- Молекулярная физиология синаптической передачи

Содержание
- 2. Содержание лекции Введение. Синаптическая пластичность Электрическое взаимодействие клеток: электрический и химический синапсы. Электрический синапс. Функции Химический
- 3. Введение.Синаптическая пластичность СП - лежат в основе обучения и памяти, но сложность взаимодействия ионных каналов, ферментов
- 4. Методы исследования синаптогенеза Two-photon laser scanning fluorescence microscopy in vivo. . Двух-фотонная микроскопия была до сих
- 5. Синаптическая пластичность
- 6. Синаптическая пластичность
- 7. Синаптическая пластичность
- 8. Электрическое взаимодействие клеток Электрический синапс Химический синапс Термин синапс был введён в 1897 г. английским физиологом
- 9. Электрический синапс. Функции 1. Быстродействие, что позволяет обеспечивать быстрые реакции организма. Например, гигантские нейроны нервных ганглиев
- 10. Принцип работы электрического синапса
- 11. Транспорт веществ через щелевой контакт Одно из важнейших свойств щелевого контакта как транспортной системы состоит в
- 12. Химический синапс Функция – передача электрического сигнала от пресинаптического нейрона на постсинаптическую клетку
- 13. Этапы реализации функции Освобождение из везикул пресинаптической зоны специфических переносчиков, называемых нейротрансмиттерами, или медиаторами. Диффузия к
- 14. Химический синапс и принцип его работы
- 15. Везикулярное высвобождение нейропередатчика в ответ на пресинаптический ПД. Конвертация химического сигнала в электрический в постсинапсе Приход
- 16. Синтез и рецикл синаптических везикул и их содержимого Аксональный транспорт: - медленный аксональный транспорт (1 мм/сутки
- 17. Синтез и рецикл синаптических везикул и их содержимого А - Биосинтез нейротрансмиттеров. I. Везикулы, предшественники пептидных
- 18. Цикл синаптических везикул : кластеризация и докинг везикул Везикулы наполненные нейропередатчиком образуют кластер в районе активной
- 19. Механизм высвобождения трансмиттера Во время деполяризации в нервном окончании возникает входящий ток Na+. Мембрана окончания аксона
- 20. Белки экзацитоза синаптических везикул Синаптические везикулы фиксируются большей частью к цитоскелету посредством протеина синапсина (synapsin), локализованного
- 21. Белки экзацитоза синаптических везикул Синаптические везикулы фиксируются большей частью к цитоскелету посредством протеина синапсина (synapsin), локализованного
- 22. Synaptotagmin Synaptotagmin 1 (или synaptotagmin) сенсор ионов Ca2+ датчик в мембране pre-synaptic axon. Кальций связывающие synaptotagmins
- 23. Neurexin Neurexin (NRXN) пресинаптическими белок, который участвует в соединении нейронов и с образованием синапса.[1] Они расположены
- 24. Neurexin and neuroligin Транс-synaptic диалог между neurexin и neuroligin организует аппозиция pre - и post-synaptic machinery
- 25. The β-neurexin-neuroligin-1 junction is the core of a newly forming synapse Бета-neurexin-neuroligin-1 junction является ядром формирования
- 26. Munc-18 Munc-18 (аббревиатура от mammalian uncoordinated-18) - белки являются гомологом у млекопитающих unc-18 белков (которые могут
- 27. Синаптофизин Синаптофизин – это гликопротеин, находящийся в пресинаптических везикулах нейронов мозга, спинного мозга, ретине, везикулах адреналиновой
- 28. Синаптобревин Синаптобревин (англ. synaptobrevin) — небольшой трансмембранный белок секреторных везикул; - компонент белкового комплекса SNARE, осуществляющего
- 29. Rab-3A Ras-связанный белок Rab-3А есть белок в организме человека, который кодируется геном RAB3A.[1][2][3] Он участвует в
- 30. Взаимодействие везикулярных белков и белков пресинаптической мембраны
- 31. Белки экзацитоза синаптических везикул Меньшая часть везикул также связана с внутренней стороной пресинаптической мембраны с помощью
- 32. Роль ионов кальция Если потенциал действия достиг пресинаптической области, и в пресинаптическом окончании концентрация Ca2+ поднялась
- 33. Роль ионов кальция Во-вторых, повышенный уровень ионов Ca2+ в пресинаптическом окончании активирует Ca2+-кальмодулин-зависимую протеинкиназу II (СаМ-киназа
- 34. Устройство синаптической везикулы Первые открытые белки синаптической везикулы: синапсинI, синаптофизин и синаптобревин (VAMP1) Синапсин связывает Везикулу
- 35. SNARE SNARE– главный компонент механизма слияния синаптической везикулы с мембраной. •Состоит из 3 синаптических белков: –Синаптобревина
- 36. SNARE и действие токсинов Бутулотоксин и столбнячный токсин (тетеноспазмин) –протеазы “разрезающие” белки SNARE комплекса
- 37. Этапы механизма высвобождения трансмиттера из везикулы синаптобревин
- 38. Слияние везикулы Слияние везикул с пресинаптической мембраной управляется белками SNARE, в результате чего происходит выделение содержимого
- 39. Exocytotic machinery
- 40. Diagram of the process of exocytosis
- 41. Цикл синаптических везикул : : возвращение в высвобождаемый пул везикул 1. Простое закрытие поры слияния и
- 42. Преобразование трансмиттера. Расщепление, удаление и обратный захват трансмиттера на примере ацетилхолина
- 43. Типы химических синапсов Синапс с ионотропным рецептором - связывание с молекулой трансмиттера, открывает ионный канал (каналы,
- 44. Перечень соединений, влияющие на никотиновые, мускариновые и холинергические синапсы
- 45. Ионотропные синапсы
- 46. Метаботропные синапсы
- 47. Механизмы работы ионотропного синапса Холин
- 48. Механизмы работы метаботропного синапса Белок G
- 49. Типы трансмиттеров и механизмы их действия Ацетилхолин – медиатор всех двигательных нейронов, многих интернейронов ЦНС, а
- 50. Типы трансмиттеров и механизмы их действия Глицин служит трансмиттером тормозных синапсов и действует как нейромодулятор. Глицин
- 51. Типы трансмиттеров и механизмы их действия γ-Аминомасляную кислоту выбрасывают в качестве трансмиттера многие тормозные интернейроны, имеющиеся
- 52. Типы трансмиттеров и механизмы их действия Серотонин (5-hydroxytryptamin) - биогенный амин, широко распространенный в ЦНС. В
- 53. Типы трансмиттеров и механизмы их действия Гистамин - моноамин, выступающий в качестве трансмиттера. Особенно важную роль
- 54. Типы трансмиттеров и механизмы их действия Дофамин - биогенный амин, наиболее представленный в базальных ганглиях, где
- 55. Типы трансмиттеров и механизмы их действия Норадреналин катехоламин, выполняющий функцию трансмиттера в ЦНС и синтезирующийся прежде
- 56. Типы трансмиттеров и механизмы их действия Олигопептиды, которые действуют как трансмиттеры или нейромодуляторы - это приемущественно
- 57. Типы трансмиттеров и механизмы их действия Другие нейропептиды: вещество Р, ангиотензин II, соматостатин, вазоактивный интестинальный полипептид
- 58. Примеры механизмов действия трансмиттеров на различные типы рецепторов
- 59. Ионотропный никотиновый холинергический синапс/Структура n-холинорецептора
- 60. Структура n-холинорецептора Рецепторный белок для нАЦХ-рецептора образует пору - ионный канал т пронизывает клеточную мембрану. Канал
- 61. Глутаматный синапс Лиганд-управляемые катионные каналы - NMDA (N-метил-D-аспартат) и AMPA (α-ами- но-3-гидрокси-5-метил-4-изоксазол-пропионовая кислота) Метаботропный mGluR1-5, действие
- 62. Активация глутаматом два ионотропный лиганд-управляемый рецептор, служащий катионным каналом AMPA, и метаботропный рецептор mGluR1-5. Повышение уровня
- 63. Фосфорилирование и дефосфорилирование в глутаматном синапсе Глутаматный синапс содержит на постсинаптической мембране два типа ионотропных рецепторов.
- 64. Глутамат и механизм работы синапса Глутамат и механизм работы синапса через ионотропные рецепторы (лигандуправляемые катионные каналы)
- 65. Глутамат и механизм работы синапса Глутамат и механизм работы синапса через ионотропные рецепторы (лигандуправляемые катионные каналы)
- 66. Механизм долговременной потенциации и долговременной синаптической депрессии
- 67. Механизм долговременной потенциации и долговременной синаптической депрессии Освобожденный из пресинаптической области глутамат активирует ионотропный рецептор -
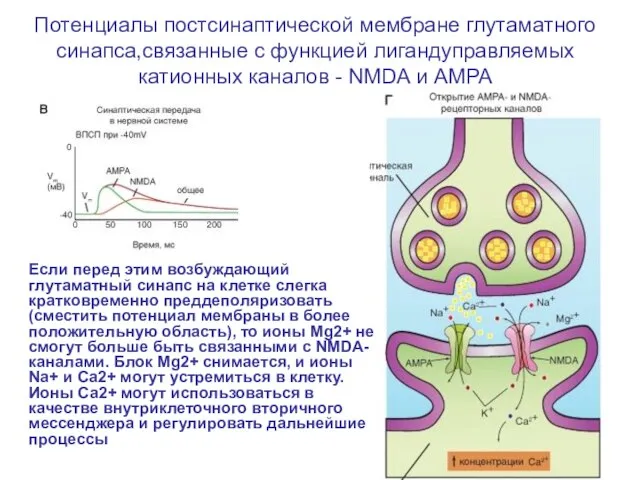
- 68. Потенциалы постсинаптической мембране глутаматного синапса,связанные с функцией лигандуправляемых катионных каналов - NMDA и AMPA Потенциалы, генерируемые
- 69. Потенциалы постсинаптической мембране глутаматного синапса,связанные с функцией лигандуправляемых катионных каналов - NMDA и AMPA Если перед
- 70. ГАМК как трансмиттер ГАМК оказывает тормозное действие на постсинаптические структуры. Существует по меньшей мере два различных
- 71. Глицин как трансмиттер Аминокислота глицин выполняет функцию трансмиттера в тормозных синапсах и действует как нейромодулятор. Глицин
- 72. NO как трансмиттер Оксид азота (NO) - это паракринный медиатор, выделяемый эндотелиальными клетками и некоторыми нейронами.
- 73. NO как трансмиттер Образование NO катализируется NO-синтазой NOS - Са2+-кальмодулин-зависимым ферментом, ускоряющим превращение аргинина в цитруллин
- 74. Постсинаптическая часть Постсинаптическая часть различается в зависимости от: 1.Типа пресинаптического нейрона (высвобождаемого нейропередатчика) 2.Типа постсинаптического нейрона
- 75. Классификация постсинапса По наличию или отсутствию шипика –синапсы шипиковые и сидячие По наличию или отсутствию постсинаптического
- 76. Типы дендритных шипиков Шипик имеет шейку и головку На головке шипика расположена активная зона (место контакта
- 77. Свойства дендритных шипиков Могут изменять свою геометрию (ширину, длину шейки, Диаметр головки) это приводит к изменению
- 78. Возбуждающий постсинаптический потенциал (ВПСП) ВПСП возникает благодаря повышению проводимости для натрия и тем самым благодаря входящему
- 79. ВПСП и ТПСП на мембране клетки взаимно влияют друг на друга Механизм возникновения постсинаптических потенциалов соответствует
- 80. Соединения с синаптическим влиянием Наряду с трансмиттером, другие соединения тоже могут влиять на рецепторный белок. Если
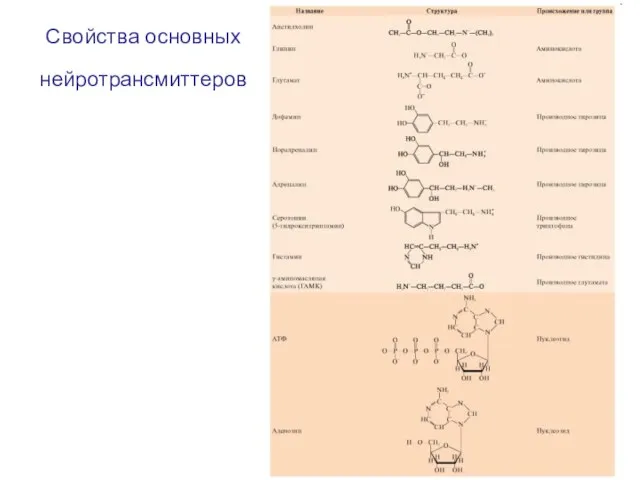
- 81. Свойства основных нейротрансмиттеров
- 82. Свойства основных нейротрансмиттеров
- 84. Скачать презентацию
Слайд 2Содержание лекции
Введение. Синаптическая пластичность
Электрическое взаимодействие клеток: электрический и химический синапсы.
Электрический синапс.
Содержание лекции
Введение. Синаптическая пластичность
Электрическое взаимодействие клеток: электрический и химический синапсы.
Электрический синапс.

