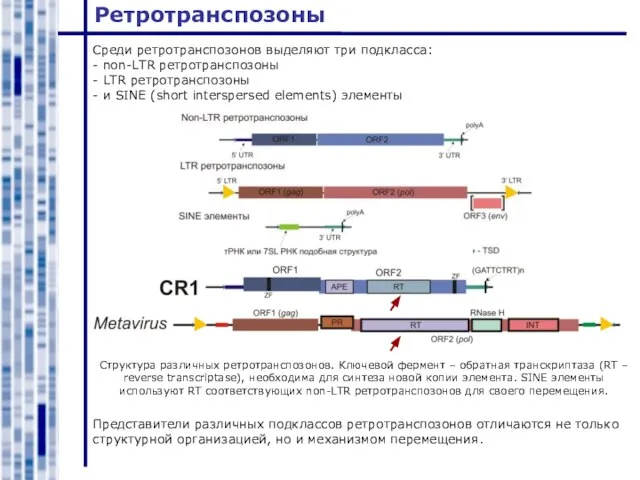
Слайд 15Non-LTR ретротранспозоны
Non-LTR ретротранспозоны используют праймер опосредованный механизм синтеза новой копии элемента (TPRT
Non-LTR ретротранспозоны
Non-LTR ретротранспозоны используют праймер опосредованный механизм синтеза новой копии элемента (TPRT
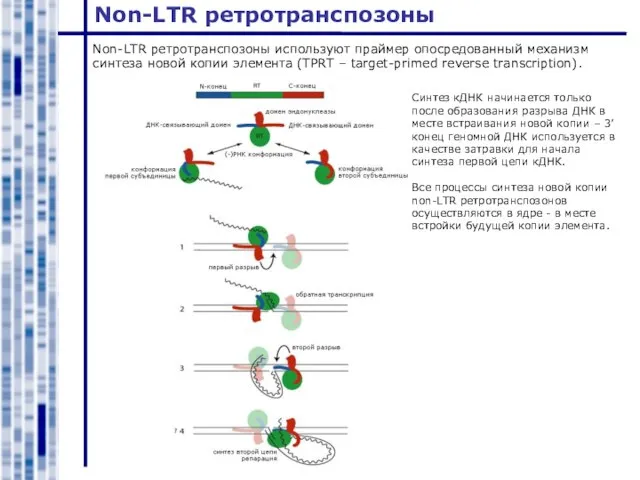
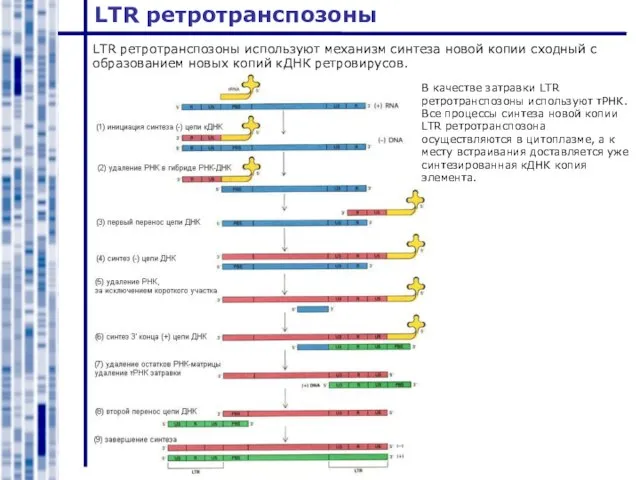
Слайд 16LTR ретротранспозоны
LTR ретротранспозоны используют механизм синтеза новой копии сходный с образованием новых
LTR ретротранспозоны
LTR ретротранспозоны используют механизм синтеза новой копии сходный с образованием новых
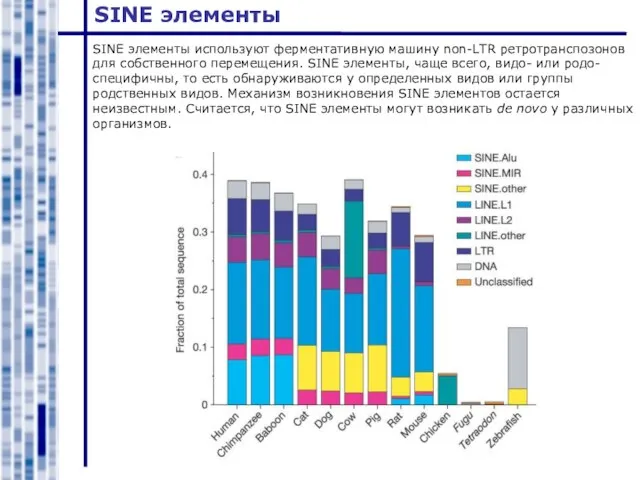
Слайд 17SINE элементы
SINE элементы используют ферментативную машину non-LTR ретротранспозонов для собственного перемещения. SINE
SINE элементы
SINE элементы используют ферментативную машину non-LTR ретротранспозонов для собственного перемещения. SINE
Слайд 18ДНК-транспозоны
ДНК-транспозоны
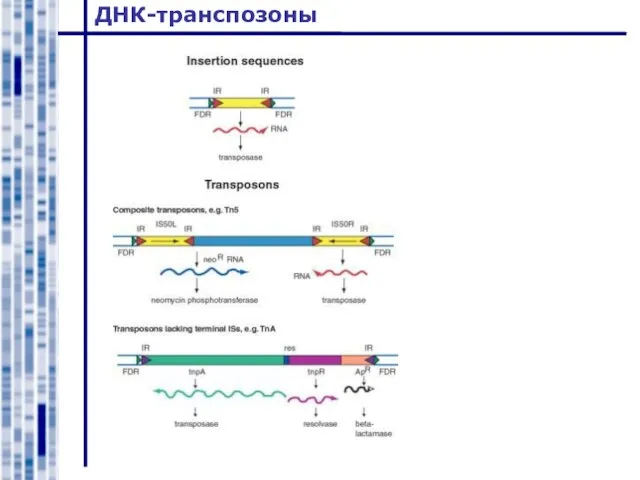
Слайд 19ДНК-транспозоны
Ранее считали, что ДНК-транспозоны эукариот делятся по механизму перемещения на два класса:
ДНК-транспозоны
Ранее считали, что ДНК-транспозоны эукариот делятся по механизму перемещения на два класса:

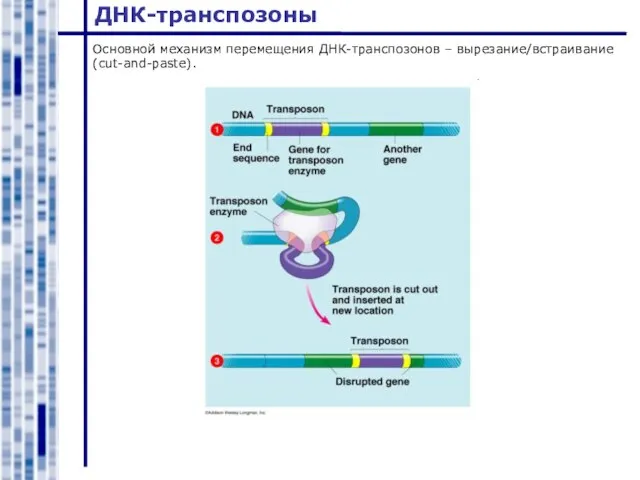
Слайд 20ДНК-транспозоны
Основной механизм перемещения ДНК-транспозонов – вырезание/встраивание (cut-and-paste).
ДНК-транспозоны
Основной механизм перемещения ДНК-транспозонов – вырезание/встраивание (cut-and-paste).
Слайд 21Мобильные элементы
Мобильные элементы в геноме человека и мыши
