- Развитие дарвинизма в XIXв – становление филогенетики

Содержание
- 2. Ведущие биологи Англии поддержали Дарвина Судьба собрала их в одно время и в одной стране. Подобно
- 3. Филогенетика Дарвин открыл механизм эволюции, но не ход реального эволюционного процесса Дарвин допускал возможность происхождения животных
- 4. Филогенетика Филогенетическое дерево по Геккелю, 1866 …Эта родословная стоит не дороже родословных героев Гомера. Э. Дюбуа-Реймон
- 5. Филогенетика: заполнение пустой оболочки Эволюционная палеонтология Кювье отрицал наличие переходных форм в ископаемых фаунах Виднейшие палеонтологи
- 6. Филогенетика: заполнение пустой оболочки Эволюционная палеонтология Первым исследовал филогенез крупной ветви позвоночных животных. Есть эволюция таксонов
- 7. Эволюционная палеонтология: «лошадиный ряд» Эволюция органа: Увеличение третьего пальца, связанное с укреплением его суставного соединения, превращение
- 8. Эволюционная палеонтология: «лошадиный ряд» и парнокопытные Скелет ноги ископаемых парнокопытных: 1 – аноплотерий, 2 – ксифодон,
- 9. Эволюционная палеонтология: «лошадиный ряд» Схема эволюции лошадиного ряда. Над схемой изображены конечности: 1 – мезогиппуса, 2
- 10. Эволюционная палеонтология: «лошадиный ряд» Эволюция лошадей: 1 – эогиппус, 2 – протерогиппус, 3 – меригиппус, 4
- 11. Эволюционная палеонтология: «лошадиный ряд» Переход от питания сочными листьями лесных растений к жёсткой траве сухих степей
- 12. Эволюционная палеонтология: «лошадиный ряд» Эволюция зубов у лошадей «Зубы Equus обладают следующими прогрессивными признаками: 1) дифференцировка