Слайд 3Введение.Синаптическая пластичность
СП - лежат в основе обучения и памяти, но сложность
Введение.Синаптическая пластичность
СП - лежат в основе обучения и памяти, но сложность

Слайд 4Методы исследования синаптогенеза
Two-photon laser scanning fluorescence
microscopy in vivo.
. Двух-фотонная микроскопия была
Методы исследования синаптогенеза
Two-photon laser scanning fluorescence
microscopy in vivo.
. Двух-фотонная микроскопия была

Слайд 5Синаптическая пластичность
Синаптическая пластичность
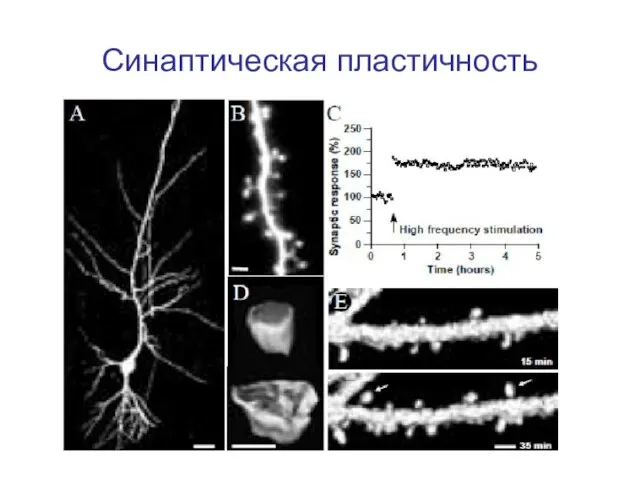
Слайд 6Синаптическая пластичность
Синаптическая пластичность
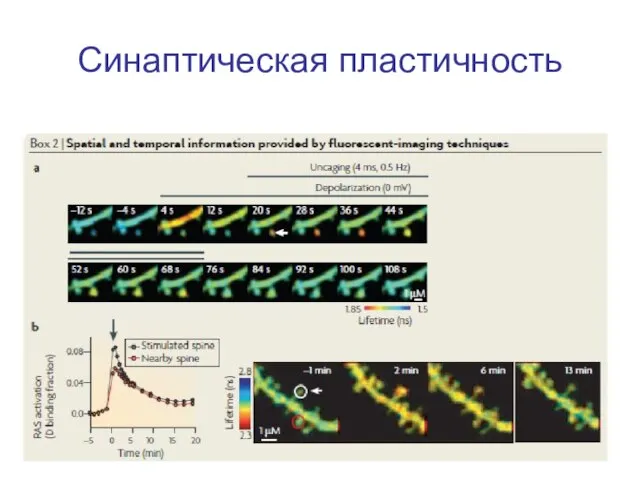
Слайд 7Синаптическая пластичность
Синаптическая пластичность
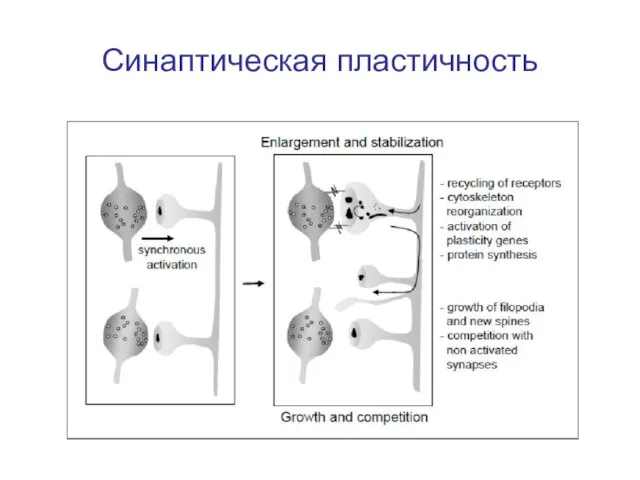
Слайд 8Электрическое взаимодействие клеток
Электрический синапс
Химический синапс
Термин синапс был введён в 1897 г.
Электрическое взаимодействие клеток
Электрический синапс
Химический синапс
Термин синапс был введён в 1897 г.

