- Пространственная модель ДНК

Содержание
- 2. Пространственная модель ДНК
- 3. Сахорофосфатные связи в цепи ДНК
- 4. Взаимодействие между цепями ДНК
- 5. Три конформации ДНК – A, B, Z
- 7. Молекула РНК дельта вируса гепатита
- 8. Нуклеосомы
- 9. Модель пространственной организации хроматина
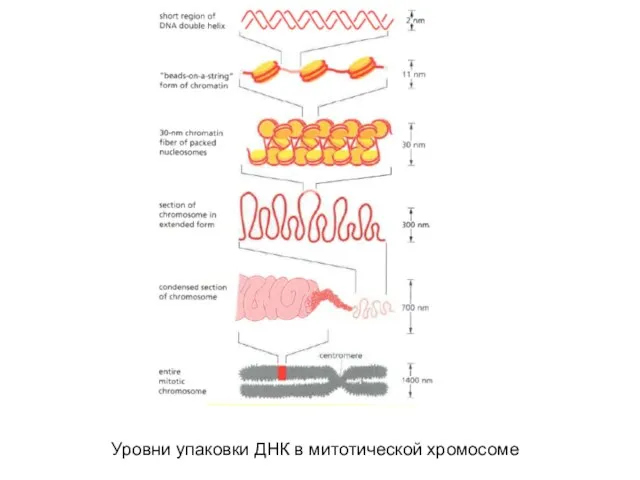
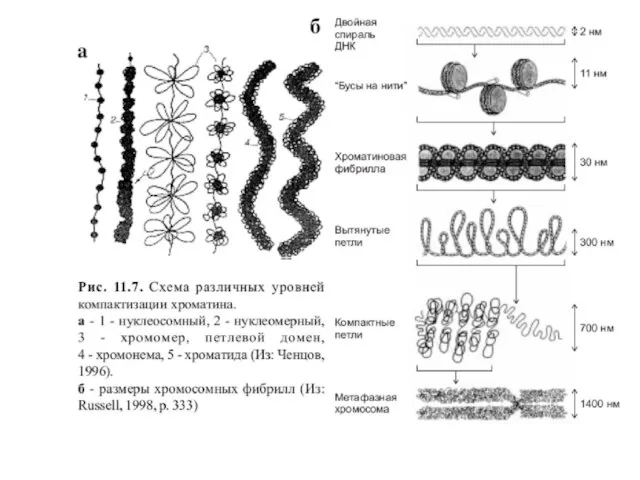
- 10. Уровни упаковки ДНК в митотической хромосоме
- 12. Митотическая хромосома в разрезе
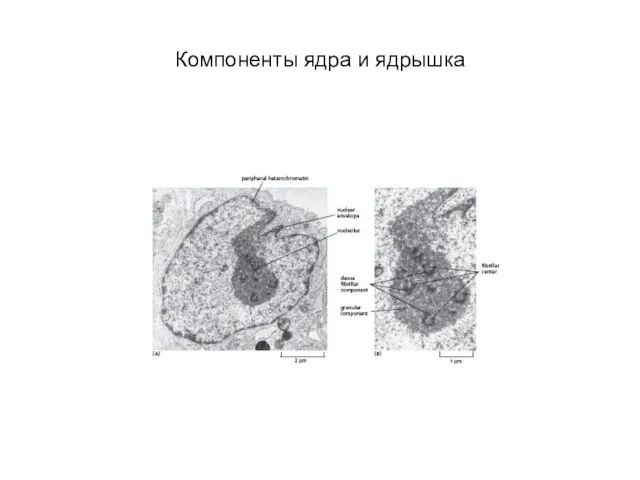
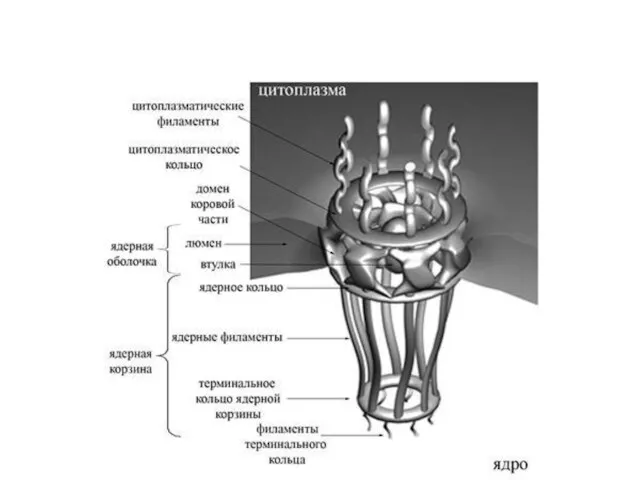
- 13. Компоненты ядра и ядрышка
- 16. Изменение формы ядра и позиционирования хромосомы 13 (зеленая) при прогерии
- 17. Синдром прогерии
- 19. Диплоидный набор хромосом человека
- 20. Кинетохор – место прикрепления микротрубочек к хроматиде
- 21. Хромосомы человека
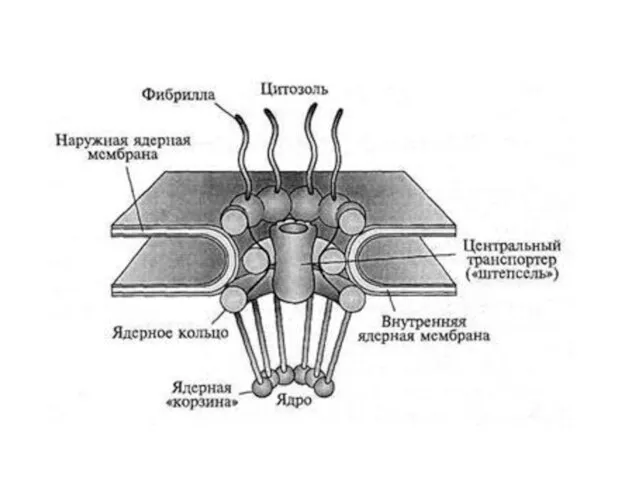
- 22. Перинуклеарное пространство - 20–40 нм
- 25. Ядерная ламина (30-100 нм) представляет собой слой белков под внутренней ядерной мембраной и состоит из белков
- 26. Ядро, окрашенное антителами к ламину В1 и В2
- 27. В состав ядерной ламины входят белки: несприн, эмерин, ламинассоциированные белки 1 и 2, рецептор ламина В
- 28. AP, attachment plate; CE, central element of the SC (synaptonemal complex); NPC, nuclear pore complex; PNS,
- 29. Brian Burke It Takes KASH to Hitch to the SUN Cell Volume 149, Issue 5, 25
- 32. Ядрышко (1-5 нм в диаметре) - место синтеза рибосом на участках хромосом, содержащих гены рибосомальной РНК
- 33. Гранулярный компонент – представляет собой гранулы 15-20 нм и расположен на периферии ядрышка Фибриллярный компонент содержит
- 34. У человека гены, кодирующие рРНК, как и у других организмов, организованы в группы тандемных повторов, расположенных
- 36. Полуконсервативный синтез ДНК
- 37. ДНК-полимераза бактериофага Т7
- 38. ДНК-полимеразы прокариот на примере E. coli
- 39. Схема ДНК-полимеразы ІІІ бактериальной клетки