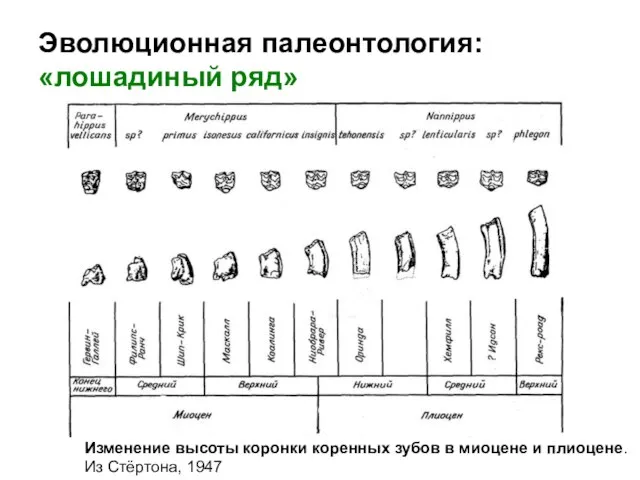
- 13. Эволюционная палеонтология: «лошадиный ряд» Изменение высоты коронки коренных зубов в миоцене и плиоцене. Из Стёртона, 1947
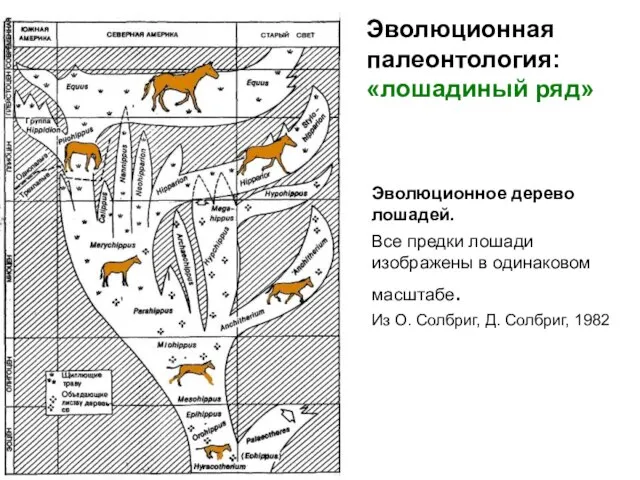
- 14. Эволюционное дерево лошадей. Все предки лошади изображены в одинаковом масштабе. Из О. Солбриг, Д. Солбриг, 1982
- 15. Филогенетика: «основной биогенетический закон» Дарвин: «Интерес эмбриологии значительно повысится, если мы будем видеть в зародыше более
- 16. Филогенетика: «основной биогенетический закон» Мюллер исходил из положения Дарвина об изменяемости в процессе эволюции любой стадии
- 17. Филогенетика: «основной биогенетический закон» Эрнст Геккель «…ведущее значение для выяснения родственных отношений между организ-мами имеют эмбриологические
- 18. Филогенетика: заполнение пустой оболочки «основной биогенетический закон» «Подобно тому, как на современном этапе развития биологии наиболее
- 19. Филогенетика: заполнение пустой оболочки «основной биогенетический закон» Изучил эмбриогенез многих беспозвоночных Обнаружил гомологи зародышевых листков у
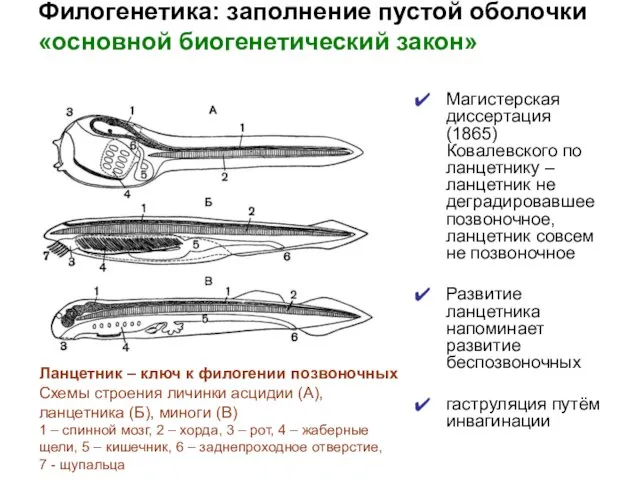
- 20. Филогенетика: заполнение пустой оболочки «основной биогенетический закон» Магистерская диссертация (1865) Ковалевского по ланцетнику – ланцетник не
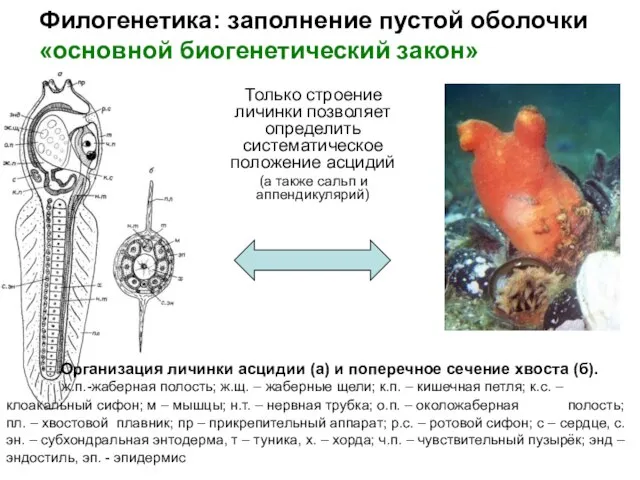
- 21. Филогенетика: заполнение пустой оболочки «основной биогенетический закон» Только строение личинки позволяет определить систематическое положение асцидий (а