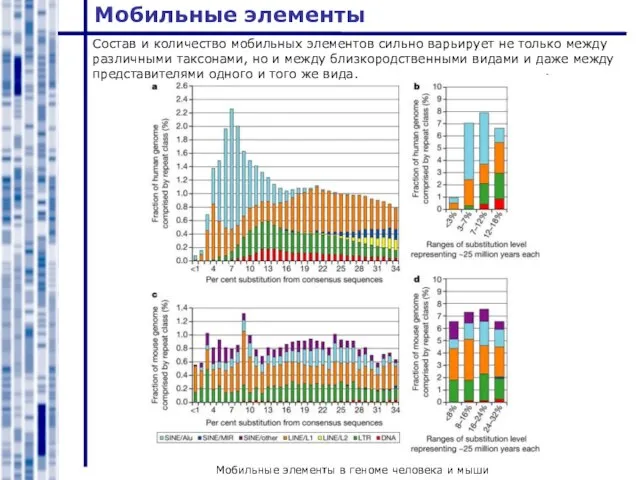
Состав и количество мобильных элементов
Мобильные элементы
Мобильные элементы в геноме человека и мыши
Состав и количество мобильных элементов
Слайд 22Мобильные элементы - роль
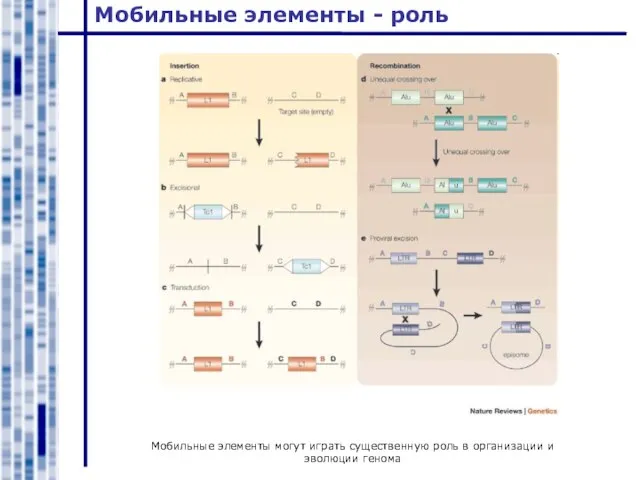
Мобильные элементы могут играть существенную роль в организации и
Мобильные элементы - роль
Мобильные элементы могут играть существенную роль в организации и
Слайд 23Мобильные элементы - роль
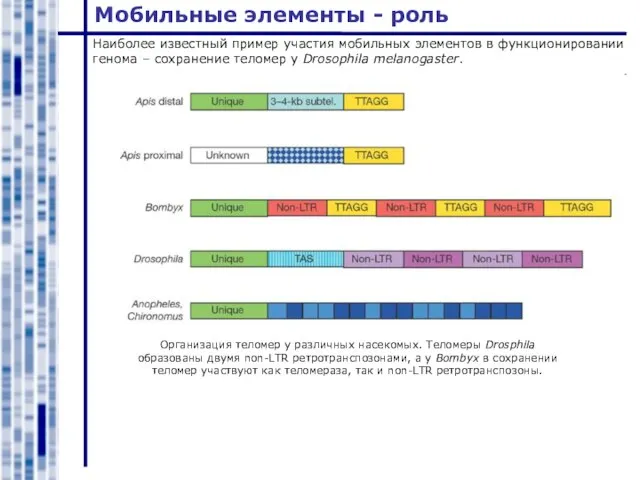
Наиболее известный пример участия мобильных элементов в функционировании генома
Мобильные элементы - роль
Наиболее известный пример участия мобильных элементов в функционировании генома
Слайд 24Мобильные элементы - роль
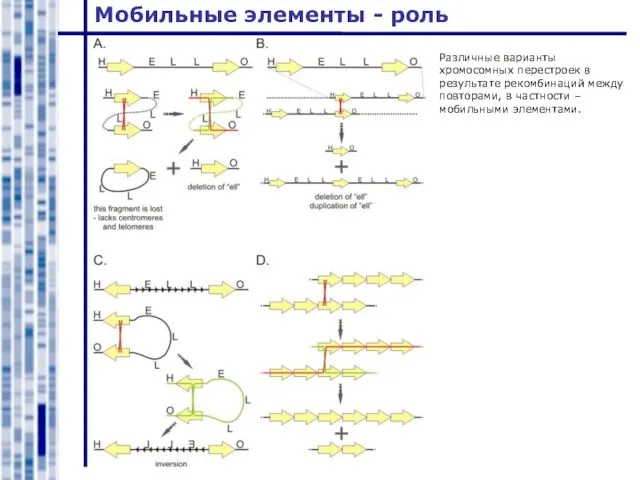
Различные варианты хромосомных перестроек в результате рекомбинаций между повторами,
Мобильные элементы - роль
Различные варианты хромосомных перестроек в результате рекомбинаций между повторами,
 Forses and Motion
Forses and Motion  Легкая атлетика.Техника бега на короткие дистанции
Легкая атлетика.Техника бега на короткие дистанции The Sources of Philosophy (India, China)
The Sources of Philosophy (India, China) Вопросы по инвестициям
Вопросы по инвестициям 1
1 Областная экспедиция Музейная коллекция, посвящённая 75-летию образования Костромской области
Областная экспедиция Музейная коллекция, посвящённая 75-летию образования Костромской области Презентация на тему Зачем люди осваивают космос
Презентация на тему Зачем люди осваивают космос  Уголовная ответственность несовершеннолетних