Слайд 9Электрический синапс. Функции
1. Быстродействие, что позволяет обеспечивать быстрые реакции организма. Например, гигантские
Электрический синапс. Функции
1. Быстродействие, что позволяет обеспечивать быстрые реакции организма. Например, гигантские

Слайд 10Принцип работы электрического синапса
Принцип работы электрического синапса
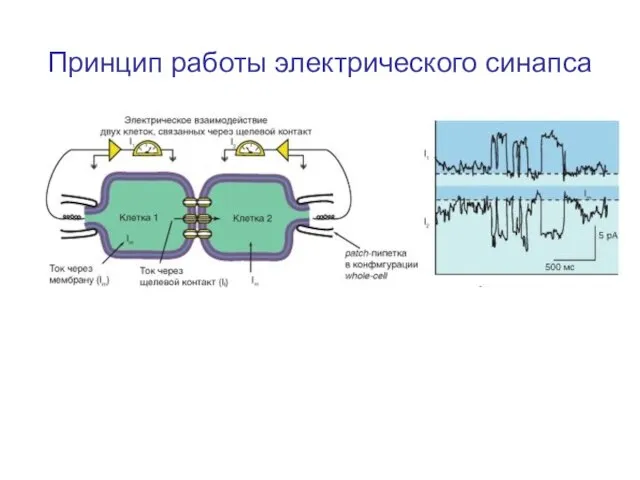
Слайд 11Транспорт веществ через щелевой контакт
Одно из важнейших свойств щелевого контакта как транспортной
Транспорт веществ через щелевой контакт
Одно из важнейших свойств щелевого контакта как транспортной

Слайд 12Химический синапс
Функция – передача электрического сигнала от пресинаптического нейрона на постсинаптическую клетку
Химический синапс
Функция – передача электрического сигнала от пресинаптического нейрона на постсинаптическую клетку

Слайд 13Этапы реализации функции
Освобождение из везикул пресинаптической зоны специфических переносчиков, называемых нейротрансмиттерами,
Этапы реализации функции
Освобождение из везикул пресинаптической зоны специфических переносчиков, называемых нейротрансмиттерами,

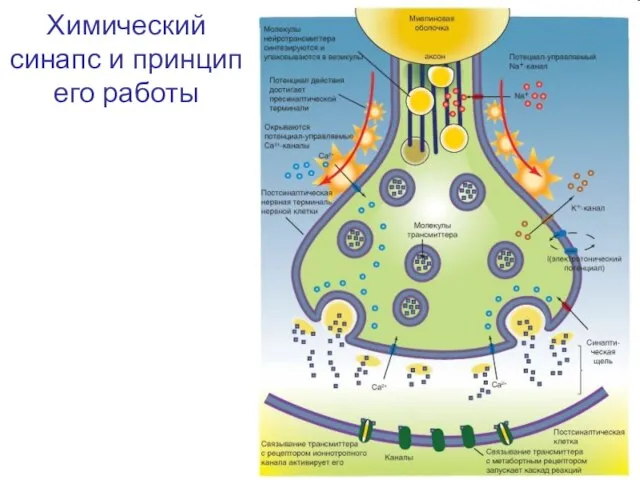
Слайд 14Химический синапс и принцип его работы
Химический синапс и принцип его работы
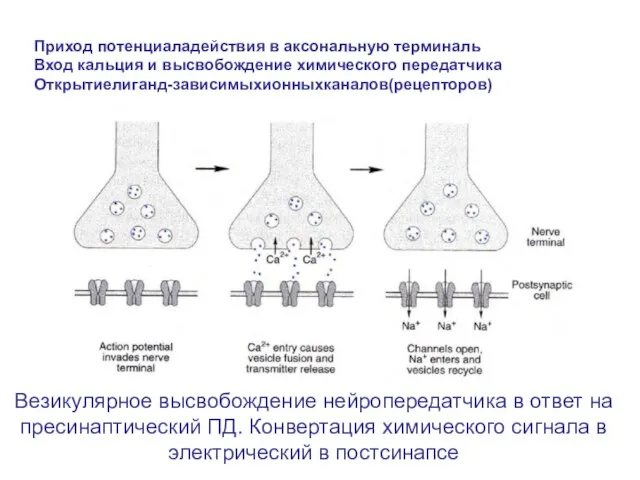
Слайд 15Везикулярное высвобождение нейропередатчика в ответ на пресинаптический ПД. Конвертация химического сигнала в
Везикулярное высвобождение нейропередатчика в ответ на пресинаптический ПД. Конвертация химического сигнала в
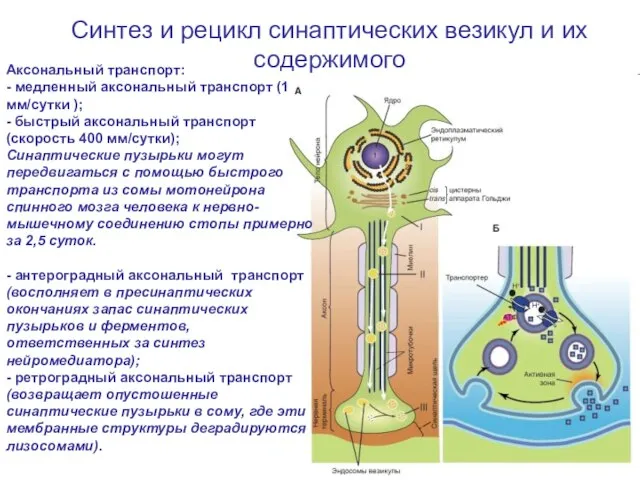
Слайд 16Синтез и рецикл синаптических везикул и их содержимого
Аксональный транспорт:
- медленный аксональный
Синтез и рецикл синаптических везикул и их содержимого
Аксональный транспорт:
- медленный аксональный
Слайд 17Синтез и рецикл синаптических везикул и их содержимого
А - Биосинтез нейротрансмиттеров.
I. Везикулы,
Синтез и рецикл синаптических везикул и их содержимого
А - Биосинтез нейротрансмиттеров.
I. Везикулы,

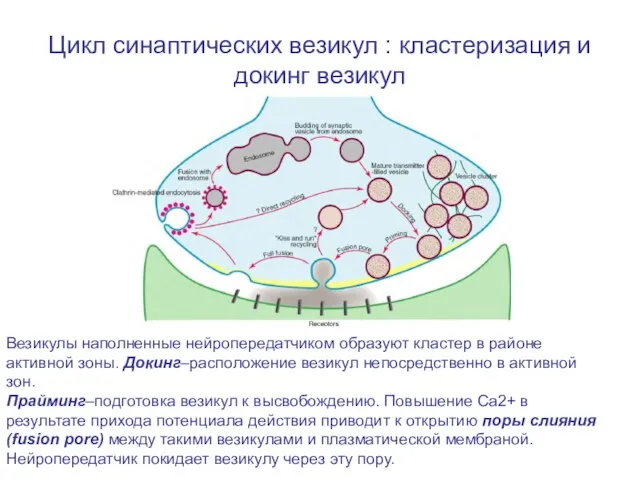
Слайд 18Цикл синаптических везикул : кластеризация и докинг везикул
Везикулы наполненные нейропередатчиком образуют кластер
Цикл синаптических везикул : кластеризация и докинг везикул
Везикулы наполненные нейропередатчиком образуют кластер
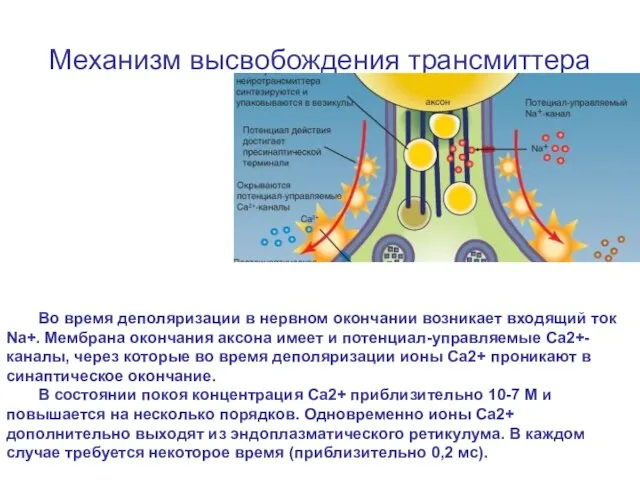
Слайд 19Механизм высвобождения трансмиттера
Во время деполяризации в нервном окончании возникает входящий ток Na+.
Механизм высвобождения трансмиттера
Во время деполяризации в нервном окончании возникает входящий ток Na+.
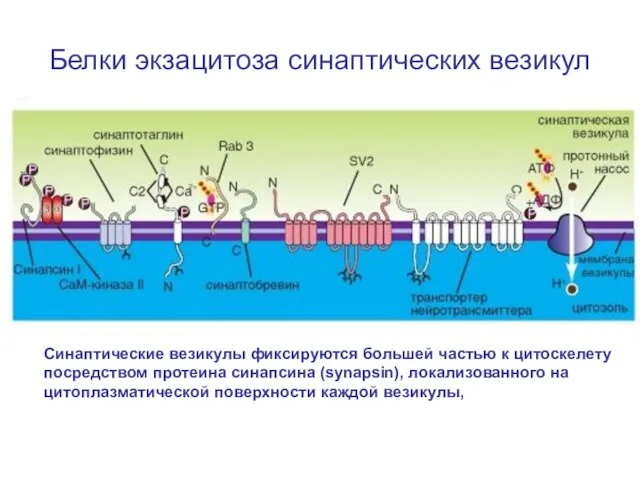
Слайд 20Белки экзацитоза синаптических везикул
Синаптические везикулы фиксируются большей частью к цитоскелету посредством протеина
Белки экзацитоза синаптических везикул
Синаптические везикулы фиксируются большей частью к цитоскелету посредством протеина
Слайд 21Белки экзацитоза синаптических везикул
Синаптические везикулы фиксируются большей частью к цитоскелету посредством протеина
Белки экзацитоза синаптических везикул
Синаптические везикулы фиксируются большей частью к цитоскелету посредством протеина

Слайд 22Synaptotagmin
Synaptotagmin 1 (или synaptotagmin) сенсор ионов Ca2+ датчик в мембране pre-synaptic axon.
Кальций
Synaptotagmin
Synaptotagmin 1 (или synaptotagmin) сенсор ионов Ca2+ датчик в мембране pre-synaptic axon.
Кальций