- 22. У ланцетника радиальное дробление яйца, которое приводит к образованию полого шарика – бластулы, из которого путём
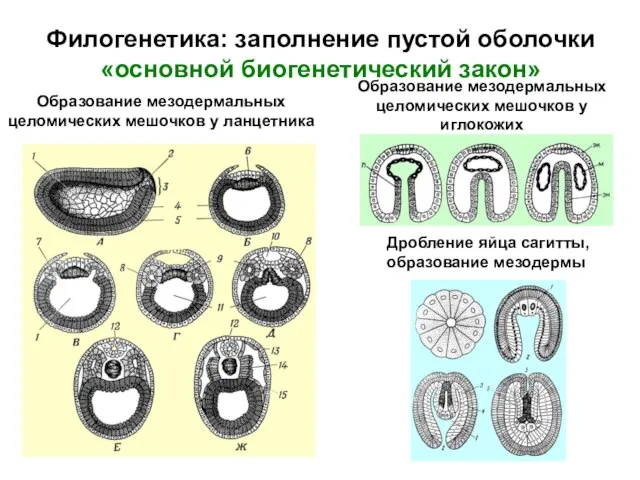
- 23. Филогенетика: заполнение пустой оболочки «основной биогенетический закон» Дробление яйца сагитты, образование мезодермы Образование мезодермальных целомических мешочков
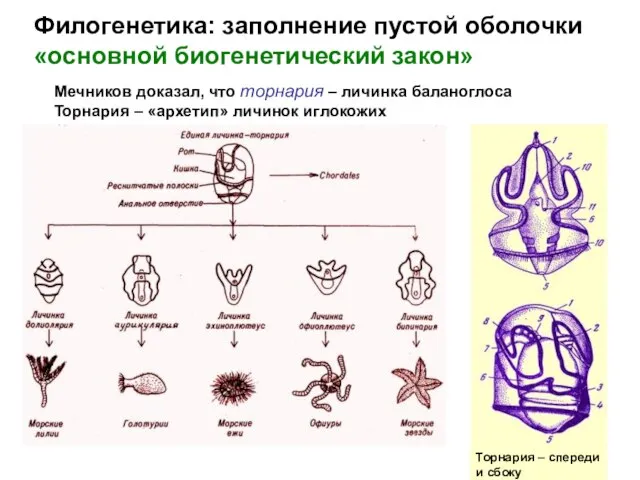
- 24. Филогенетика: заполнение пустой оболочки «основной биогенетический закон» Мечников доказал, что торнария – личинка баланоглоса Торнария –
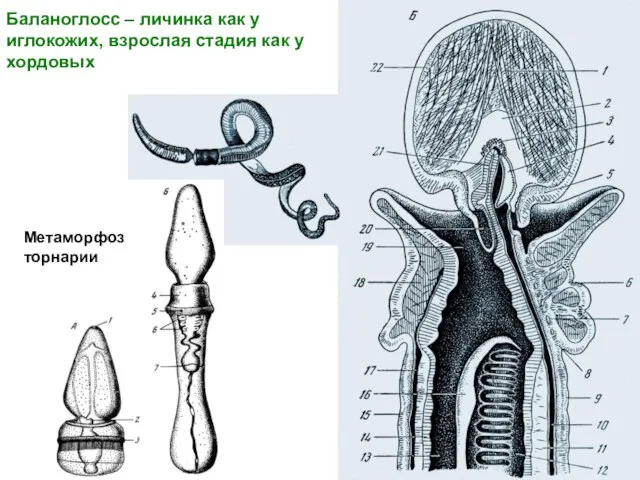
- 25. Баланоглосс – личинка как у иглокожих, взрослая стадия как у хордовых Метаморфоз торнарии
- 26. Филогенетика: заполнение пустой оболочки «основной биогенетический закон» Геккель считал открытые Ковалевским стадии развития зародышей вторичноротых рекапитуляцией
- 27. Научная жизнь Мечникова делится на русский, сравнительно-эмбриологический период и французский, иммунологический. Работая совместно с А.О. Ковалевским
- 28. Филогенетика: заполнение пустой оболочки «основной биогенетический закон» Эволюция форм гаструляции (из Ивановой-Казас) Типы бластул и связанные
- 29. Филогенетика: заполнение пустой оболочки «основной биогенетический закон» Главнейшие предполагаемые стадии развития Metazoa по Иванову (1968) А.Б
- 30. Филогенетика: заполнение пустой оболочки «основной биогенетический закон» …эти зародышевые пласты остаются гомологичными друг другу у всех
- 31. Филогенетика: заполнение пустой оболочки «основной биогенетический закон» СЕВЕРЦОВ Алексей Николаевич (1866-1936) капитальные труды по филогенезу низших
- 33. Скачать презентацию