- 40. Схема репликативной вилки
- 42. Репликационные фабрики в ядре мыши и дрожжей Крупные репликационные фабрики в ядре мыши составлены из нескольких
- 43. Репликационные фабрики в ядрах СНО, активированных в экстракте ооцитов шпорцевой лягушки Xenopus laevis Roscovitine – ингибитор
- 44. Пространственная модель РНК-полимеразы прокариотической клетки
- 45. Пространственная модель РНК-полимеразы II клетки дрожжей (12 субъединиц)
- 46. Синтез иРНК у прокариот
- 47. Промотор для РНК-полимеразы II
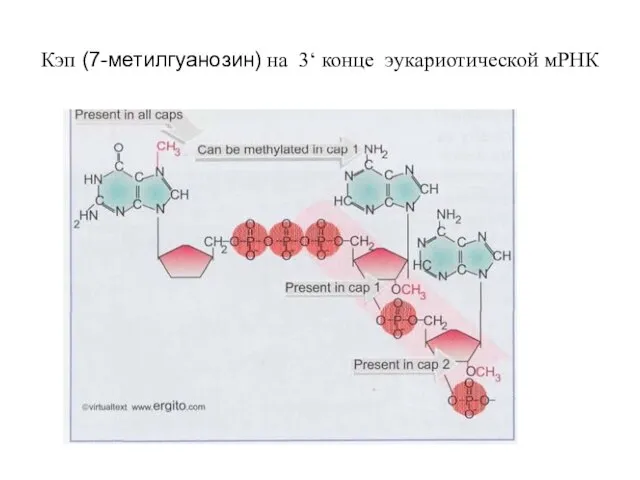
- 48. Кэп (7-метилгуанозин) на 3‘ конце эукариотической мРНК
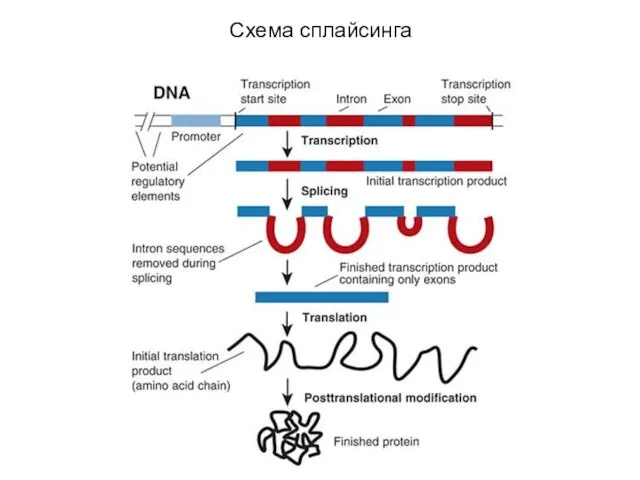
- 49. Схема сплайсинга
- 50. Схема строения транскрипционной фабрики Диаметр фабрики – 56, 75 нм, по данным разных авторов В клетках
- 51. Схема изменений в состоянии фосфорилирования большой субъединицы РНК-полимеразы -II процессе транскрипции Большая субъединица ( RPB1) полимеразы
- 52. Схема транскрипционной фабрики Зеленым цветом отмечены 8 молекул РНК-полимеразы, желтым - РНК
- 54. Две возможные схемы сплайсинга генов, транскрибируемых на двух транскрипционных фабриках с – транскрипция двух генов идет
- 56. Транскрипционные фабрики в клетках HeLa Транскрипционные фабрики указаны стрелочками, интерхроматиновые гранулы (сплайсосомы) - звездочками
- 57. Транскрипционные фабрики в клетках бактерий
- 58. Транскрипция двух генов в эритробластах человека SC35 – фактор сплайсинга
- 59. Транскрипция глобиновых генов в эритробластах человека Красный цвет – РНК-пол II Зеленый –РНК Два глобиновых гена
- 60. Интерхроматиновые гранулы (speckles, 20-25 нм) в клетках HeLa Размер черточки – 5 мкм Гранулы содержат мяРНК
- 61. Возможные функции некодирующих РНК (нкРНК) и малых интерференционных РНК (миРНК) Small interfering RNA (siRNA)
- 62. Схема образования малых ядерных РНК путем разрезания нк РНК
- 63. Регуляция трансляции с помощью микро РНК (miRNA) Предшественники miRNA транскрибируются и подвергаются процессингу РНКазами Drosha и