Слайд 23Neurexin
Neurexin (NRXN) пресинаптическими белок, который участвует в соединении нейронов и с образованием
Neurexin
Neurexin (NRXN) пресинаптическими белок, который участвует в соединении нейронов и с образованием

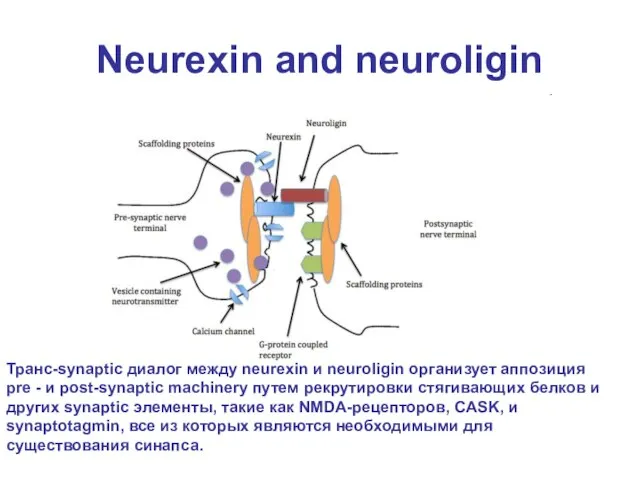
Слайд 24Neurexin and neuroligin
Транс-synaptic диалог между neurexin и neuroligin организует аппозиция pre -
Neurexin and neuroligin
Транс-synaptic диалог между neurexin и neuroligin организует аппозиция pre -
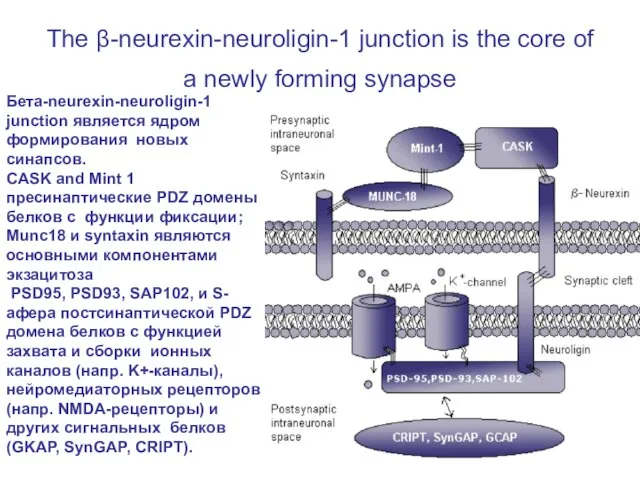
Слайд 25The β-neurexin-neuroligin-1 junction is the core of a newly forming synapse
Бета-neurexin-neuroligin-1
The β-neurexin-neuroligin-1 junction is the core of a newly forming synapse
Бета-neurexin-neuroligin-1
Слайд 26Munc-18
Munc-18 (аббревиатура от mammalian uncoordinated-18) - белки являются гомологом у млекопитающих unc-18
Munc-18
Munc-18 (аббревиатура от mammalian uncoordinated-18) - белки являются гомологом у млекопитающих unc-18

Слайд 27Синаптофизин
Синаптофизин – это гликопротеин, находящийся в пресинаптических везикулах нейронов мозга, спинного мозга,
Синаптофизин
Синаптофизин – это гликопротеин, находящийся в пресинаптических везикулах нейронов мозга, спинного мозга,

Слайд 28Синаптобревин
Синаптобревин (англ. synaptobrevin) — небольшой трансмембранный белок секреторных везикул;
- компонент
Синаптобревин
Синаптобревин (англ. synaptobrevin) — небольшой трансмембранный белок секреторных везикул;
- компонент

Слайд 29Rab-3A
Ras-связанный белок Rab-3А есть белок в организме человека, который кодируется геном RAB3A.[1][2][3]
Rab-3A
Ras-связанный белок Rab-3А есть белок в организме человека, который кодируется геном RAB3A.[1][2][3]

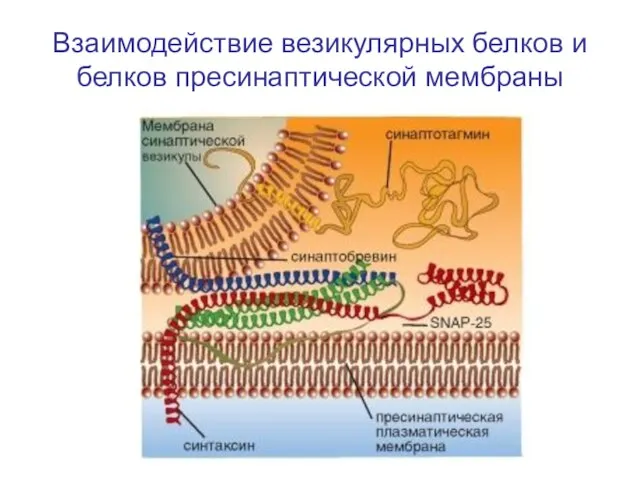
Слайд 30Взаимодействие везикулярных белков и белков пресинаптической мембраны
Взаимодействие везикулярных белков и белков пресинаптической мембраны
Слайд 31Белки экзацитоза синаптических везикул
Меньшая часть везикул также связана с внутренней стороной пресинаптической
Белки экзацитоза синаптических везикул
Меньшая часть везикул также связана с внутренней стороной пресинаптической

Слайд 32Роль ионов кальция
Если потенциал действия достиг пресинаптической области, и в пресинаптическом
Роль ионов кальция
Если потенциал действия достиг пресинаптической области, и в пресинаптическом

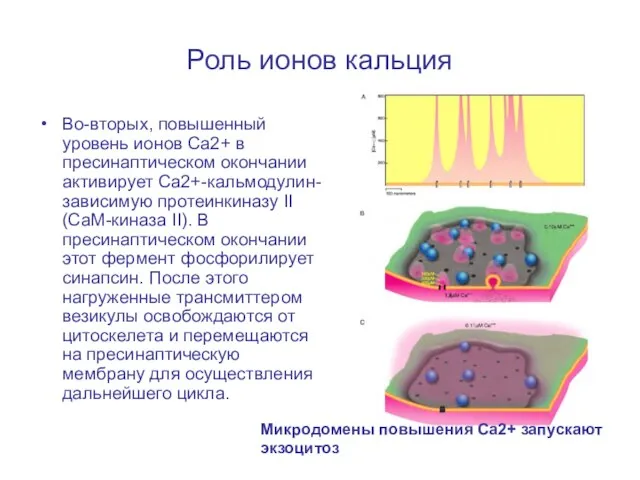
Слайд 33Роль ионов кальция
Во-вторых, повышенный уровень ионов Ca2+ в пресинаптическом окончании активирует Ca2+-кальмодулин-зависимую
Роль ионов кальция
Во-вторых, повышенный уровень ионов Ca2+ в пресинаптическом окончании активирует Ca2+-кальмодулин-зависимую
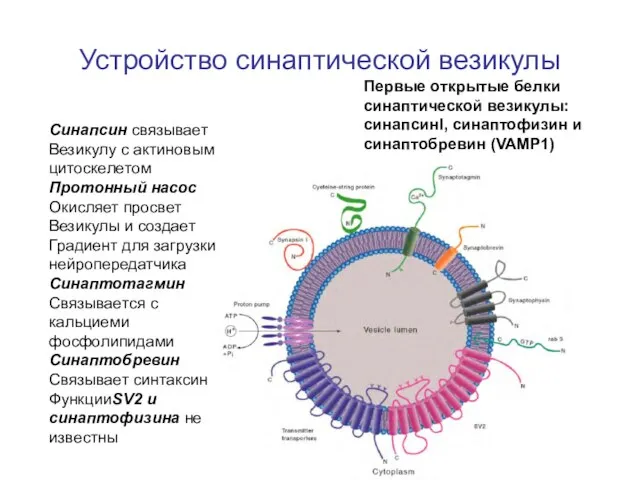
Слайд 34Устройство синаптической везикулы
Первые открытые белки синаптической везикулы:
синапсинI, синаптофизин и синаптобревин (VAMP1)
Синапсин связывает
Везикулу
Устройство синаптической везикулы
Первые открытые белки синаптической везикулы:
синапсинI, синаптофизин и синаптобревин (VAMP1)
Синапсин связывает
Везикулу
Слайд 35SNARE
SNARE– главный компонент механизма слияния синаптической везикулы с мембраной.
•Состоит из 3
SNARE
SNARE– главный компонент механизма слияния синаптической везикулы с мембраной.
•Состоит из 3