Слайд 3Филогенетика
Дарвин открыл механизм эволюции, но не ход реального эволюционного процесса
Дарвин допускал возможность
Филогенетика
Дарвин открыл механизм эволюции, но не ход реального эволюционного процесса
Дарвин допускал возможность

Слайд 4Филогенетика
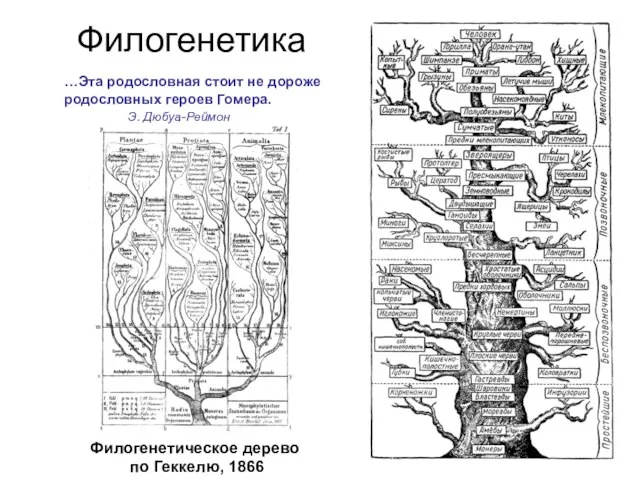
Филогенетическое дерево
по Геккелю, 1866
…Эта родословная стоит не дороже
родословных героев Гомера.
Э. Дюбуа-Реймон
Филогенетика
Филогенетическое дерево
по Геккелю, 1866
…Эта родословная стоит не дороже
родословных героев Гомера.
Э. Дюбуа-Реймон
Слайд 5Филогенетика: заполнение пустой оболочки
Эволюционная палеонтология
Кювье отрицал наличие переходных форм в ископаемых фаунах
Виднейшие
Филогенетика: заполнение пустой оболочки
Эволюционная палеонтология
Кювье отрицал наличие переходных форм в ископаемых фаунах
Виднейшие

Слайд 6Филогенетика: заполнение пустой оболочки
Эволюционная палеонтология
Первым исследовал филогенез крупной ветви позвоночных животных.
Есть
Филогенетика: заполнение пустой оболочки
Эволюционная палеонтология
Первым исследовал филогенез крупной ветви позвоночных животных.
Есть

Слайд 7Эволюционная палеонтология:
«лошадиный ряд»
Эволюция органа: Увеличение третьего пальца, связанное с укреплением его суставного
Эволюционная палеонтология:
«лошадиный ряд»
Эволюция органа: Увеличение третьего пальца, связанное с укреплением его суставного