- 64. Деградация РНК вируса c помощью белка Argonaute-2 РНКаза Dicer-2 разрезает двухцепочечную dsRNA на фрагменты малых интерфенирующих
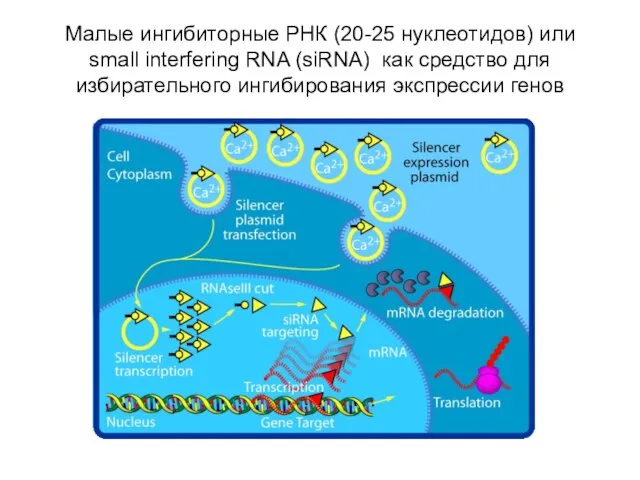
- 65. Малые ингибиторные РНК (20-25 нуклеотидов) или small interfering RNA (siRNA) как средство для избирательного ингибирования экспрессии
- 67. Скачать презентацию
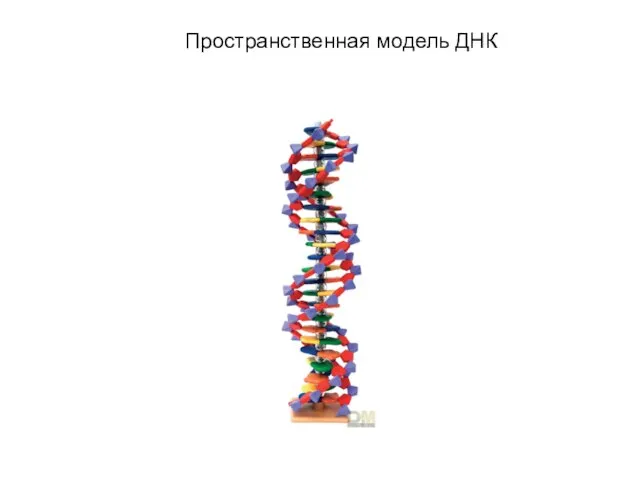
Слайд 3Сахорофосфатные связи в цепи ДНК
Сахорофосфатные связи в цепи ДНК
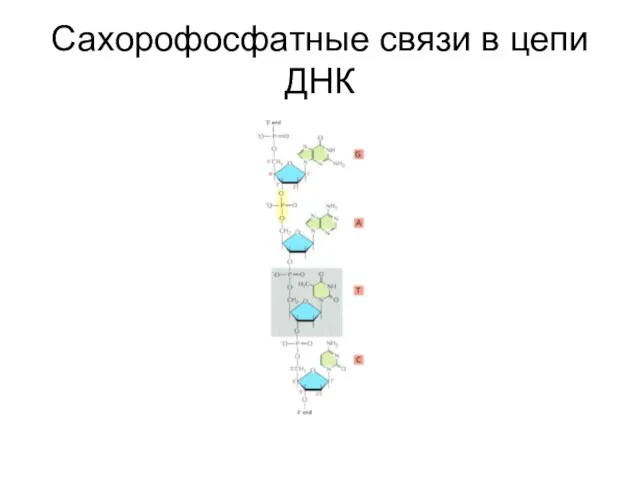
Слайд 4Взаимодействие между цепями ДНК
Взаимодействие между цепями ДНК
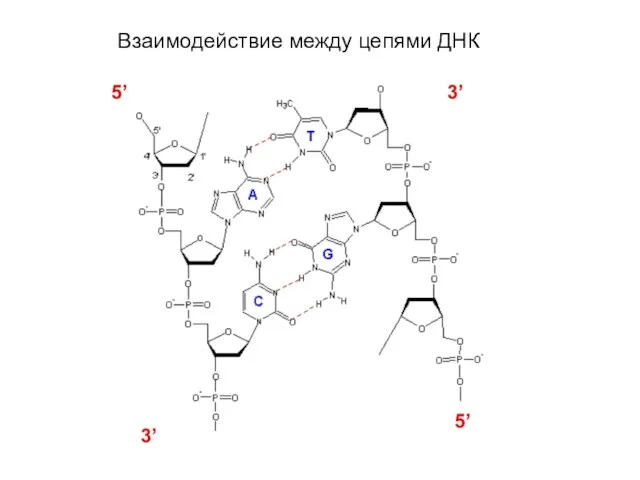
Слайд 5 Три конформации ДНК – A, B, Z
Три конформации ДНК – A, B, Z
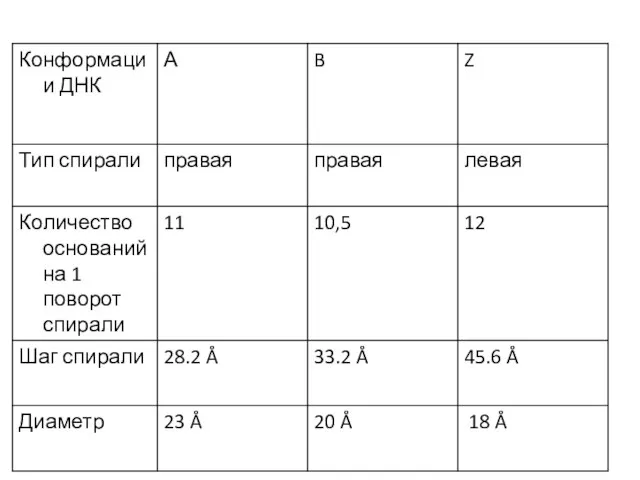
Слайд 7Молекула РНК дельта вируса гепатита
Молекула РНК дельта вируса гепатита
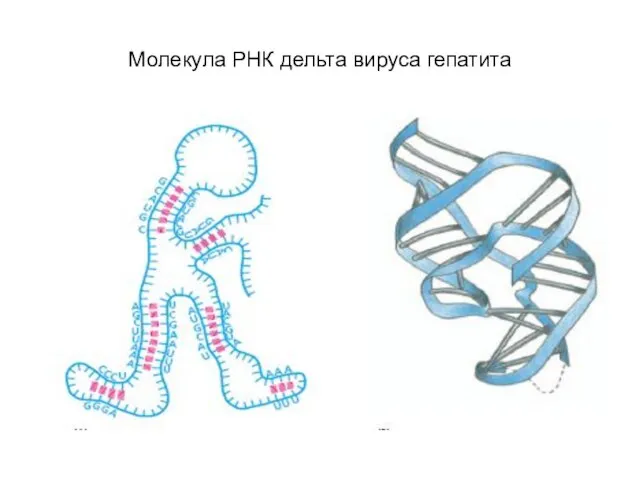
Слайд 8Нуклеосомы
Нуклеосомы
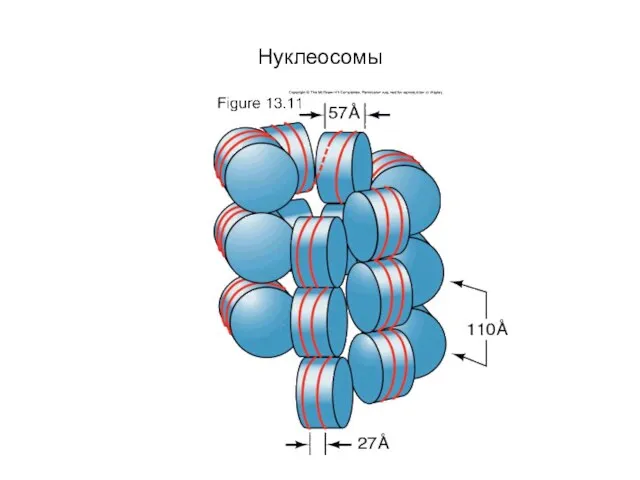
Слайд 9Модель пространственной организации хроматина
Модель пространственной организации хроматина
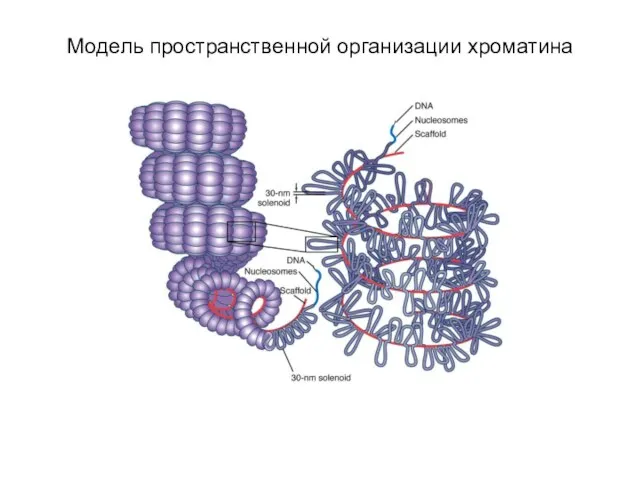
Слайд 10Уровни упаковки ДНК в митотической хромосоме
Уровни упаковки ДНК в митотической хромосоме
Слайд 12Митотическая хромосома в разрезе
Митотическая хромосома в разрезе

Слайд 13Компоненты ядра и ядрышка
Компоненты ядра и ядрышка
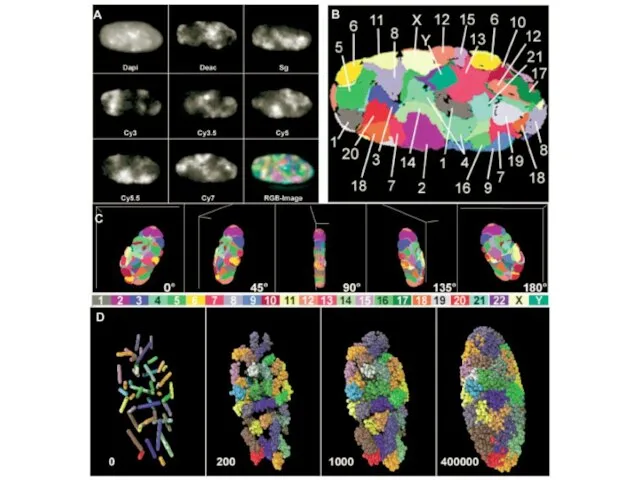
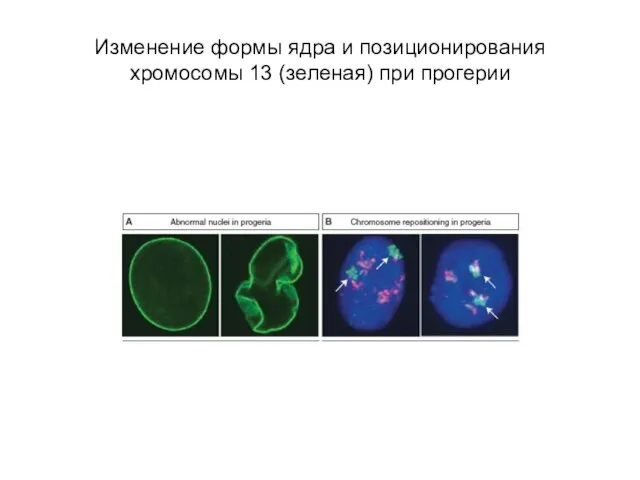
Слайд 16Изменение формы ядра и позиционирования хромосомы 13 (зеленая) при прогерии
Изменение формы ядра и позиционирования хромосомы 13 (зеленая) при прогерии
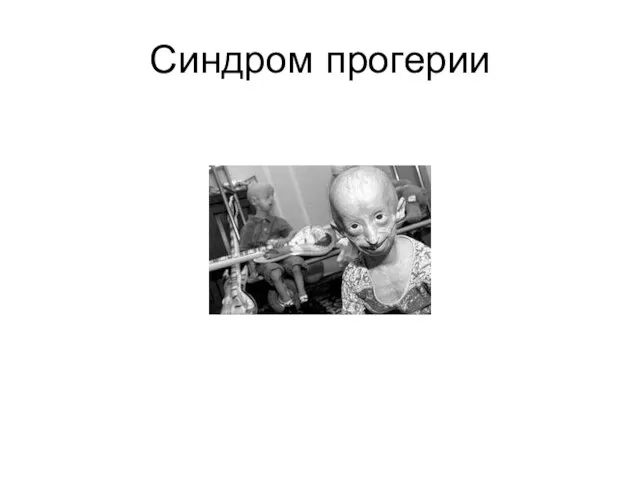
Слайд 17Синдром прогерии
Синдром прогерии
Слайд 19Диплоидный набор хромосом человека
Диплоидный набор хромосом человека

Слайд 20Кинетохор – место прикрепления микротрубочек к хроматиде
Кинетохор – место прикрепления микротрубочек к хроматиде

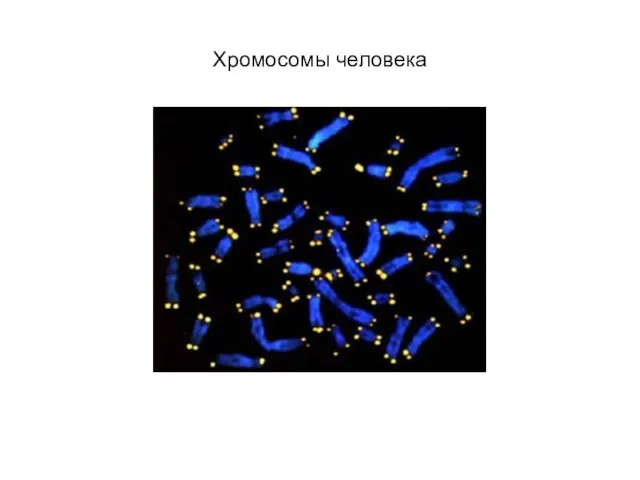