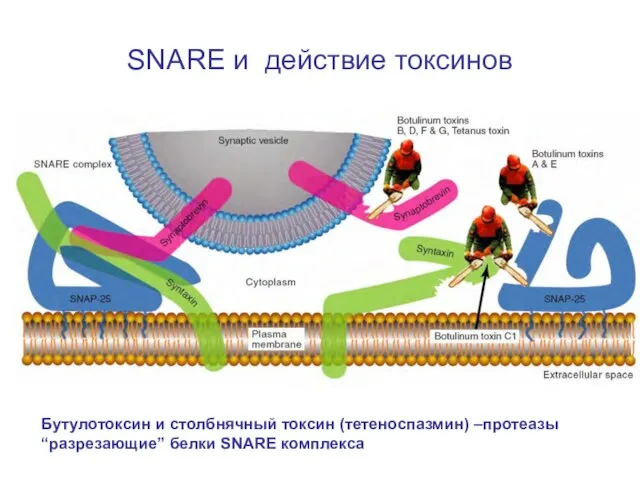
Слайд 36SNARE и действие токсинов
Бутулотоксин и столбнячный токсин (тетеноспазмин) –протеазы “разрезающие” белки SNARE
SNARE и действие токсинов
Бутулотоксин и столбнячный токсин (тетеноспазмин) –протеазы “разрезающие” белки SNARE
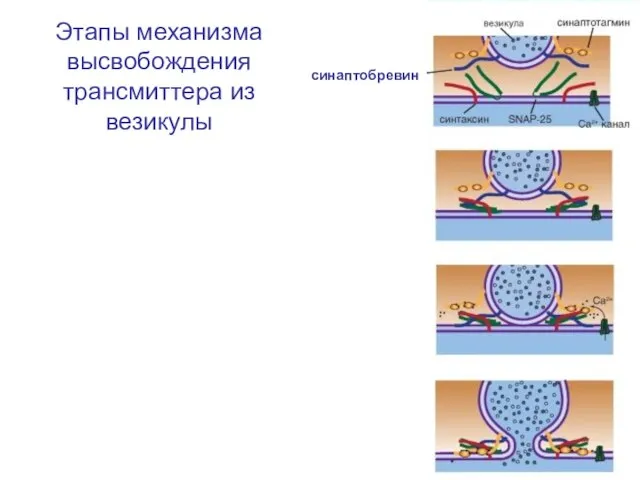
Слайд 37Этапы механизма высвобождения трансмиттера из везикулы
синаптобревин
Этапы механизма высвобождения трансмиттера из везикулы
синаптобревин
Слайд 38Слияние везикулы
Слияние везикул с пресинаптической мембраной управляется белками SNARE, в результате чего
Слияние везикулы
Слияние везикул с пресинаптической мембраной управляется белками SNARE, в результате чего

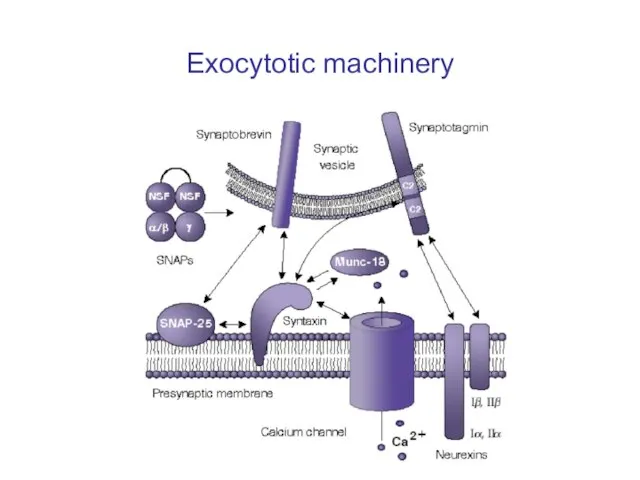
Слайд 39Exocytotic machinery
Exocytotic machinery
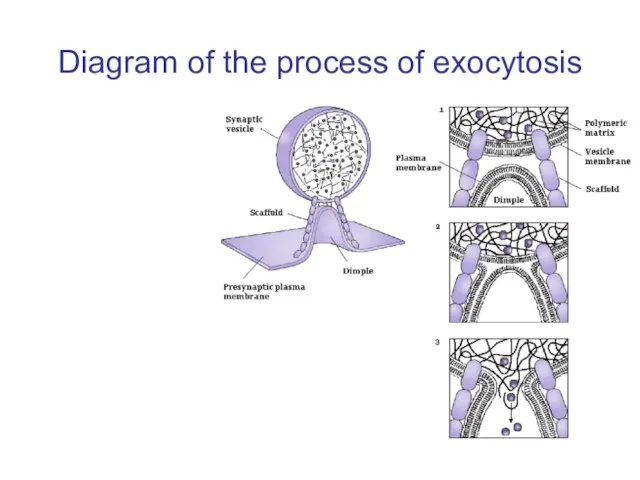
Слайд 40Diagram of the process of exocytosis
Diagram of the process of exocytosis
Слайд 41Цикл синаптических везикул : : возвращение в высвобождаемый пул везикул
1. Простое закрытие
Цикл синаптических везикул : : возвращение в высвобождаемый пул везикул
1. Простое закрытие

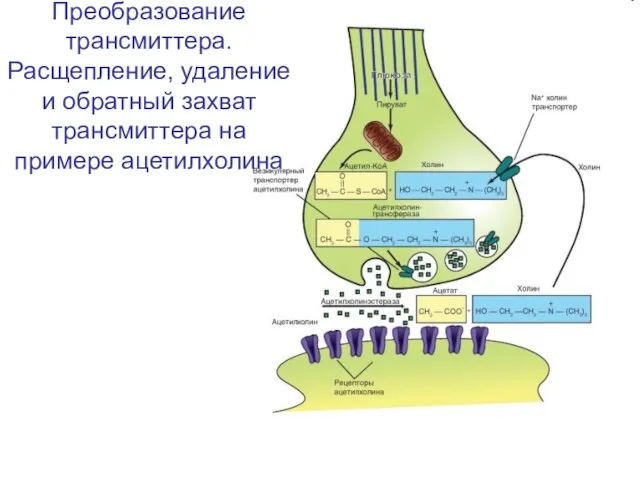
Слайд 42Преобразование трансмиттера.
Расщепление, удаление и обратный захват трансмиттера на примере ацетилхолина
Преобразование трансмиттера.
Расщепление, удаление и обратный захват трансмиттера на примере ацетилхолина
Слайд 43Типы химических синапсов
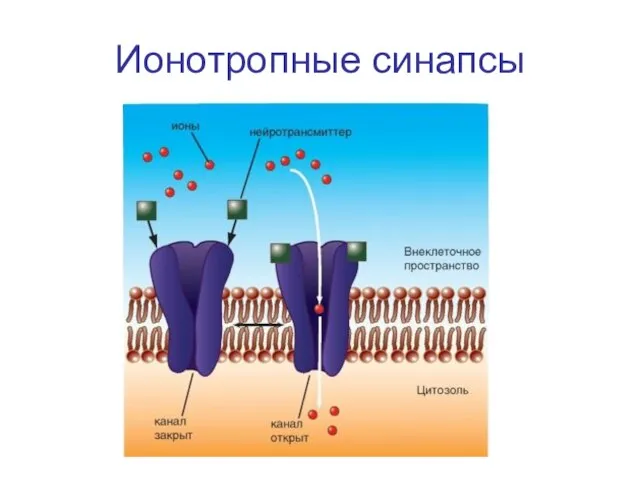
Синапс с ионотропным рецептором - связывание с молекулой трансмиттера, открывает
Типы химических синапсов
Синапс с ионотропным рецептором - связывание с молекулой трансмиттера, открывает

Слайд 44Перечень соединений, влияющие на никотиновые, мускариновые и холинергические синапсы
Перечень соединений, влияющие на никотиновые, мускариновые и холинергические синапсы

Слайд 45Ионотропные синапсы
Ионотропные синапсы
Слайд 46Метаботропные синапсы
Метаботропные синапсы
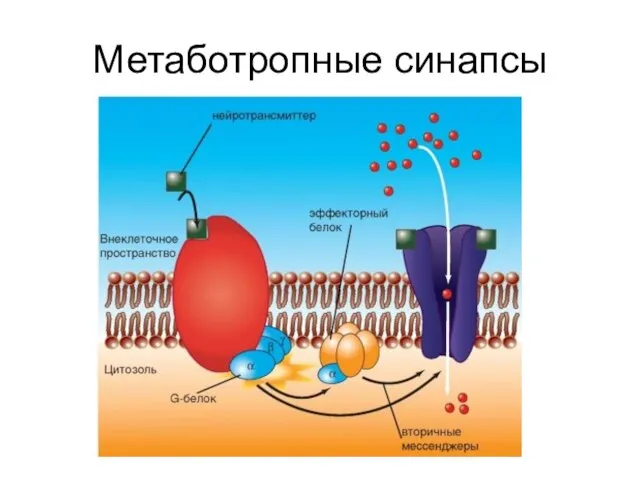
Слайд 47Механизмы работы ионотропного синапса
Холин
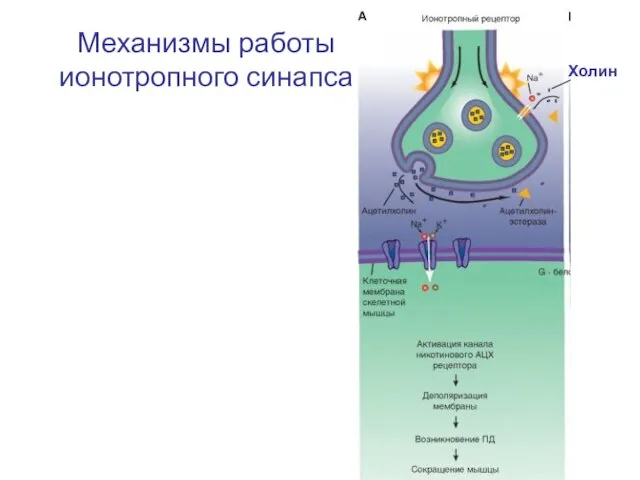
Механизмы работы ионотропного синапса
Холин
Слайд 48Механизмы работы метаботропного синапса
Белок G
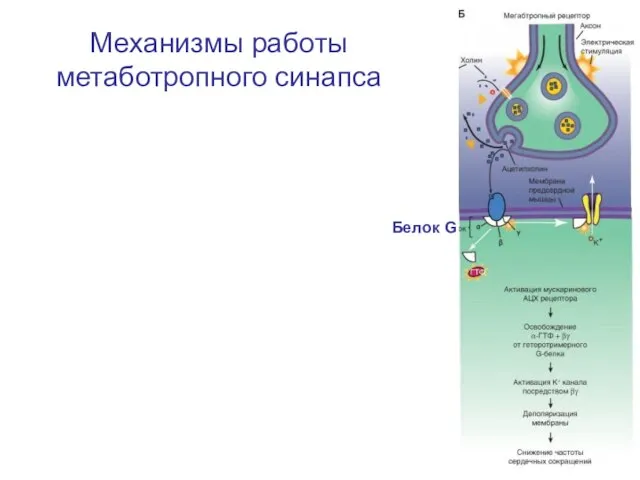
Механизмы работы метаботропного синапса
Белок G
Слайд 49Типы трансмиттеров и механизмы их действия
Ацетилхолин – медиатор всех двигательных нейронов, многих
Типы трансмиттеров и механизмы их действия
Ацетилхолин – медиатор всех двигательных нейронов, многих