Слайд 8Эволюционная палеонтология:
«лошадиный ряд» и парнокопытные
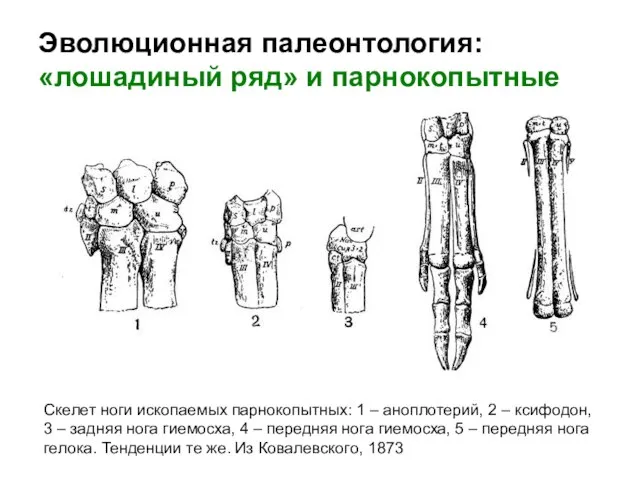
Скелет ноги ископаемых парнокопытных: 1 – аноплотерий, 2
Эволюционная палеонтология:
«лошадиный ряд» и парнокопытные
Скелет ноги ископаемых парнокопытных: 1 – аноплотерий, 2
Слайд 9Эволюционная палеонтология:
«лошадиный ряд»
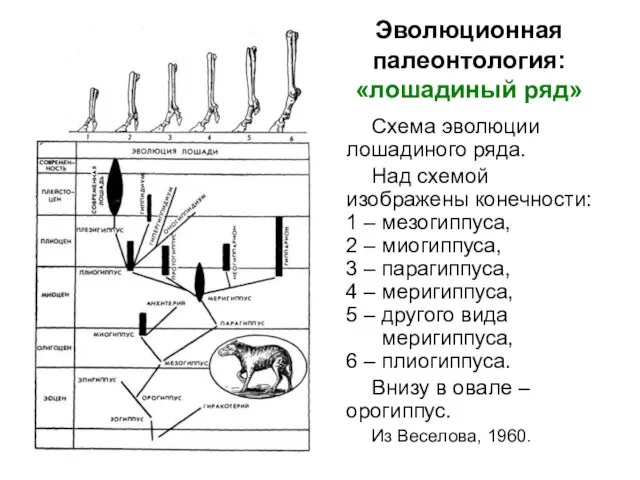
Схема эволюции лошадиного ряда.
Над схемой изображены конечности: 1 – мезогиппуса,
Эволюционная палеонтология:
«лошадиный ряд»
Схема эволюции лошадиного ряда.
Над схемой изображены конечности: 1 – мезогиппуса,
Слайд 10Эволюционная палеонтология:
«лошадиный ряд»
Эволюция лошадей:
1 – эогиппус,
2 – протерогиппус,
3 –
Эволюционная палеонтология:
«лошадиный ряд»
Эволюция лошадей:
1 – эогиппус,
2 – протерогиппус,
3 –

Слайд 11Эволюционная палеонтология:
«лошадиный ряд»
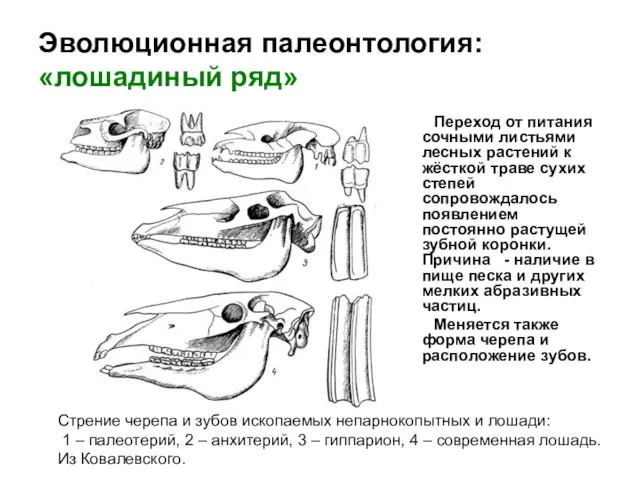
Переход от питания сочными листьями лесных растений к жёсткой траве
Эволюционная палеонтология:
«лошадиный ряд»
Переход от питания сочными листьями лесных растений к жёсткой траве
Слайд 12Эволюционная палеонтология:
«лошадиный ряд»
Эволюция зубов у лошадей
«Зубы Equus обладают следующими прогрессивными признаками:
1)
Эволюционная палеонтология:
«лошадиный ряд»
Эволюция зубов у лошадей
«Зубы Equus обладают следующими прогрессивными признаками:
1)