Слайд 21Хромосомы человека
Хромосомы человека
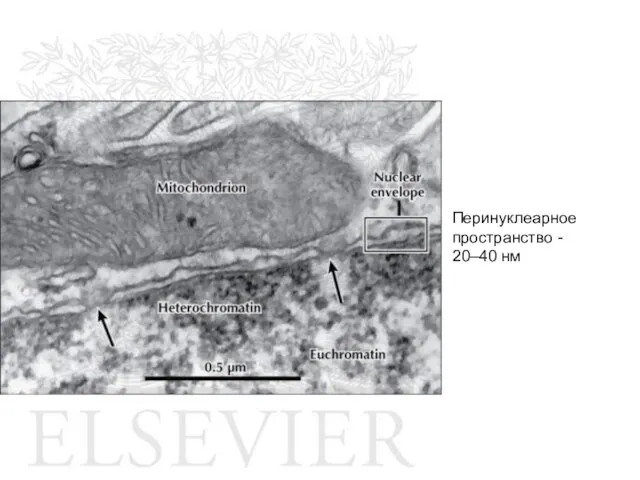
Слайд 22Перинуклеарное пространство - 20–40 нм
Перинуклеарное пространство - 20–40 нм
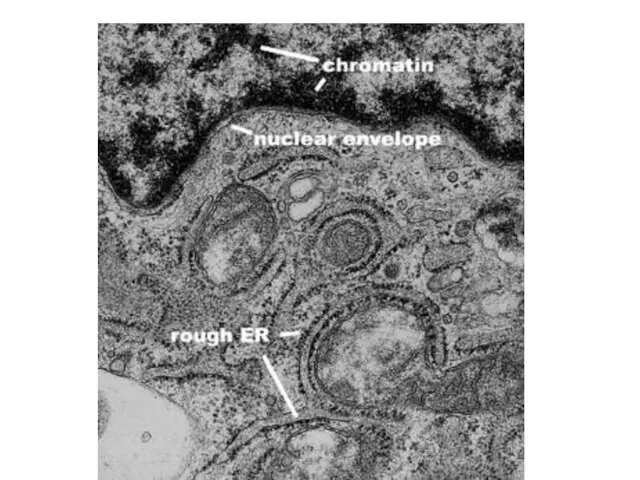

Слайд 25Ядерная ламина (30-100 нм) представляет собой слой белков под внутренней ядерной мембраной
Ядерная ламина (30-100 нм) представляет собой слой белков под внутренней ядерной мембраной

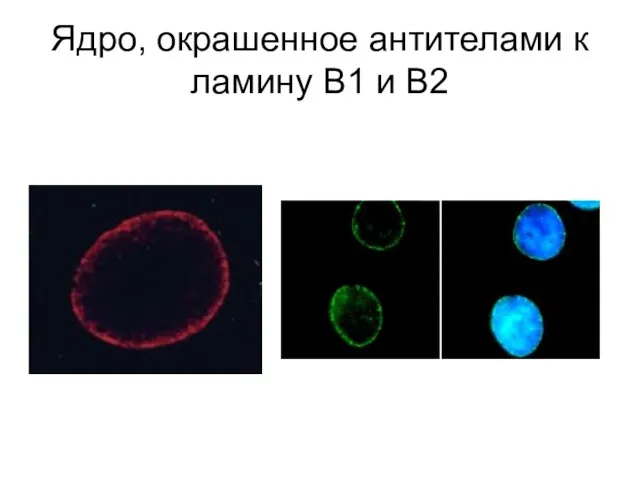
Слайд 26Ядро, окрашенное антителами к ламину В1 и В2
Ядро, окрашенное антителами к ламину В1 и В2
Слайд 27В состав ядерной ламины входят белки: несприн, эмерин, ламинассоциированные белки 1 и
В состав ядерной ламины входят белки: несприн, эмерин, ламинассоциированные белки 1 и
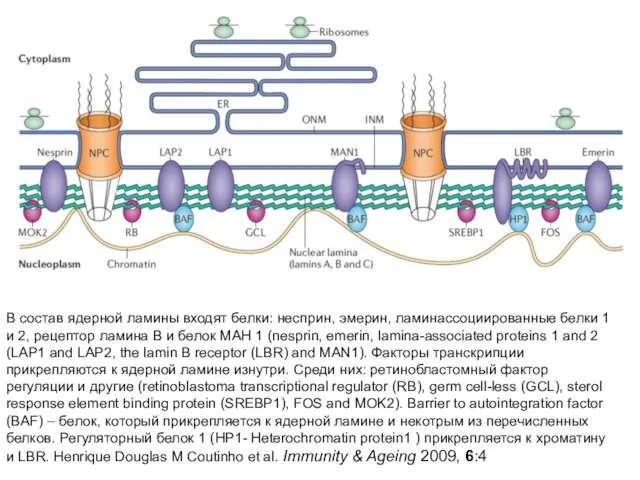
Слайд 28AP, attachment plate; CE, central element of the SC (synaptonemal complex); NPC,
AP, attachment plate; CE, central element of the SC (synaptonemal complex); NPC,
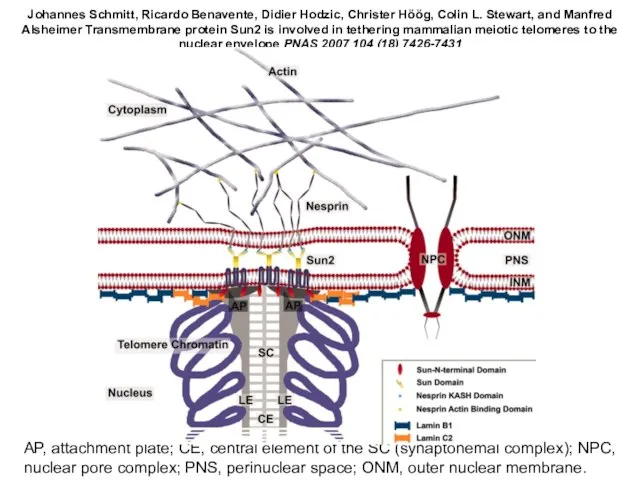
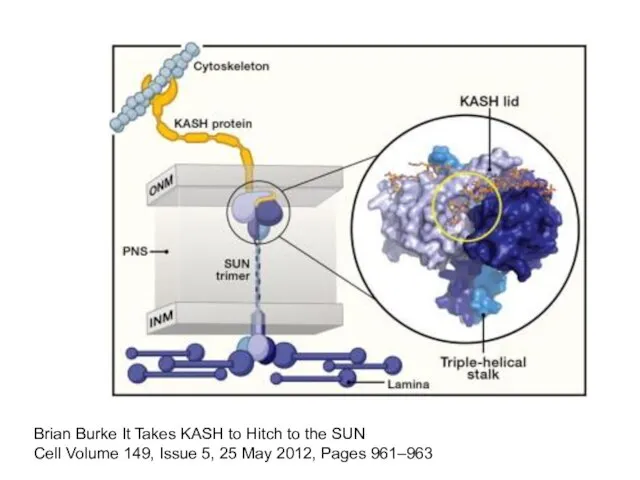
Слайд 29Brian Burke It Takes KASH to Hitch to the SUN
Cell Volume 149,
Brian Burke It Takes KASH to Hitch to the SUN
Cell Volume 149,
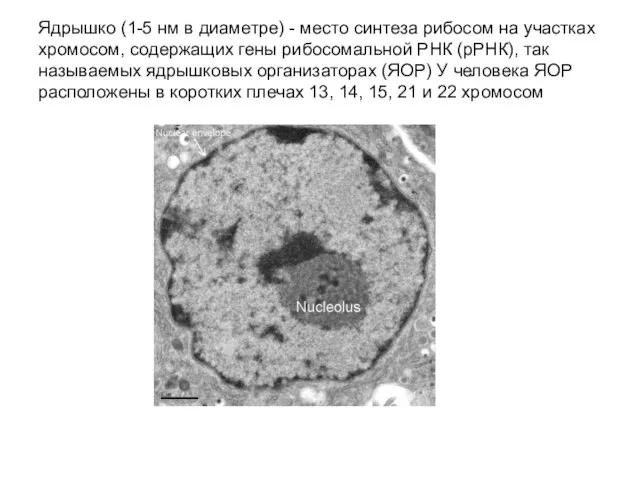
Слайд 32Ядрышко (1-5 нм в диаметре) - место синтеза рибосом на участках хромосом,
Ядрышко (1-5 нм в диаметре) - место синтеза рибосом на участках хромосом,
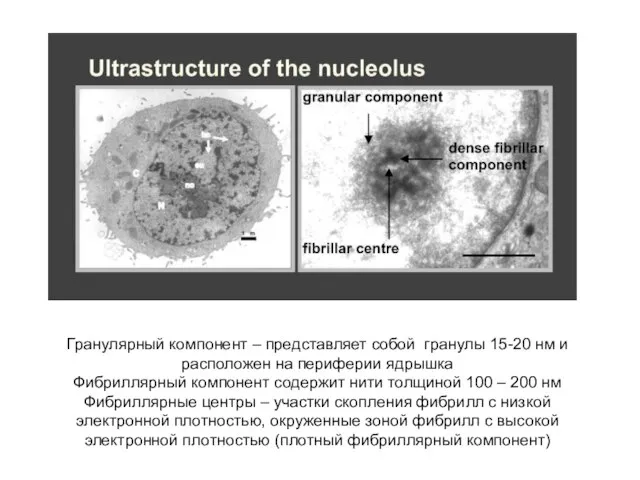
Слайд 33Гранулярный компонент – представляет собой гранулы 15-20 нм и расположен на периферии
Гранулярный компонент – представляет собой гранулы 15-20 нм и расположен на периферии
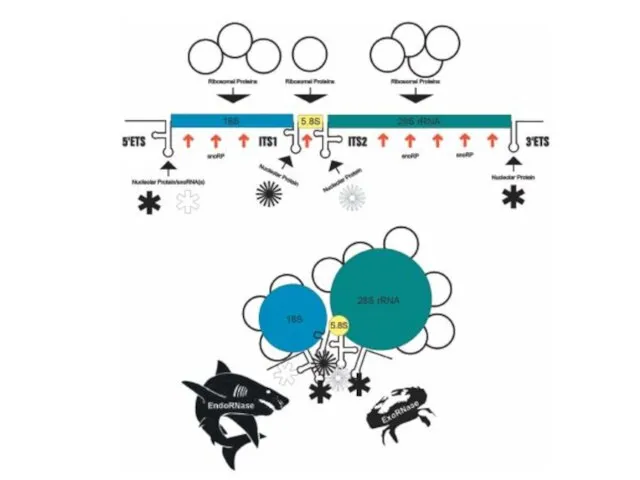
Слайд 34У человека гены, кодирующие рРНК, как и у других организмов, организованы в
У человека гены, кодирующие рРНК, как и у других организмов, организованы в

Слайд 36Полуконсервативный синтез ДНК
Полуконсервативный синтез ДНК

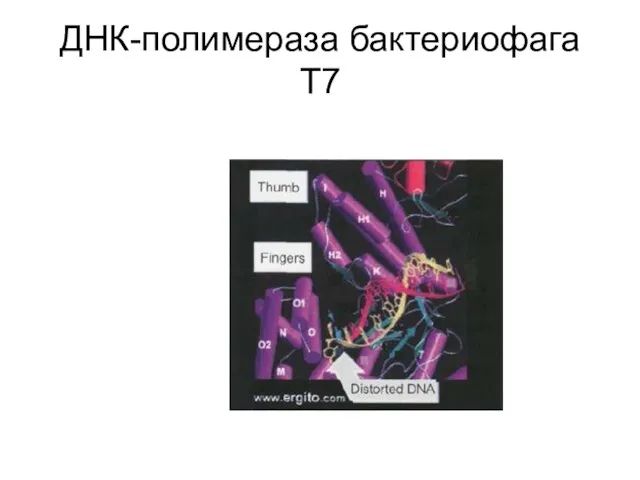
Слайд 37ДНК-полимераза бактериофага Т7