Слайд 50Типы трансмиттеров и механизмы их действия
Глицин служит трансмиттером тормозных синапсов и действует
Типы трансмиттеров и механизмы их действия
Глицин служит трансмиттером тормозных синапсов и действует

Слайд 51Типы трансмиттеров и механизмы их действия
γ-Аминомасляную кислоту выбрасывают в качестве трансмиттера многие
Типы трансмиттеров и механизмы их действия
γ-Аминомасляную кислоту выбрасывают в качестве трансмиттера многие

Слайд 52Типы трансмиттеров и механизмы их действия
Серотонин (5-hydroxytryptamin) - биогенный амин, широко распространенный
Типы трансмиттеров и механизмы их действия
Серотонин (5-hydroxytryptamin) - биогенный амин, широко распространенный

Слайд 53Типы трансмиттеров и механизмы их действия
Гистамин - моноамин, выступающий в качестве трансмиттера.
Типы трансмиттеров и механизмы их действия
Гистамин - моноамин, выступающий в качестве трансмиттера.

Слайд 54Типы трансмиттеров и механизмы их действия
Дофамин - биогенный амин, наиболее представленный в
Типы трансмиттеров и механизмы их действия
Дофамин - биогенный амин, наиболее представленный в

Слайд 55Типы трансмиттеров и механизмы их действия
Норадреналин катехоламин, выполняющий функцию трансмиттера в ЦНС
Типы трансмиттеров и механизмы их действия
Норадреналин катехоламин, выполняющий функцию трансмиттера в ЦНС

Слайд 56Типы трансмиттеров и механизмы их действия
Олигопептиды, которые действуют как трансмиттеры или нейромодуляторы
Типы трансмиттеров и механизмы их действия
Олигопептиды, которые действуют как трансмиттеры или нейромодуляторы

Слайд 57Типы трансмиттеров и механизмы их действия
Другие нейропептиды: вещество Р, ангиотензин II, соматостатин,
Типы трансмиттеров и механизмы их действия
Другие нейропептиды: вещество Р, ангиотензин II, соматостатин,

Слайд 58Примеры механизмов действия трансмиттеров на различные типы рецепторов
Примеры механизмов действия трансмиттеров на различные типы рецепторов

Слайд 59Ионотропный никотиновый холинергический синапс/Структура n-холинорецептора
Ионотропный никотиновый холинергический синапс/Структура n-холинорецептора
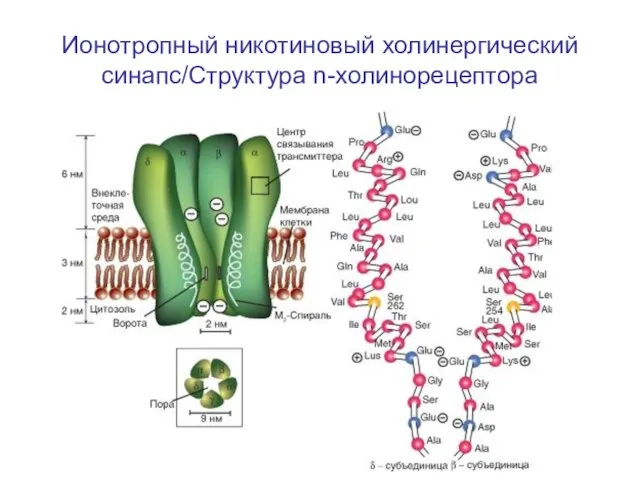
Слайд 60Структура n-холинорецептора
Рецепторный белок для нАЦХ-рецептора образует пору - ионный канал т пронизывает
Структура n-холинорецептора
Рецепторный белок для нАЦХ-рецептора образует пору - ионный канал т пронизывает
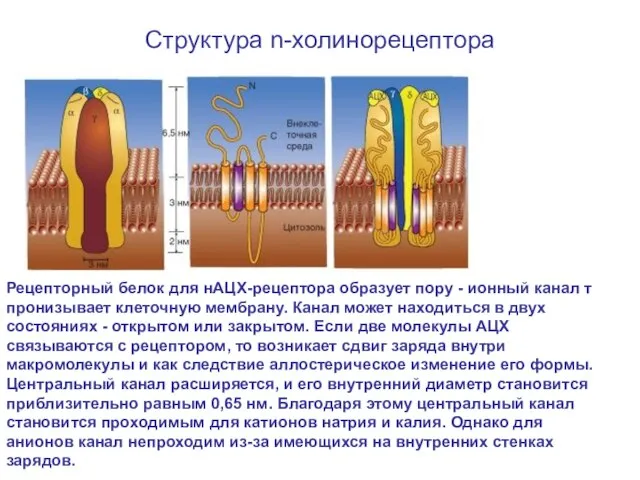
Слайд 61Глутаматный синапс
Лиганд-управляемые катионные каналы - NMDA (N-метил-D-аспартат) и
AMPA (α-ами- но-3-гидрокси-5-метил-4-изоксазол-пропионовая кислота)
Глутаматный синапс
Лиганд-управляемые катионные каналы - NMDA (N-метил-D-аспартат) и
AMPA (α-ами- но-3-гидрокси-5-метил-4-изоксазол-пропионовая кислота)
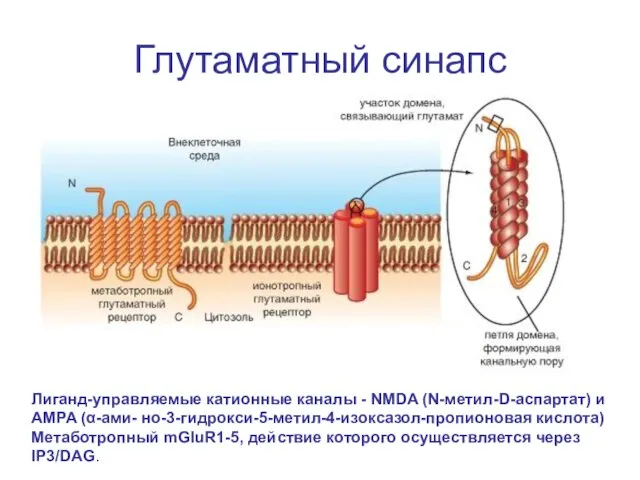
Слайд 62Активация глутаматом два ионотропный лиганд-управляемый рецептор, служащий катионным каналом AMPA, и метаботропный
Активация глутаматом два ионотропный лиганд-управляемый рецептор, служащий катионным каналом AMPA, и метаботропный
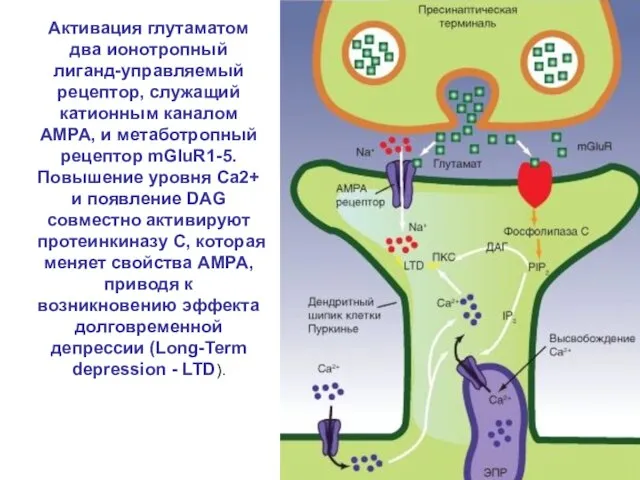
Слайд 63Фосфорилирование и дефосфорилирование в глутаматном синапсе
Глутаматный синапс содержит на постсинаптической мембране два
Фосфорилирование и дефосфорилирование в глутаматном синапсе
Глутаматный синапс содержит на постсинаптической мембране два