Слайд 13Эволюционная палеонтология:
«лошадиный ряд»
Изменение высоты коронки коренных зубов в миоцене и плиоцене.
Из
Эволюционная палеонтология:
«лошадиный ряд»
Изменение высоты коронки коренных зубов в миоцене и плиоцене.
Из
Слайд 14Эволюционное дерево лошадей.
Все предки лошади изображены в одинаковом масштабе.
Из О. Солбриг,
Все предки лошади изображены в одинаковом масштабе.
Из О. Солбриг,
Слайд 15Филогенетика:
«основной биогенетический закон»
Дарвин: «Интерес эмбриологии значительно повысится, если мы будем видеть
Филогенетика:
«основной биогенетический закон»
Дарвин: «Интерес эмбриологии значительно повысится, если мы будем видеть

Слайд 16Филогенетика:
«основной биогенетический закон»
Мюллер исходил из положения Дарвина об изменяемости в процессе
Филогенетика:
«основной биогенетический закон»
Мюллер исходил из положения Дарвина об изменяемости в процессе

Слайд 17Филогенетика:
«основной биогенетический закон»
Эрнст Геккель
«…ведущее значение для выяснения родственных отношений между организ-мами
Филогенетика:
«основной биогенетический закон»
Эрнст Геккель
«…ведущее значение для выяснения родственных отношений между организ-мами

Слайд 18Филогенетика: заполнение пустой оболочки
«основной биогенетический закон»
«Подобно тому, как на современном этапе развития
Филогенетика: заполнение пустой оболочки
«основной биогенетический закон»
«Подобно тому, как на современном этапе развития

Слайд 19Филогенетика: заполнение пустой оболочки
«основной биогенетический закон»
Изучил эмбриогенез многих беспозвоночных
Обнаружил гомологи зародышевых листков
Филогенетика: заполнение пустой оболочки
«основной биогенетический закон»
Изучил эмбриогенез многих беспозвоночных
Обнаружил гомологи зародышевых листков