ДНК-полимераза бактериофага Т7
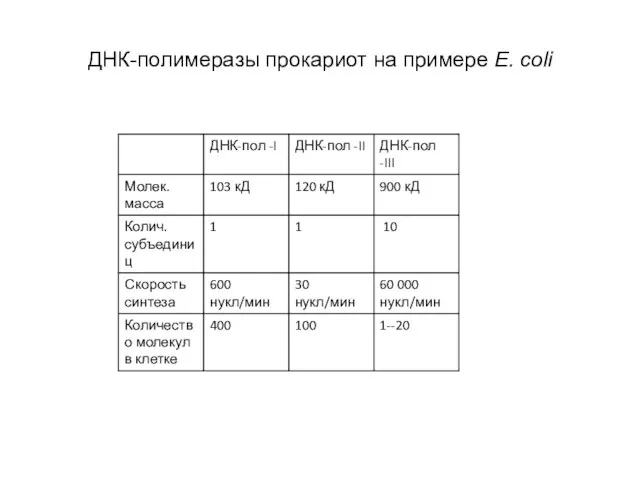
Слайд 38ДНК-полимеразы прокариот на примере E. coli
ДНК-полимеразы прокариот на примере E. coli
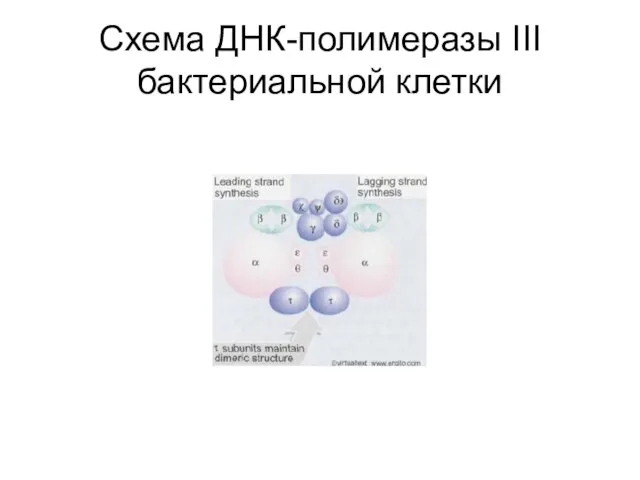
Слайд 39Схема ДНК-полимеразы ІІІ бактериальной клетки
Схема ДНК-полимеразы ІІІ бактериальной клетки
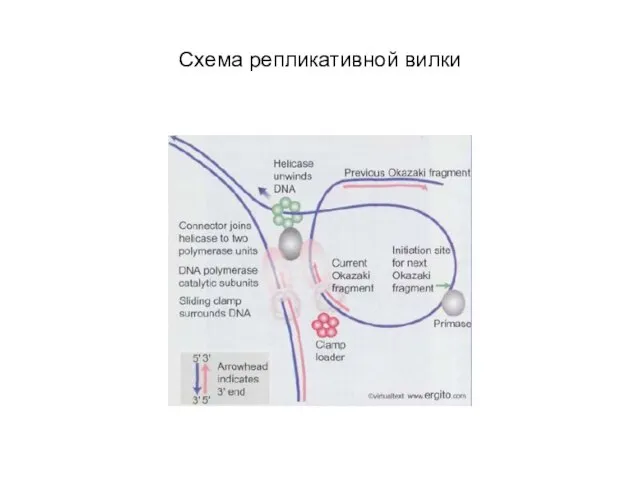
Слайд 40Схема репликативной вилки
Схема репликативной вилки
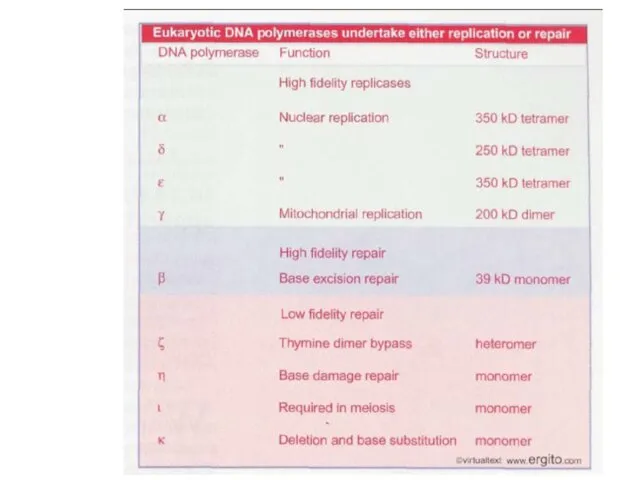
Слайд 42Репликационные фабрики в ядре мыши и дрожжей
Крупные репликационные фабрики в ядре
мыши
Репликационные фабрики в ядре мыши и дрожжей
Крупные репликационные фабрики в ядре
мыши
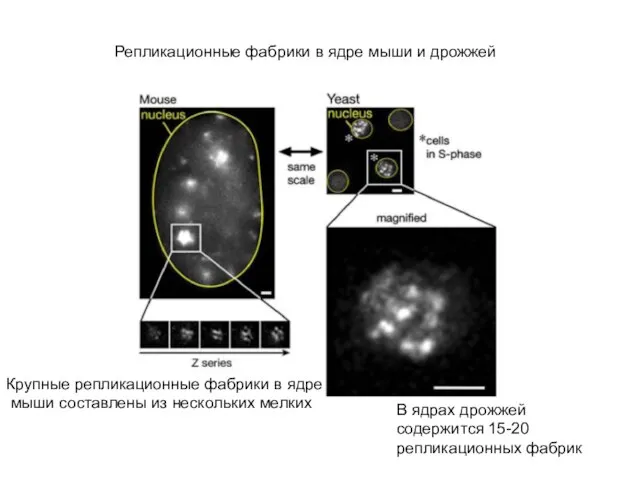
Слайд 43Репликационные фабрики в ядрах СНО, активированных в экстракте ооцитов шпорцевой лягушки Xenopus
Репликационные фабрики в ядрах СНО, активированных в экстракте ооцитов шпорцевой лягушки Xenopus
Слайд 44Пространственная модель РНК-полимеразы прокариотической клетки
Пространственная модель РНК-полимеразы прокариотической клетки

Слайд 45Пространственная модель РНК-полимеразы II клетки дрожжей (12 субъединиц)
Пространственная модель РНК-полимеразы II клетки дрожжей (12 субъединиц)

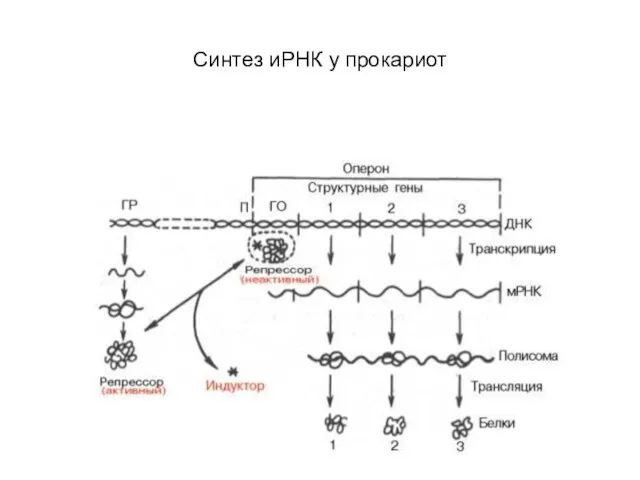
Слайд 46Синтез иРНК у прокариот
Синтез иРНК у прокариот
Слайд 47Промотор для РНК-полимеразы II
Промотор для РНК-полимеразы II

Слайд 48Кэп (7-метилгуанозин) на 3‘ конце эукариотической мРНК
Кэп (7-метилгуанозин) на 3‘ конце эукариотической мРНК
Слайд 49Схема сплайсинга
Схема сплайсинга
Слайд 50Схема строения транскрипционной фабрики
Диаметр фабрики – 56, 75 нм, по данным разных
Схема строения транскрипционной фабрики
Диаметр фабрики – 56, 75 нм, по данным разных

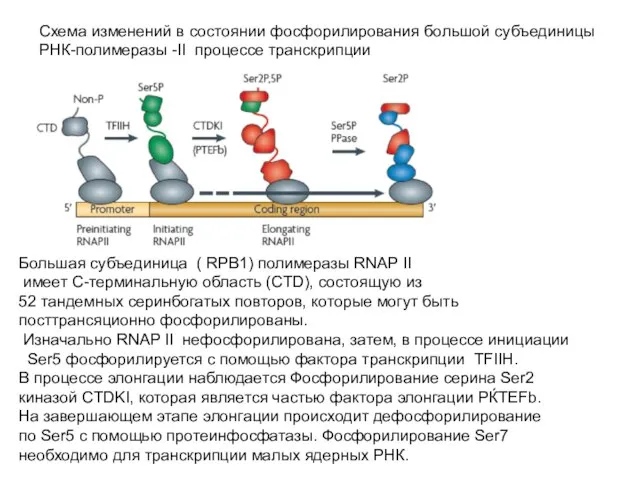
Слайд 51Схема изменений в состоянии фосфорилирования большой субъединицы
РНК-полимеразы -II процессе транскрипции
Большая субъединица
Схема изменений в состоянии фосфорилирования большой субъединицы