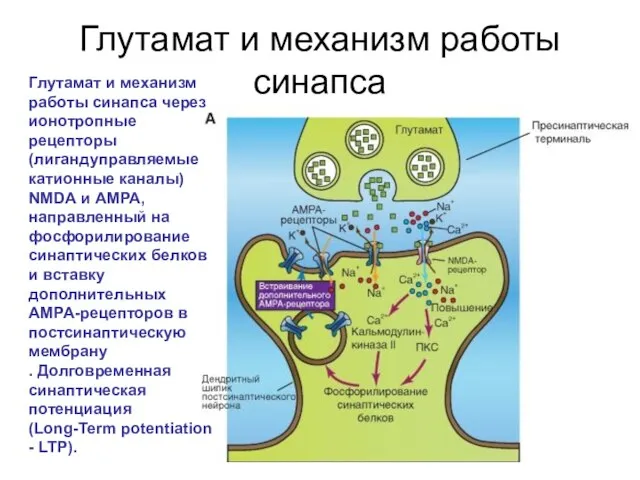
Слайд 64Глутамат и механизм работы синапса
Глутамат и механизм работы синапса через ионотропные рецепторы
Глутамат и механизм работы синапса
Глутамат и механизм работы синапса через ионотропные рецепторы
Слайд 65Глутамат и механизм работы синапса
Глутамат и механизм работы синапса через ионотропные рецепторы
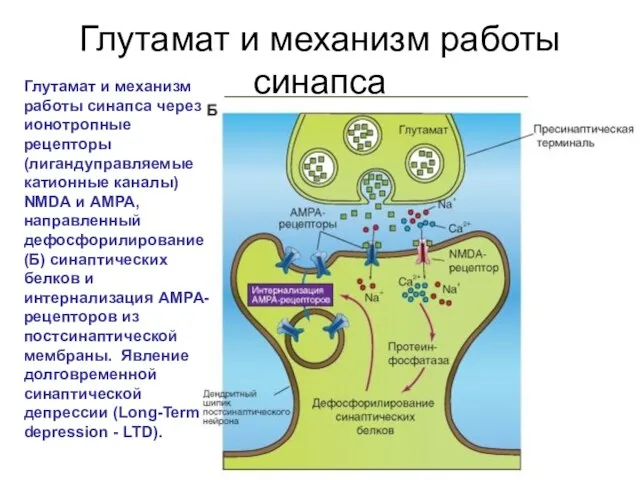
Глутамат и механизм работы синапса
Глутамат и механизм работы синапса через ионотропные рецепторы
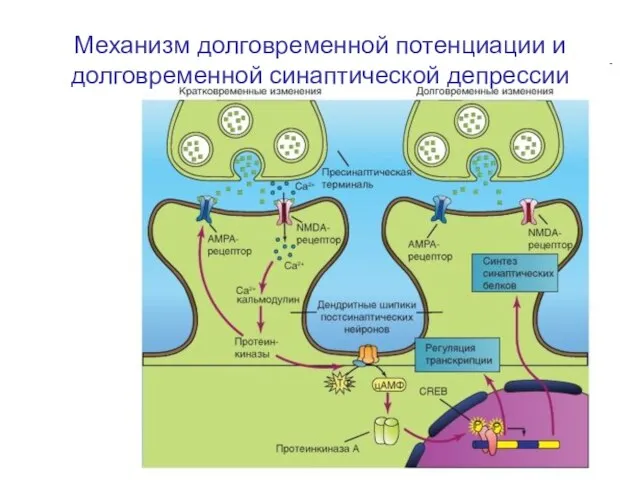
Слайд 66Механизм долговременной потенциации и долговременной синаптической депрессии
Механизм долговременной потенциации и долговременной синаптической депрессии
Слайд 67Механизм долговременной потенциации и долговременной синаптической депрессии
Освобожденный из пресинаптической области глутамат активирует
Механизм долговременной потенциации и долговременной синаптической депрессии
Освобожденный из пресинаптической области глутамат активирует

Слайд 68Потенциалы постсинаптической мембране глутаматного синапса,связанные с функцией лигандуправляемых катионных каналов - NMDA
Потенциалы постсинаптической мембране глутаматного синапса,связанные с функцией лигандуправляемых катионных каналов - NMDA
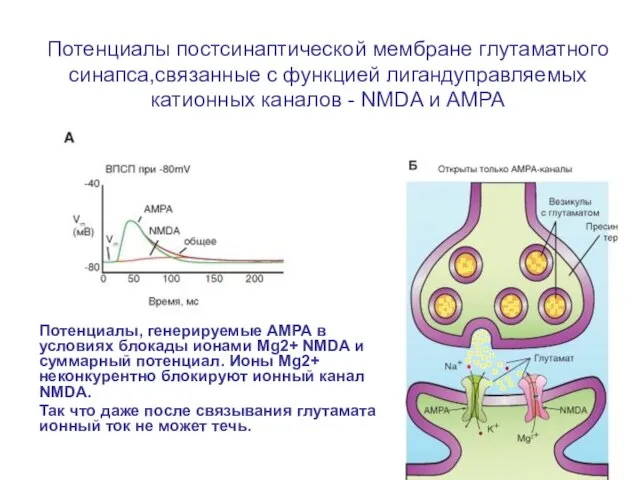
Слайд 69Потенциалы постсинаптической мембране глутаматного синапса,связанные с функцией лигандуправляемых катионных каналов - NMDA
Потенциалы постсинаптической мембране глутаматного синапса,связанные с функцией лигандуправляемых катионных каналов - NMDA
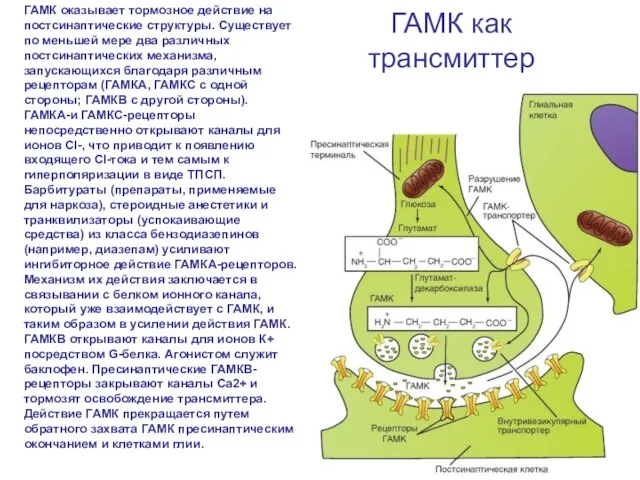
Слайд 70ГАМК как трансмиттер
ГАМК оказывает тормозное действие на постсинаптические структуры. Существует по меньшей
ГАМК как трансмиттер
ГАМК оказывает тормозное действие на постсинаптические структуры. Существует по меньшей
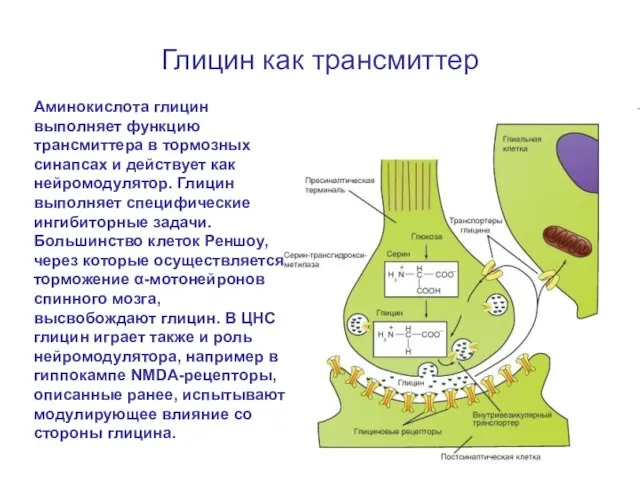
Слайд 71Глицин как трансмиттер
Аминокислота глицин выполняет функцию трансмиттера в тормозных синапсах и действует
Глицин как трансмиттер
Аминокислота глицин выполняет функцию трансмиттера в тормозных синапсах и действует
Слайд 72NO как трансмиттер
Оксид азота (NO) - это паракринный медиатор, выделяемый эндотелиальными клетками
NO как трансмиттер
Оксид азота (NO) - это паракринный медиатор, выделяемый эндотелиальными клетками
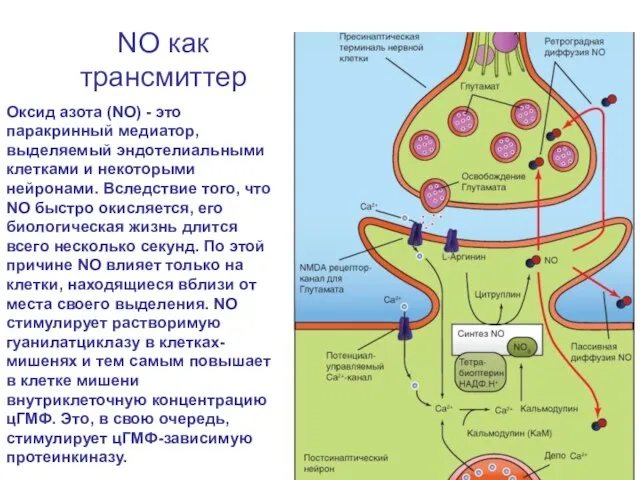
Слайд 73NO как трансмиттер
Образование NO катализируется NO-синтазой NOS - Са2+-кальмодулин-зависимым ферментом, ускоряющим превращение
NO как трансмиттер
Образование NO катализируется NO-синтазой NOS - Са2+-кальмодулин-зависимым ферментом, ускоряющим превращение