Слайд 20Филогенетика: заполнение пустой оболочки
«основной биогенетический закон»
Магистерская диссертация (1865) Ковалевского по ланцетнику –
Филогенетика: заполнение пустой оболочки
«основной биогенетический закон»
Магистерская диссертация (1865) Ковалевского по ланцетнику –
Слайд 21Филогенетика: заполнение пустой оболочки
«основной биогенетический закон»
Только строение личинки позволяет определить систематическое положение
Филогенетика: заполнение пустой оболочки
«основной биогенетический закон»
Только строение личинки позволяет определить систематическое положение
Слайд 22У ланцетника радиальное дробление яйца, которое приводит к образованию полого шарика –
У ланцетника радиальное дробление яйца, которое приводит к образованию полого шарика –
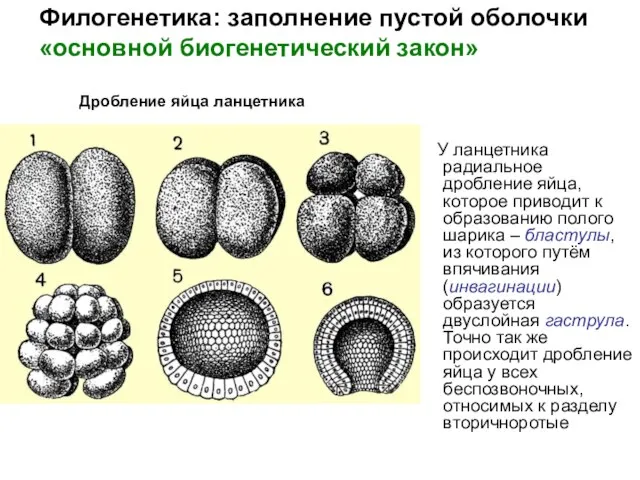
Слайд 23Филогенетика: заполнение пустой оболочки
«основной биогенетический закон»
Дробление яйца сагитты,
образование мезодермы
Образование мезодермальных
целомических мешочков у
иглокожих
Образование
Филогенетика: заполнение пустой оболочки
«основной биогенетический закон»
Дробление яйца сагитты,
образование мезодермы
Образование мезодермальных
целомических мешочков у
иглокожих
Образование
Слайд 24Филогенетика: заполнение пустой оболочки
«основной биогенетический закон»
Мечников доказал, что торнария – личинка баланоглоса
Торнария
Филогенетика: заполнение пустой оболочки
«основной биогенетический закон»
Мечников доказал, что торнария – личинка баланоглоса
Торнария
Слайд 25Баланоглосс – личинка как у
иглокожих, взрослая стадия как у хордовых
Метаморфоз
торнарии
Баланоглосс – личинка как у
иглокожих, взрослая стадия как у хордовых
Метаморфоз
торнарии
Слайд 26Филогенетика: заполнение пустой оболочки
«основной биогенетический закон»
Геккель считал открытые Ковалевским стадии развития зародышей
Филогенетика: заполнение пустой оболочки
«основной биогенетический закон»
Геккель считал открытые Ковалевским стадии развития зародышей

Слайд 27Научная жизнь Мечникова делится на русский, сравнительно-эмбриологический период и французский, иммунологический.
Работая совместно
Научная жизнь Мечникова делится на русский, сравнительно-эмбриологический период и французский, иммунологический.
Работая совместно

Слайд 28Филогенетика: заполнение пустой оболочки
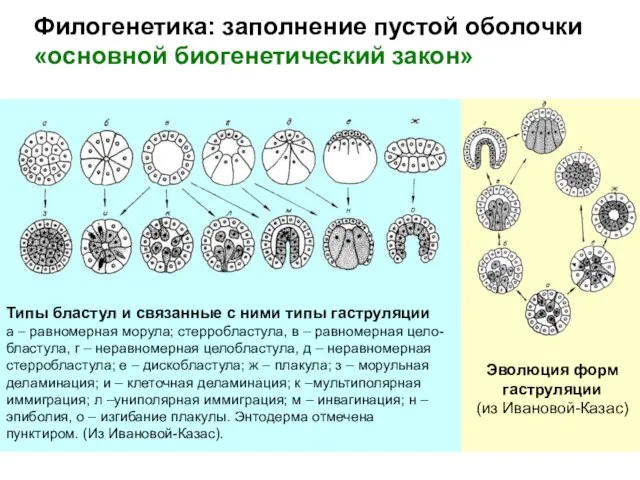
«основной биогенетический закон»
Эволюция форм
гаструляции
(из Ивановой-Казас)
Типы бластул и связанные с
Филогенетика: заполнение пустой оболочки
«основной биогенетический закон»
Эволюция форм
гаструляции
(из Ивановой-Казас)
Типы бластул и связанные с
Слайд 29Филогенетика: заполнение пустой
оболочки
«основной биогенетический закон»
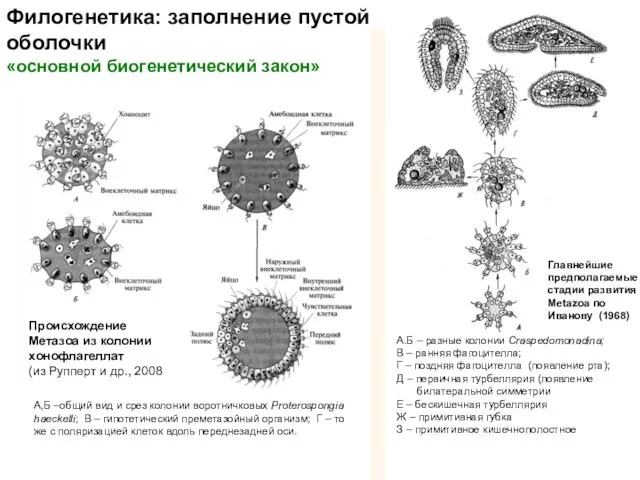
Главнейшие
предполагаемые стадии развития Metazoa по Иванову
Филогенетика: заполнение пустой
оболочки
«основной биогенетический закон»
Главнейшие
предполагаемые стадии развития Metazoa по Иванову
Слайд 30Филогенетика: заполнение пустой
оболочки
«основной биогенетический закон»
…эти зародышевые пласты остаются гомологичными друг другу
Филогенетика: заполнение пустой
оболочки
«основной биогенетический закон»
…эти зародышевые пласты остаются гомологичными друг другу