Уголовная ответственность несовершеннолетних Культура и образование в информационный век: направления сотрудничества
Культура и образование в информационный век: направления сотрудничества Севастопольская городская молодежная организация
Севастопольская городская молодежная организация SRO #3. SmartCat (2)
SRO #3. SmartCat (2) Университетский округ инновационных образовательных учреждений при ПГПУ
Университетский округ инновационных образовательных учреждений при ПГПУ Вот и кончилось лето
Вот и кончилось лето Наши клиенты
Наши клиенты Сегментация изображений
Сегментация изображений Доли. Обыкновенные дроби
Доли. Обыкновенные дроби Внедрение информационных технологийв образовательный процесс начальной школы.
Внедрение информационных технологийв образовательный процесс начальной школы. Всемирный день качества
Всемирный день качества Клиентоориентированность. Рыбный мир 55
Клиентоориентированность. Рыбный мир 55 Факультет экономики и права ФГБОУ ВО Оренбургский ГАУ
Факультет экономики и права ФГБОУ ВО Оренбургский ГАУ ИЗО
ИЗО Влияет ли одежда на здоровье подростка ?
Влияет ли одежда на здоровье подростка ? QNX Software Systems
QNX Software Systems поликарбонатные профили +система монтажа сотового поликарбоната
поликарбонатные профили +система монтажа сотового поликарбоната 03 - System Specifications Rev B
03 - System Specifications Rev B Презентация на тему Иммунная система организма
Презентация на тему Иммунная система организма  South Korea (фотографии)
South Korea (фотографии) Виды воды
Виды воды