РНК-полимеразы -II процессе транскрипции
Большая субъединица
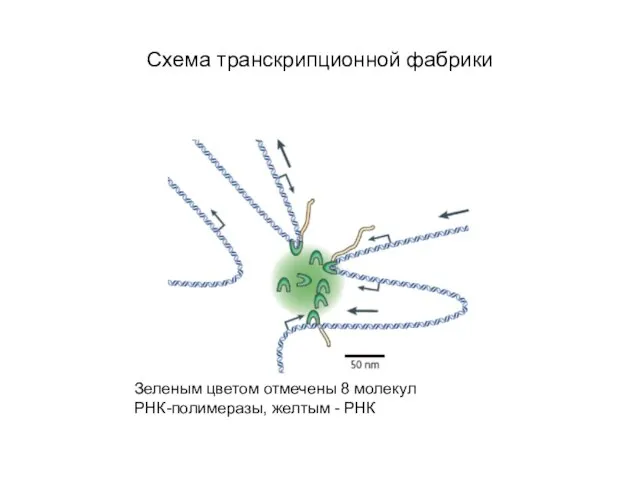
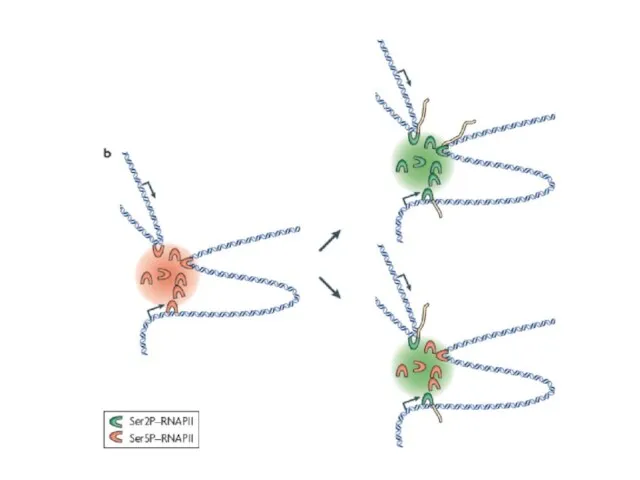
Слайд 52Схема транскрипционной фабрики
Зеленым цветом отмечены 8 молекул РНК-полимеразы, желтым - РНК
Схема транскрипционной фабрики
Зеленым цветом отмечены 8 молекул РНК-полимеразы, желтым - РНК
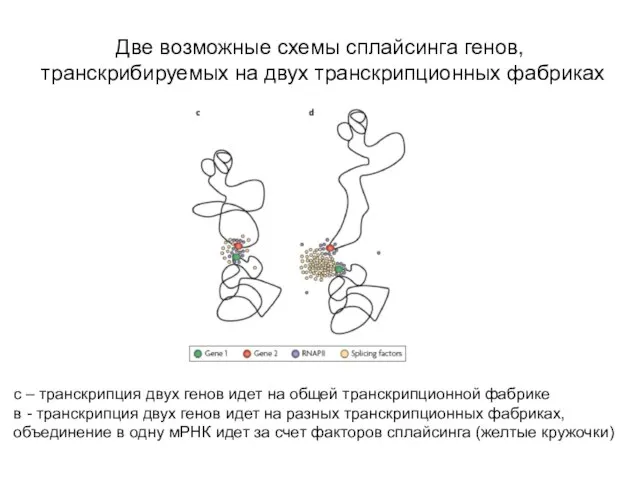
Слайд 54Две возможные схемы сплайсинга генов, транскрибируемых на двух транскрипционных фабриках
с – транскрипция
Две возможные схемы сплайсинга генов, транскрибируемых на двух транскрипционных фабриках
с – транскрипция
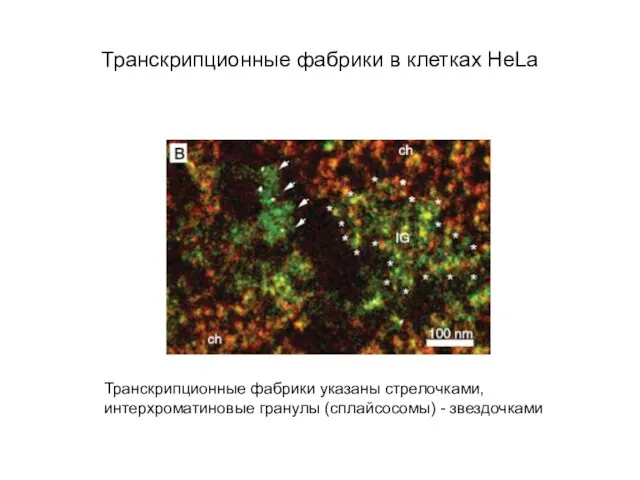
Слайд 56Транскрипционные фабрики в клетках HeLa
Транскрипционные фабрики указаны стрелочками,
интерхроматиновые гранулы (сплайсосомы) - звездочками
Транскрипционные фабрики в клетках HeLa
Транскрипционные фабрики указаны стрелочками,
интерхроматиновые гранулы (сплайсосомы) - звездочками
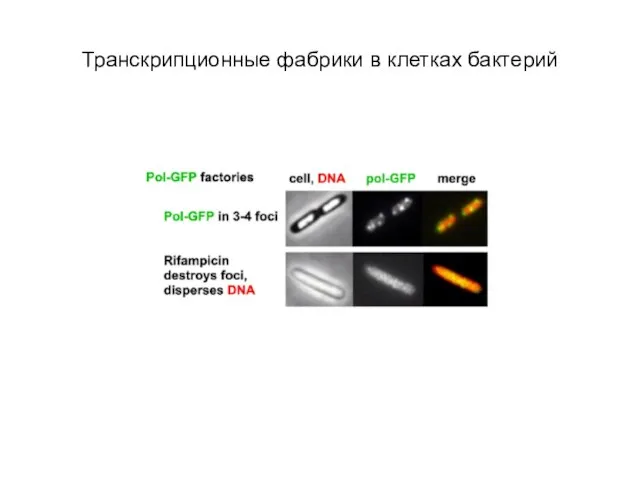
Слайд 57Транскрипционные фабрики в клетках бактерий
Транскрипционные фабрики в клетках бактерий
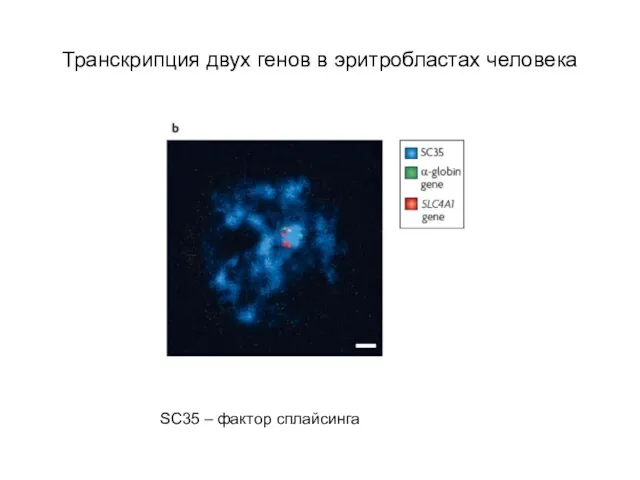
Слайд 58Транскрипция двух генов в эритробластах человека
SC35 – фактор сплайсинга
Транскрипция двух генов в эритробластах человека
SC35 – фактор сплайсинга
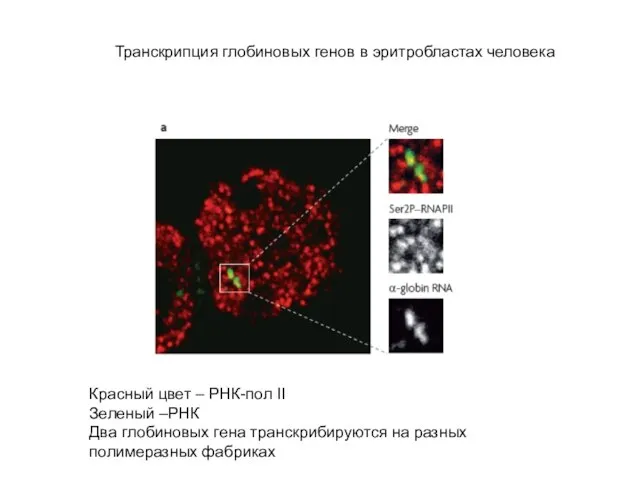
Слайд 59Транскрипция глобиновых генов в эритробластах человека
Красный цвет – РНК-пол II
Зеленый –РНК
Два глобиновых
Транскрипция глобиновых генов в эритробластах человека
Красный цвет – РНК-пол II
Зеленый –РНК
Два глобиновых
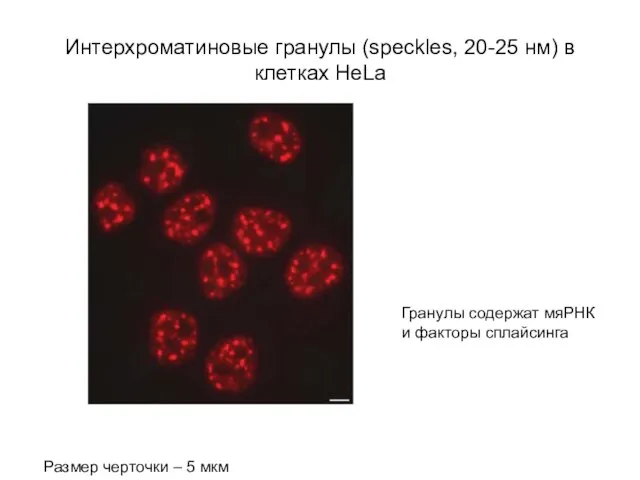
Слайд 60Интерхроматиновые гранулы (speckles, 20-25 нм) в клетках HeLa
Размер черточки – 5 мкм
Гранулы
Интерхроматиновые гранулы (speckles, 20-25 нм) в клетках HeLa
Размер черточки – 5 мкм
Гранулы
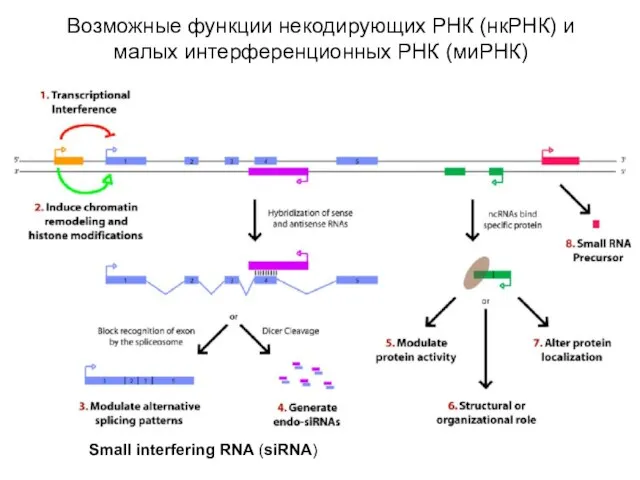
Слайд 61Возможные функции некодирующих РНК (нкРНК) и малых интерференционных РНК (миРНК)
Small interfering RNA
