- ПРИРОДА ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ

Содержание
- 2. 1. Введение Гипотеза «1 ген – 1 фермент», обоснованная в 40-х годах, допускала два возможных объяснения
- 3. 1. Гены и белки Взаимосвязь между генами и строением белков впервые была показана при изучении одной
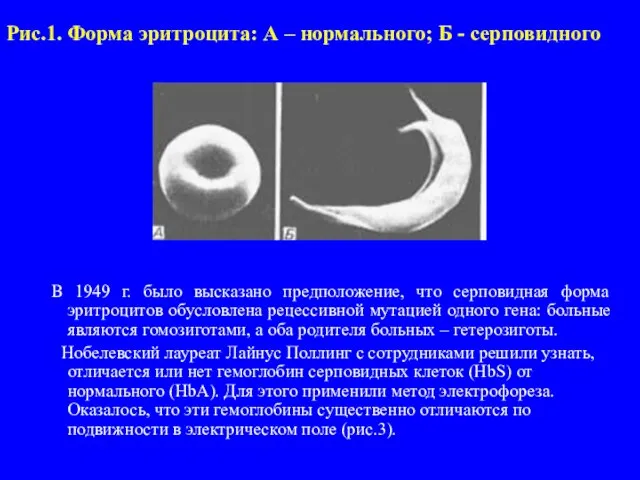
- 4. Рис.1. Форма эритроцита: А – нормального; Б - серповидного В 1949 г. было высказано предположение, что
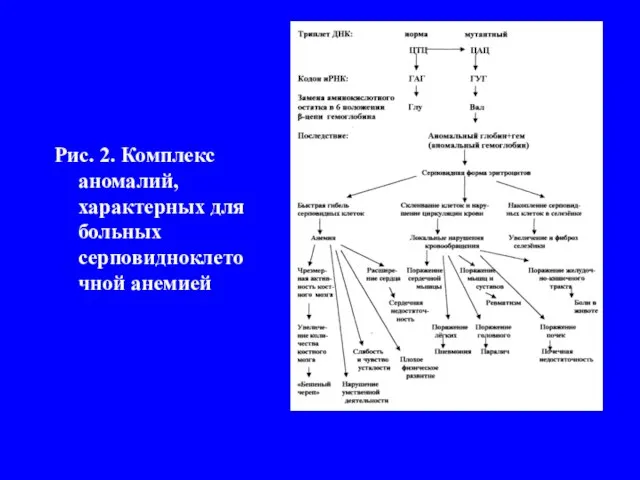
- 5. Рис. 2. Комплекс аномалий, характерных для больных серповидноклеточной анемией
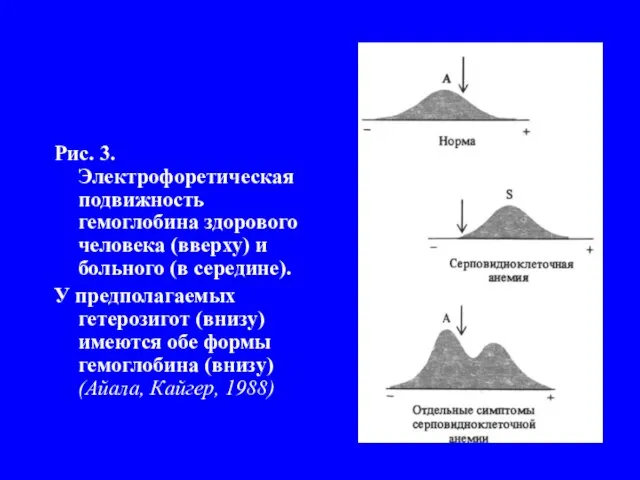
- 6. Рис. 3. Электрофоретическая подвижность гемоглобина здорового человека (вверху) и больного (в середине). У предполагаемых гетерозигот (внизу)
- 7. Очевидно, мутация, вызывающая серповидно-клеточность, связана с изменениями белка – гемоглобина. У гомозигот в эритроцитах имеются только
- 8. В 1957 году предположение Л. Поллинга было экспериментально доказано Верноном Ингрэмом. Он определил, что в шестом
- 9. Таким образом, стало очевидно, что генетическая информация – это информация о последовательности аминокислотных остатков в полипептидных
- 10. Многие белки состоят из двух и более различающихся полипептидных цепей, каждая из которых кодируется своим геном.
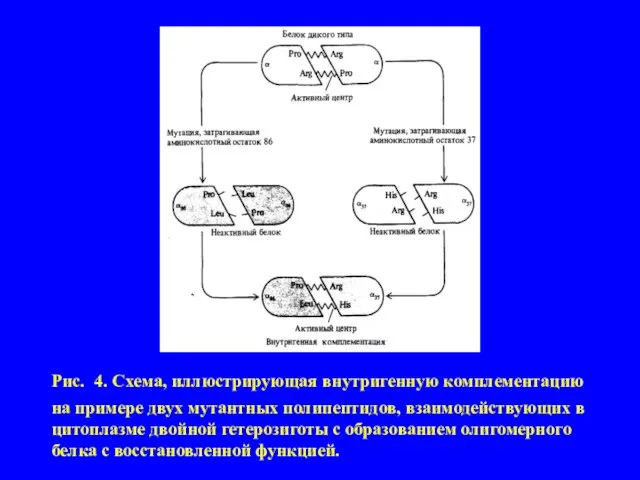
- 11. 2. Внутригенная комплементация Многие белки состоят из двух или нескольких идентичных полипептид-ных цепей, образующих функционально активную
- 12. Рис. 4. Схема, иллюстрирующая внутригенную комплементацию на примере двух мутантных полипептидов, взаимодействующих в цитоплазме двойной гетерозиготы
- 14. Скачать презентацию