Слайд 31Филогенетика: заполнение пустой оболочки
«основной биогенетический закон»
СЕВЕРЦОВ
Алексей Николаевич
(1866-1936)
капитальные труды по филогенезу низших позвоночных
Филэмбриогенезы
Филогенетика: заполнение пустой оболочки
«основной биогенетический закон»
СЕВЕРЦОВ
Алексей Николаевич
(1866-1936)
капитальные труды по филогенезу низших позвоночных
Филэмбриогенезы

 Средства автоматизации сметных расчетов
Средства автоматизации сметных расчетов Публичный доклад МОУ «Нижнекулойская средняя общеобразовательная школа» за 2009 – 2010 учебный год
Публичный доклад МОУ «Нижнекулойская средняя общеобразовательная школа» за 2009 – 2010 учебный год Информация и знания Чувственное познание окружающего мира
Информация и знания Чувственное познание окружающего мира  Чертежи деталей и изделий из древесины
Чертежи деталей и изделий из древесины Презентация на тему Звук, ультразвук, инфразвук и их использование
Презентация на тему Звук, ультразвук, инфразвук и их использование Презентация на тему Амазонка
Презентация на тему Амазонка Энергетический голод
Энергетический голод SIMECO ОСНОВНЫЕ ПРОЕКТЫ 2001-2011Version: v8
SIMECO ОСНОВНЫЕ ПРОЕКТЫ 2001-2011Version: v8 Мой образовательный маршрут
Мой образовательный маршрут Исследование свойства цвета светлота
Исследование свойства цвета светлота Молодежная наркомания
Молодежная наркомания  1-я Эмоция
1-я Эмоция Путешествие к хлудневским мастерам
Путешествие к хлудневским мастерам Касимов
Касимов Сотрудничество с Орто-М
Сотрудничество с Орто-М Для друга 2.0
Для друга 2.0 Презентация на тему Энциклопедия слова - земляника
Презентация на тему Энциклопедия слова - земляника «Бизнес-Продовольствие 2011»
«Бизнес-Продовольствие 2011» День Матери
День Матери Лесные производители.
Лесные производители. Учебно-методические комплексы (УМК) для начальной школы
Учебно-методические комплексы (УМК) для начальной школы Сетевой подход в маркетинге как основа коммерциализации инноваций
Сетевой подход в маркетинге как основа коммерциализации инноваций Vegas night. Баннер
Vegas night. Баннер HUMAN RESOURCE MANAGEMENT
HUMAN RESOURCE MANAGEMENT Расчёт пути и времени движения
Расчёт пути и времени движения Планета Земля. Пятый век. Коммунистическая эра
Планета Земля. Пятый век. Коммунистическая эра Звон колоколов
Звон колоколов Презентация на тему Концепции и глобальные модели будущего мира
Презентация на тему Концепции и глобальные модели будущего мира