Слайд 74Постсинаптическая часть
Постсинаптическая часть различается в зависимости от:
1.Типа пресинаптического нейрона (высвобождаемого нейропередатчика)
2.Типа
Постсинаптическая часть
Постсинаптическая часть различается в зависимости от:
1.Типа пресинаптического нейрона (высвобождаемого нейропередатчика)
2.Типа

Слайд 75Классификация постсинапса
По наличию или отсутствию шипика –синапсы шипиковые и сидячие
По наличию
Классификация постсинапса
По наличию или отсутствию шипика –синапсы шипиковые и сидячие
По наличию

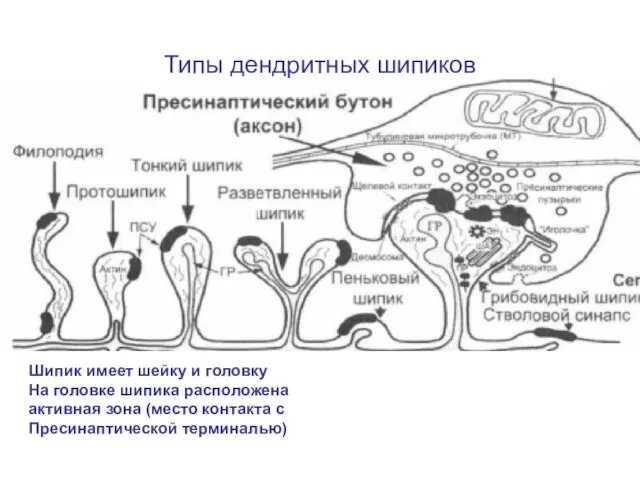
Слайд 76Типы дендритных шипиков
Шипик имеет шейку и головку
На головке шипика расположена активная
Типы дендритных шипиков
Шипик имеет шейку и головку
На головке шипика расположена активная
Слайд 77Свойства дендритных шипиков
Могут изменять свою геометрию (ширину, длину шейки,
Диаметр головки)
Свойства дендритных шипиков
Могут изменять свою геометрию (ширину, длину шейки,
Диаметр головки)

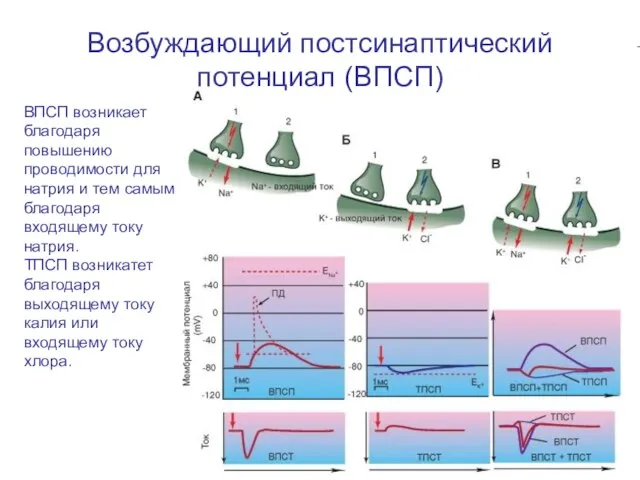
Слайд 78Возбуждающий постсинаптический потенциал (ВПСП)
ВПСП возникает благодаря повышению проводимости для натрия и тем
Возбуждающий постсинаптический потенциал (ВПСП)
ВПСП возникает благодаря повышению проводимости для натрия и тем
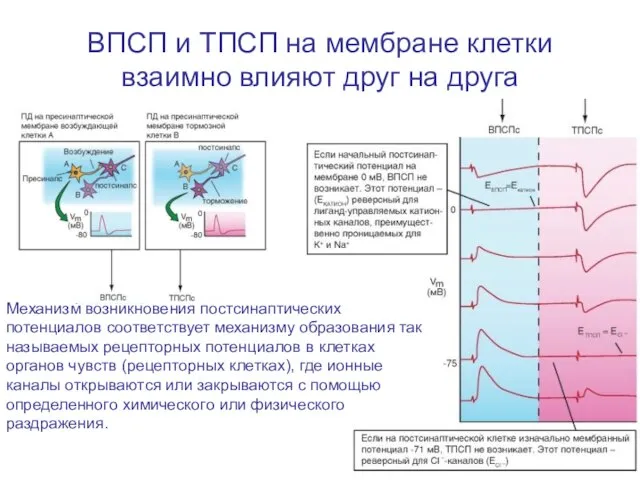
Слайд 79ВПСП и ТПСП на мембране клетки взаимно влияют друг на друга
Механизм возникновения
ВПСП и ТПСП на мембране клетки взаимно влияют друг на друга
Механизм возникновения
Слайд 80Соединения с синаптическим влиянием
Наряду с трансмиттером, другие соединения тоже могут влиять на
Соединения с синаптическим влиянием
Наряду с трансмиттером, другие соединения тоже могут влиять на

Слайд 81Свойства основных нейротрансмиттеров
Свойства основных нейротрансмиттеров
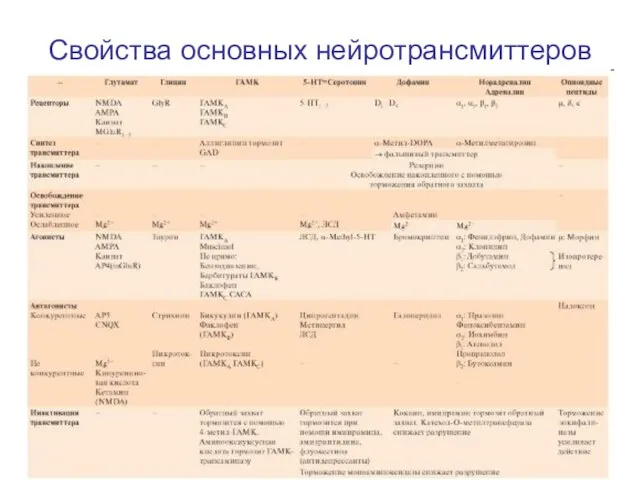
Слайд 82Свойства основных нейротрансмиттеров
Свойства основных нейротрансмиттеров
 Интерактивная игра Моя Россия
Интерактивная игра Моя Россия Презентация на тему Кавказский заповедник
Презентация на тему Кавказский заповедник Один день из жизни Е.Онегина
Один день из жизни Е.Онегина Презентация на тему Личность и интернет
Презентация на тему Личность и интернет  Профком ОГАУ
Профком ОГАУ Tsifrovye_servisy_i_instrumenty_dlya_sozdania_obuchayuschikh_video
Tsifrovye_servisy_i_instrumenty_dlya_sozdania_obuchayuschikh_video Изложение2 класс
Изложение2 класс Океаны и их обитатели
Океаны и их обитатели Маленькие герои большой войны
Маленькие герои большой войны Лекция 1
Лекция 1 Профессия 220301 Автоматизация технологических процессов и производств (по отраслям)
Профессия 220301 Автоматизация технологических процессов и производств (по отраслям) Презентация на тему Публичное выступление
Презентация на тему Публичное выступление Мониторинг как средство оценки результатов обучения
Мониторинг как средство оценки результатов обучения ОПЫТ РАЗРАБОТКИ ДОКУМЕНТАЦИИ СЗПДн. МОДЕЛЬ УГРОЗ БЕЗОПАСНОСТИ ПДн.
ОПЫТ РАЗРАБОТКИ ДОКУМЕНТАЦИИ СЗПДн. МОДЕЛЬ УГРОЗ БЕЗОПАСНОСТИ ПДн. Особенности строения земноводных
Особенности строения земноводных Сложение чисел с разными знаками
Сложение чисел с разными знаками Biznesa plāns. Sabiedrība ar ierobežotu atbildību SIA TAPTI
Biznesa plāns. Sabiedrība ar ierobežotu atbildību SIA TAPTI self promo
self promo Ревизия (от позднелат. revisio пересмотр) так называлась в России 1819 вв. перепись населения.
Ревизия (от позднелат. revisio пересмотр) так называлась в России 1819 вв. перепись населения. Федоскинская лаковая миниатюра
Федоскинская лаковая миниатюра Возвышение Рима
Возвышение Рима П.Я.Бакланов Природопользование в стратегии развития Дальнего Востока России Хабаровск, 2011
П.Я.Бакланов Природопользование в стратегии развития Дальнего Востока России Хабаровск, 2011 кровяные инфекции (сыпной и возвратный малярия и др.)
кровяные инфекции (сыпной и возвратный малярия и др.) Патрина Наталья Николаевна МОУ «Лицей № 6» г. Миасс
Патрина Наталья Николаевна МОУ «Лицей № 6» г. Миасс Учимся считать семейные доходы
Учимся считать семейные доходы Музеи Азии
Музеи Азии Мое хобби - иностранные языки. Практическая работа
Мое хобби - иностранные языки. Практическая работа Придаточные предложения
Придаточные предложения