Слайд 31. Гены и белки
Взаимосвязь между генами и строением белков впервые
1. Гены и белки
Взаимосвязь между генами и строением белков впервые

Слайд 4Рис.1. Форма эритроцита: А – нормального; Б - серповидного
В
Рис.1. Форма эритроцита: А – нормального; Б - серповидного
В
Слайд 5Рис. 2. Комплекс аномалий, характерных для больных серповидноклеточной анемией
Рис. 2. Комплекс аномалий, характерных для больных серповидноклеточной анемией
Слайд 6Рис. 3. Электрофоретическая подвижность гемоглобина здорового человека (вверху) и больного (в середине).
Рис. 3. Электрофоретическая подвижность гемоглобина здорового человека (вверху) и больного (в середине).
Слайд 7 Очевидно, мутация, вызывающая серповидно-клеточность, связана с изменениями белка – гемоглобина. У
Очевидно, мутация, вызывающая серповидно-клеточность, связана с изменениями белка – гемоглобина. У

Слайд 8 В 1957 году предположение Л. Поллинга было экспериментально доказано Верноном Ингрэмом.
В 1957 году предположение Л. Поллинга было экспериментально доказано Верноном Ингрэмом.

Слайд 9Таким образом, стало очевидно, что генетическая информация – это информация о последовательности
Таким образом, стало очевидно, что генетическая информация – это информация о последовательности

Слайд 10 Многие белки состоят из двух и более различающихся полипептидных цепей, каждая
Многие белки состоят из двух и более различающихся полипептидных цепей, каждая

Слайд 112. Внутригенная комплементация
Многие белки состоят из двух или нескольких идентичных
2. Внутригенная комплементация
Многие белки состоят из двух или нескольких идентичных

Слайд 12Рис. 4. Схема, иллюстрирующая внутригенную комплементацию на примере двух мутантных полипептидов, взаимодействующих
Рис. 4. Схема, иллюстрирующая внутригенную комплементацию на примере двух мутантных полипептидов, взаимодействующих
 Современное искусство
Современное искусство Исследование и анализ ошибок при определении стоимости в оценочной деятельности
Исследование и анализ ошибок при определении стоимости в оценочной деятельности Ценность рода и семьи
Ценность рода и семьи Активная и пассивная безопасность автомобиля
Активная и пассивная безопасность автомобиля Ценообразование в логистических системах
Ценообразование в логистических системах Диагностика финансового состояния организации по критериям банкротства
Диагностика финансового состояния организации по критериям банкротства Лучевая семиотика заболеваний сердца и сосудов
Лучевая семиотика заболеваний сердца и сосудов Право в системе социальных норм
Право в системе социальных норм Презентация на тему Законы Кеплера (11 класс)
Презентация на тему Законы Кеплера (11 класс) Что дарим на праздники? Открытка
Что дарим на праздники? Открытка Гойя Франсиско Хосе де испанский живописец, гравер
Гойя Франсиско Хосе де испанский живописец, гравер Secondary school
Secondary school ФЕДЕРАЛЬНАЯ АНТИМОНОПОЛЬНАЯ СЛУЖБА Анализ результатов мониторинга цен на лекарственные средства по итогам 1 квартала 2010 года Упр
ФЕДЕРАЛЬНАЯ АНТИМОНОПОЛЬНАЯ СЛУЖБА Анализ результатов мониторинга цен на лекарственные средства по итогам 1 квартала 2010 года Упр Успешные кейсы развития бизнеса с помощью интернета Константин Миска 24.11.2011. - презентация
Успешные кейсы развития бизнеса с помощью интернета Константин Миска 24.11.2011. - презентация Работа с концептами как одна из форм реализации лингвокультурологического подхода
Работа с концептами как одна из форм реализации лингвокультурологического подхода ИНФОРМАЦИОНН ОЕОБЕСПЕЧЕНИЕ ИС
ИНФОРМАЦИОНН ОЕОБЕСПЕЧЕНИЕ ИС Методы обеспечения продольной устойчивости самолета в режиме скоростей от 0,85м до 2,5м
Методы обеспечения продольной устойчивости самолета в режиме скоростей от 0,85м до 2,5м План Путина – победа России
План Путина – победа России Цветоведение
Цветоведение ТЕМА: Наглядное представление статистической информации
ТЕМА: Наглядное представление статистической информации 1
1 Microsoft TechDays Константин Трещев MCITP: Enterprise Administrator
Microsoft TechDays Константин Трещев MCITP: Enterprise Administrator Восточнохристианский и исламский миры
Восточнохристианский и исламский миры «На Кубани родной живем единой семьей»
«На Кубани родной живем единой семьей» Шляпы и короны королевы Великобритании
Шляпы и короны королевы Великобритании Новообразования костей лицевого скелета у детей
Новообразования костей лицевого скелета у детей Изобретение радио Александром Степановичем Поповым
Изобретение радио Александром Степановичем Поповым Влияние цвета на настроение человека
Влияние цвета на настроение человека