Возможные функции некодирующих РНК (нкРНК) и малых интерференционных РНК (миРНК)
Small interfering RNA
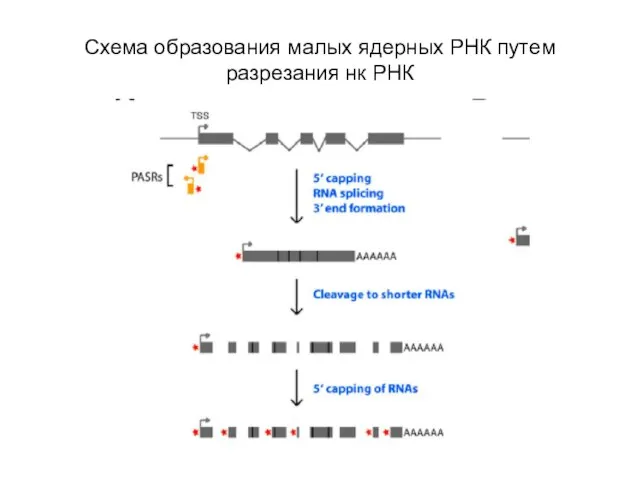
Слайд 62Схема образования малых ядерных РНК путем разрезания нк РНК
Схема образования малых ядерных РНК путем разрезания нк РНК
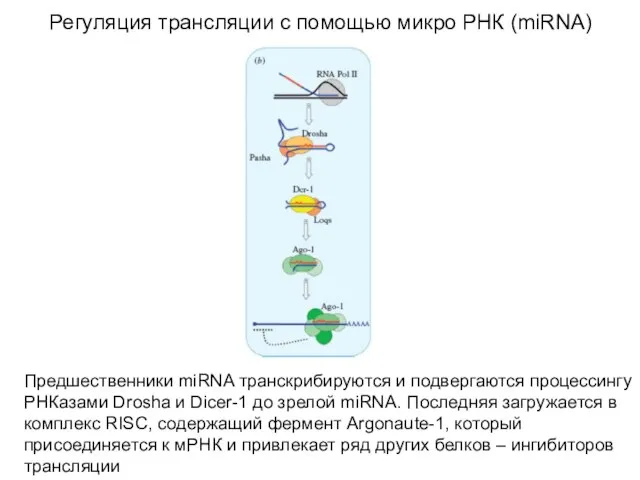
Слайд 63Регуляция трансляции с помощью микро РНК (miRNA)
Предшественники miRNA транскрибируются и подвергаются процессингу
Регуляция трансляции с помощью микро РНК (miRNA)
Предшественники miRNA транскрибируются и подвергаются процессингу
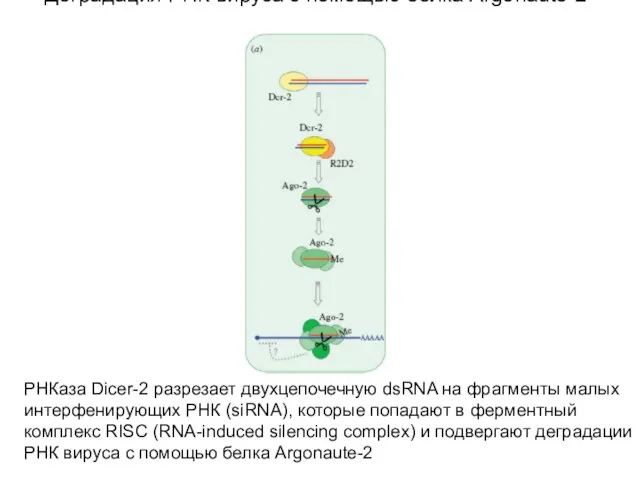
Слайд 64Деградация РНК вируса c помощью белка Argonaute-2
РНКаза Dicer-2 разрезает двухцепочечную dsRNA на
Деградация РНК вируса c помощью белка Argonaute-2
РНКаза Dicer-2 разрезает двухцепочечную dsRNA на
Слайд 65Малые ингибиторные РНК (20-25 нуклеотидов) или small interfering RNA (siRNA) как средство
Малые ингибиторные РНК (20-25 нуклеотидов) или small interfering RNA (siRNA) как средство
 Свобода и ответственность (4 класс)
Свобода и ответственность (4 класс) Презентация на тему Новый год. История праздника
Презентация на тему Новый год. История праздника  Загрязнение автотранспортом
Загрязнение автотранспортом Муниципальное автономное дошкольное образовательное учреждение «Центр развития ребенка – детский сад №50 «Родничок»б
Муниципальное автономное дошкольное образовательное учреждение «Центр развития ребенка – детский сад №50 «Родничок»б Энергетическое использование биомассы в Украине: правовые аспекты
Энергетическое использование биомассы в Украине: правовые аспекты Приоритетными направлениями деятельности общественного движения «Добрые дети мира» в 2010 году будут являться:
Приоритетными направлениями деятельности общественного движения «Добрые дети мира» в 2010 году будут являться: Воркшоп по Мастерской будущего Содержание
Воркшоп по Мастерской будущего Содержание Учителя – участники ВОВ
Учителя – участники ВОВ Цитомегаловирусы
Цитомегаловирусы Политическое лидерство: Дональд Трамп
Политическое лидерство: Дональд Трамп Всегда великая Россия
Всегда великая Россия Подготовка к ГИА по русскому языку часть 2 (тестовые задания и задания с кратким ответом)
Подготовка к ГИА по русскому языку часть 2 (тестовые задания и задания с кратким ответом) Биологическая и психологическая подструктура личности
Биологическая и психологическая подструктура личности Старая печатная русскоязычная реклама парфюмерии
Старая печатная русскоязычная реклама парфюмерии Тема 12.Правонарушения
Тема 12.Правонарушения Творения мира
Творения мира Коллективное управление смежными правами Глава III. Смежные права (Закон РК «Об авторском праве и смежных правах» от 10 июня 1996 года N
Коллективное управление смежными правами Глава III. Смежные права (Закон РК «Об авторском праве и смежных правах» от 10 июня 1996 года N OOP, metaprogramming, blocks, iterators
OOP, metaprogramming, blocks, iterators Независимый депозитарий с сегрегированными счетами и арбитражем сделок для трейдеров и брокеров
Независимый депозитарий с сегрегированными счетами и арбитражем сделок для трейдеров и брокеров Мат одинокому королю
Мат одинокому королю Панно средствами народного примитивного орнаментирования
Панно средствами народного примитивного орнаментирования Имя урока
Имя урока Установка локальной сети в школе
Установка локальной сети в школе Образовательный проект Промсвязьбанка
Образовательный проект Промсвязьбанка Презентация на тему Тюменская область история, география, экономика
Презентация на тему Тюменская область история, география, экономика Санаторий Лесники. Медицинские услуги
Санаторий Лесники. Медицинские услуги Опыт взаимодействия Церкви и государства в трезвенном просвещении
Опыт взаимодействия Церкви и государства в трезвенном просвещении Чытачам Брылеўскай бібліятэкі Красавік 2019 г
Чытачам Брылеўскай бібліятэкі Красавік 2019 г